
ABSTRACT
Introduction: The retentive niches of deep caries lesions have a distinct biome. Methods: We evaluated the site-specific (occlusal and proximal) Candida-biome of Severe-Early Childhood Caries (S-ECC) in 66- children (132 lesions). Asymptomatic primary molars fitting the definition of the International Caries Detection and Assessment-(ICDAS)-caries-code 5/6 were analyzed. Deep-dentinal sampling and simultaneous assessment of pH were performed. Clinical isolates were speciated using multiplex-PCR and evaluated for their acidogenic and aciduric potential.Results: Surprisingly, a high prevalence of Candida species (72.7%), either singly or in combination, was noted from both the proximal and occlusal cavities. C. tropicalis was the most prevalent species (47%; 34/72), followed by C. krusei (43.1%; 31/72) and C. albicans (40.3%; 29/72), with C. glabrata being the least (9.7%; 7/72). Over 45% low-pH niches (pH <7) of both sites yielded either dual or triple species of Candida. Genotyping revealed three distinct C. albicans genotypes (A, B, and C) with (14/29; 48.3%) of strains belonging to Genotype A. All four evaluated Candida species exhibited acidogenic and aciduric potential, C. tropicalis being the most potent.Conclusion: This, the first report of the high-density, multispecies, yeast colonization of deep-dentinal lesions in S-ECC, suggests that the Candida-biome plays a significant etiologic role in the condition, possibly due to their profound acidogenicity in milieus rich in dietary carbohydrates.
Introduction
Early Childhood Caries (ECC) is perhaps the most globally prevalent, insidious form of dental caries affecting virtually millions of primary teeth of preschool children [Citation1,Citation2]. A hypervirulent variant of this intractable infection is called severe-early childhood caries (S-ECC). If untreated, the disease progresses into extensive cavitation, resulting in painful pulp involvement, loss of teeth, and possibly, systemic infection [Citation3,Citation4]. The major etiological agent of ECC is the plaque biofilm microbiome comprising a bacteriome and a mycobiome [Citation5].
Factors such as the diet and the individual’s oral hygiene critically impact the composition and functionality of this poly-microbial biofilm community with diverse inter-kingdom interactions [Citation6,Citation7]. Dietary carbohydrates, in particular, play a key role in promoting the recruitment and colonization of the biofilm matrix - including pathogens such as the mutans-streptococci, which are both acidogenic and aciduric. Subsequent acidification of the matrix-microenvironment by the cariogens leads to initial enamel demineralization and the eventual lesion [Citation8–10].
It is also likely that the specific anatomical location in which these cariogens reside mainly the occlusal and proximal tooth surfaces may modify the disease process, and some have suggested differential initiation and progression rates of caries in occlusal and proximal lesions of ECC. For instance, Vanderas et al. (2006), in a 4-year longitudinal study, observed that the caries progression was more rapid on the proximal surfaces of teeth compared to the occlusal surfaces [Citation11]. Allison et al. (2003) found a strong correlation between enclosed, proximal spaces in the primary dentition and caries incidence [Citation12]. Others have suggested that the rapid progression of proximal caries lesions, in comparison to the occlusal lesions, may be due to the increased rate of plaque accumulation due to difficulty in access for oral hygiene measures, sequestration from the neutralizing effect of salivary flow, compounded by the highly aciduric/acidophilic resident microbes, thriving at a low pH location [Citation13–16].
Streptococcus mutans and lactobacilli have traditionally been cited as the prime drivers of ECC [Citation17,Citation18], although reports from recent clinical studies have indicated the intriguingly high prevalence of Candida species in plaque biofilm in ECC [Citation19–22]. Indeed, several researchers studying the cariogenic process have surmised that the Candida species cohabiting with S. mutans may play a significant causal role in the initiation and progression of dental caries [Citation1,Citation2,Citation9]. This is evidenced by the fact that Candida spp, in general, have a remarkable ability to adhere and colonize dental enamel and exposed dentine surfaces while producing copious quantities of carboxylic acids through dietary carbohydrate metabolism [Citation23–25]. Moreover, the intense aciduric and acidogenic attributes of Candida spp. ensure their continued survival and growth in a low pH, dysbiotic ecosystem, hostile for healthy, biofilm propagation [Citation25–27].
Based on this background, and in the absence of any data in the literature on the yeast carriage of proximal and occlusal lesions of S-ECC, we hypothesized that the prevalence of yeasts in the latter two niches differ in terms of their Candida-biome. Hence, in a cohort of Emirati children with S-ECC, we evaluated the site-specific prevalence of candidal load in infected dentine harvested from primary molars. As acidogenicity is a surrogate indicator of the cariogenic potential of a pathogen, and the opportunity was also taken to evaluate in vitro the relative acidogenicity of a select group of 35, C. albicans (10 isolates), C. tropicalis (10), C. krusei (10) and C. glabrata (5) isolates from S-ECC lesions.
Material and Methods
Study subjects
A cohort of 66 Emirati children, aged 48 months to 72-months-old, attending the routine pedodontics teaching clinic at the University Dental Hospital Sharjah, UAE, were invited to participate in the study. After obtaining informed consent from the parent of each child participant, under a protocol approved by the Research Ethics Committee (REC), University of Sharjah (REC-18-02-18-03), all healthy, cooperative participants underwent a full dental examination. Exclusion criteria included i) the likelihood of pulp exposure during the carie excavation process, ii) receipt of antibiotics 4 weeks prior to the study, iii) those wearing orthodontic appliance/s/, iv) congenital tooth anomaly, (v) dentine samples from endodontically treated teeth, or bleeding from the cavity during the sample collection process.
Caries diagnosis, sample collection, microbiological and molecular biological analyses were performed as described below, with minor variations to the protocol from a recently published study [Citation28].
Caries diagnosis
Caries status was documented using the World Health Organization (WHO) criterion of decayed, missing, and filled (dmft) index via an oral examination. The severity of occlusal and proximal caries lesions was assessed and documented according to the International Caries Detection and Assessment System (ICDAS)-classification criteria codes [Citation29]. A trained paediatric dentist (KSF) conducted all the clinical investigations. Children with more than five decayed teeth and at least two asymptomatic primary molars with occlusal or proximal cavitated carious lesions involved were selected. The severity of cavitated lesions was classified according to the ICDAS code 5/6 (code 5-distinct cavity with visible dentine; code 6-extensive and distinct cavity with visible dentine affecting more than half of the surface) [Citation29].
Sample collection
Each selected symptom-free, caries active, deep-dentin, occlusal, and proximal lesions were isolated with cotton wool rolls to obviate salivary contamination. The pH of both the occlusal and proximal cavities was evaluated using pH indicator strip (Spezialindikator, pH 4.0–7.0, Merck, Germany) by a single operator (KSF) according to the protocol described by Carlen et al. [Citation30].
After cleaning and drying the occlusal and proximal cavities with a prophy brush without using a prophy paste supplement, the infected-dentine samples were collected by excavating soft dentine from the cavitated dentine lesions. Each sample was split into two aliquots, and one aliquot was put in a 1.5 ml microcentrifuge tube containing 300 µl of Phosphate Buffered Saline (PBS) for multiplex PCR. The second aliquot was dispersed in Brain Heart Infusion Broth (BHI) (Thermo Scientific Remel, USA) and was immediately frozen at −20°C prior to evaluation by microbial culture.
Microbiological analysis
In the laboratory setting, an aliquot in BHI broth was cultured aerobically on chloramphenicol supplemented (50 mg/mL) Sabourauds dextrose agar (SDA) at 37°C for 48 h, and the resultant growth was observed. Five colonies of each sample that yielded yeast growth on the SDA plate were then sub-cultured on CHROMagar (HiCrome™ Candida Differential Agar, M1297A) for 24 h. Pure cultures of different species were then obtained by selecting colony forming units (CFUs) based on their colony appearance on CHROM agar. The different candidal species obtained from each sample were then sub-cultured in Sabouraud dextrose broth containing 100 mM glucose for 24 hours to evaluate their acidogenic potential as well.
DNA isolation and multiplex PCR
DNA extraction of the infected-dentine samples was performed using MasterPure™ Complete DNA and RNA Purification (Epicenter, USA) following the manufacturer’s instructions. The extracted DNA quality and quantity was assessed using the Colibri Microvolume Spectrometer (Titertek-Berthold Detection Systems GmbH, Germany). DNA samples were considered pure if the A260/280 ratio were more than 1.8 and the A260/230 values were in the range of 1 to 2.2.
Clinical isolates were identified and confirmed by the multiplex PCR amplification, which allowed the identification of six common, clinically pathogenic yeasts of the Candida genus, namely C. albicans, C. glabrata, C. tropicalis, C. krusei, C. parapsilosis, and C. dubliniensis. The working method was established on the amplification of the two fragments from the ITS1 and ITS2 regions by the combination of two yeast specific and six species-specific primers [Citation31] using PCR ().
Table 1. Amplicon sizes (base pairs) results from multiplex PCR amplification using yeast specific (Universal-UNI1 and UNI2) and corresponding species-specific primers of Candida spp
PCR involved the following thermal cycling conditions: 40 cycles of 15 s at 94°C, then 30 s at 55°C, and 45 s at 65°C, following an initial 10-min period of DNA denaturation and enzyme activation at 94°C [Citation32]. All amplicons were assessed by electrophoresis in 2.0% (w/v) agarose gels run at 90 V/cm2 for 60 mins. Samples containing multiple species were further re-confirmed by quantitative PCR analysis using species-specific primers for the identified phenotypes. This was particularly necessary for the case as discrimination of C. tropicalis and C. albicans within multispecies samples was difficult using gel electrophoresis due to the relatively close proximity of bands derived from multiplex PCR amplification.
C. albicans genotyping
Candida albicans sequences were characterized into five genotypes A, B, C, D, and E based on the band patterns, with genotype A-450 bp, genotype B-840 bp, genotype C-450-840 bp, genotype D-1080 bp, and genotype E-1400bp. C. albicans genotyping was performed using the following primers: CA-INT- L (5ʹ-ATAAGGGAAGTCGG-CAAAATAGATCCGTAA-3ʹ) and CA-INT-R (5ʹCCTTGGCTGTGGTTTCGCTAGATAGTAGAT-3ʹ) (). The amplification process follows an initial denaturation at 93°C for 5 mins, then 40 cycles of denaturation for 30 s at 93°C, with primer annealing at 55°C for 45 s and extension at for 45 s at 72°C, with a final extension for 7 mins at 72°C [Citation33]. The PCR products were loaded into a 2% (w/v) agarose gel (Bio-Rad, Hercules, CA, USA) and electrophoresed at 100 V/cm2 for 30 mins and stained using ethidium bromide solution.
Table 2. Primers used for the determination of Candida albicans genotypes
Acid production and Acid tolerance
Investigations on acid production and acid tolerance involved 35 randomly selected isolates, comprising 10 strains each of C. albicans, C. tropicalis, C. krusei, and five strains of C. glabrata. A protocol previously described by Qiu et al., with some modifications was used for this purpose [Citation20].
Candida was suspended in PBS and adjusted to an optical density (OD) of 1.0 at 530 nm (1×108 cells/mL) for both assays. Sabouraud dextrose broth (SDB) containing 100 mM glucose was adjusted to pH 4.0, 5.0, 5.5, 6.0, and 7.0 using sterile HCl and NaOH. Then, 50 µl of suspension of the selected isolate was inoculated into 5 mL SDB and cultured at 37°C for 48-h. After incubation, the growth was centrifuged at 5000 rpm for 5 min at 4°C, and the pH of the supernatant was measured using a pH meter (JK-PHM-002 Desktop pH meter). Acid production was assessed as change of pH over the incubation period.
To determine acid-tolerance of Candida spp., under different pH values, the growth of different species was evaluated by washing the precipitate (above) three times with PBS and then resuspending in 2 mL PBS. Finally, the turbidity was measured at 530 nm (OD530) using a spectrophotometer (Genova Bio Spectrophotometer).
Higher ∆pH and OD530 values were taken as a surrogate indicator of acidogenicity and survival potency in an acidic milieu, respectively. All tests were done in triplicate on two separate occasions.
Acidogenicity in glucose supplemented media
The relative acidogenic potential of the 35 clinical isolates of Candida species in dietary sucrose (as opposed to glucose) was also evaluated. For this purpose, 100 mM sucrose supplemented SDB in multiwell plates was used, and pH reduction assessed at 24 h and 48 h time intervals as per the method of Samaranayake et al. (1983) with some modifications [Citation26]. Separate overnight growth of yeast suspension from each of the four species was adjusted to an optical density (OD) of 1.0 at 530 nm (1×108 cells/ml), and 50 µl of each of the selected isolates were added to each well of 5 ml SDB containing 100 mM sucrose (adjusted to pH 7.0 using NaOH) followed by incubation for 48 h at 37°C. At the above time points, the pH was assessed using a pH meter (Portable pH meter-H1991, Hanna, USA).
Statistical analyses
Results were presented as mean ± SD, and data were evaluated using t-tests. Chi-square test, Fischer exact tests, and analysis of variance (ANOVA) for comparing group differences between Candida species were used. Statistical significance was considered significant at p ≤ 0.05.
Results
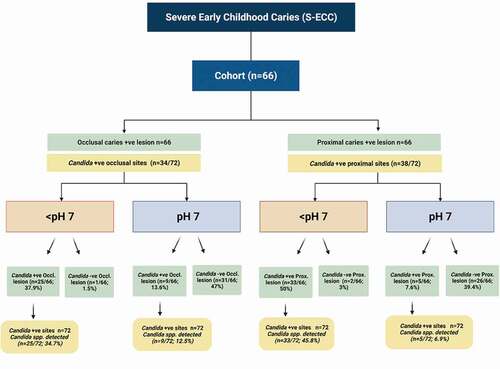
Candida species were detected in occlusal and proximal deep caries lesions of 48/66 (72.7%) children (mean-age 5.3 ± 0.78 years) with S-ECC. The mean decayed teeth per child was 8.32 ± 2.66, while the subset of children who were yeast positive had 8.44 ± 2.77 mean-decayed teeth. In total 72/132 (54.5%) proximal and occlusal sites harboured yeasts either as single or multiple species (). Among children with Candida positive caries lesions, 25/72 (34.7%), and 9/72 (12.5%) of the occlusal lesions at pH <7 and pH 7, respectively, harboured single or dual/multispecies Candida.
Table 3. Frequency distribution of prevalence of different Candida species in terms of their occurrences as single, dual and triple species in total 72 Candida positive sites (34 occlusal and 38 proximal)
Of children with deep caries lesions (66-occlusal and 66-proximal lesions; total 132), 26/66 (39.4%) of occlusal lesions and 35/66 (53%) proximal lesions were acidic, i.e. pH <7 (; ). Similarly, 33 (45.8%) and 5 (6.9%) of the proximal lesions, each at pH <7 and pH 7, respectively, yielded single or dual/multispecies Candida. Thus, the total isolation frequency of yeasts from both the proximal and occlusal lesions was 72/132 (54.5%) ().
Figure 1. Prevalence of Candida in proximal and occlusal S-ECC categorized as per the pH of the lesions
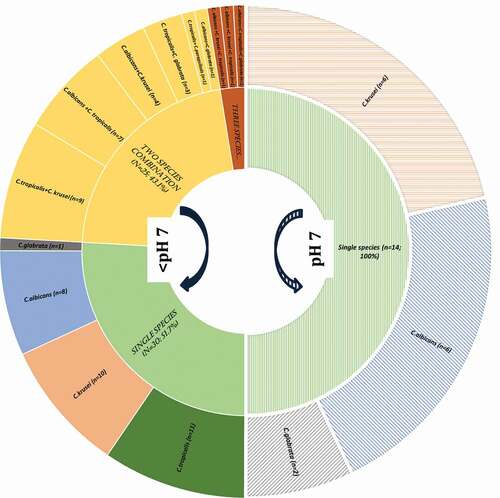
All Candida isolates from both the proximal and occlusal lesions belonged to four species, C. krusei, C. albicans, C. tropicalis, and C. glabrata (). The highly prevalent species were C. tropicalis, C. krusei, and C. albicans isolated from 34/72 (47%), 31/72 (43.1%), and 29/72 (40.3%) of the clinical samples, respectively (p > 0.05); C. glabrata species was the least prevalent, being present only in 7/72 (9.7%) samples ().
Figure 2. Candida species habitation at <pH 7 (acidic) and pH7 of deep-dentine caries lesions -ICDAS caries code 5 and 6
A predominance of C. albicans in proximal lesions in 19/29 (65.5%) samples was evident, either singly or in cohabitation with another Candida species but mostly with C. tropicalis (; p < 0.001). Interestingly, C. krusei, when isolated, was found on significantly more occasions in occlusal lesions compared with proximal lesions (22/72, 30.6%; p < 0.001) either as a single species or in combination with C. tropicalis, p < 0.05) (). There was no significant difference in the isolation frequency of yeasts between occlusal and proximal lesions ().
Significant variations in candidal species colonization profiles could be discerned depending on the alkalinity or the acidity of deep-dentinal lesions. In general, the low pH niches (<pH 7) tended to harbour multispecies of yeasts, while mono-species Candida was mainly seen in high pH localities (pH 7). In comparison to 33/72 (45.8%) occasions of multispecies of Candida in the acidic milieu of proximal lesions, and none of the alkaline lesions yielded multispecies of Candida. Only 5/72 (6.9%) yielded single species of Candida at an alkaline pH (p = 0.001; ; ).
C. tropicalis, C. albicans, and C. krusei demonstrated a high propensity for colonization of acidic pH locales of both the occlusal and proximal caries lesions (; ; p < 0.001). Curiously, C tropicalis was exceptional in that they were noted only in acidic pH lesions ().
Interestingly, this study provided some indications of candidal interspecies interactions in occlusal and proximal caries lesions. Only 9/72 (12.5%) occlusal lesions at pH 7 were yeast positive, compared to 25/72 (34.7%) of occlusal lesions with an acidic niche (pH < 7; p = 0.001). Moreover, low pH environments of deep, proximal lesions exhibited either dual or triple species communities of Candida. Notably, we found C. albicans cohabited almost exclusively with C. tropicalis in relatively more acidic, proximal cavities ().
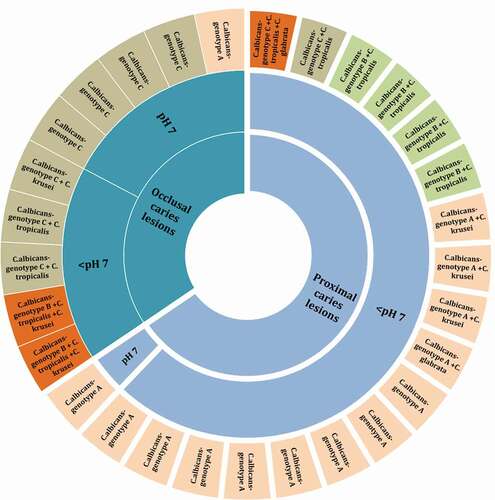
All C. albicans isolated fell into either A, B or C, genotypes, while the genotypes D and E were not found among our isolates. Genotype A was the most predominant (14/29; 48.3%) followed by the genotypes C (9/29; 31.0%), and B (6/29; 20.7%). We noted a high prevalence of strains belonging to genotype A in the proximal isolates, whereas the genotypes B and C were predominantly found in occlusal cavities (). In terms of dual-species interactions, Genotype A and B were predominantly present in proximal caries lesions with an acidic pH and cohabited almost exclusively with C. krusei and C. tropicalis, respectively.
Figure 3. Distribution of C. albicans (n = 29) genotypes (A, B and C) as single, dual, and mixed species at neutral pH 7 and acidic <pH 7 in occlusal and proximal -dentine caries lesions
All four isolated Candida species exhibited a high degree of acidogenic potential in metabolizing the dietary carbohydrate glucose, depressing the pH of the growth milieu from pH 7 to pH 4 (). This was the case when the experiment was repeated with varying starter pH values of the suspensions. Among all four Candida spp, C. tropicalis demonstrated the highest potency for acid production, followed by C. albicans. Significant interspecies differences in the acidogenic potential were noted between C. tropicalis and C. glabrata (p ≤ 0.05). Additionally, all yeast isolates exhibited a relatively high acid tolerance and continued to grow at a low pH 4 when pulsed with 100 mM of glucose (). C. tropicalis was noticeably more aciduric than C. glabrata despite varying the initial pH levels (p ≤ 0.05).
Table 4. The in vitro acidogenic potential of clinical isolates of Candida species and genotypes of C. albicans from ECC lesions at 24 hours- post exposure to 100 mM glucose (mean ± SD)
Table 5. Aciduric potential of clinical isolates of Candida species and genotypes of C. albicans at 24 hours- post consumption 100 mM of glucose (mean ± SD)
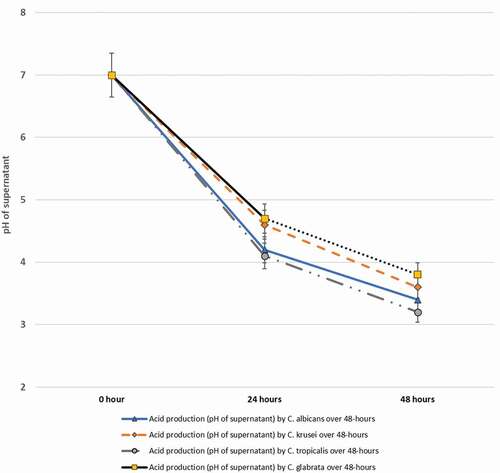
In terms of acidogenicity of the four yeast species in dietary sucrose (100 mM) supplemented media, all isolates uniformly reduced the pH from 7.0 to 3.2–3.8 (). C. glabrata, once again, appeared to exhibit the highest acidogenic potential, although no significant inter-species differences were noted among the four species ().
Figure 4. In vitro acid production by a total of 35 isolates belonging to C. albicans (10 isolates), C. krusei (10) C. tropicalis (10) and C. glabrata (5) isolated from S-ECC samples, over a 48-hour period in 100 mM sucrose supplemented media
When the acidogenicity of the different genotypes were assessed, all three genotypes A, B, and C of C. albicans displayed varying degrees of acidogenicity and aciduricity and grew uniformly well in a milieu rich in glucose. However, C. albicans genotypes B and C of were significantly more acidogenic than genotype A, particularly at pH 4, after pairwise comparisons (p < 0.05; ). No significant difference in ∆pH was observed between genotypes B and C.
Discussion
Early Childhood Caries (ECC), and in particular its hypervirulent variant, severe-early childhood caries (S-ECC) affects virtually millions of primary teeth of preschool children globally [Citation1,Citation2]. Although the classic microbial pathogens associated with caries are mutans-streptococci and lactobacilli [Citation34], there are an increasing number of studies that indicate the intimate association of Candida species, an opportunistic oral commensal, with caries etiology [Citation35]. As there were no published reports, to our knowledge, on the prevalence of Candida species in S-ECC, we embarked on the current study to obtain baseline data on the presence of oral yeasts in S-ECC in a cohort of Emirati children with a very high prevalence of the disease [Citation28]. An experienced pedodontist performed site-specific mycobiological sampling of a total of 132 (occlusal and proximal) lesions in 66 children. Subsequent species-specific molecular analyses were performed on purified isolates to elicit their genotypes, followed by phenotypic analyses to evaluate in vitro aciduricity and acidogenicity.
There were three major observations in this study. The first was the surprisingly high oral yeast isolation rate from S-ECC lesions, with almost three-quarters of the lesions (72.7%) harbouring Candida species. Second, the predominance of non-albicans Candida species (NACS) in the lesions, and finally, the multispecies communities of Candida species, mainly in acidic caries niches.
In general, oral yeast carriage rates in healthy populations are reported to range from 2.0 to 71.0%, although these figures can vary considerably depending on factors such as the age, the wearing of oral appliances, and the nutritional status of individuals, including a high carbohydrate diet [Citation36]. In one of the most comprehensive 4-year longitudinal study to date on the oral carriage of yeast in 116 Chinese primary schoolchildren in Hong Kong, Sedgley et al. (1997) noted the oral prevalence of yeasts for each consecutive year to be 7.7%, 12.0%, 14.4%, and 15.5%, respectively, together with a weighted mean of 12.5%. Although the caries status of the children was not recorded, the vast majority (84%) of the isolates were C. albicans as opposed to the current findings [Citation37]. It would appear that in comparison the yeast prevalence in S-ECC lesions, and by extension, the oral yeast carriage in over one-half of our cohort was remarkably high. As discussed below, such high yeast prevalence may be related to the multiple, stagnant deep caries lesions with acidic pH milieus.
Although C. albicans is thought to be a key driver of the caries process aided and abetted by other cariogens [Citation35], a predominance of NACS (particularly C. krusei and C. tropicalis), were noted in the caries samples from our cohort. Candida krusei in particular, was the predominant species in occlusal samples, isolated either singly or in combination with other Candida species. In a comprehensive review of 44 publications for which statistics on rates of human carriage of C. krusei are available, Samaranayake and Samaranayake noted that the highest oral carriage rates for C. krusei were 6.1% either in health or in disease [Citation38] a figure significantly lower than 43.1% noted here.
The predominant oral prevalence of C. krusei has, to our knowledge, been reported only on a single previous occasion in a group of recluse leprosy patients in Chiang Mai, Thailand [Citation39]. The authors of this latter paper could not attribute a specific reason for this observation, although sharing food and utensils in the secluded community was suspected. Such reasons, however, would not explain the unusually high prevalence of C. krusei in our cohort, although it is tempting to speculate that, in general, poor oral hygiene in relatively inaccessible deep, particularly in proximal, caries lesions, as well as high sucrose dietary regimens, may have contributed to this phenomenon.
Compared to other medically significant Candida species, C. krusei has been isolated from a wide range of natural habitats. These include the atmosphere, fruits, sewage, silage, soil, wine, and beer [Citation40]. Recently, C. krusei has been identified as an important agent of nosocomial candidiasis owing to its inherent resistance to azoles, especially fluconazole [Citation38]. Hence, it is considered to be a facultative saprophyte which may cause opportunistic infections when host defences are impaired.
Candida tropicalis, together with C. krusei and C. albicans, was the second most predominant species, particularly in the high acidic caries milieus (below pH 7), of both occlusal and proximal-cavitated lesions. It is known that C. albicans and C. tropicalis, are genetically similar, and group together in phenotypic assays relative to other pathogenic Candida species [Citation41]. In addition, both of these species appear to have the capacity for cell-wall remodelling for survival, in the face of adverse ecological pH [Citation42], in addition, to possessing twin attributes of surface adherence and robust biofilm-producing potential essential for successful colonization [Citation43–47]. These may plausibly explain why C. albicans and C. tropicalis species are effective pathogens in S-ECC in comparison to their counterparts. Further work, however, is required to confirm or refute our nascent findings and the reasons for the preponderance of specific yeast species in S-ECC.
The thriving and cohabiting commune habitats of Candida spp. in S-ECC were another unexpected finding of our study (). Multispecies communities of Candida in the oral cavity were first reported by Samaranayake et al. over three decades ago [Citation48]. These authors noted the phenomenon in 15.3% of 150 clinical oral samples, and the most common cohabitants were reported to be C. albicans and C. glabrata. Others have subsequently confirmed these findings in health and disease, with various species combinations, in different body regions, including the oral cavity [Citation49,Citation50].
With regard to multispecies yeast carriage in the current study, there were two striking findings. First, none of the dentinal lesions with alkaline or neutral pH had multispecies carriage, and the phenomenon was exclusively seen in cavities with an acidic pH (). Second, proximal cavities had a higher mixed-species fungal incidence compared to occlusal-deep dentine niches. These observations tend to concur with the fact that low-pH habitats promote yeast growth, and the proximal lesions with low accessibility for oral hygiene with possible entrapped cariogenic, nutritional food sources [Citation9] combined with a stagnant salivary flow devoid of its flushing action, may have all fostered the aciduric communities of yeasts [Citation16,Citation51].
Filamentation being a foremost contributor to their biofilm formation and virulence [Citation52]. In a recent study, Pathirana and colleagues (2019) elegantly demonstrated that, in dual-species co-cultures of C. albicans and C. tropicalis, biofilm formation was associated with intense filamentation and suggested that this promoted a mutualistic survival advantage for the yeasts [Citation53]. Others, however, have suggested antagonistic behaviour between these species in co-culture and observed that C. tropicalis negatively impacted on growth and virulence of C. albicans [Citation52]. The precise role of their interspecies interplay in the caries pathogenesis needs further analysis.
As previously observed [Citation20,Citation33,Citation54], we also noted that the commonest oral C. albicans genotypes in S-ECC lesions were A, B, and C. This preliminary observation needs to be confirmed due to the small sample size of our study.
The cariogenic process is intimately intertwined with acid production and acid-stress tolerance of the cariogens [Citation26,Citation27,Citation55]. Aciduricity is a complex trait that involves many contributing factors regulating adaptive acid resistance [Citation56–60]. Our in vitro growth data confirm previous reports where, in glucose and sucrose containing environments, all tested Candida species produce robust growth and were acid-tolerant to pH 3.00, with few interspecies variations [Citation26,Citation27,Citation61]. Interestingly, C. tropicalis was the most acidogenic of the tested species, both in glucose and sucrose supplemented media, closely followed by C. albicans. Considering that the enamel demineralization process that initiates the carious process begins at pH 5.5 [Citation62,Citation63], it is entirely plausible that yeasts in these lesions contribute to S-ECC. However, for better insight of yeast survival potential in a low pH environment as well as their total contribution to the caries process, it would be useful to evaluate the colony forming units (CFUs) from the primary isolation plates in future studies. Alternately, quantitative PCR (qPCR) or more accurate, next-generation sequencing data of the mycological burden would be of added value.
Acidogenicity and aciduricity of C. albicans are well-known hallmark traits of these opportunists [Citation26,Citation27]. Our data indicated that all three genotypes A, B, and C, were acidogenic and were well adapted to grow in a low acidic milieu. Interestingly the acidogenicity of genotypes B and C differed significantly from that of genotype A. An identical observation was made by Qiu et al. [Citation20], who studied the genotypic diversity and carcinogenicity of C. albicans isolated from Chinese children with and without ECC (but not S-ECC). This previous investigation also detected genotypes B and C more frequently in the ECC group than in the caries-free group. Yet, whether genotypic diversity is related to the virulence of C. albicans is unclear, as there are contrasting reports. Karahan et al. [Citation64] showed that in patients with deep-seated infections, genotype A was more invasive, compared with genotypes B and C. In contrast, another study found that genotype C was the most invasive and genotype A the least invasive in a selection of clinical isolates [Citation65]. Others could not discern any difference in invasiveness of the genotypes in patients with bloodborne candidal infections and concluded that the genotypic distribution of C. albicans was unrelated to invasiveness [Citation66].
Finally, the presence of a significant oral reservoir of multiple Candida species in this cohort of children is of concern, as it may serve as a nexus for systemic infections, especially in compromised individuals who may be at risk [Citation4,Citation18]. There have been a number of reports where oral fungal pathogens originating from an oral niche have been implicated in systemic infections ranging from infective endocarditis to various fungaemia [Citation67–70]. Thus, the high prevalence of fluconazole-resistant C. krusei species in our cohort is particularly noteworthy in this context.
To the best of our knowledge, this is the first comprehensive report highlighting the Candida mycobiome in the infected dentine-niche of occlusal and proximal cavities of S-ECC. Furthermore, we have unequivocally demonstrated the presence of multiple Candida species in deep dentinal caries lesions of S-ECC. However, further work is needed to assess the contribution of Candida species in the initiation and progression (S-ECC) as well as their impact, particularly on the therapeutic management of the condition.
Author Contributions
KSF together with HE and LPS performed data collation analysis and manuscript writing; HCN and SP critically examined and edited the final versions of the manuscript. All authors gave approval for the final version to be published.
Acknowledgments
Dr. Fakhruddin gratefully acknowledges the visiting researcher position offered by the Tohoku University Graduate School of Dentistry, and the advice and guidance of Professor Egusa. Professor Samaranayake gratefully acknowledges the Thammasat University of Thailand for the award of a Bualuang ASEAN Chair Professorship to support this research.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Correction Statement
This article has been republished with minor changes. These changes do not impact the academic content of the article.
References
- Hajishengallis E, Parsaei Y, Klein MI, et al. Advances in the microbial etiology and pathogenesis of early childhood caries. Mol Oral Microbiol. 2017;32(1):24–12. Epub 2015/ 12/31. PubMed PMID: 26714612; PubMed Central PMCID: PMCPMC4929038.
- Falsetta ML, Klein MI, Colonne PM, et al. Symbiotic relationship between Streptococcus mutans and Candida albicans synergizes virulence of plaque biofilms in vivo. Infect Immun. 2014;82(5):1968–1981. PubMed PMID: 24566629
- Casamassimo PS, Thikkurissy S, Edelstein BL, et al. Beyond the dmft: the human and economic cost of early childhood caries. J Am Dent Assoc. 2009;140(6):650–657. Epub 2009/ 06/06. PubMed PMID: 19491160.
- Hossain H, Ansari F, Schulz-Weidner N, et al. Clonal identity of Candida albicans in the oral cavity and the gastrointestinal tract of pre-school children. Oral Microbiol Immunol. 2003;18(5):302–308.
- Baker JL, Bor B, Agnello M, et al. Ecology of the Oral Microbiome: beyond Bacteria. Trends Microbiol. 2017;25(5):362–374. Epub 2017/ 01/17. PubMed PMID: 28089325; PubMed Central PMCID: PMCPMC5687246.
- Gomez A, Nelson KE. The Oral Microbiome of Children: development, Disease, and Implications Beyond Oral Health. Microb Ecol. 2017;73(2):492–503. Epub 2016/ 09/14. PubMed PMID: 27628595.
- Jenkinson HF, Lamont RJ. Oral microbial communities in sickness and in health. Trends Microbiol. 2005;13(12):589–595.
- Bowen WH. Dental caries – not just holes in teeth! A perspective. Mol Oral Microbiol. 2016;31(3):228–233.
- Bowen WH, Burne RA, Wu H, et al. Oral Biofilms: pathogens, Matrix, and Polymicrobial Interactions in Microenvironments. Trends Microbiol. 2018;26(3):229–242. Epub 2017/ 11/04. PubMed PMID: 29097091; PubMed Central PMCID: PMCPMC5834367.
- Lamont RJ, Koo H, Hajishengallis G. The oral microbiota: dynamic communities and host interactions. Nature Rev Microbiol. 2018;16(12):745–759.
- Vanderas AP, Gizani S, Papagiannoulis L. Progression of proximal caries in children with different caries indices: a 4-year radiographic study. Eur Arch Paediatr Dent. 2006;7(3):148–152. Epub 2006/ 12/05. PubMed PMID: 17140544.
- Allison PJ, Schwartz S. Interproximal contact points and proximal caries in posterior primary teeth. Pediatr Dent. 2003;25(4):334–340. Epub 2003/ 09/19. PubMed PMID: 13678098.
- Dirksen TR, Little MF, Bibby BG. The pH of carious cavities-II. The pH at different depths in isolated cavities. Arch Oral Biol. 1963 PubMed PMID: 14028031;8:91–97. Epub 1963/ 03/01.
- Ekstrand KR, Bruun G, Bruun M. Plaque and gingival status as indicators for caries progression on approximal surfaces. PubMed PMID: 9438570. 1998. 32(1):41–45. Caries Res. Epub 1998/ 01/23.
- Novaes TF, Matos R, Braga MM, et al. Performance of a pen-type laser fluorescence device and conventional methods in detecting approximal caries lesions in primary teeth–in vivo study. Caries Res. 2009;43(1):36–42. Epub 2009/ 01/13. PubMed PMID: 19136830.
- Marsh PD, Martin MV. Oral Microbiology Textbook ed Lewis MA. Edinburgh, London, NewYork, Oxford: Churchill Livingstone Elsevier;2009 8–23. 2019.
- Mattos-Graner RO, Klein MI, Smith DJ. Lessons Learned from Clinical Studies: roles of Mutans Streptococci in the Pathogenesis of Dental Caries. Curr Oral Health Rep. 2014;1(1):70–78.
- Caufield PW, Schön CN, Saraithong P, et al. Oral Lactobacilli and Dental Caries: a Model for Niche Adaptation in Humans. J Dent Res. 2015;94(9Suppl):110S–8S. PubMed PMID: 25758458
- de Carvalho FG, Silva DS, Hebling J, et al. Presence of mutans streptococci and Candida spp. in dental plaque/dentine of carious teeth and early childhood caries. Arch Oral Biol. 2006;51(11):1024–1028. Epub 2006/ 08/08. PubMed PMID: 16890907.
- Qiu R, Li W, Lin Y, et al. Genotypic diversity and cariogenicity of Candida albicans from children with early childhood caries and caries-free children. BMC Oral Health. 2015;15(1):144.
- Yang XQ, Zhang Q, Lu LY, et al. Genotypic distribution of Candida albicans in dental biofilm of Chinese children associated with severe early childhood caries. Arch Oral Biol. 2012;57(8):1048–1053.
- Raja M, Hannan A, Ali K. Association of oral candidal carriage with dental caries in children. Caries Res. 2010;44(3):272–276. Epub 2010/ 06/03. PubMed PMID: 20516688.
- DdSAV B, Vicente VA, Fraiz FC, et al. Analysis of the in vitro adherence of Streptococcus mutans and Candida albicans. Braz J Microbiol. 2007;38:624–631.
- Ene IV, Heilmann CJ, Sorgo AG, et al. Carbon source-induced reprogramming of the cell wall proteome and secretome modulates the adherence and drug resistance of the fungal pathogen Candida albicans. Proteomics. 2012;12(21):3164–3179. PubMed PMID: 22997008; PubMed Central PMCID: PMCPMC3569869. Epub 2012/ 09/22.
- Klinke T, Guggenheim B, Klimm W, et al. Dental caries in rats associated with Candida albicans. Caries Res. 2011;45(2):100–106. Epub 2011/ 03/18. PubMed PMID: 21412001.
- Samaranayake LP, Geddes DA, Weetman DA, et al. Growth and acid production of Candida albicans in carbohydrate supplemented media. Microbios. 1983;37(148):105–115. Epub 1983/ 01/01. PubMed PMID: 6353167.
- Samaranayake LP, Hughes A, Weetman DA, et al. Growth and acid production of Candida species in human saliva supplemented with glucose. J Oral Pathol. 1986;15(5):251–254. Epub 1986/ 05/01. PubMed PMID: 3091791.
- Fakhruddin KS, Perera Samaranayake L, Egusa H, et al. Candida biome of severe early childhood caries (S-ECC) and its cariogenic virulence traits. J Oral Microbiol. 2020;12(1):1724484.
- Ekstrand KR, Martignon S, Ricketts DJ, et al. Detection and activity assessment of primary coronal caries lesions: a methodologic study. Oper Dent. 2007;32(3):225–235. Epub 2007/ 06/09. PubMed PMID: 17555173.
- Carlén A, Hassan H, The LP. ‘Strip Method’: a Simple Method for Plaque pH Assessment. Caries Res. 2010;44(4):341–344.
- Trost A, Graf B, Eucker J, et al. Identification of clinically relevant yeasts by PCR/RFLP. J Microbiol Methods. 2004;56(2):201–211. Epub 2004/ 01/28 PubMed PMID: 14744449.
- Carvalho A, Costa-De-Oliveira S, Martins ML, et al. Multiplex PCR identification of eight clinically relevant Candida species. Med Mycol. 2007;45(7):619–627.
- Qiong Yang X, Zhang Q, Ying LL, et al. Genotypic distribution of Candida albicans in dental biofilm of Chinese children associated with severe early childhood caries 2012. 1048–1053 p.
- Essential SL. Microbiology for Dentistry. Vol. 2018. Lakshman Samaranayake. Essential Microbiology for Dentistry. 5th Edition ed. Elsevier; 2018. p. 1–400.
- Pereira D, Seneviratne CJ, Koga-Ito CY, et al. Is the oral fungal pathogen Candida albicans a carcinogen? Oral Dis. 2018;24(4):518–526. Epub 2017/ 05/18. PubMed PMID: 28513096.
- Samaranayake LP. Nutritional factors and oral candidosis. J Oral Pathol Med. 1986;15(2):61–65.
- Sedgley CM, Samaranayake LP, Chan JC, et al. A 4-year longitudinal study of the oral prevalence of enteric gram-negative rods and yeasts in Chinese children. Oral Microbiol Immunol. 1997;12(3):183–188. Epub 1997/ 06/01. PubMed PMID: 9467406.
- SAMARANAYAKE YH, SAMARANAYAKE LP. Candida krusei: biology, epidemiology, pathogenicity and clinical manifestations of an emerging pathogen. J Med Microbiol. 1994;41(5):295–310.
- Reichart PA, Samaranayake LP, Samaranayake YH, et al. High oral prevalence of Candida krusei in leprosy patients in northern Thailand. J Clin Microbiol. 2002;40(12):4479–4485. Epub 2002/ 11/28. PubMed PMID: 12454139; PubMed Central PMCID: PMCPMC154582.
- Yeasts D-SL. Distribution of yeasts in nature. In: AH R, JS H editor. The yeasts, biology of yeasts, Vol. 1st edn.:1. Academic Press, London. 1969;1(1, :79–105.
- Butler G, Rasmussen MD, Lin MF, et al. Evolution of pathogenicity and sexual reproduction in eight Candida genomes. Nature. 2009;459(7247):657–662. PubMed PMID: 19465905; PubMed Central PMCID: PMCPMC2834264. Epub 2009/ 05/26.
- Sherrington SL, Sorsby E, Mahtey N, et al. Adaptation of Candida albicans to environmental pH induces cell wall remodelling and enhances innate immune recognition. PLoS Pathog. 2017;13(5):e1006403. PubMed PMID: 28542528; PubMed Central PMCID: PMCPMC5456412. Epub 2017/ 05/26.
- da Costa KR, Ferreira JC, Lavrador MA, et al. Virulence attributes and genetic variability of oral Candida albicans and Candida tropicalis isolates. Mycoses. 2012;55(3):e97–e105. Epub 2011/ 11/01. PubMed PMID: 22035510.
- Chaves GM, Diniz MG, da Silva-Rocha WP, et al. Species distribution and virulence factors of Candida spp. isolated from the oral cavity of kidney transplant recipients in Brazil. Mycopathologia. 2013;175(3–4):255–263. PubMed PMID: 23539354. Epub 2013/ 03/30.
- Zuza-Alves DL, Silva-Rocha WP, Chaves GM. An Update on Candida tropicalis Based on Basic and Clinical Approaches. Front Microbiol. 2017;8:1927. PubMed PMID: 29081766
- Marcos-Zambrano LJ, Escribano P, Bouza E, et al. Production of biofilm by Candida and non-Candida spp. isolates causing fungemia: comparison of biomass production and metabolic activity and development of cut-off points. Int J Med Microbiol. 2014;304(8):1192–1198. Epub 2014/ 09/17. PubMed PMID: 25224357.
- Pannanusorn S, Fernandez V, Romling U. Prevalence of biofilm formation in clinical isolates of Candida species causing bloodstream infection. Mycoses. 2013;56(3):264–272. Epub 2012/ 11/02. PubMed PMID: 23113805.
- Samaranayake LP, MacFarlane TW, Williamson MI. Comparison of Sabouraud dextrose and Pagano-Levin agar media for detection and isolation of yeasts from oral samples. J Clin Microbiol. 1987;25(1):162–164. Epub 1987/ 01/01. PubMed PMID: 3539988; PubMed Central PMCID: PMCPMC265849.
- Samaranayake L. Commensal oral Candida in Asian cohorts. Int J Oral Sci. 2009;1(1):2–5. Epub 2009/ 03/01. PubMed PMID: 20690497; PubMed Central PMCID: PMCPMC3470111.
- Keten HS, Keten D, Ucer H, et al. Prevalence of oral Candida carriage and Candida species among cigarette and maras powder users. Int J Clin Exp Med. 2015;8(6):9847–9854. PubMed PMID: 26309667
- Bourgeois D, David A, Inquimbert C, et al. Quantification of carious pathogens in the interdental microbiota of young caries-free adults. PLOS ONE. 2017;12(10):e0185804.
- de Barros PP, Rossoni RD, Freire F, et al. Candida tropicalis affects the virulence profile of Candida albicans: an in vitro and in vivo study. Pathog Dis. 2018;76(2). Epub 2018/ 04/05. DOI:https://doi.org/10.1093/femspd/fty014. PubMed PMID: 29617858.
- Pathirana RU, McCall AD, Norris HL, et al. Candida Species Adhere to Candida albicans and Benefit From Dual Biofilm Growth. Front Microbiol. 2019;10:1188.
- Zhang W, Li Y, Lin J, et al. Cariogenicity of Candida albicans of distinct genotypes among 3-5-year-old Uygur children in Kashgar, China- a case-control study. BMC Oral Health. 2018;18(1):203.
- Klinke T, Kneist S, de Soet JJ, et al. Acid Production by Oral Strains of Candida albicans and Lactobacilli. Caries Res. 2009;43(2):83–91.
- Bowman BJ, Bowman EJ H+-ATPases from mitochondria, plasma membranes, and vacuoles of fungal cells. J Membr Bio l. 1986;94(2):83–97. Epub 1986/ 01/01. PubMed PMID: 2882028.
- Vylkova S, Carman AJ, Danhof HA, et al. The Fungal Pathogen Candida albicans Autoinduces Hyphal Morphogenesis by Raising Extracellular pH. mBio. 2011;2(3):e00055–11.
- Zhou J, Liu L, Improved CJ. ATP supply enhances acid tolerance of Candida glabrata during pyruvic acid production. J Appl Microbiol. 2011;110(1):44–53.
- Schmidt P, Walker J, Selway L, et al. Proteomic analysis of the pH response in the fungal pathogen Candida glabrata. Proteomics. 2008;8(3):534–544. PubMed PMID: 18186024. Epub 2008/ 01/11.
- Wei P, Li Z, He P, et al. Genome shuffling in the ethanologenic yeast Candida krusei to improve acetic acid tolerance. Biotechnol Appl Biochem. 2008;49(Pt2):113–120. Epub 2007/ 07/17. PubMed PMID: 17630953.
- Van Ende M, Wijnants S, Van Dijck P, et al. Signaling in Candida albicans and Candida glabrata. Front Microbiol. 2019;10:99.
- Takahashi N, Nyvad B. Ecological Hypothesis of Dentin and Root Caries. Caries Res. 2016;50(4):422–431.
- Abou Neel EA, Aljabo A, Strange A, et al. Demineralization-remineralization dynamics in teeth and bone. Int J Nanomedicine. 2016;11:4743–4763. PubMed PMID: 27695330
- ZC K, Güriz H, Ağırbaşlı H, et al. Genotype distribution of Candida albicans isolates by 25S intron analysis with regard to invasiveness. Mycoses. 2004;47(11‐12):465–469.
- Gurbuz M, Kaleli I. Molecular analysis of Candida albicans isolates from clinical specimens. Mycopathologia. 2010;169(4):261–267. Epub 2009/ 12/17. PubMed PMID: 20012366.
- Millar BC, Moore JE, Xu J, et al. Genotypic subgrouping of clinical isolates of Candida albicans and Candida dubliniensis by 25S intron analysis. Lett Appl Microbiol. 2002;35(2):102–106.
- Loesche WJ Association of the oral flora with important medical diseases. Curr Opin Periodontol. 1997;4:21–28. Epub 1997/ 01/01. PubMed PMID: 9655017.
- Andrutis KA, Riggle PJ, Kumamoto CA, et al. Intestinal lesions associated with disseminated candidiasis in an experimental animal model. J Clin Microbiol. 2000;38(6):2317–2323. Epub 2000/ 06/02. PubMed PMID: 10834995; PubMed Central PMCID: PMCPMC86791.
- Kuboniwa M, Tribble GD, Hendrickson EL, et al. Insights into the virulence of oral biofilms: discoveries from proteomics. Expert Rev Proteomics. 2012;9(3):311–323.
- Parahitiyawa NB, Jin LJ, Leung WK, et al. Microbiology of odontogenic bacteremia: beyond endocarditis. Clin Microbiol Rev. 2009;22(1):46–64. PubMed PMID: 19136433