
SUMMARY
Colorectal cancer (CRC) is the second most common cause of cancer deaths in men and women combined. Colon-tumor growth is multistage and the result of the accumulation of spontaneous mutations and epigenetic events that silence tumor-suppressor genes and activate oncogenes. Environmental factors are primary contributors to these somatic gene alterations, which account for the increase in incidence of CRC in western countries. In recent decades, gut microbiota and their metabolites have been recognized as essential contributing factors to CRC, and now serve as biomarkers for the diagnosis and prognosis of CRC. In the present review, we highlight holistic approaches to understanding how gut microbiota contributes to CRC. We particularly focus herein on bacteria-related changes in host DNA methylation and the risk for CRC.
KEYWORDS:
Introduction
Colorectal carcinoma (CRC) is one of the three most common cancers, with more than 1.2 million new cases and about 600,000 deaths worldwide every year.Citation1 CRC tumors result from an accumulation of DNA alterations that alter key signaling pathways. This forms part of a multistage carcinogenesis process that involves epithelial hyperproliferation, development of aberrant crypt foci (ACF), adenoma to carcinoma transition, tumor invasion, and metastasis. Most CRCs are of a sporadic nature and less than 5% are hereditary,Citation2 which suggests that environmental factors are the common underlying cause of CRC carcinogenesis, rather than inherited genetic defects. Environmental exposure is considered an underlying feature of the increasing prevalence of sporadic CRCs in the western world. Although they have been investigated for a long time, exhaustive analysis of environmental factors is a daunting task and appears almost impossible. A major breakthrough has come with the association of gut microbiota (GM) to CRC. The causative role of GM in disease was first illustrated with mouse models, in studies of obesity and nutrition.Citation3,Citation4 A community of stool bacteria from obese mice that had been submitted to hyperphagia transmitted obesity to germ-free recipients while they were submitted to normophagia.Citation4 GM has now become a focus of investigations as biomarkers for both host and bacteria adaptation to the environment. Ongoing research has linked GM with various diseases, including diabetes, inflammatory bowel disease (IBD), and CRC. This review analyzes GM as a novel tool for studying colon carcinogenesis, focusing on changes in host DNA methylation as a specific etiologic mechanism.
Mutations in CRC
The genetic basis of cancer was firmly established by the discovery and cloning of the adenomatous polyposis coli (APC) gene, whose loss of function is likely the most common cancer initiator in CRC.Citation5–9 This was followed by the recognition that CRC carcinogenesis involves several genetic events and is a multistage process.Citation10,Citation11 Individuals that are born with a heterozygous APC deficiency develop familial polyposis (FAP), which, if left untreated, develops into colon cancer. A second form of hereditary colon cancer is caused by defects in the DNA mismatch repair system (MMR). Heritable MMR gene mutations are the primary cause of hereditary nonpolyposis colorectal cancer (HNPCC), which is known as Lynch syndrome.Citation12 However, both APC and MMR gene silencing similarly contribute to sporadic CRC development.
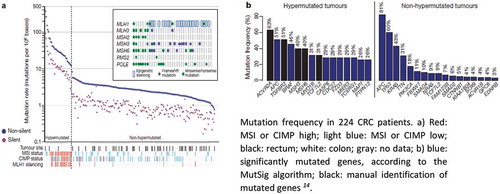
Several critical genes and pathways that are essential in CRC initiation and progression have now been described. Accordingly, consensus molecular CRC subtypes have also been described,Citation13 and a cancer genome atlas on CRC has been established.Citation14 Overall, 24 host genes frequently mutate in CRC, including APC, TP53, SMAD4, PIK3CA, KRAS, ARID1A, SOX9, FAM123B, TGF-β, and DCC. Additionally, MMR genes exhibit recurrent copy-number alterations with drug-targetable amplifications of ERBB2, amplification of IGF2, and recurrent chromosomal translocations with the fusion of NAV2 and WNT-pathway member TCF7L1. Integrative analyses suggest new markers for aggressive CRC and an essential role for MYC-directed transcriptional activation and repression. Particularly, several of these DNA mutations can be detected in precancerous lesions (). Based on mutation rates within tumor tissues, the Cancer Genome Atlas projectCitation14 divided CRCs into two groups: those with a mutation rate of <8.24 per 106 (84% of cases, which are designated hypomutated) and remaining cases with mutation rates of >12 per 106 (which are designated hypermutated) (). Overall, 16% of CRCs are hypermutated, and most of these fall into the category of tumors with microsatellite instability (MSI), which is either due to mutations in the MMR or promoter methylation of the MLH1 gene (CIMP).Citation15,Citation16 By contrast, the majority of CRC tumors have markedly fewer mutations but exhibit chromosomal instability (CIN), which is primarily caused by hereditary or spontaneous inactivating mutations in the APC gene.
Table 1. Colorectal carcinogenic pathways and main genetic alterations.
Figure 1. Frequency and type of mutations in human CRC tissues.
DNA Methylation as a marker of global health and mortality
DNA methylation in palindromic CpG dinucleotides (CpG islands) plays a pivotal role in mammalian genomic stability and gene expression.Citation17 In normal physiology, de novo methylation events are secondary to gene repression. They serve as a long-term memory of earlier gene expression decisions that have defined cell fate and function, which are intended to prevent gene reactivation in subsequent cell generations.Citation18,Citation19 By contrast, the active demethylation of gene regulatory sequences is an integral component of the mechanisms that drive cellular differentiation through gene expression.Citation20,Citation21
The covalent bonding of methyl groups at the five positions of cytosine (5mC) is catalyzed by DNA methyltransferases (DNMTs). The addition of methyl groups to unmethylated cytosines, de novo methylation, is catalyzed by DNMT3 family enzymes. While de novo methylation of CpG islands in gene promoters results in gene silencing, intergenic methylation controls spurious transcription initiation and regulates alternate promoter usage and RNA alternative splicing, which often enhance gene expression. After establishing DNA methylation, the methylation pattern is maintained during DNA replication by the maintenance DNA methyltransferase DNMT1, through a process that is guided by hemi-methylated DNA and histone marks. DNMT1 plays a critical role in preserving epigenetic memory during cell division. The removal of methylation is important for cellular differentiation and is mediated by ten-eleven translocation (TET) proteins.Citation20,Citation21 Active genome-wide DNA demethylation allows for the pluripotency of germ cells during the early stages of embryonic development, which is followed by remethylation prior to birth in males and after birth in females.Citation17 Likewise, TET proteins and global demethylation have been linked to the dedifferentiation process in cancer.Citation22 Passive demethylation occurs when the DNMT1 function is inhibited during DNA replication, or when there is a shortage of cofactors like S-adenosylmethyonine (SAM).
Based on thorough comparative analysis of different CpG island sets from different tissues and age spectra, DNA methylation has emerged as a promising estimate of age and age-related conditions.Citation23,Citation24 The aging process has been associated with what appears to be stochastic changes in patterns of DNA methylation in multiple tissues, which begin in the embryo and slowly continue over time. This process includes de novo methylation of the CpG island set that is bound by the polycomb repressor and demethylation of other large areas of the genome, which is reviewed by Dor and Cedar.Citation24 By comparing methylated loci under physiological and pathological conditions from 30 different tissues and cell types from adults and children, HorvathCitation25 demonstrated that DNA methylation events occur in a highly specific manner, and form tissue-specific patterns of biological aging. Similarly, HannumCitation26 derived a highly accurate age estimator on the basis of 71 CpGs from adult blood DNA, which became a predictor of human health commonly known as “Hannum’s clock”. Subsequently, Horvath used a weighted average of 10 clinical blood biomarkers of chronological age to select blood CpGs that estimate biological age and predict mortality.Citation27
As an output, methylation markers can be used to estimate disease-related age and mortality. Indeed, all-cause mortality is widely studied in association with DNA methylation age. In a study of 5,124 older individuals from the Northeast US, the Framingham Heart Study demonstrated that methylation-related age was associated with mortality,Citation28 which was corroborated by newer studies.Citation29,Citation30 Additionally, a study of 86 Danish twins found that the twin with the higher DNA methylation age at baseline displayed a 69% probability of dying first, over a 10-year follow-up period. In a German population-based cohort, 5-year higher DNA methylation age was associated with a 23% (CI = 1.10–1.38) and 10% (CI = 0.94–1.29) increased risk of all-cause mortality, respectively, after adjustment for technical and clinical confounders.Citation31.
Thus, applying these age estimators to the general population can predict outliers; that is to say, it can identify individuals that are associated with a greater number of age-related conditions and diseases, including cancer. Consequently, DNA methylation-based measures of biological aging have been associated with increased cancer risk and shorter cancer survival, independently of major health risk factors.Citation32,Citation33 Since DNA methylation levels that are associated with cellular senescence relate to cancer occurrence, the CancerClock predictive model of age was recently proposed, which includes methylations of 282 sites from different tumor samples.Citation34
DNA Methylation in CRC
Changes in DNA methylation patterns and histone modifications play key pathophysiological roles in the etiology of cancer, as discussed below. Hypermethylation silences the expression of tumor-suppressor genes and directly impact cancer initiation and progression. Much of the research on the impact of hypermethylation in cancer has focused on CRC. In 1999, Toyota et al.Citation35 reported that some CRCs display a significantly high frequency of aberrant DNA methylation in specific CpG islands, named the CpG island methylator phenotype (CIMP). Serrated polyps, which are the immediate precursors to CIMP tumors, evolve through activation of the MAPK-ERK pathway by BRAF/KRAS mutations and are distinguished by methylation-mediated transcriptional inactivation of various genes that belong to the β-catenin/WNT pathway (SFRP family, CDX2, and MCC), the insulin-like growth factor signaling pathway (IGFBP7), cell-cycle control proteins (CDKN2A), and the DNA mismatch repair (MLH-1) familyCitation36,Citation37, CIMP-positive CRC shows DNA hypermethylation of the MLH1 mismatch repair gene, Citation38 and is highly enriched for the BRAF V600E-activating mutation. BRAF is an upstream activator of the DNMT genes.Citation39 More recent studies indicate that DNMT1, DNMT3B, and EZH2 histone (H3K27) methyltransferase are all upregulated in both hereditary and sporadic MSI tumors.Citation40
Promoter hypermethylation of specific tumor-suppressor genes is emerging as a new biomarker of CRC,Citation41,Citation42 including CDKN2A (p16INK4A and p14ARF)Citation43,Citation44 and APC,Citation45 as reviewed by Okugawa and colleagues.Citation3,Citation6,Citation46 A higher incidence of hypermethylation and a low expression of the secreted frizzled-related proteins (SFRP) genes, which are negative regulators of the WNT pathway, is observed in the normal colonic mucosa of patients with CRC, in comparison with healthy individuals.Citation47 Higher methylation levels of age-related markers, such as ESR1 and MyD88, were observed in the normal colonic mucosa of patients with CIMP-positive CRC, compared to individuals without CRC. Genome-wide DNA methylation analysis has revealed that the gene methylation levels involved in the metabolic pathways of carbohydrates, lipids, and amino acids are significantly different among normal colonic mucosa specimens that are obtained from patients with CRC and controls. Extensive sequencing data obtained from primary and metastatic tumor tissues reveal that genetic mutations alone are insufficient for predicting metastatic potential, while epigenetic changes contribute to the acquisition of additional properties that are necessary for cancer metastasis.Citation48,Citation49 Thus, DNA methylation is emerging as a promising area of investigation for cancer prediction, prevention, and therapy.
Various environmental factors have been investigated for their association with specific patterns of DNA methylation and cancer predisposition or incidence. Among these, chronic inflammation is recognized as a significant inducer of aberrant DNA methylation, as demonstrated by the analysis of non-cancerous tissues, such as colonic mucosae in ulcerative colitis.Citation50 Observations in mouse models are starting to provide mechanistic cause and effect insights into how inflammation and DNA methylation contribute to the etiology of CRC. In a mouse colitis model induced by dextran sodium sulfate, aberrant CpG island methylation in colonic epithelial cells was shown to accumulate gradually,Citation51 and to be heritable.Citation52 Dnmt3b, which is frequently activated in human tumors, promotes tumor development in APCMin/+ mice,Citation53,Citation54 establishing a cause and effect relation. Interestingly, the transgenic expression of Dnmt3b, in the mouse colon initiates de novo DNA methylation of genes that resemble those becoming methylated in human colon cancer. This indicates that aberrant methylation in cancer targets specific sequences and is not random.Citation55 In our recent studies, described in more detail below, transfer of gut microbiota from CRC patients to germ free mice produced similar changes in methylation patterns of oncogenes and tumor suppressor genes in the mouse colon.Citation56 Drugs for the modulation of DNA methylation have also shown preclinical promise in terms of slowing tumor progression.Citation57 Thus, mouse models are a valuable tool for understanding the link between the environment, and in particular the microbiome, and host methylation events that increase CRC risk and treatment outcomes.
Significance of hypomethylation
DNA hypomethylation refers to the loss of the methyl group in the 5-methycytosine nucleotide. The first evidence that demethylation results in the reactivation of silenced genes in vivo came from the treatment of cultured mammalian cells with the demethylating agent deoxyazacytidine, as well as from mutagenesis-driven selection of temperature-sensitive mutants in DNA hypomethylation.Citation58 There is accumulating evidence that DNA hypomethylation and gene reactivation play a major role in the etiology of aging and cancer. While CpGs, at which methylation significantly associates with transcription (eCpG dinucleotides) show heterogeneous distribution, they demonstrate aberrant hypomethylation in agingCitation59 and cancers.Citation60 In normal physiology, aging is known to reduce global methylation of DNA. Indeed, centenarian DNA has a lower DNA methylation content throughout the genome, with more hypomethylated CpGs in promoter, exonic, intronic, and intergenic regions, compared to newborn DNA.Citation61 Interestingly, this global hypomethylation was associated with a greater level of DNA methylation in the CpG island promoters of a few genesCitation61 that have similar features to the cancer epigenome. Some examples of hypomethylation and gene reactivation in cancer include the loss of imprinting in IGF2 as an early event in CRC carcinogenesis.Citation62 Another example is MAGE gene families, which are physiologically repressed by promoter methylation in normal somatic tissues and activated through promoter hypomethylation in several types of cancers.Citation63 Several other examples of the hypomethylation of key procarcinogenic genes and their overexpression have been reported,Citation64 such as the BCL2 gene in lymphocytic lymphoma, RRAS in gastric cancers, MAGE family genes, and GPR17 in lung, head, and neck cancers.Citation57 The association of DNA hypomethylation with high HIF-1α expression levels has critical implications for energy metabolism, angiogenesis, cell survival, and tumor invasion.Citation57 The HIF-1α promoter is an auto-transactivating gene that enables the HIF-1α protein to bind to its own promoter, which explains why hypomethylation of this gene activates tumor growth. Thus, widespread DNA hypomethylation and focal hypermethylation characterize both aging and cancer epigenomes.
DNA hypomethylation can produce chromosomal instability, loss of imprinting, and the reactivation of endogenous transposable elements and retroviruses.Citation65 However, the experimental outcomes of hypomethylation can vary, depending on the approach used. While treatment with 5-aza-deoxycytidine reduces tumor burden in mouse models of hereditary polyposis,Citation66 DNMT1 deficiency predisposes mice to developing lymphomas.Citation67 Additional studies indicate that, while DNA hypermethylation of specific loci drives CRC tumorigenesis,Citation68,Citation69 genome-wide hypomethylation is associated with chromosomal instability in CRC.Citation70 Thus, mouse models are also beginning to demonstrate the nonrandomness of DNA hypomethylation and its likely association with aging and cancer predisposition (; ).
Table 2. Hallmarks of cancer and examples of genes silenced by aberrant methylation.
Figure 2. Mechanisms of DNA demethylation. (a) Passive demethylation. This process occurs during replication, wherein one or more limiting factors (i.e., compromised DNMT function, absence of SAM) prevents methylation maintenance and results in the subsequent loss of 5mC residues. (b) Active demethylation. The figure shows TET enzymes (TET1, TET2, or TET3) (teal) catalyzing stepwise oxidation of 5mC, which is first converted into 5-hydoxymethylcytosine (5hmC), further oxidized into 5-formylcytosine (5fC), and finally converted into 5-carbocylcytosine (5caC). 5fC and 5caC intermediates can be recognized and removed by thymine DNA glycosylase (TDG) (violet). They are then replaced with an unmethylated cytosine nucleotide to complete the base excision repair (BER) process (figure from ref).Citation57
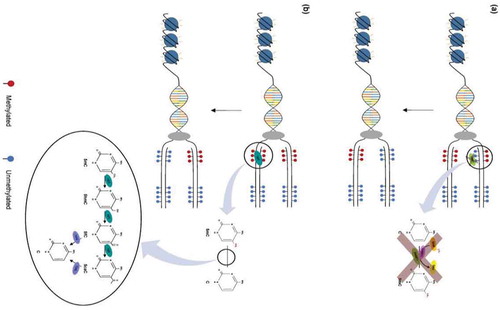
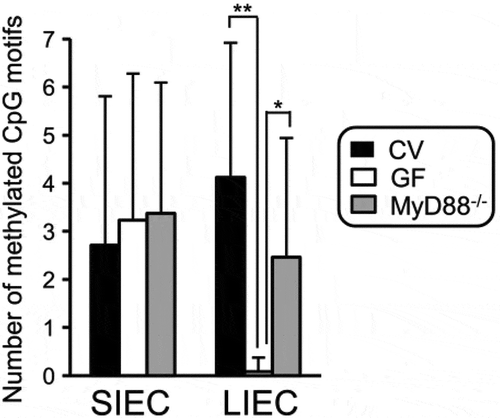
Figure 3. IEC: intestinal epithelial cell; SIEC: small; IECL: IEC large intestine; CV: conventional; GF: germ-free; IEC: intestinal epithelial cell; SIEC:small; LIEC: large intestine; CV:conventional; GF: germ-free; from Takashi et al Ref. Citation94.
Microbes and CRC
The characterization of colonic gut microbiota that is causatively linked to CRC raises two main difficulties. The immense number of microbes (1012 toCitation14/gr) and proportion of anaerobes in the stool make it difficult to culture them all. Thus, GM is usually characterized by using 16S rRNA and whole metagenomic sequencing procedures. We previously showed, by using principal component analyses, that GM from CRC patients and controlsCitation71 display statistically significant separation, as also reported by others.Citation72 However, a variety of bacterial species, not a single microbe, including commensal and virulent bacteria, were associated with tumors. Also, based on the characterization of tissue-adherent bacteria, we found that microbial organization and networking were different in normal and tumor tissues, even in the same individual.Citation71,Citation73 Some microbial species were more frequently present in tumor tissue than in homologous normal tissue throughout the entire progression of a disease, from early tumor development to metastasis.Citation74 At this time, however, cause and effect have not been established.
To understand the mechanisms through which microbiota may be involved in colon carcinogenesis, it was first necessary to clarify the “chicken or egg” relationship. Several groups have shown that unselected bacteria from CRC patients’ microbiota can initiate and/or promote colon carcinogenesis.Citation56,Citation75 As effective causative evidence, PEG, a laxative component that is known to impede the adherence of bacteria to the mucosa, was shown to reduce the development of aberrant Crypt foci (ACF) in experimental mouse models.Citation56 This was also shown in humans, as demonstrated in a randomized double-blind placebo trial.Citation76 In all these studies, the carcinogenic effect of the microbiota was associated with various abnormalities, such as epithelial cell hyperproliferation, inflammatory cytokines release, and inflammatory and immune cell infiltration within the colonic mucosa. In addition, injuries in the mucosa were found to be significantly linked to microbial composition, regardless of whether they were associated with precancerous and cancerous lesions. Such abnormalities within tissues have been observed in the normal macroscopic colonic mucosa of various other emerging diseases, such as obesity, IBD, and insulin resistance.Citation73,Citation77,Citation78 Thus, it became necessary to address how microbial-related effects could be specific to CRC.
Results from studies done based on single bacterium–cell or bacterium–tissue interactions have enriched our knowledge of how microbiota impact host homeostasis or promote pathology. Disruption of cell–cell junctions in the colonic mucosa and enhanced mutations in the host DNA have been attributed to Escherichia coli strains, Bacteroides fragilis, and Fusobacterium nucleatum.Citation78,Citation79However, the putative action of each bacterium remains questionable, due to the large number ofbacteria and difficulties in distinguishing direct and indirect effects, such as DNA damage,chromosomal instability, inflammation, and metabolism.Citation80 One attractive approach to moving from individual species to more complex microbiota is to focus on microbial metabolites, such as H2SCitation81,Citation82 and secondary bile acidsCitation83, both of which can cause DNA damage as procarcinogenic agents and alter immunity.Citation78
Virulent microbes and host DNA mutations
Multiple studies analyzing colonic mucosa samples from CRC patients have reported an association between virulent pro-inflammatory bacteria, such as the pathogenic strains of E. coli, and CRC through the depletion of MMR protein network in host cells. For example, enteropathogenic E. Coli (EPEC) produces the EspF toxin that depletes MMR proteins MSH-2 and MLH-1 in cultured colonic cellsCitation84 Bacteria–cell interaction leads to the rapid accumulation of spontaneous somatic mutations throughout the genome, particularly in long repeated sequences of 1–4 nucleotides, which are termed microsatellites.Citation15,Citation85 In addition to causing microsatellite instability (MSI), MMR disruption enhances the somatic mutation of tumor-suppressor genes, such as APC and P53, which are mutated in the majority of CRCs. Whether EPEC induces somatic mutations in MMR genes themselves has not yet been established.
E. coli is divided into various phylotypes (A, B1, B2, D, E, and F). Although significant levels of E. coli were detected in CRC tissues, compared to healthy colonic tissues, the E. coli from CRC cases (57%) belonged to different phylotypes.Citation86 All Phylotype A E. coli strains are commensal, but B2 is the most prevalent haplotype in feces. Based on virulence factors, all the strains belonging to the phylotypes B2, D (with an exception), E, and F cluster together.Citation87 Whether this is implicated in colon carcinogenesis remains unclear. However, there is a high likelihood that virulent bacteria work as consortia rather than alone. For example, colibactin produced by pks+ E. coli works synergistically with fragilysin a toxin produced by enterotoxigenic Bacteroides fragilis (ETBF), to damage DNA in colonocytes and stimulate pro-inflammatory response in mice.Citation88
Overall, and at the genus level, 20–30 bacterial genera have been found to be significantly different between CRC patients and controls, and at the OTU level, more than 80 genera have marked differences.Citation56,Citation89 However, fewer shifts were observed in a Chinese cohort.Citation90 We reported noticeable shifts in Prevotella/Bacteroides in the stool samples of CRC patients versus controls.Citation71 Lower diversity in CRC patients’ stool was reported in most studies, with enrichment of Fusobacterium, Porphyromonas, P. micra, and B. fragilis.Citation78,Citation79,Citation91 This suggests that, for each of these bacteria, various species may preferentially be linked with CRC, which indicates that it is highly unlikely that a single causative bacterium with one or several mutagenic effects on the colonic mucosa will be identified. It has not yet been clarified whether bacteria overgrowth exerts a direct effect or damages in the colonic mucosa result from chronic inflammation. The growth of virulent bacteria seems to be favored by meat-enriched diets,Citation92 however the impact of tumor growth and changes in immune response on enrichment of such harmful strains needs more investigation.
Commensal habitants and the significance of host DNA methylation
Commensal bacteria inhabit the colon, where they contribute to host nutrition, prevent pathogen colonization, and are involved in the regulation of various physiological functions. DNA methylation is one of the tools by which they influence physiological functions or cause pathology. Exposure to GM drives distinct methylome and transcriptome changes in intestinal epithelial cells during human post-natal development.Citation93 In adults, decreased expression of specific toll-like receptors (TLRs) in colonocytes limits their responsiveness to microbiota, which prevents autoimmunity.Citation79 Increased expression of TLR4 and an elevated response to commensal organisms are observed in patients with IBD.
Takashi et al.Citation94 investigated the methylation level of the TLR4 gene in the intestinal epithelial cells (IEC) of conventional mice (CV) and compared results to those of germ-free (GF) mice that did not have intestinal commensal bacteria. First, the methylation level of the TLR4 gene was significantly lower in the colonocytes of GF mice than in those of CV. Second, DNA methylation in the TLR4 gene promotor in CV mice was higher in IECs than in splenic cells in CV. Third, this methylation was dependent on the differentiation state of the IECs, as the differentiated IEC population display greater methylation and lower expression of the TLR4 gene than the undifferentiated population. Fourth, overexpression of CDX2, which is a transcription factor that is required for the development of the intestine, decreased the methylation level of the TLR4 gene promoter and increased gene expression and responsiveness to TLR4 liganchanges in intestinal epithelial cells during in the small intestine, which does not host commensal bacteria, was similar in both CV and GF mice.Citation94 These findings show that although IECs are continuously exposed to commensal bacteria, they remain relatively insensitive to them, which avoids an excessive inflammatory reaction in the colonic mucosa. Human IEC lines are hyperresponsive to LPS in vitro because of the down-regulation of TLR4 gene expression, which is silenced through several epigenetic mechanisms. By controlling epigenetic modification of the host genes in the large intestine, the commensal bacteria maintain intestinal symbiosis and may contribute to healthy mild inflammation within the mucosa. Thus, epigenetic-mediated mechanisms, including DNA methylation and histone deacetylation, are suggested as the main pathway by which commensal bacteria regulate colonocyte maturation and responsiveness to the microbiome.Citation50
Moreover, altered DNA methylation in gut epithelial cells can have pathological consequences. A well-known example is the hypermethylation of MMR genes that are predisposed to sporadic MSI colon cancer. Hypermethylation of CpG islands in the promotor of hMLH1, which is one of the main functional genes of the MMR system, mimics MMR system failure and resembles Lynch syndrome.Citation95 Thus, we suggest that DNA methylation plays a pivotal role in maintaining symbiosis between the mucosa and commensal bacteria in the intestine regulating host DNA mutations and repair that are caused by invasive and virulent bacteria.
Microbiota-induced methylation and immune response to CRC
Although CRC was the first neoplasia found to be under immune surveillance,Citation96 it is largely resistant to immunotherapy.Citation97 This can be partly explained by the immune suppressive microenvironment of CRC tumors, which includes expression of immune checkpoint moleculesCitation98,Citation99 and a local T-cell suppressive inflammatory infiltrate.Citation100 Paradoxically, DNA hypermutation that occurs in relatively rare hereditary MMR or sporadic MSI tumors improves antitumor immune responses, compared to more frequent CIN tumors, which have fewer mutations. DNA hypermutation generates neoantigensCitation101 and enhances T-cell responses to the tumor, which increases the densities of tumor-infiltrating T-cellsCitation102-104 that respond to mutation. These findings have led to the description of an Immunoscore, which measures tumor-infiltrating T-cell density as a predictive marker of cancer outcome, and its incorporation into the classical CRC tumor staging.Citation105,Citation106 Following this rationale, pharmacologic demethylation of tumor-associated DNA may boost protective T-cell responses through the activation of endogenous retroviruses and cancer-testis antigens, which are normally suppressed in somatic cells. Activation of these genes can give rise to neoantigens in treated cells, which increase the T-cell immunosurveillance capability of the host and thus improve clinical outcomes.Citation107
Changes in the DNA methylation patterns in the blood (PBMC and free circulating DNA) are emerging as a characteristic feature of CRC. In a recent study, we linked changes in methylation patterns of blood mononuclear cells with CRC-associated microbiota.Citation56 We found that several gene promoters, including SFRP1, 2, and 3, PENK, NPY, ALX4, SEPT9, and WIF1, were hypermethylated in the CRC tumoral tissues and blood, compared to healthy donor tissues and blood. Based on these reults, an easily reproducible blood test was developed, and validated in a large cohort of 1,000 individuals, including 187 with advanced adenomas or invasive cancer in their colon and coloscopy, with no significant abnormalities in the remaining cases. The blood test that included methylation levels of WIF1, PENK, and NPY genes, which is called the cumulative methylation index (CMI), was closely associated with both CRC and dysbiosis. Further, CMI appeared to be an independent risk factor for CRC diagnosis, as shown by multivariate analysis that included a fecal immunochemical blood test using a mass-screening program, too. Overall, individuals with higher CMI in their blood were identified as having a greater abundance of pro-epigenomic bacteria in their stools. To relate methylation changes in the host with gut microbiota, we took fresh fecal samples from nine subjects with normal colonoscopy and from nine sex- and age-matched CRC patients, and transferred these microbiota (FMT) into germ-free (GF) mice. To accelerate any oncogenic events produced by the microbiota, separate groups of these mice were treated with low levels of AOM. Fecal and colonic mucosa features were measured, using histopathological and molecular techniques. Between seven and 14 weeks after FMT, CRC-associated microbiota induced higher numbers of aberrant crypt foci (ACF) as compared to healthy donor microbiota. The increase in ACF incidence corresponded to higher levels of hypermethylated genes in the colonic mucosa of the mice that received CRC-microbiota relative to healthy control microbiota recipients, but no significant difference in DNA mutation was observed between the animal groups. Thus, we conclude that the panel of bacteria that is associated with high levels of CMI can be potentially used for screening and therapy surveillance in CRC patients.Citation56
Other independent studies have found correlations between DNA methylation patterns of peripheral blood and incidence of CRC. In one study, the DNA methylation changes were found in genes associated with control of metastasis (AOX-114 and RARB215), angiogenesis (RERG16 and ADAMTS917), and autoimmunity (IRF4 and FOXE-1).Citation108 Hypermutation of FOXE-1 had been earlier reported to be a predictive biomarker of CRC in biopsies obtained by colonoscopy.Citation109 In another study, hypermutated promoter regions of a panel of 30 CRC-associated genes was assayed in circulating DNA and proposed to be a biomarker of CRC.Citation110 To what extent these blood markers are derived from circulating tumor cells, versus PBMC, is yet to be determined. However, changes in the gut microbiota that are associated with distinct pathologies other than cancer, such as obesity, also translate into distinctly altered global peripheral blood mononuclear cells (PBMC) DNA methylation patterns.Citation111 Thus, it is highly likely that microbiota regulate host immunity by altering methylation patterns of circulating immune cells.
It is very tempting to consider how microbiota-induced changes in the methylation patterns of immune cells can contribute to the nature of antitumor or pro-tumor immunity in CRC. GM have been proposed to contribute to immunological failures, which lead to disease predisposition and tumor growthCitation112, as well as to effective immune surveillance and response to therapy.Citation113 Numerous studies have established the role of GM in the maturation of the immune compartment of developing mammals and in determining the nature of immune responses in adults. Indeed, immune response is typically associated with alterations in DNA methylation. For example, the response of cancer patients to neoadjuvant chemotherapyCitation114 or peptide vaccinationCitation115 can be monitored by changes in the patterns of DNA methylation in their PBMC. Altered methylation patterns of circulating CD8+ T-cells have been associated with expression of PD1 and T-cell exhaustion, as reviewed by Emran and colleagues.Citation39 DNA methylation patterns define a population of anergic CD4+ effector T-cells that can differentiate into regulatory T-cells (Tregs).Citation116 A study of the PBMC of pregnant women found an association between the differential methylation of promoter regions of gene associated with cardiovascular disease and Firmicutes and Bacteroidetes but not Proteobacteria.Citation117
Perspectives
We hypothesize that microbes and colonocytes develop adaptive processes to deal with their environment. By characterizing microbiota dysbiosis, we can establish a combined risk factor per individual that includes environmental and host-adaptive processes. The genetic mutations and epigenetic changes including DNA methylation that control gene functions are the main markers of such adaptive processes. Thus, we propose a holistic evaluation of CRC risk and outcomes that is based on individual environment-related risk factors, their impact on GM, and the methylation pattern of host tissues, including blood. Guan et al.Citation118 characterized these risks for breast cancer incidence by evaluating and comparing the predictive performance of whole‐blood DNA methylation and genetic and environment risks in a prospective cohort with up to 14 years of follow‐up. They showed that all three types of risk scores were predictive of breast cancer occurrence. In their study, a genetic risk score that was based on multiple common variants (269 SNPs) predicted cancer incidence with much higher accuracy than either methylation risk that was based on previously identified CpGs or an environmental risk score that was derived from previous studies. They also showed that the combination of three risks enhanced risk prediction, with an “area under the cure” (AUC) statistic value of approximately 0.64. Similar predictive accuracy of either individual or combined risk scores was observed in specific subgroups that were defined by time to diagnosis. We believe that microbiome analysis should improve environmental risk markers and can help to identify CpG methylation targets that relate to cancer risk. Thus, physicians may validate a combination of microbial and molecular (genetic and epigenetic) markers to diagnose tumors and to predict related prognosis.
Whether microbiota exerts a direct or indirect effect on the epigenome remains unclear. Nevertheless, it is likely that much of the impact of microbiota on the tumor is through modulation of the immune response. In addition to bacterial products, such as LPS, the metabolism of nutriments and bile acids by bacteria impact the host’s innate and adaptive immunity. These include short-chain fatty acids (SCFAs), hydrogen sulfide (H2S), and secondary bile acids, all of which alter the genome or epigenome of immune cells.Citation78 The SCFAs (acetate, propionate, and butyrate) enhance the induction of Tregs and certain bile acids, while bile acid metabolites alter inflammation by inducing the expression of the canonical T helper 17 transcription factor and retinoic acid-related orphan receptor gamma (RORγt) in the Tregs,Citation119 and serve as ligands to RORγt.Citation120 Future research will establish the role of bacterial metabolites in determining the fate of immune cells through DNA methylation.
Acknowldegements
We thank Dr. Michael Sadowsky (University of Minnesota) for critical comments and edits. The Minnesota Colorectal Cancer Research Foundation and the National Institute of Health (RO1 AI 108682) are gratefully acknowledged for their support.
References
- Jemal A, Bray F, Center MM, Ferlay J, Ward E, Forman D. Global cancer statistics. CA Cancer J Clin. 2011;61(2):69–15. doi:10.3322/caac.20107.
- Foulkes WD. Inherited susceptibility to common cancers. N Engl J Med. 2008;359(20):2143–2153. doi:10.1056/NEJMra0802968.
- Turnbaugh PJ, Ley RE, Mahowald MA, Magrini V, Mardis ER, Gordon JI. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature. 2006;444(7122):1027–1031. doi:10.1038/nature05414.
- Ley RE, Turnbaugh PJ, Klein S, Gordon JI. Microbial ecology: human gut microbes associated with obesity. Nature. 2006;444(7122):1022–1023. doi:10.1038/4441022a.
- Kinzler KW, Nilbert MC, Su LK, Vogelstein B, Bryan TM, Levy DB, Smith K, Preisinger A, Hedge P, McKechnie D, et al. Identification of FAP locus genes from chromosome 5q21. Science. 1991;253(5020):661–665. doi:10.1126/science.1651562.
- Kinzler KW, Nilbert MC, Vogelstein B, Bryan TM, Levy DB, Smith KJ, Preisinger A, Hedge P, Markham A, et al. Identification of a gene located at chromosome 5q21 that is mutated in colorectal cancers. Science. 1991;251(4999):1366–1370. doi:10.1126/science.1848370.
- Nakamura Y, Nishisho I, Kinzler KW, Vogelstein B, Miyoshi Y, Miki Y, Ando H, Horii A, Nagase H. Mutations of the adenomatous polyposis coli gene in familial polyposis coli patients and sporadic colorectal tumors.. Princess Takamatsu Symposia. 1991;22:285–292.
- Nakamura Y, Nishisho I, Kinzler KW, Vogelstein B, Miyoshi Y, Miki Y, ANDO H, HORII A. Mutations of the APC(Adenomatous Polyposis Coli) Gene in FAP(Familial Polyposis Coli) Patients and in Sporadic Colorectal Tumors.. Tohoku J Exp Med. 1992;168(2):141–147. doi:10.1620/tjem.168.141.
- Powell SM, Zilz N, Beazer-Barclay Y, Bryan TM, Hamilton SR, Thibodeau SN, Vogelstein B, Kinzler KW. APC mutations occur early during colorectal tumorigenesis. Nature. 1992;359(6392):235–237. doi:10.1038/359235a0.
- Vogelstein B, Kinzler KW. The multistep nature of cancer. Trends Genet. 1993;9(4):138–141. doi:10.1016/0168-9525(93)90209-Z.
- Kinzler KW, Vogelstein B. Landscaping the cancer terrain. Science. 1998;280:1036–1037. doi:10.1126/science.280.5366.1036.
- Burton-Chase AM, Hovick SR, Peterson SK, Marani SK, Vernon SW, Amos CI, Frazier ML, Lynch PM, Gritz ER. Changes in screening behaviors and attitudes toward screening from pre-test genetic counseling to post-disclosure in Lynch syndrome families. Clin Genet. 2013;83(3):215–220. doi:10.1111/cge.12091.
- Guinney J, Dienstmann R, Wang X, de Reynies A, Schlicker A, Soneson C, Marisa L, Roepman P, Nyamundanda G, Angelino P. The consensus molecular subtypes of colorectal cancer. Nat Med. 2015;21(11):1350–1356. doi:10.1038/nm.3967.
- The Cancer Genome Atlas Network. Comprehensive molecular characterization of human colon and rectal cancer. Nat Med. 2012;487(7407):330–337. doi:10.1038/nature11252.
- Kane MF, Loda M, Gaida GM, Lipman J, Mishra R, Goldman H, Jessup JM, Kolodner R. Methylation of the hMLH1 promoter correlates with lack of expression of hMLH1 in sporadic colon tumors and mismatch repair-defective human tumor cell lines. Cancer Res. 1997;57:808–811.
- Parsons MT, Buchanan DD, Thompson B, Young JP, Spurdle AB. Correlation of tumour BRAF mutations and MLH1 methylation with germline mismatch repair (MMR) gene mutation status: a literature review assessing utility of tumour features for MMR variant classification. J Med Genet. 2012;49(3):151–157. doi:10.1136/jmedgenet-2011-100714.
- Chen Z, Zhang Y. Role of Mammalian DNA Methyltransferases in Development. Annu Rev Biochem. 2020 Jun 20;89:135–158. doi: 10.1146/annurev-biochem-103019-102815.
- Epsztejn-Litman S, Feldman N, Abu-Remaileh M, Shufaro Y, Gerson A, Ueda J, Deplus R, Fuks F, Shinkai Y, Cedar H. De novo DNA methylation promoted by G9a prevents reprogramming of embryonically silenced genes. Nat Struct Mol Biol. 2008;15(11):1176–1183. doi:10.1038/nsmb.1476.
- Feldman N, Gerson A, Fang J, Li E, Zhang Y, Shinkai Y, Cedar H, Bergman Y. G9a-mediated irreversible epigenetic inactivation of Oct-3/4 during early embryogenesis. Nat Cell Biol. 2006;8(2):188–194. doi:10.1038/ncb1353.
- Orlanski S, Labi V, Reizel Y, Spiro A, Lichtenstein M, Levin-Klein R, Koralov SB, Skversky Y, Rajewsky K, Cedar H. Tissue-specific DNA demethylation is required for proper B-cell differentiation and function. Proc Natl Acad Sci U S A. 2016;113(18):5018–5023. doi:10.1073/pnas.1604365113.
- Lu F, Liu Y, Jiang L, Yamaguchi S, Zhang Y. Role of Tet proteins in enhancer activity and telomere elongation. Genes Dev. 2014;28(19):2103–2119. doi:10.1101/gad.248005.114.
- Abdel-Wahab O, Mullally A, Hedvat C, Garcia-Manero G, Patel J, Wadleigh M, Malinge S, Yao J, Kilpivaara O, Bhat R, et al. Genetic characterization of TET1, TET2, and TET3 alterations in myeloid malignancies. Blood. 2009;114(1):144–147. doi:10.1182/blood-2009-03-210039.
- Horvath S, Raj K. DNA methylation-based biomarkers and the epigenetic clock theory of ageing. Nat Rev Genet. 2018;19(6):371–384. doi:10.1038/s41576-018-0004-3.
- Dor Y, Cedar H. Principles of DNA methylation and their implications for biology and medicine. Lancet. 2018;392(10149):777–786. doi:10.1016/S0140-6736(18)31268-6.
- Horvath S. DNA methylation age of human tissues and cell types. Genome Biology. 1994;145:R115. doi:10.1186/gb-2013-14-10-r115.
- Hannum G, Guinney J, Zhao L, Zhang L, Hughes G, Sadda S, Klotzle B, Bibikova M, Fan J-B, Gao Y, et al. Genome-wide methylation profiles reveal quantitative views of human aging rates. Mol Cell. 2013;49(2):359–367. doi:10.1016/j.molcel.2012.10.016.
- Levine ME, Lu AT, Quach A, Chen BH, Assimes TL, Bandinelli S, Hou L, Baccarelli AA, Stewart JD, Li Y, et al. An epigenetic biomarker of aging for lifespan and healthspan. Aging (Albany NY). 2018;10(4):573–591. doi:10.18632/aging.101414.
- Murabito JM, Rong J, Lunetta KL, Huan T, Lin H, Zhao Q, Freedman JE, Tanriverdi K, Levy D, Larson MG, et al. Cross-sectional relations of whole-blood miRNA expression levels and hand grip strength in a community sample. Aging Cell. 2017;16(4):888–894. doi:10.1111/acel.12622.
- Mills JA, Beach SRH, Dogan M, Simons RL, Gibbons FX, Long JD, et al. A Direct Comparison of the Relationship of Epigenetic Aging and Epigenetic Substance Consumption Markers to Mortality in the Framingham Heart Study. Genes (Basel). 2019 Jan 15;10(1):51. doi: 10.3390/genes10010051.
- Philibert RA, Dogan MV, Mills JA, Long JD. AHRR Methylation is a Significant Predictor of Mortality Risk in Framingham Heart Study. J Insur Med. 2019;48(1):79–89. doi:10.17849/insm-48-1-1-11.1. Epub 2019 Oct 16.
- Dhingra R, Nwanaji-Enwerem JC, Samet M, Ward-Caviness CK. DNA Methylation Age—Environmental Influences, Health Impacts, and Its Role in Environmental Epidemiology. Curr Environ Health Rep. 2019;10:317–327. doi:10.1007/s40572-018-0203-2.
- Zheng Y, Joyce BT, Colicino E, Liu L, Zhang W, Dai Q, Shrubsole MJ, Kibbe WA, Gao T, Zhang Z, et al. Blood Epigenetic Age may Predict Cancer Incidence and Mortality. EBioMedicine. 2016;5:68–73. doi:10.1016/j.ebiom.2016.02.008.
- Dugue P-A, Bassett JK, Joo JE, Jung C-H, Ming Wong E, Moreno-Betancur M, Schmidt D, Makalic E, Li S, Severi G, et al. DNA methylation-based biological aging and cancer risk and survival: pooled analysis of seven prospective studies. Int J Cancer. 2018;142(8):1611–1619. doi:10.1002/ijc.31189.
- Zhu T, Gao Y, Wang J, Li X, Shang S, Wang Y, Guo S, Zhou H, Liu H, Sun D, et al. CancerClock: A DNA Methylation Age Predictor to Identify and Characterize Aging Clock in Pan-Cancer. Front Bioeng Biotechnol. 2019;7:388. doi:10.3389/fbioe.2019.00388.
- Toyota M, Ho C, Ahuja N, KW J, Li Q, Ohe-Toyota M. The proto CpG island methylator phenotype of sessile serrated adenomas/polyps. Epigenetics. 2018;13(10–11):1088–1105. doi:10.1080/15592294.2018.1543504.
- Okugawa Y, Grady WM, Goel A. Epigenetic Alterations in Colorectal Cancer: Emerging Biomarkers. Gastroenterology 2015; 149:1204–25 e12.
- Parker HR, Orjuela S, Martinho Oliveira A, Cereatti F, Sauter M, Heinrich H, et al. The proto CpG island methylator phenotype of sessile serrated adenomas/polyps. Epigenetics 2018; 13:1088–105.
- Cunningham JM, Christensen ER, Tester DJ, Kim CY, Roche PC, Burgart LJ, Thibodeau SN. Hypermethylation of the hMLH1 promoter in colon cancer with microsatellite instability.. Cancer Res. 1998;58:3455–3460.
- Emran AA, Chatterjee A, Rodger EJ, Tiffen JC, Gallagher SJ, Eccles MR, Hersey P. Targeting DNA Methylation and EZH2 Activity to Overcome Melanoma Resistance to Immunotherapy. Trends Immunol. 2019;40(4):328–344. doi:10.1016/j.it.2019.02.004.
- Joensuu EI, Nieminen TT, Lotsari JE, Pavicic W, Abdel-Rahman WM, Peltomaki P. Methyltransferase expression and tumor suppressor gene methylation in sporadic and familial colorectal cancer. Genes Chromosomes Cancer. 2015;54(12):776–787. doi:10.1002/gcc.22289.
- Ng JM, Yu J. Promoter Hypermethylation of Tumour Suppressor Genes as Potential Biomarkers in Colorectal Cancer. Int J Mol Sci. 2015;16(2):2472–2496. doi:10.3390/ijms16022472.
- O’Connell MR, Sarkar S, Luthra GK, Okugawa Y, Toiyama Y, Gajjar AH, Qiu S, Goel A, Singh P. Epigenetic changes and alternate promoter usage by human colon cancers for expressing DCLK1-isoforms: clinical Implications. Scientific Reports. 2015;5(1):14983. doi:10.1038/srep14983.
- Bihl MP, Foerster A, Lugli A, Zlobec I. Characterization of CDKN2A(p16) methylation and impact in colorectal cancer: systematic analysis using pyrosequencing. Journal of Translational Medicine. 2012;10(1):173. doi:10.1186/1479-5876-10-173.
- Esteller M, Tortola S, Toyota M, Capella G, Peinado MA, Baylin SB, Herman JG. Hypermethylation-associated inactivation of p14(ARF) is independent of p16(INK4a) methylation and p53 mutational status.. Cancer Res. 2000;60:129–133.
- Liang T-J, Wang H-X, Zheng -Y-Y, Cao Y-Q, Wu X, Zhou X, Dong S-X. APC hypermethylation for early diagnosis of colorectal cancer: a meta-analysis and literature review. Oncotarget. 2017;8(28):46468–46479. doi:10.18632/oncotarget.17576.
- Baylin SB, Jones PA. A decade of exploring the cancer epigenome — biological and translational implications. Nat Rev Cancer. 2011;11(10):726–734. doi:10.1038/nrc3130.
- Jung G, Hernandez-Illan E, Moreira L, Balaguer F, Goel A. Epigenetics of colorectal cancer: biomarker and therapeutic potential. Nat Rev Gastroenterol Hepatol. 2020;17(2):111–130. doi:10.1038/s41575-019-0230-y.
- Chatterjee A, Rodger EJ, Eccles MR. Epigenetic drivers of tumourigenesis and cancer metastasis. Semin Cancer Biol. 2018;51:149–159. doi:10.1016/j.semcancer.2017.08.004.
- Vogelstein B, Papadopoulos N, Velculescu VE, Zhou S, Diaz LA Jr., Kinzler KW. Cancer genome landscapes. Science. 2013;339(6127):1546–1558. doi:10.1126/science.1235122.
- Howell KJ, Kraiczy J, Nayak KM, Gasparetto M, Ross A, Lee C, Mak TN, Koo B-K, Kumar N, Lawley T. DNA Methylation and Transcription Patterns in Intestinal Epithelial Cells From Pediatric Patients With Inflammatory Bowel Diseases Differentiate Disease Subtypes and Associate With Outcome. Gastroenterology. 2018;154(3):585–598. doi:10.1053/j.gastro.2017.10.007.
- Kellermayer R, Balasa A, Zhang W, Lee S, Mirza S, Chakravarty A, Szigeti R, Laritsky E, Tatevian N, Smith CW, et al. Epigenetic maturation in colonic mucosa continues beyond infancy in mice. Hum Mol Genet. 2010;19(11):2168–2176. doi:10.1093/hmg/ddq095.
- Tschurtschenthaler M, Kachroo P, Heinsen F-A, Adolph TE, Ruhlemann MC, Klughammer J, Offner FA, Ammerpohl O, Krueger F, Smallwood S, et al. Paternal chronic colitis causes epigenetic inheritance of susceptibility to colitis. Scientific Reports. 2016;6(1):31640. doi:10.1038/srep31640.
- Nosho K, Shima K, Irahara N, Kure S, Baba Y, Kirkner GJ, Chen L, Gokhale S, Hazra A, Spiegelman D, et al. DNMT3B expression might contribute to CpG island methylator phenotype in colorectal cancer. Clin Cancer Res. 2009;15(11):3663–3671. doi:10.1158/1078-0432.CCR-08-2383.
- Linhart HG, Lin H, Yamada Y, Moran E, Steine EJ, Gokhale S, Lo G, Cantu E, Ehrich M, He T, et al. Dnmt3b promotes tumorigenesis in vivo by gene-specific de novo methylation and transcriptional silencing. Genes Dev. 2007;21(23):3110–3122. doi:10.1101/gad.1594007.
- Steine EJ, Ehrich M, Bell GW, Raj A, Reddy S, van Oudenaarden A,
- Sobhani I, Bergsten E, Couffin S, Amiot A, Nebbad B, Barau C, et al. Colorectal cancer-associated microbiota contributes to oncogenic epigenetic signatures. Proc Natl Acad Sci U S A. 2019;116(48):24285–24295. doi:10.1073/pnas.1912129116.
- Urbano A, Smith J, Weeks RJ, Chatterjee A. Gene-Specific Targeting of DNA Methylation in the Mammalian Genome. Cancers (Basel). 2019 Oct 9;11(10):1515. doi:10.3390/cancers11101515.
- Gounari F, Banks GR, Khazaie K, Jeggo PA, Holliday R. Gene reactivation: a tool for the isolation of mammalian DNA methylation mutants.. Genes Dev. 1987;1(9):899–912. doi:10.1101/gad.1.9.899.
- Unnikrishnan A, Hadad N, Masser DR, Jackson J, Freeman WM, Richardson A. Revisiting the genomic hypomethylation hypothesis of aging. Ann N Y Acad Sci. 2018;1418(1):69–79. doi:10.1111/nyas.13533.
- Ehrlich M. DNA hypomethylation in cancer cells. Epigenomics. 2009;1(2):239–259. doi:10.2217/epi.09.33.
- Heyn H, Li N, Ferreira HJ, Moran S, Pisano DG, Gomez A, Diez J, Sanchez-Mut JV, Setien F, Carmona FJ, et al. Distinct DNA methylomes of newborns and centenarians. Proc Natl Acad Sci U S A. 2012;109(26):10522–10527. doi:10.1073/pnas.1120658109.
- Cheng Y-W, Idrees K, Shattock R, Khan SA, Zeng Z, Brennan CW, Paty P, Barany F. Loss of imprinting and marked gene elevation are 2 forms of aberrant IGF2 expression in colorectal cancer. Int J Cancer. 2010;127(3):568–577. doi:10.1002/ijc.25086.
- Weon JL, Potts PR. The MAGE protein family and cancer. Curr Opin Cell Biol. 2015;37:1–8. doi:10.1016/j.ceb.2015.08.002.
- Kidane D, Chae WJ, Czochor J, Eckert KA, Glazer PM, Bothwell AL, Sweasy JB. Interplay between DNA repair and inflammation, and the link to cancer. Crit Rev Biochem Mol Biol. 2014;49(2):116–139. doi:10.3109/10409238.2013.875514.
- Esteller M. Epigenetics in cancer. N Engl J Med. 2008;358(11):1148–1159. doi:10.1056/NEJMra072067.
- Laird PW, Jackson-Grusby L, Fazeli A, Dickinson SL, Jung WE, Li E, Weinberg RA, Jaenisch R. Suppression of intestinal neoplasia by DNA hypomethylation. Cell. 1995;81(2):197–205. doi:10.1016/0092-8674(95)90329-1.
- Gaudet F. Induction of tumors in mice by genomic hypomethylation. Science. 2003;300(5618):489–492. doi:10.1126/science.1083558.
- Sakai E. Accumulation of aberrant DNA methylation during colorectal cancer development. World J Gastroenterol. 2014;20(4):978–987. doi:10.3748/wjg.v20.i4.978.
- Sun D, Chen Y, Fang J-Y. Influence of the microbiota on epigenetics in colorectal cancer. National Science Review. 2019;6(6):1138–1148. doi:10.1093/nsr/nwy160.
- Suter CM, Martin DI, Ward RL. Hypomethylation of L1 retrotransposons in colorectal cancer and adjacent normal tissue. Int J Colorectal Dis. 2004;19(2):95–101. doi:10.1007/s00384-003-0539-3.
- Sobhani I, Tap J, Roudot-Thoraval F, Roperch JP, Letulle S, Langella P, Corthier G, Van Nhieu JT, Furet JP, et al. Microbial dysbiosis in colorectal cancer (CRC) patients. PLoS One. 2011;6(1):e16393. doi:10.1371/journal.pone.0016393.
- Duvallet C, Gibbons SM, Gurry T, Irizarry RA, Alm EJ. Meta-analysis of gut microbiome studies identifies disease-specific and shared responses. Nat Commun. 2017;8(1):1784. doi:10.1038/s41467-017-01973-8.
- Sobhani I, Amiot A, Le Baleur Y, Levy M, Auriault ML, Van Nhieu JT, et al. Microbial dysbiosis and colon carcinogenesis: could colon cancer be considered a bacteria-related disease? Therapeutic advances in gastroenterology 2013; 6:215–229.
- Nakatsu G, Li X, Zhou H, Sheng J, Wong SH, Wu WK, et al. Gut mucosal microbiome across stages of colorectal carcinogenesis. Nat Commun. 2015;6(1):8727. doi:10.1038/ncomms9727.
- Wong SH, Zhao L, Zhang X, Nakatsu G, Han J, Xu W, Xiao X, Kwong TNY, Tsoi H, Wu WKK, et al. Gavage of Fecal Samples From Patients With Colorectal Cancer Promotes Intestinal Carcinogenesis in Germ-Free and Conventional Mice. Gastroenterology. 2017;153(6):1621–33 e6. doi:10.1053/j.gastro.2017.08.022.
- Remya KR, Chandran S, Mani S, John A, Ramesh P. Hybrid polycaprolactone/polyethylene oxide scaffolds with tunable fiber surface morphology, improved hydrophilicity and biodegradability for bone tissue engineering applications. J Biomater Sci Polym Ed. 2018;29(12):1444–1462. doi:10.1080/09205063.2018.1465664.
- Sandler RS, Baron JA, Tosteson TD, Mandel JS, Haile RW. Rectal mucosal proliferation and risk of colorectal adenomas: results from a randomized controlled trial.. Cancer Epidemiol Biomarkers Prev. 2000;9:653–656.
- Kostic AD, Chun E, Robertson L, Glickman JN, Gallini CA, Michaud M, Clancy T, Chung D, Lochhead P, Hold G, et al. Fusobacterium nucleatum potentiates intestinal tumorigenesis and modulates the tumor-immune microenvironment. Cell Host Microbe. 2013;14(2):207–215. doi:10.1016/j.chom.2013.07.007.
- Allen J, Sears CL. Impact of the gut microbiome on the genome and epigenome of colon epithelial cells: contributions to colorectal cancer development. Genome Med. 2019;11(1):11. doi:10.1186/s13073-019-0621-2.
- Rubinstein MR, Wang X, Liu W, Hao Y, Cai G, Han YW. Fusobacterium nucleatum Promotes Colorectal Carcinogenesis by Modulating E-Cadherin/β-Catenin Signaling via its FadA Adhesin. Cell Host Microbe. 2013;14(2):195–206. doi:10.1016/j.chom.2013.07.012.
- Attene-Ramos MS, Nava GM, Muellner MG, Wagner ED, Plewa MJ, Gaskins HR. DNA damage and toxicogenomic analyses of hydrogen sulfide in human intestinal epithelial FHs 74 Int cells.. Environmental and Molecular Mutagenesis. 2010;51(4):304–314. doi:10.1002/em.20546.
- Yazici C, Wolf PG, Kim H, Cross TWL, Vermillion K, Carroll T, et al. Methylation of the hMLH1 promoter correlates with lack of expression of hMLH1 in sporadic colon tumors and mismatch repair-defective human tumor cell lines. Gut. 2017 Nov;66(11):1983-1994. doi:10.1136/gutjnl-2016-313321.
- Ridlon JM, Wolf PG, Gaskins HR. Taurocholic acid metabolism by gut microbes and colon cancer. Gut Microbes. 2016;7(3):201–215. doi:10.1080/19490976.2016.1150414.
- Maddocks OD, Scanlon KM, Donnenberg MS. An Escherichia coli effector protein promotes host mutation via depletion of DNA mismatch repair proteins. MBio. 2018;19:e00152–13. doi:10.1128/mBio.00152-13.
- Hackman P, Tannergard P, Osei-Mensa S, Chen J, Kane MF, Kolodner R, Lambert B, Hellgren D, Lindblom A. A human compound heterozygote for two MLH1 missense mutations. Nat Genet. 1997;17(2):135–136. doi:10.1038/ng1097-135.
- Raisch J. Colon cancer-associated B2 Escherichia coli colonize gut mucosa and promote cell proliferation. World J Gastroenterol. 2014;20(21):6560–6572. doi:10.3748/wjg.v20.i21.6560.
- Wassenaar TM. E. Coli and Colorectal Cancer: A Complex Relationship that Deserves a Critical Mindset. Crit Rev Microbiol. 2018 Sep;44(5):619–632. doi:10.1080/1040841X.2018.1481013.
- Dejea CM, Fathi P, Craig JM, Boleij A, Taddese R, Geis AL, Wu X, DeStefano Shields CE, Hechenbleikner EM, Huso DL, et al. Patients with familial adenomatous polyposis harbor colonic biofilms containing tumorigenic bacteria. Science. 2018;359(6375):592–597. doi:10.1126/science.aah3648.
- Zeller G, Tap J, Voigt AY, Sunagawa S, Kultima JR, Costea PI, Amiot A, Böhm J, Brunetti F, Habermann N, et al. Potential of fecal microbiota for early-stage detection of colorectal cancer. Molecular Systems Biology. 2014;10(11):766. doi:10.15252/msb.20145645.
- Wang T, Cai G, Qiu Y, Fei N, Zhang M, Pang X, et al. Structural segregation of gut microbiota between colorectal cancer patients and healthy volunteers. The ISME journal 2012;6:320–329.
- O'Keefe SJ, Li JV, Lahti L, Ou J, et al. Fat, fibre and cancer risk in African Americans and rural Africans. Nat Commun. 2015;6(1):6342. doi:10.1038/ncomms7342.
- Pan W-H, Sommer F, Falk-Paulsen M, Ulas T, Best P, Fazio A, Kachroo P, Luzius A, Jentzsch M, Rehman A, et al. Exposure to the gut microbiota drives distinct methylome and transcriptome changes in intestinal epithelial cells during postnatal development. Genome Med. 2018;10(1):27. doi:10.1186/s13073-018-0534-5.
- Abreu MT, Vora P, Faure E, Thomas LS, Arnold ET, Arditi M. Decreased expression of Toll-like receptor-4 and MD-2 correlates with intestinal epithelial cell pro-tection against dysregulated proinflammatory gene expression in response to bacterial lipopolysaccharide. J Immunol. 2001;167(3):1609–1616. doi: 10.4049/jimmunol.167.3.1609.
- Takahashi K, Sugi Y, Nakano K, Tsuda M, Kurihara K, Hosono A, Kaminogawa S. Epigenetic control of the host gene by commensal bacteria in large intestinal epithelial cells. J Biol Chem. 2011;286(41):35755–35762. doi:10.1074/jbc.M111.271007.
- Liu B, Nicolaides NC, Markowitz S, Willson JK, Parsons RE, Jen J, Papadopolous N, Peltomäki P, de la Chapelle A, Hamilton SR. Mismatch repair gene defects in sporadic colorectal cancers with microsatellite instability. Nat Genet. 1995;9(1):48–55. doi:10.1038/ng0195-48.
- Galon J, Costes A, Sanchez-Cabo F, et al. Type, density, and location of immune cells within human colorectal tumors predict clinical outcome. Science 2006; 313:1960-4.Sep 29;313(5795):1960–functional genes of the MMR system, mimics M1964. doi:10.1126/science.1129139.
- Kroemer G, Galluzzi L, Zitvogel L, Fridman WH. Colorectal cancer: the first neoplasia found to be under immunosurveillance and the last one to respond to immunotherapy? Oncoimmunology. 2015;4(7):e1058597. doi:10.1080/2162402X.2015.1058597.
- Llosa NJ, Cruise M, Tam A, Wicks EC, Hechenbleikner EM, Taube JM, Blosser RL, Fan H, Wang H, Luber BS, et al. The vigorous immune microenvironment of microsatellite instable colon cancer is balanced by multiple counter-inhibitory checkpoints. Cancer Discov. 2015;5(1):43–51. doi:10.1158/2159-8290.CD-14-0863.
- Masugi Y, Nishihara R, Yang J, Mima K, da Silva A, Shi Y, Inamura K, Cao Y, Song M, Nowak JA, et al. Tumour CD274 (PD-L1) expression and T cells in colorectal cancer. Gut. 2017;66(8):1463–1473. doi:10.1136/gutjnl-2016-311421.
- Lasry A, Zinger A, Ben-Neriah Y. Inflammatory networks underlying colorectal cancer. Nat Immunol. 2016;17(3):230–240. doi:10.1038/ni.3384.
- Maby P, Tougeron D, Hamieh M, Mlecnik B, Kora H, Bindea G, Angell HK, Fredriksen T, Elie N, Fauquembergue E, et al. Correlation between Density of CD8+ T-cell Infiltrate in Microsatellite Unstable Colorectal Cancers and Frameshift Mutations: A Rationale for Personalized Immunotherapy. Cancer Res. 2015;75(17):3446–3455. doi:10.1158/0008-5472.CAN-14-3051.
- Kim H, Jen J, Vogelstein B, Hamilton SR. Clinical and pathological characteristics of sporadic colorectal carcinomas with DNA replication errors in microsatellite sequences.. Am J Pathol. 1994;145:148–156.
- Smyrk TC, Watson P, Kaul K, Lynch HT. Tumor-infiltrating lymphocytes are a marker for microsatellite instability in colorectal carcinoma. Cancer. 2001;91(12):2417–2422. doi:10.1002/1097-0142(20010615)91:12<2417::AID-CNCR1276>3.0.CO;2-U.
- Dolcetti R, Viel A, Doglioni C, Russo A, Guidoboni M, Capozzi E, Vecchiato N, Macrì E, Fornasarig M, Boiocchi M, et al. High prevalence of activated intraepithelial cytotoxic T lymphocytes and increased neoplastic cell apoptosis in colorectal carcinomas with microsatellite instability. Am J Pathol. 1999;154(6):1805–1813. doi:10.1016/S0002-9440(10)65436-3.
- Mlecnik B, Tosolini M, Kirilovsky A, Berger A, Bindea G, Meatchi T, Bruneval P, Trajanoski Z, Fridman W-H, Pagès F, et al. Histopathologic-based prognostic factors of colorectal cancers are associated with the state of the local immune reaction. J Clin Oncol. 2011;29(6):610–618. doi:10.1200/JCO.2010.30.5425.
- Angell H, Galon J. From the immune contexture to the Immunoscore: the role of prognostic and predictive immune markers in cancer. Curr Opin Immunol. 2013;25(2):261–267. doi:10.1016/j.coi.2013.03.004.
- Hardy MP, Vincent K, Perreault C. The Genomic Landscape of Antigenic Targets for T Cell-Based Leukemia Immunotherapy. Front Immunol. 2019;10:2934. doi:10.3389/fimmu.2019.02934.
- Luo X, Huang R, Sun H, Liu Y, Bi H, Li J, Yu H, Sun J, Lin S, Cui B, et al. Methylation of a panel of genes in peripheral blood leukocytes is associated with colorectal cancer. Scientific Reports. 2016;6(1):29922. doi:10.1038/srep29922.
- Papadia C, Louwagie J, Del Rio P, Grooteclaes M, Coruzzi A, Montana C, Novelli M, Bordi C, De’ Angelis GL, Bassett P, et al. FOXE1 and SYNE1 genes hypermethylation panel as promising biomarker in colitis-associated colorectal neoplasia. Inflamm Bowel Dis. 2014;20(2):271–277. doi:10.1097/01.MIB.0000435443.07237.ed.
- Rasmussen SL, Krarup HB, Sunesen KG, Johansen MB, Stender MT, Pedersen IS. Hypermethylated DNA, a circulating biomarker for colorectal cancer detection. PLoS One. 2017;12:e0180809.
- Ramos-Molina B, Sanchez-Alcoholado L, Cabrera-Mulero A, Lopez-Dominguez R, CarmonaSaez P, Garcia-Fuentes E, et al. Gut Microbiota Composition Is Associated With the Global DNA Methylation Pattern in Obesity. Front Genet. 2019;10:613. doi:10.3389/fgene.2019.00613
- Hurtado CG, Wan F, Housseau F, Sears CL. Roles for Interleukin 17 and Adaptive Immunity in Pathogenesis of Colorectal Cancer. Gastroenterology 2018;155:1706–15 6 doi:10.1053/j.gastro.2018.08.056
- Viaud S, Saccheri F, Mignot G, Yamazaki T, Daillere R, Hannani D, et al. The intestinal microbiota modulates the anticancer immune effects of cyclophosphamide. Science 2013; 342:971–6
- Hsu P-C, Kadlubar SA, Siegel ER, Rogers LJ, Todorova VK, Su LJ, Makhoul I. Genome-wide DNA methylation signatures to predict pathologic complete response from combined neoadjuvant chemotherapy with bevacizumab in breast cancer. PLoS One. 2020;15(4):e0230248. doi:10.1371/journal.pone.0230248.
- Hernandez Puente CV, Hsu P-C, Rogers LJ, Jousheghany F, et al. Association of DNA-Methylation Profiles With Immune Responses Elicited in Breast Cancer Patients Immunized With a Carbohydrate-Mimicking Peptide: A Pilot Study. Front Oncol. 2020;10:879. doi:10.3389/fonc2020.
- Kalekar LA, Schmiel SE, Nandiwada SL, Lam WY, Barsness LO, Zhang N, Stritesky GL, Malhotra D, Pauken KE, Linehan JL. CD4+ T cell anergy prevents autoimmunity and generates regulatory T cell precursors. Nat Immunol. 2016;17(3):304–314. doi:10.1038/ni.3331.
- Kumar H, Lund R, Laiho A, et al. Gut microbiota as an epigenetic regulator: pilot study based on whole-genome methylation analysis. MBio. 2014; 5.Dec 16;5(6):e02113-14. doi: 10.1128/mBio.02113-14.
- Guan Z, Raut JR, Weigl K, Schottker B, Holleczek B, Zhang Y, Brenner H. Individual and joint performance of DNA methylation profiles, genetic risk score and environmental risk scores for predicting breast cancer risk. Mol Oncol. 2020;14:42–53. doi:10.1002/1878-0261.12594.
- Song X, Sun X, Oh SF, Wu M, Zhang Y, Zheng W, Geva-Zatorsky N, Jupp R, Mathis D, Benoist C. Microbial bile acid metabolites modulate gut RORγ+ regulatory T cell homeostasis. Nature. 2020;577(7790):410–415. doi:10.1038/s41586-019-1865-0.
- Hang S, Paik D, Yao L, Kim E, Trinath J, Lu J, Ha S, Nelson BN, Kelly SP, Wu L, et al. Bile acid metabolites control TH17 and Treg cell differentiation. Nature. 2019;576(7785):143–148. doi:10.1038/s41586-019-1785-z.