
ABSTRACT
Copper is one of the metals described to bind the Parkinson disease-related protein α-synuclein (aSyn), and to promote its aggregation. Although histidine at position 50 in the aSyn sequence is one of the most studied copper-anchoring sites, its precise role in copper binding and aSyn aggregation is still unclear. Previous studies suggested that this residue does not significantly affect copper-mediated aSyn aggregation. However, our findings showed that the aggregation of the pathological H50Q aSyn mutant is enhanced by copper hints otherwise. Despite the inexistence of a model for aSyn H50Q-copper complexation, we discuss possible mechanisms by which this metal contributes to the misfolding and self-assembly of this particular aSyn mutant. Considering the genetic association of the H50Q mutation with familial forms of Parkinson disease, and the fact that copper homeostasis is deregulated in this disorder, understanding the interplay between both factors will shed light into the molecular and cellular mechanisms triggering the development and spreading of the aSyn pathology.
The abnormal accumulation of α-synuclein (aSyn) is a characteristic of various neurodegenerative diseases, such as Parkinson disease (PD), that are collectively known as synucleinopathies.Citation1 However, the pathological consequences of this process are still not clear, neither the molecular mechanisms promoting and/or causing aSyn aggregation. Factors that promote aSyn misfolding and self-assembly include point mutations, found in genetic forms of synucleinopathies, and the presence of environmental stimuli, such as pro-aggregation metals.Citation2,3 We recently exploited the unique combination of the H50Q pathological aSyn mutation and the presence of copper(II) (Cu2+), a metal whose homeostasis is deregulated in many neurodegenerative conditions, and found that the neuronal damage induced by the aSyn species used in our study is not correlated with the accumulation of intracellular aSyn inclusions, supporting the dissociation between toxicity and the accumulation of large inclusion bodies.Citation4
Since the recent discovery of the H50Q mutation in familial forms of PD, various studies reported that it enhances the aggregation of aSyn in vitro.Citation5-7 Additionally, Cu2+ and other metals are known to accelerate aSyn fibrillation.Citation8 Cu2+ interacts with the negatively-charged C-terminal region of the protein, but binds the N-terminal region with higher affinity.Citation9,10 Two main possible complexation modes in the N-terminus are suggested: one consider the first few residues and His50 (together with neighboring residues) as 2 independent binding sites;Citation10 the other considers the formation of a long-range bridge in the polypeptide chain through the binding of a single metal ion, in which His 50 would act as a coordination switch ().Citation9 Cu2+ coordination provokes compaction in aSyn.Citation11,12 These conformational changes, even if subtle, may reduce the free energy barrier and trigger aggregation. However, these hypotheses are challenged by the recent finding that the aggregation-inducing activity of Cu2+ is stronger in the H50Q mutant than in the wild-type (WT).Citation4 This conclusion is derived not only from cell-free aggregation experiments but also from a cellular model where a panel of pathological mutations was expressed and exposed to different metals and only the combination of Cu2+ and H50Q resulted in the formation of inclusion bodies.Citation4 This uncovers a plethora of questions regarding the binding mode of Cu2+ in this particular mutant of aSyn, and why such event causes the rapid formation of aSyn aggregates ().
Figure 1. The H50 residue is key for anchoring Cu2+ binding to aSyn. Quantum Mechanics / Molecular Mechanics (QM/MM) simulationsCitation21 (21) were performed to test the stability of all the possible coordination geometries of Cu2+ at H50 site, fitting available experimental evidences.Citation14,17,20 Models A and B results to be the most plausible. The Cu2+ ion binds to H50 side chain in both of them. The 3 additional ligands are the H50 amide group, a water molecule, and either V48 carbonyl O (model A) or V49 deprotonated amide (model B). All of the coordination bonds exhibited relatively small fluctuations around their average values, except for the Cu2+ -V48 carbonyl and Cu2+ -V49 amide. This suggests an interconversion between the 2 forms. Residues and water molecules are shown in ball-and-stick representation: red, blue, gray and white for oxygen, nitrogen, carbon and hydrogen, respectively. Cu2+ is shown as a golden ball.
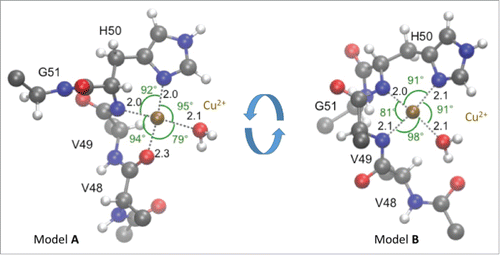
Table 1. Effects of Cu2+ on H50Q vs. WT aSyn.
Existing data indicate that the binding profile of aSyn-Cu2+ is not significantly disturbed in the H50Q mutant, despite the absence of the His50 site, suggesting the absence of coordination of Cu2+ with Gln50.Citation5 However, the faster aggregation of H50Q in the presence of Cu2+, when compared with WT aSyn, indicates that position 50 likely plays a role in modulating the effect of Cu2+. One possibility is that His50 lies on a “strategical” position in the sequence, in such a way that the binding of Cu2+ to this residue may induce a local turn-like structure, thereby favoring β-sheet nucleation.Citation13 The lack of this process in the H50Q variant, due to the lack of Cu2+ binding at position 50, would trigger other misfolding mechanisms, resulting in different aggregation characteristics. Another possibility is that, indeed, the differences between Cu2+−promoted aggregation of WT and H50Q do not depend on conformational changes in the beginning of the aggregation process, but rather on the subsequent elongation stages. However, previous studies reported that, at least with WT aSyn, Cu2+ mainly influences the lag phase of aggregation, promoting accelerated nucleation.Citation14 Similarly, although the Cu2+ effect on WT aSyn seems restricted to intramolecular phenomenaCitation11 promotion of intermolecular aberrant interactions could occur in the H50Q. Additionally, it is also possible that the increased flexibility of the C-terminal region in the H50Q mutantCitation15 could change the Cu2+-binding features in this domain and, thereby affect the overall aggregation process. Although the absence of the imidazole ring of His50 may disturb Cu2+-triggered aSyn aggregation, Gln50 may also promote a toxic gain-of-function role. The aggregation of different aSyn point-mutants at position 50 revealed that, while H50Q was the more aggregation-prone among the mutants tested, a positive charge at this site (WT or H50R) probably protects from fibrilization.Citation15 A similar experiment performed in the presence of Cu2+ would contribute to defining the effect of Gln50 in Cu2+ binding. Even if the Gln50 is not directly involved in metal coordination, it could play a role in stabilizing protein-metal complexation.Citation16
Electron microscopy confirmed that fibrils formed by the H50Q mutant resemble those formed by WT aSyn.Citation4-6 Cu2+ also does not alter the morphology of fibers formed by WT aSyn.Citation14 However, the combination of the H50Q mutation and Cu2+ results in the formation of amorphous aggregates.Citation4 Although this could be the mere consequence of the very rapid self-assembly rate, it will be worthwhile exploring this issue further to understand when and where those morphological changes arise.
Finally, certain physiologic circumstances may also cause the loss of Cu2+-binding, such as N-terminal acetylation of aSyn or a reduction in the micro-environmental pH.Citation17,18 The latter would result in protonation of the His50 imidazole ring, impairing metal binding. In this case, the H50Q substitution could create a similar scenario, forcing Cu2+ to coordinate only to the N-terminal cluster. The lack of a potential intramolecular bridge supported by a Cu2+ ion might alter the entire conformation, probably inducing more open Cu2+-aSyn complexation modes, affecting the aggregation process and eventually the toxicity of the resultant species.
In the future, when further addressing the role of Cu2+ as an enhancer of the H50Q aggregation, physiologic micro-environments and posttranslational modifications will have to be considered. A molecular understanding of the interplay between genetic factors, such as aSyn mutations and environmental factors, such as the presence of Cu2+, might greatly contribute for our understanding of the pathological events causing PD.
Disclosure of potential conflicts of interest
No potential conflicts of interest were disclosed.
Funding
AV-P is supported by the Georg-August–Universität Göttingen under the Dorothea Schlözer Program. PC and GR were supported in this project by the Deutsche Forschungsgemeinschaft (DFG) (CA 973/14–1), as well as, by 270,000 core hours on the JUROPA supercomputer in the Jülich Supercomputing Center through the NIC program (NIC: 5720 and 6695). TFO is supported by the DFG Center for Nanoscale Microscopy and Molecular Physiology of the Brain. S.V. has been granted an ICREA ACADEMIA award. TFO and SV are supported by a grant from Fundación La Marato de TV3 (Ref. 20144330).
References
- McCann H, Stevens CH, Cartwright H, Halliday GM. α-Synucleinopathy phenotypes. Park Relat Disord 2014; 20:S62-7; http://dx.doi.org/10.1016/S1353-8020(13)70017-8
- Santner A, Uversky VN. Metalloproteomics and metal toxicology of alpha-synuclein. Metallomics 2010; 2:378-92; PMID:21072383; http://dx.doi.org/10.1039/b926659c
- Petrucci S, Ginevrino M, Valente EM. Phenotypic spectrum of alpha-synuclein mutations: New insights from patients and cellular models. Parkinsonism Relat Disord 2016; 22:S16-20; PMID:26341711; http://dx.doi.org/10.1016/j.parkreldis.2015.08.015
- Villar-Piqué A, Lopes da Fonseca T, Sant'Anna R, Szegö ÉM, Fonseca-Ornelas L, Pinho R, Carija A, Gerhardt E, Masaracchia C, Abad Gonzalez E, et al. Environmental and genetic factors support the dissociation between α-synuclein aggregation and toxicity. Proc Natl Acad Sci USA 2016; In press. http://www.pnas.org/content/early/2016/10/04/1606791113.abstract
- Khalaf O, Fauvet B, Oueslati A, Dikiy I, Mahul-Mellier AL, Ruggeri FS, Mbefo MK, Vercruysse F, Dietler G, Lee SJ, et al. The H50Q mutation enhances alpha-synuclein aggregation, secretion, and toxicity. J Biol Chem 2014; 289:21856-76; PMID:24936070; http://dx.doi.org/10.1074/jbc.M114.553297
- Rutherford NJ, Moore BD, Golde TE, Giasson BI. Divergent effects of the H50Q and G51D SNCA mutations on the aggregation of α-synuclein. J Neurochem 2014; 131:859-67; PMID:24984882; http://dx.doi.org/10.1111/jnc.12806
- Ghosh D, Mondal M, Mohite GM, Singh PK, Ranjan P, Anoop A, Ghosh S, Jha NN, Kumar A, Maji SK. The parkinson's disease-associated H50Q mutation accelerates α-synuclein aggregation in vitro. Biochemistry 2013; 52:6925-7; PMID:24047453; http://dx.doi.org/10.1021/bi400999d
- Uversky VN, Li J, Fink AL. Metal-triggered structural transformations, aggregation, and fibrillation of human alpha-synuclein: A possible molecular link between parkinson's disease and heavy metal exposure. J Biol Chem 2001; 276:44284-96; PMID:11553618; http://dx.doi.org/10.1074/jbc.M105343200
- Dudzik CG, Walter ED, Millhauser GL. Coordination features and affinity of the Cu(2+) site in the alpha-synuclein protein of Parkinson's disease. Biochemistry 2011; 50:1771-7; PMID:21319811; http://dx.doi.org/10.1021/bi101912q
- Sung YH, Rospigliosi C, Eliezer D. NMR mapping of copper binding sites in alpha-synuclein. Biochim Biophys Acta 2006; 1764:5-12; PMID:16338184; http://dx.doi.org/10.1016/j.bbapap.2005.11.003
- Ahmad A, Burns CS, Fink AL, Uversky VN. Peculiarities of copper binding to alpha-synuclein. J Biomol Struct Dyn 2012; 29:825-42; PMID:22208282; http://dx.doi.org/10.1080/073911012010525023
- Binolfi A, Rodriguez EE, Valensin D, D'Amelio N, Ippoliti E, Obal G, Duran R, Magistrato A, Pritsch O, Zweckstetter M, et al. Bioinorganic chemistry of Parkinson's disease: Structural determinants for the copper-mediated amyloid formation of alpha-synuclein. Inorg Chem 2010; 49:10668-79; PMID:20964419; http://dx.doi.org/10.1021/ic1016752
- Rose F, Hodak M, Bernholc J. Mechanism of copper(II)-induced misfolding of Parkinson's disease protein. Sci Rep 2011; 1:11; PMID:22355530; http://dx.doi.org/10.1038/srep00011
- Rasia RM, Bertoncini CW, Marsh D, Hoyer W, Cherny D, Zweckstetter M, Griesinger C, Jovin TM, Fernández CO. Structural characterization of copper(II) binding to alpha-synuclein: Insights into the bioinorganic chemistry of Parkinson's disease. Proc Natl Acad Sci U S A 2005; 102:4294-9; PMID:15767574; http://dx.doi.org/10.1073/pnas.0407881102
- Chi YC, Armstrong GS, Jones DNM, Eisenmesser EZ, Liu CW. Residue histidine 50 plays a key role in protecting α-Synuclein from aggregation at physiological pH. J Biol Chem 2014; 289:15474-81; PMID:24742669; http://dx.doi.org/10.1074/jbc.M113.544049
- Chiera NM, Rowinska-Zyrek M, Wieczorek R, Guerrini R, Witkowska D, Remelli M, Kozlowski H. Unexpected impact of the number of glutamine residues on metal complex stability. Metallomics 2013; 5:214-21; PMID:23370132; http://dx.doi.org/10.1039/c3mt20166j
- De Ricco R, Valensin D, Dell'Acqua S, Casella L, Dorlet P, Faller P, Hureau C. Remote His50 acts as a coordination switch in the high-affinity N-terminal centered copper(II) Site of α-Synuclein. Inorg Chem 2015; 54:4744-51; PMID:25926427; http://dx.doi.org/10.1021/acs.inorgchem.5b00120
- Mason RJ, Paskins AR, Dalton CF, Smith DP. Copper binding and subsequent aggregation of alpha-synuclein are modulated by N-terminal acetylation and ablated by the H50Q missense mutation. Biochemistry 2016; 55:4737-41; PMID:27517125; http://dx.doi.org/10.1021/acs.biochem.6b00708
- Proukakis C, Dudzik CG, Brier T, MacKay DS, Cooper JM, Millhauser GL, Houlden H, Schapira AH. A novel alpha-synuclein missense mutation in Parkinson disease. Neurology 2013; 80:1062-4; PMID:23427326; http://dx.doi.org/10.1212/WNL.0b013e31828727ba
- Valensin D, Camponeschi F, Luczkowski M, Baratto MC, Remelli M, Valensin G, Kozlowski H. The role of His-50 of alpha-synuclein in binding Cu(II): pH dependence, speciation, thermodynamics and structure. Metallomics 2011; 3(3):292-302; PMID:21212878; http://dx.doi.org/10.1039/c0mt00068j
- Laio A, VandeVondele J, Rothlisberger U. A Hamiltonian electrostatic coupling scheme for hybrid Car-Parrinello molecular dynamics simulations. J Chem Phys 2002; 116(16):6941-6947