
Abstract
Candida albicans is both a commensal microorganism in healthy individuals and a major fungal pathogen causing high mortality in immunocompromised patients. Yeast-hypha morphological transition is a well known virulence trait of C. albicans. Host innate immunity to C. albicans critically requires pattern recognition receptors (PRRs). In this review, we summarize the PRRs involved in the recognition of C. albicans in epithelial cells, endothelial cells, and phagocytic cells separately. We figure out the differential recognition of yeasts and hyphae, the findings on PRR-deficient mice, and the discoveries on human PRR-related single nucleotide polymorphisms (SNPs).
Candida Albicans and Host Pattern Recognition Receptors (PRRs)
Humans encounter fungi every day, while only a few fungal species may cause infections. In the past 3–4 decades, with the advent of organ transplantation, haematopoietic stem cell transplantation, immunosuppression, chemotherapy, and the dissemination of HIV, the incidence of fungal infections has been rising.Citation1-3 Presently, fungi have become the fourth main cause of hospital-acquired infections.Citation4,5 Among the fungal pathogens, Candida spp is prominent, and Candida albicans is a major pathogen.Citation1,6
C. albicans is a commensal fungus that colonize on gastrointestinal/genital mucosa of mammalians without causing disease in most healthy individuals, but in case the host defense is weakened under certain circumstances can C. albicans become pathogenic.Citation7-10 C. albicans may cause 2 types of infections: superficial infections (such as oral or vaginal candidiasis), and systemic infections (such as life-threatening bloodstream infections/candidaemia).Citation11 Polymorphological transition is the widely known virulence trait of C. albicans,Citation12 and the fungus can grow as white-phase yeast cells, GUT cells, opaque-phase cells, gray-phase cells,Citation13 chlamydospores, ture hyphae, and pseudohyphae.Citation14 C. albicans normally grows in white-phase yeast form when it colonizes on mammalian mucosal surfaces.Citation15 Yeast-to-hypha transition usually indicates a pathogenic status.Citation8,16 In accordance, the yeast form is tolerated by the host immune system, while the invasive hyphal form may induce robust immune responses.Citation17
The cell wall of C. albicans provides targets for host immune system to sense the pathogen and trigger immune response. The cell wall is a matrix of 3 components: chitin, glucans, and mannans. Chitin locates at the most inside of the cell wall and covalently linked to β-glucan. Chitin and β-glucan form hydrogen bonds with each other to construct a tough 3-dimensional inner layer network of microfibrils. The outer layer is constituted by O-linked and N-linked mannose polymers (mannans), with highly glycosylated cell wall proteins attached. Although the basic components of the cell wall are similar, the precise structure and chemical properties are different in different forms of C. albicans, which may facilitate host immune system to recognize different cell forms.Citation18,19
Host innate immunity to C. albicans critically requires pattern recognition receptors (PRRs). PRRs is fundamental in discriminating self from non-self via recognizing vital conserved chemical signatures of pathogens, called pathogen-associated molecular patterns (PAMPs).Citation20As for C. albicans, amounts of cell wall components are PAMPs, including β-glucan, mannans, and cell wall proteins. Besides, some intracellular components, such as specific DNA and RNA can also be recognized as PAMPs. For all kinds of pathogens, 4 major classes of PRRs have been identified: Toll-like receptors (TLRs), C-type lectin receptors (CLRs), Nucleotide-binding oligomerization domain (NOD)-like receptors (NLRs) and retinoic-acid-inducible gene I (RIGI)-like receptors (RLRs).Citation21 TLRs can be divided into cell-membrane-associated receptors (TLR1, TLR2, TLR4, TLR5, and TLR6) and endosomal receptors (TLR3, TLR7, TLR8, and TLR9).Citation21 CLRs are mainly membrane-bound receptors.Citation21NLRs and RLRs are both intracellular receptors.Citation21 In addition, some circulating proteins, such as mannose-binding lectin (MBL), are also considered as PRRs.Citation22 TLRs and CLRs play major roles for the recognition of C. albicans PAMPs; NLRs are also involved in the recognition; while little is known about the role of RLR in the recognition of C. albicans PAMPs up to now (). TLR2, TLR4, and TLR9 directly recognize phospholipomannan (PLM),Citation23 O-linked mannosyl residues,Citation24 and CpG DNACitation25 respectively in C. albicans. TLR7 is required for the recognition of single-stranded RNA of C. albicans.Citation26,27 Of note, some CLRs are involved in the recognition of C. albicans, including Dectin-1, Dectin-2, Dectin-3 (originally named murine macrophage C-type lectin, MCL), the dendritic cell-specific ICAM3-grabbing non-integrin (DC-SIGN), the macrophage mannose receptor (MR), Galectin-3, and the macrophage-inducible C-type lectin (Mincle).Citation28,29 Dectin-1 recognizes β-(1,3)-glucan, while the rest CLRs recognize different mannose-relative structures.Citation28 MBL is considered a soluble CLR. It mediates opsonization and uptake of Candida by phagocytes via binding to both Candida mannan and the C1q receptor on the surface of phagocytes.Citation30 NLRs are involved in the recognition of intracellullar pathogens.Citation31 Interestingly, NOD1 and NOD2, 2 main NLRs important in bacterial peptidoglycans recognition, are not involved in the recognition of C. albicans.Citation32 Nevertheless, NLRP3 plays an important role in C. albicans-induced inflammation.Citation33-37 Several other NLRs, such as NLRP10 and NLRC4, are also revealed to be related to anti-C. albicans responses.Citation38,39 NLRP10 is required to control the disseminated C. albicans infection,Citation38 and NLRC4 functions within the mucosal stroma to control oral C. albicans infection.Citation39
Table 1. Pattern recognition receptors and C. albicans PAMPs
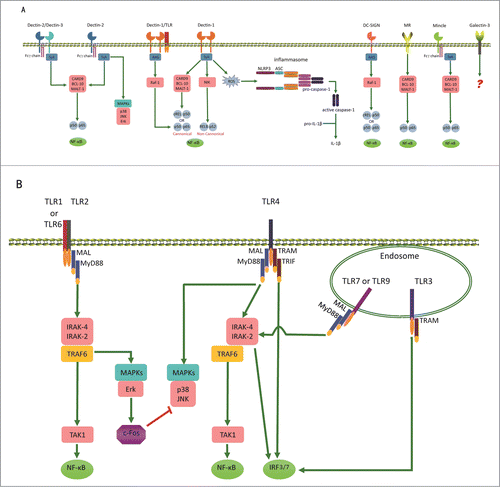
After recognizing C. albicans PAMPs, the recognition signals can be transducted through multiple pathways (). In TLRs signaling pathway, 2 crucial signaling adaptors, MyD88 and TRIF (also known as TICAM-1) are involved. MyD88 binds to almost all TLRs except TLR3.Citation40,41 TRIF binds to TLR3 and TRL4.Citation40,42,43 The activation of MyD88 and TRIF lead to MyD88-dependent pathway and the MyD88-independent (TRIF-dependent) pathway respectively.Citation44 In contrast, CLRs are mostly associated with the spleen tyrosine kinase (SYK) (), which activates MAPK (mitogen-activated protein kinase).Citation45-47 Dectin-1 signals are associated with Syk directly, while Dectin-2 and Mincle couple Syk via the common γ-chain of Fc receptor.Citation48-50 Of note, in the Syk signaling pathway, CARD9 plays a crucial role to induce cytokine induction.Citation51-53 Raf-1 is another kinase involved in CLRs signal transduction, which can be activated by Dectin-1 or DC-SIGN.Citation54,55 Interestingly, most PRRs, if not all, activate the transcription factor NF-κB or IRF3/7 to induce immune responses.Citation56,57 ().
Figure 1. Overview of PRRs for C. albicans recognition. Recognition of C. albicans is mainly mediated by Toll-like receptors (TLRs) and C-type lectin receptors (CLRs). (A) The recognition of C. albicans by some CLRs can stimulate receptor phosphorylation and recruitment of the spleen tyrosine kinase (SYK). The association of dectin-1 with SYK activates assembly of the CARD complex (CARD9, BCL-10 and MALT-1). This results in the release of NF-κB. Syk activation also induces the noncanonical NF-κB pathway mediated by NF-κB-inducing kinase (NIK). Only the Dectin-1 and DC-SIGN recruit Ras for signal transducing, which leads to the release of NF-κB. Dectin-1 recognition of C. albicans can also activate the NLRP3 inflammasome through a mechanism that involves Syk and ROS. (B) The recognition of C. albicans by some TLRs can stimulate MyD88-dependent or TRIF-dependent pathways, leading to the release of NF-κB or the activation of IRF3/7.
Innate immunity is the first defense of host against C. albicans, where several types of cells work against the pathogen: epithelial cells, endothelial cells, and phagocytic cells. These types of cells work in cooperation to sense, kill, and present antigens of the invasive C. albicans. They express specific PRRs, which determine their specific interactions with C. albicans. Here, we summarize the role of PRRs in recognition of C. albicans in each cell type.
PRRs of Epithelial Cells
Mucosal epithelial cells are engaged in the first line of anti-fungal defense. Mucosal epithelial cells not only function as a passive physical barrier to restrain pathogen from invading, but also sense the pathogen and trigger immune responses. As key cells in innate immunity, mucosal epithelial cells express a wide range of PRRs, including TLR1-6 and TLR8-10, Dectin-1, Galectins, and NOD1.Citation58 Although epithelial cells from different sites express similar TLRs, the expression levels may be different. For example, TLR2 mRNA is most abundant in human female genital tract, fallopian tubes and cervical tissues, followed by the endometrium and ectocervix.Citation59 The expression of TLR4 in the epithelial cells of the human female genital tract is controversial. Some observed the presence of TLR4,Citation60-62 while others reported the absence.Citation63,64 In normal human intestinal epithelial cells, TLR2 and TLR4 are expressed at low level.Citation65-68 The differential expression of PRRs at different sites may be related to the different microenvironments and the different roles of PRRs.Citation69,70 For example, TLR2 is revealed to be associated with epithelial growth, survival, and repair.Citation69,70 Of note, although epithelial cells express a wide range of PRRs, not all PRRs are engaged in C. albicans recognition, and only certain TLRs and CLRs on epithelial surfaces are reported to be engaged.Citation71 After culturing epithelial cells with C. albicans yeasts, TLR2, TLR4, TLR6, and TLR9 genes' expression were activated.Citation72 Nevertheless, the activation of human epithelial cell immune response is not likely to be initiated by TLRs, as blocking of these receptors with antibodies didn't alter the epithelial cytokine profile.Citation73 Dectin-1 and Dectin-2 are 2 confirmed receptors for C. albicans recognition.Citation74 However, only Dectin-1 is found expressed in epithelial cells.Citation75 Although some studies on other fungal pathogens have revealed the up-regulation of Dectin-1 in human epithelial cells upon fungal infection,Citation76,77 few studies work on the role of Dectin-1 in C. albicans recognition in epithelial cells up to now.
MAPK and NF-κB pathways are crucial in epithelial cells response to C. albicans.Citation17 MAPK pathway is revealed to identify the invasive C. albicans, while NF-κB pathway is in charge of anti-fungal responses.Citation17 The activation of the pathways would lead to series of biological effects, and the effects can be divided into 2 categories. One is to activate weak but direct anti-fungal defense by secreting antimicrobial peptides (such as β-defensins and LL-37).Citation78,79 The antimicrobial peptides can recruit immune cells to sites of infection/proliferation and additionally can bind to PRRs and influences responses.Citation80 The other is to secrete a profile of pro-inflammatory cytokines and chemokines, which facilitates epithelial cells to cooperate with other types of cells at mucosal surface, including dendritic cells and neutrophils.Citation81 C. albicans stimulates epithelial cells to produce IL-1α/β, IL-6, G-CSF, GM-CSF, TNF-α, and IL-8, while IL-12, IFN-γ, IL-4, and IL-13 are not included.Citation73,81-84 For example, IL-8 was released at high level by C. albicans-infected human oral epithelial cells and it actively recruits neutrophils into mucosal tissues, leading to an anti-fungal defense.Citation73 In primary human keratinocytes, IL-22 plus TNF-α effectively inhibit the growth of C. albicans and maintain the survival of epithelia.Citation85 In addition, human vaginal epithelial cells could express S100 calcium-binding proteins to recruit polymorphonuclear neutrophils (PMNs) to the C. albicans-infected vagina.Citation86 Recently it was shown that S100A8 alarmin is sufficient, but not necessary, to induce PMN migration during experimental vaginal candidiasis.Citation87
PRRs of Endothelial Cells
Endothelium forms a semi-permeable barrier between blood/lymph and the surrounding tissues. Once hematogenously disseminated candidiasis initiates, C. albicans must adhere to and invade the endothelial cell lining to infect the deep tissues.Citation88 Endothelial cells express many PRRs. In detail, immune responsive endothelial cells in healthy arteries express low levels of TLR2 and TLR4.Citation89,90 Human umbilical vein endothelial cells (HUVECs) express TLR3 and TLR9.Citation91-93 Human dermal microvascular endothelial cells express MR.Citation94 Besides, Galection-3 was detected on cultured endothelial cells.Citation95 The expression of Dectin-1 on endothelial cells is controversial. In contrast to its major role on phagocytic cells for fungal β-glucan recognition, Dectin-1 is generally believed not expressed by endothelial cells previously.Citation88,96 However, a recent study observed Dectin-1 on HUVECs.Citation97
C. albicans has a family of specialized proteins (adhesins) with agglutinin-like sequence (ALS). The family contains 8 members (Als1-7 and Als9), and Als3 is vital for C. albicans adherence and invasion.Citation11 Some studies using HUVECs revealed that the ALS proteins mediate the adherence to endothelial cells. After adherence, C. albicans invades endothelial cells through endocytosis. Als3 and Ssa1 on the surface of C. albicans hyphae bind to N-cadherin and other receptors on the surface of endothelial cell, which mediates the endocytosis.Citation98-100 In contrast to the ALS proteins and endocytosis, the interaction between C. albicans PAMPs and PRRs on endothelial cells was rarely investigated. A study revealed that C. albicans triggers proinflammatory gene expression in primary human endothelial cells through TLR3 signaling, NF-κB, and p38 MAPK pathways in response to C. albicans invasion.Citation96 While, CLRs on endothelial cells rarely studied.
PRRs of Phagocytic Cells
Phagocytic cells, including neutrophils, monocytes/macrophages, and dendritic cells (DCs), are believed to be most effective for controlling and clearing C. albicans infection. Neutrophils are considered principal effector cells, followed by monocytes/macrophages.Citation101 Monocytes/macrophages can also present antigens, but they only play an integral role in anti-Candida defense in infection locations. DCs are professional antigen-presenting cells (APCs), which ingest C. albicans PAMPs and present antigens via major histocompatibility complex (MHC) class II molecules. Till now, most findings on the interaction between PRRs and C. albicans PAMPs are obtained through investigations on phagocytic cells.
PRRs of neutrophils and monocytes/macrophages
Neutrophils and monocytes/macrophages are effective in killing invasive fungal cells. They are rapidly recruited at the infection locations.Citation102-105 Monocytes mainly recognize invading fungi in circulation, expressing high levels of TLRs and moderate levels of CLRs.Citation21 When monocytes reside at infected tissues and differentiate into macrophages, they keep on expressing TLRs and up-regulate the expression of CLRs.Citation21 As the expression of PRRs in macrophages can be influenced by cytokines and some other factors, the levels of PRRs in macrophages are variable.Citation21 Neutrophils express TLRs, CLRs, and phagocytic receptors including complement receptor 3 (CR3) and Fcγ receptors (FcγRs).Citation21 Collectively, in these phagocytic cells, TLRs (including TLR2, TLR4, and TLR6), CLRs (including Dectin-1, Dectin-2, MR, Galectin-3, and Mincle), and other fungal-relevant receptors (including FcγR, CR3, CD36, and SCARF1) are expressed, which play roles in recognize various PAMPs from C. albicans.
Some PRRs recognize C. albicans mannans. MR on the surface of macrophages recognizes the N-bound mannans,Citation24 and TLR4 recognizes the O-bound mannans.Citation106 TLR2 and TLR6 interact with phospholipomannan (PLM) directly to initiate proinflammatory cytokine production in the mouse macrophage-like cell line J774.Citation23 Galectin-3 on the surface of murine macrophages can bind to the β-1,2 mannosides of C. albicans.Citation107,108 In human neutrophil, Galectin-3 plays an important role in phagocytosing C. albicans hyphae, but not C. albicans yeasts.Citation109 Dectin-2 is another PRR for phagocytic cells to recognize C. albicans mannans.Citation110-112 By using mouse leukemic monocyte macrophage cell line RAW 264.7, Dectin-2 is found mainly involved in the recognition of C. albicans hyphae and induce intracellular signals through FcγR.Citation49 A recent study revealed that Dectin-3 recognized α-mannans on the surfaces of C. albicans hyphae and induced NF-κB activation in bone marrow-derived macrophages from C57B/L6 mice.Citation29 Dectin-3 constantly forms heterodimers with Dectin-2 for recognizing C. albicans hyphae. Compared to their respective homodimers, Dectin-3 and Dectin-2 heterodimers bound α-mannans more effectively, leading to potent inflammatory responses against fungal infections.Citation29 Another study indicated that Dectin-3 can mediate endocytosis.Citation113 Mincle, another C-type lectin, is expressed predominantly on macrophages. It is shown to play a role in murine macrophage responses to yeast-form C. albicans.Citation114 Similar to Dectin-2, Mincle recognizes C. albicans by selectively binding α-mannose with the association of FcγR.Citation114 In human monocyte and neutrophil, Mincle expression is up-regulated upon C. albicans stimulus and further immune responses are thereby regulated.Citation115
CR3 and Dectin-1 are revealed to recognize C. albicans β-glucans. CR3 is a widely expressed β2-integrin. In human kidney 293 fibroblastoid cell line, CR3 mediates recognition of both the yeast and hyphal forms of C. albicans.Citation116 Moreover, the phagocytosis of C. albicans by human polymorphonuclear leukocytes (PMN) is mainly mediated by CR3.Citation117 Dectin-1 is a myeloid-expressed receptor for β-glucans recognition and expressed systematically on phagocytic cells. On C. albicans yeasts, budding and cell separation may expose β-glucans, thereby Dectin-1 can recognize C. albicans yeasts easily.Citation118,119 Dectin-1 is revealed to be important for the activation of murine PMN by Candida.Citation120 Besides, some investigations indicated that CD36 and SCARF1 may bind C. albicans β-glucan.Citation121 SCARF1 is expressed on macrophages and endothelial cells. To test the ability of SCARF1 to bind and phagocytose fungi, investigators isolated the SCARF1 cDNA from a human endothelial cell cDNA library and established a stable Chinese hamster ovary (CHO) cell line expressing SCARF1 (CHO-SCARF1). They found that CHO-SCARF1 cells bind C. albicans in a β-glucan–dependent manner. Moreover, CD36 is a class B scavenger receptor that is a sensor for endogenous molecules and microbial products that signal via TLR2.Citation121 CHO-CD36 cells bind C. albicans also in a β-glucan–dependent manner.Citation121 CD36-deficient macrophages showed an ∼50% reduction in binding C. albicans relative to WT macrophage,Citation121 and CD36−/− macrophages stimulated with C. albicans had a marked reduction in the expression of IL-1β, TNF, IL-12p40, MIP-2, MIP-1α, MIP-1β, and RANTES.Citation121
The chitin recognition receptors have also been investigated. On human peripheral blood mononuclear cells (PBMCs) and murine macrophages, purified chitin from C. albicans could block the recognition of C. albicans yeast cells.Citation19 A recent study revealed that purified chitin particles derived from C. albicans led to the selective secretion of the anti-inflammatory cytokine IL-10 on murine bone marrow-derived macrophages,Citation122 and NOD2, TLR9 and MR are essential fungal chitin-recognition receptors for this response.Citation122
The recognition of C. albicans by PRRs is generally thought to occur at the phagocytic cell surface and this process leads to the phagocytosis of C. albicans and the formation of an intracellular vacuole called phagosome. Dectin-1,Citation48,123 CR3,Citation117 MR,Citation124 Galectin-3,Citation109 and possibly TLR2 Citation109 have been identified as receptors involved in phagocytosis of C. albicans. For example, Galectin-3 antibody significantly inhibited neutrophil phagocytosis of C. albicans hyphae,Citation109 and exogenous galectin-3 increases phagocytosis of C. albicans yeast.Citation109 Actually, PRRs not only mediate the opsonised fungi uptake, but also the recognition of C. albicans components at the phagosomes. Dectin-1 not only controls internalization of β-1,3-glucan containing phagosomes and triggers proinflammatory cytokines, but also acts as a master regulator for subsequent phagolysosomal maturation through Syk activation.Citation125 TLR9 and TLR7 are both endosomal receptors, and not expressed on cell surface. TLR9 mediate the sensing of C. albicans unmethylated genomic DNA.Citation25 By using TLR9 knockout (TLR9KO) macrophages, TLR9 is revealed to be activated by fungal DNA and modulate macrophage anti-fungal effect.Citation126 TLR7 is required for the recognition of single-stranded RNA from C. albicans and the mice lacking TLR7 were hypersusceptible to systemic C. albicans infection.Citation26,27 Of note, inflammasome plays an important role in antifungal immune response. Inflammasome is a cytoplasmic proteolytic multimeric protein complex expressed in myeloid cells, and consists of NLRs and several adaptors.Citation127 Two NLRs, NLRP3 and NLRC4, are implicated in mediating responses to C. albicans, while only NLRP3 is involved in preventing subsequent dissemination of this pathogen.Citation34,36,37,39,128 C. albicans are able to trigger proinflammatory cytokine IL-1β and IL-18 production via NLRP3 inflammasome in murine monocytes, macrophages and dendritic cells.Citation129 Consistently, downregulation of NLRP3 by RNA interference strongly reduced the secretion of bioactive IL-1β.Citation129 Pyroptosis is an inflammasome-mediated programmed cell death. In bone marrow-derived macrophages and murine J774 macrophages, NLRP3 can be triggered by C. albicans, leading to NLRP3-mediated pyroptosis.Citation130
As multiple PRRs are involved in the recognition of C. albicans, it is reasonable that “cross-talk” can occur between these receptors.Citation131 Dectin-1 is shown to collaborate with TLR2, 4, 5, 7 or 9 to synergistically influence the secretion of many cytokines (inducing IL-23, while repressing IL-12).Citation132,133 In human PBMCs, Dectin-1/TLR2 pathway was able to amplify MR-induced IL-17 production, a vital cytokine upon C. albicans stimulus. This finding provides an evidence for the interaction between C-type lectin receptors and TLRs.Citation134 Galectin-3 on the surface of murine macrophages specifically recognizes C. albicans, which needs the association of TLR2 for signaling.Citation108 Interestingly, Galectin-3 also works in association with Dectin-1 on macrophages.Citation135 When macrophages expressing Dectin-1 are exposed to C. albicans mutants with increased exposure of β-glucan, the loss of Galectin-3 dramatically accentuates the failure to trigger an appropriate TNF-α response.Citation135 Moreover, the collaboration in immune response may also involve other cell receptors, such as peroxisome proliferator-activated receptor (PPAR)-γ and CD44. PPAR-γ is expressed in various immune cells and acts as a transcriptional repressor to inhibit the transcription of many proinflammatory cytokines.Citation136 PPAR-γ has been implicated in the negative regulation of IFN-β production in TLR3- and TLR4-stimulated peritoneal primary macrophages.Citation136 CD44 is a major hyaluronan receptor distributed in different tissues and alerts cell to injury. CD44 is found to regulate TLR2-mediated immune responses in murine bone marrow-derived macrophages.Citation137 The collaborations between the receptors, downstream signaling pathways, and cytokine production are complicated. More studies are needed to reveal much more underlying “cross-talk.”
After recognition of invasive fungi, neutrophils and monocytes/macrophages use a number of oxidative and non-oxidative mechanisms to kill extracellular and internalized fungi. The respiratory burst is thought to be a major anti-fungal defense mechanism. Besides producing reactive oxygen intermediates, the production of reactive nitrogen intermediates is another oxidative system possessing fungicidal activity. This system can be induced by PRRs and cytokines, leading to the killing of fungi.Citation138,139 The interaction between phagocyte PPRs and C. albicans plays an important role in determining the cytokine and chemokine profiles. For example, Dectin-1 induces the production of inflammatory mediators, including eicosanoids, TNF-α, IL-1β, IL-6, IL-23, CCL2, CXCL1 and CCL3.Citation138,140,141 Fungal recognition by Dectin-2 can induce the production of numerous cytokines, including TNF-α, IL-1Ra, IL-2, IL-10, IL-6, IL-1β, IL-12 and IL-23, and possibly cysteinyl leukotrienes.Citation49,51,142,143 Similarly, fungal recognition by Mincle induces the production of cytokines, including MIP-2, KC, IL-10 and TNF-α.Citation107,114
PRRs of dendritic cells (DCs)
DCs are specific phagocytic cells, existing in tissues in contact with external environment, such as the skin and mucosal surface. Besides ingesting and killing C. albicans, DCs mainly act as professional antigen-presenting cells (APCs), which bridge innate and adaptive immunity by shaping the T cell response following PRR-dependent cytokine production. Only DCs are able to prime naive T cells to generate life-long memory immunity. DCs express a myriad of PRRs involved in C. albicans recognition, including TLRs, CLRs, FcγR, and CR3. The expression of PRRs on DCs is variable in different situations. For example, immature DCs express high level of FcγR, CRs, and TLRs, while mature DCs express less. Human plasmacytoid DCs express endosomal TLR7 and TLR9, but lack the expression of TLR4.Citation144 However, whether the differences influence the recognition of C. albicans is rarely investigated.
The PRRs on DCs for C. albicans recognition are similar with those on phagocytic cells. Here, we emphatically summarize the findings on DCs. TLR2 and TLR4 can recognize a 65 kDa cell surface mannoprotein (MP65) from C. albicans.Citation145 After the recognition via TLR2 and TLR4, DCs increase the secretion of TNF-α, IL-6, IL-12 and the expression of CD14 and FcγR.Citation145 TLR1 was indicated not involved in the recognition of C. albicans, while TLR6 might play a role in modulating the balance between Th1 and Th2 cytokines.Citation146 The TLR6 knock-out mice displayed a defective production of IL-10 and an increased IFN-γ release.Citation146 These results indirectly suggest that DCs might regulate adaptive immunity through TLR6. Whether the phagosomal TLRs (such as TLR3 and TLR7) on DCs were involved in recognizing RNA of C. albicans is still unclear. Nevertheless, murine conventional DCs mount a type-I IFN response against C. albicans requiring phagosomal TLR7-mediated IFN-β signaling.Citation147 Moreover, it is suggested that C. albicans activates murine bone marrow-derived myeloid DCs through a TLR9-mediated signaling pathway.Citation25 Dectin-1 was well characterized on DCs. It was found that Dectin-1 was high expressed on the DC cell line XS52, but less on the macrophage cell line J774.Citation148 A recent study showed that Dectin-1 on mouse renal DCs senses C. albicans and induces IFN-β production through Dectin-1-Syk-IRF5 signaling.Citation149 Dectin-2 also contributes to DCs activation.Citation142 Dectin-2 is important in murine immune defense against C. albicans by inducing Th17 cell differentiation via Fcγ chain and Syk-CARD9-NF-κB-depedent signaling pathway.Citation51 Although Dectin-2 was found to bind hyphal components of C. albicans preferentially, both yeast and hyphal C. albicans can induce Th17 differentiation.Citation51 DC-SIGN also plays roles in the immune response to C. albicans. It was demonstrated that DC-SIGN is able to bind C. albicans in human monocyte-derived DCs.Citation150 The binding was shown to be time- as well as concentration-dependent, and live as well as heat-inactivated C. albicans were bound to the same extent.Citation150 Interestingly, N-linked mannan, but not O-linked or phosphomannan, is specifically recognized by DC-SIGN on human DCs and directly influences the production of the proinflammatory cytokine IL-6.Citation151 Galectin-3 on murine bone marrow-derived DCs was recently found to be able to modulate Th17 responses to C. albicans.Citation152 Galectin-3 is encoded by GAL3 gene. Under condition that C. albicans was mixed with gal3-/- or gal3+/+ DCs for 3 days, and then intravenously injected into wild-type mice, the levels of IL-6, TGF-β, IL-23, and IL-17 in mice receiving gal3-/- DCs were significantly higher than those receiving gal3+/+ DCs,Citation152 indicating the modulatory role of Galectin-3. MR is widely expressed on DC subsets and has been shown to play a role in the recognition of Candida.Citation153 A recent study revealed that MR promotes DCs to form “fungipod” upon contacting yeast-form Candida. “fungipod” is a dorsal pseudopodial protrusion and phagocytic structure of DCs, and the formation of this structure is propelled by the robust actin cytoskeleton growth at the DC-yeast contact site. MR is required for the formation of “fungipod,” while Dectin-1 is not.Citation154 Of note, not all Candida species can induce the formation of “fungipod." The human pathogen C. parapsilosis induces DC fungipod formation strongly, but the response is species specific since the related fungal pathogens C. tropicalis and C. albicans induce very few and no fungipods, respectively. Besides, NLRP3 is also involved in C. albicans recognition of DCs. In mouse BM-DCs, β-glucan can activate NLRP3 inflammasome, and this activation is dispensable for antigen specific Th1 and Th17 polarization.Citation35
Previously, it is a general thought that DCs have little effect on innate resistance to fungal infection. However, a recent study revealed that selective loss of Syk in DCs abrogates innate resistance to acute systemic C. albicans infection in mice,Citation155 which indicates that a single kinase in DCs might orchestrate a complex series of molecular and cellular events in innate resistance to C. albicans. Actually, full protection against most pathogens requires an adaptive response, which is initiated and directed by DCs.Citation156 DCs respond to in situ and released C. albicans PAMPs via PRRs (MR, Dectin-1, TLR2, Galectin-3 etc).Citation134 The PAMPs of C. albicans are then taken up, processed, and presented in an MHC class II-restricted fashion to naive T cells. An in-vitro study showed that the addition of β-glucan to the DCs promoted the activation and maturation of human DCs.Citation157 DCs would present the C. albicans-specific antigens to T cells. In addition to priming T cell responses via antigen presentation, DCs also shape T cell responses through secretion of cytokines.Citation158 The presentation of C. albicans antigen by DCs leads to the activation of CD4+ and CD8+ T cells.Citation159,160 CD8+ T cells process direct cytolytic activity and inhibit the growth of C. albicans hyphae in vitro,Citation160 while CD4+ T-cells are thought to be predominant in adaptive response to C. albicans infection. CD4+ T cells are mainly polarized into specific subsets characterized as Th1, Th2, Th17 or Tregs, each of which is dictated by the cytokines and microenvironment.Citation159 In contrast to the phenotype of Th2, which is closely associated with increased growth and dissemination of the fungus, Th17/Th1 cells are important in reactions against C. albicans.Citation161 IL-23 promotes terminal differentiation and expansion of the Th17 cells,Citation162 and accumulating evidence indicates that Th17 cells are key cells in the response to C. albicans infection.Citation45,163,164 Th17 cells secrete numerous cytokines including IL-17A, IL-17F and IL-22, and play vital roles for immune protection against C. albicans at the majority of mucosal sites in the body.Citation165,166 The importance of Th17 cells in anti-Candida responses is not only portrayed by data from mice models, but also from human patients. Patients with deficiency in Th17-mediated adaptive immunity frequently suffer from chronic mucocutaneous candidiasis.Citation167-171
Differential Recognition of Yeasts and Hyphae
Invasive hyphal form of C. albicans is more pathogenic and tends to induce robust immune responses, while yeast form is more likely to lead to commensalism.Citation172 More specifically, C. albicans hyphae are believed as an infection form to penetrate the epithelial cell layers, whereas yeast cells are generally found either on the epithelial cell surface or at the local tissues after the penetration.Citation173,174 Host cells discriminate the morphologies of C. albicans and response accordingly. Furthermore, there is a threshold for the amount of C. albicans that can be tolerated by the host.Citation175 A study revealed that oral epithelial cells orchestrate an innate response to C. albicans via NF-κB and a biphasic MAPK response. Activation of NF-κB and the first MAPK phase, constituting c-Jun activation, is independent of morphology and due to fungal cell wall recognition. Activation of the second MAPK phase, constituting MKP1 and c-Fos activation, is dependent upon hypha formation and fungal burdens and correlates with proinflammatory responses. MAPK/MKP1/c-Fos activation may be critical for identifying and responding to the pathogenic switch of commensal microbes.Citation17
An open question is what exact PAMPs on different forms of C. albicans are presented to immune cells. To date, this question is partly answered. Glucans are key fungal PAMPs.Citation18,19,118 A recent study revealed that glucans from C. albicans hyphae are different from those from yeast cells. Hyphal glucan induces robust immune responses in human PBMCs and macrophages via a Dectin-1-dependent mechanism. In contrast, C. albicans yeast glucan is a much less potent stimulus.Citation176 On ordinary yeast cells, cell wall β-glucan of C. albicans is largely shielded by outer wall components.Citation118 Neverthelss, the yeast budding and cell separation create permanent scars, which expose β-glucan to trigger antimicrobial responses.Citation118 Unmasking of C. albicans β-glucan to PRRs during yeast-to-hypha switch may lead to robust immune responses. A recent study demonstrated that phospholipids phosphatidylserine (PS) is an component masking β-glucan of C. albicans.Citation177 The mutant species lacking PS exhibits increases in exposure of β-(1-3)-glucan, which leads to greater binding by Dectin-1 in both yeast and hyphal forms.Citation177 The unmasking of β-(1-3)-glucan results in increased elicitation of TNF-α from macrophages in a Dectin-1-dependent manner.Citation177 In addition to glucans, PLMs of C. albicans are able to evoke a proinflammatory state in murine macrophage, which, however, in part depends on their glycosylation status.Citation178 Thus, it can be inferred that the PRRs described above might be able to discriminate the subtle differences of PAMPs after morphological transition and lead to corresponding immune responses. Further studies are expected to reveal the detailed mechanism of PRRs-PAMP interactions during morphological transitions.
The Roles of PRRs: Findings Using PRR-Deficient Mice
In the sections above, the roles of PRRs in the recognition of C. albicans in various cell types are discussed in detail. In this section, we update the findings using in-vivo murine models ().
Table 2. Comparision of PRR-deficient mice and human PRRs genetic polymorphisms
TLR2-/- mice infected with C. albicans intraperitoneally or intravenously exhibit significantly decreased survival compared with the wild-type mice. And this effect is associated with decreased production of TNF-α and chemokines.Citation179 Accordingly, in an intraperitoneal model of candidiasis, fungal clearance is delayed in mice lacking TLR2.Citation180 However, a few other studies showed controversial results. Upon intravenous C. albicans infection, mice lacking TLR2 have an improved fungal clearance and better survival compared to wild-type mice.Citation181,182 The susceptibility of TLR4-lacking mice to C. albicans infection depends on the C. albicans infection route and growth form. TLR4-defective C3H/HeJ mice have been reported to be more susceptible to disseminated candidiasis.Citation183 Similar results were obtained in models of intragastric infection and intravenous re-infection.Citation182 However in intravenous infection models, TLR4-/- mice survived longer upon C. albicans hyphae infection,Citation182 while no difference was observed between the TLR4-/- and wild-type mice upon C. albicans yeasts infection.Citation183 TLR7-deficient mice are more susceptible to systemic infections by low doses of C. albicans. However, when challenged with higher doses, no significant difference was observed between TLR7-deficient mice and wild-type mice.Citation27 Similarly, using high dose challenges, TLR9-/- mice showed no significant alteration in survival upon clinical C. albicans isolate infection.Citation25,27 However, increased susceptibility to systemic candidiasis and impaired fungal clearance in TLR9-/- mice were observed upon challenge with a lower dose of C. albicans.Citation27
Taylor PR et al.Citation184 investigatedβ-glucan recognition and control of fungal infection using Dectin1-/- mice, and revealed that cytokine release, phagocyte recruitment, phagocytosis and microbial killing are all impaired in the Dectin1-/- mice compared with wild-type mice in disseminated candidiasis model. However, another study carried out by Saijo S et al.Citation185 using Dectin1-/- mice suggested that Dectin-1 is not required for host defense against C. albicans. The different conclusion may be caused by different experimental design. Saijo inoculated much more C. albicans to the mice intravenously (1 × 106 or 5 × 105 C. albicans) than Taylor PR et al did (1 × 104 or 1 × 105 C. albicans), which may lead to the different conclusions. Dectin-3-deficient mice are highly susceptible to C. albicans infection.Citation29 With a low dose of C. albicans infection, the survival rate of Dectin-3-deficient mice is significant lower than wild-type mice. But with a higher dose, only a slight difference was observed regarding the survival rate of wild-type mice and Dectin-3-deficient mice.Citation29 MR-/- mice have also been used in studies. After challenging MR-/- mice and wild-type mice intraperitoneally with C. albicans, no significant difference was found in survival between the mice. However, MR-/- mice has higher average fungal burdens in some of the organs but exhibited more competence in inflammatory cell recruitment and antibody production.Citation179 Galectin-3-deficient mice (gal3-/-) were more susceptible to C. albicans infection than wild-type (WT) mice. More specifically, gal3-/- mice died significantly faster and exhibited a trend toward increased fungal burden and increased abscess formation in infected brains compared to WT mice.Citation186
NLRP3 has been found critical for the control of C. albicans infections.Citation35 Mice deficient in NLRP3 displayed diminished serum IL-1β, reduced survival and higher fungal burdens in kidney, spleen, liver, and lung upon C. albicans infection.Citation34,37,187 In a murine model of oral C. albicans infection, NLRP3 inflammasome was necessary to prevent systemic dissemination of C. albicans.Citation37 The NLRP3 inflammasome modulates not only the innate immunity, but also the adaptive immunity. After exposed to disseminated candidiasis, the mice deficient in NLRP3 inflammasome display diminished Th1/Th17 responses, followed by increased fungal outgrowth and lower survival.Citation36 In addition, NLRP10 was found essential for protective anti-fungal adaptive immunity against C. albicans.Citation38 NLRP10-deficient mice had increased susceptibility to disseminated candidiasis, which is indicated by decreased survival and increased fungal burdens. Further investigation revealed that NLRP10-deficient mice also displayed a defect in the generation of Th1/Th17 responses.Citation38
As many PPRs share same adaptors for signal introduction, the mice lacking these adaptors also influence host responses to C. albicans. MyD88 is an important adaptor of TLRs for signal introduction. Moreover, MyD88 is also the receptor of IL-1, IL-18, and IL-33.Citation182,188 Thus, MyD88 plays a key role in innate immune system. Mice lacking MyD88 are hypersensitive to systemic C. albicans infection.Citation188 TRIF is another important signaling adaptor of TLRs. In intra-gastric infection models, mice lacking TRIF fail to prevent C. albicans from infecting peripheral organs.Citation189 Some data support that TRIF pathways promote tolerance to C. albicans, whereas MyD88 is engaged in anti-fungal response.Citation190 CARD9 plays an essential role in Dectin1-Syk-CARD9 pathway.Citation53 CARD9-/- mice are more susceptible to disseminated candidiasis.Citation53 Collectively, MyD88, TRIF, and CARD9 are all important in response to C. albicans infection based on findings on gene knockout mice.
More in-vivo studies are still needed to better understand the role of PRRs. Firstly, the conclusions on the role of some specific PRRs are still inconsistent. The inconsistence may be caused by the differences in experimental design and materials. Another example is that there are conflicting results regarding the presence of Dectin-1 on human PBMCs in vitro and animal Candida infection models.Citation191,192 Increased Dectin-1 expression was demonstrated in animal fungal sepsis models.Citation191 In contrast, in vitro dectin-1 expression on cell surfaces was diminished because of internalization of β-glucan coupled with the corresponding Dectin-1 receptors.Citation192 More in-vivo studies are needed to reveal the exact roles of PRRs. One more example is that live C. albicans and heat-killed C. albicans may lead to different Th17 responses.Citation193 In mammalian cells, 2 pathways of tryptophan metabolization have been described. One pathway leads to the L-kynurenine synthesis and thereafter to niacin as the end metabolite. The second pathway is mediated by the enzymatic activity of tryptophan hydroxylase, producing 5-hydroxytryptophan, followed by further metabolization into serotonin and melatonin.Citation193 Under conditions that PBMCs were incubated with C. albicans, live C. albicans can shift tryptophan metabolism away from kynurenines and toward 5-hydroxytryptophan metabolites. Five-hydroxytryptophan metabolites inhibit IL-17 production.Citation193 Thus, live C. albicans may inhibit host Th17 responses, and experiments using live C. albicans, heat-killed C. albicans or PAMPs may lead to different conclusions. Secondly, more in-vivo research data are needed to better understand oral/vaginal candidiasis and host immune responses. Presently, most in-vivo studies use disseminated candidiasis animal models, while few studies focus on oral or vaginal candidiasis. C. albicans colonizes on gastrointestinal or genital mucosa as a constituent of microbiota. The situation at local mucosa is rather complicated. For example, a recent report revealed that Lactobacillus crispatus modulates epithelial cell defense against C. albicans through TLR2 and TLR4, IL-8 and human β-defensins 2 and 3.Citation194 It can be inferred that more in-vivo studies with the consideration of microbiota on mucosa will shed new insights into the interaction between C. albicans and host immunity. Thirdly, some studies have opened a door to study the collaborations/coordinations among PRRs, while more in-vivo studies are needed to provide more meaningful information.
The Roles of PRRs: Studies on Human SNPs
Regarding the immunity against C. albicans, differences do exist between human and mice (). Although studies using murine models have provide a lot of information on the role of PRRs, findings on human beings are especially valuable. After analyzing SNPs in genes encoding TLRs, MyD88 and Mal in patients with candidemia and controls, TLR1 SNPs (R80T, S248N, I602S) were found associated with candidemia susceptibility, while no association was revealed between the susceptibility and SNPs in TLR2, TLR4, TLR6, TLR9, MyD88 and Mal.Citation195 Another study revealed that L412F, a TLR3 SNP, is associated with increased prevalence of chronic candidiasis.Citation196 By testing patients’ PBMCs for secretion of cytokines, cells carrying the L412F variant showed reduced IFN-γ and TNF-α secretion in responses to C. albicans.Citation196 A study on patients with recurrent vulvovaginal candidiasis (RVVC) revealed that a non-synonymous polymorphism in TLR2 (P631H) significantly increase susceptibility to RVVC, while SNPs in TLR1, TLR4, and CARD9 did not affect the susceptibility to RVVC.Citation197 The Dectin-1 Y238X polymorphism was revealed to express defective Dectin-1.Citation198 Patients with the homozygous mutation are more susceptible to mucocutaneous fungal infections (RVVC or onychomycosis).Citation198 After performing genetic studies on a family with susceptibility to fungal infections, a homozygous Q295X mutation in CARD9 is associated with susceptibility to chronic mucocutaneous candidiasis.Citation170
These studies indicate that PRRs are actively involved in antifungal response in humans, and the roles of PRRs in humans and mice are different. There is no doubt that TLR2 is essential in defense against C. albicans in mice.Citation179,181,182 In contrast to the findings on mice, the study on human SNPs suggested that mutations of TLR2 did not increase the risk to candidemia.Citation195 Few studies reported the importance of TLR1 and TLR3 in defense against C. albicans in murine models, but the mutations of TLR1 or TLR3 increase the susceptibility to C. albicans in humans.Citation195,196 Mice lacking MyD88 are hypersensitive to systemic C. albicans infection,Citation188 but humans with autosomal recessive MyD88 deficiency are resistant to fungal infections normally.Citation199
Of note, it is still immature to make conclusions on the role of most PRRs in human and further studies are necessary. Firstly, only a small number of patients/volunteers are included in most studies up to now, and the evidence is not as solid as we expected. Secondly, the roles of PRRs in different sites of human bodies may be different. For example, patients with mutations of TLR1, but not TLR2, were found susceptibile to candidemia,Citation195 while a non-synonymous polymorphism in TLR2, but not TLR1, significantly enhances susceptibility to mucocutaneous C. albicans infection.Citation197
Conclusions and Future Perspectives
The roles of many PRRs in innate recognition of C. albicans have been investigted. Some in-vitro studies indicate that TLRs and CLRs play a non-negligible role in C. albicans recognition, while NLRs (especially NLRP3), and some other receptors (such as FcγR, CR3, CD36, and SCARF1) are involved in. Nevertheless, further studies are expected to reveal the different recognition mechanisms between yeast and hyphal cells, and more in-vivo studies are necessary to better understand the exact roles of PPRs. Beside, the cross-talks between various PRRs as well as between epithelial cells and phagocytic cells should draw more attentions. Of note, there are some crucial questions remain unanswered. For example, what are the differences between human and mice regarding innate recognition of C. albicans? Further studies on the role of PRRs will open up new avenues for preventing and treating Candida infections.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Funding
This work was supported by National Natural Science Foundation of China (81273558), National Key Basic Research Program of China (2013CB531602), major project of National Natural Science Foundation of China (81330083), and the Shanghai Pujiang Program (14PJD001).
References
- Pfaller MA, Diekema DJ. Epidemiology of invasive candidiasis: a persistent public health problem. Clin Microbiol Rev 2007; 20:133-63; PMID:17223626; http://dx.doi.org/10.1128/CMR.00029-06
- Pfaller MA, Diekema DJ. Epidemiology of invasive mycoses in North America. Crit Rev Microbiol 2010; 36:1-53; PMID:20088682; http://dx.doi.org/10.3109/10408410903241444
- Pagano L, Caira M, Nosari A, Van Lint MT, Candoni A, Offidani M, Aloisi T, Irrera G, Bonini A, Picardi M, et al. Fungal infections in recipients of hematopoietic stem cell transplants: results of the SEIFEM B-2004 study–Sorveglianza Epidemiologica Infezioni Fungine Nelle Emopatie Maligne. Clin Infect Dis 2007; 45:1161-70; PMID:17918077; http://dx.doi.org/10.1086/522189
- Perlin DS. Current perspectives on echinocandin class drugs. Future Microbiol 2011; 6:441-57; PMID:21526945; http://dx.doi.org/10.2217/fmb.11.19
- Pfaller MA. Antifungal drug resistance: mechanisms, epidemiology, and consequences for treatment. Am J Med 2012; 125:S3-13; PMID:22196207; http://dx.doi.org/10.1016/j.amjmed.2011.11.001
- Yapar N. Epidemiology and risk factors for invasive candidiasis. Ther Clin Risk Manag 2014; 10:95-105; PMID:24611015; http://dx.doi.org/10.2147/TCRM.S40160
- Iliev ID, Funari VA, Taylor KD, Nguyen Q, Reyes CN, Strom SP, Brown J, Becker CA, Fleshner PR, Dubinsky M, et al. Interactions between commensal fungi and the C-type lectin receptor Dectin-1 influence colitis. Science 2012; 336:1314-7; PMID:22674328; http://dx.doi.org/10.1126/science.1221789
- Wachtler B, Wilson D, Haedicke K, Dalle F, Hube B. From attachment to damage: defined genes of Candida albicans mediate adhesion, invasion and damage during interaction with oral epithelial cells. PLoS One 2011; 6:e17046; PMID:21407800; http://dx.doi.org/10.1371/journal.pone.0017046
- Ghannoum MA, Jurevic RJ, Mukherjee PK, Cui F, Sikaroodi M, Naqvi A, Gillevet PM. Characterization of the oral fungal microbiome (mycobiome) in healthy individuals. PLoS Pathog 2010; 6:e1000713; PMID:20072605; http://dx.doi.org/10.1371/journal.ppat.1000713
- White SJ, Rosenbach A, Lephart P, Nguyen D, Benjamin A, Tzipori S, Whiteway M, Mecsas J, Kumamoto CA. Self-regulation of Candida albicans population size during GI colonization. PLoS Pathog 2007; 3:e184; PMID:18069889; http://dx.doi.org/10.1371/journal.ppat.0030184
- Mayer FL, Wilson D, Hube B. Candida albicans pathogenicity mechanisms. Virulence 2013; 4:119-28; PMID:23302789; http://dx.doi.org/10.4161/viru.22913
- Sudbery P, Gow N, Berman J. The distinct morphogenic states of Candida albicans. Trends Microbiol 2004; 12:317-24; PMID:15223059; http://dx.doi.org/10.1016/j.tim.2004.05.008
- Tao L, Du H, Guan G, Dai Y, Nobile CJ, Liang W, Cao C, Zhang Q, Zhong J, Huang G. Discovery of a "white-gray-opaque" tristable phenotypic switching system in candida albicans: roles of non-genetic diversity in host adaptation. PLoS Biol 2014; 12:e1001830; PMID:24691005; http://dx.doi.org/10.1371/journal.pbio.1001830
- Gow NA. A developmental program for Candida commensalism. Nat Genet 2013; 45:967-8; PMID:23985683; http://dx.doi.org/10.1038/ng.2737
- Klein BS, Tebbets B. Dimorphism and virulence in fungi. Curr Opin Microbiol 2007; 10:314-9; PMID:17719267; http://dx.doi.org/10.1016/j.mib.2007.04.002
- Dwivedi P, Thompson A, Xie Z, Kashleva H, Ganguly S, Mitchell AP, Dongari-Bagtzoglou A. Role of Bcr1-activated genes Hwp1 and Hyr1 in Candida albicans oral mucosal biofilms and neutrophil evasion. PLoS One 2011; 6:e16218; PMID:21283544; http://dx.doi.org/10.1371/journal.pone.0016218
- Moyes DL, Runglall M, Murciano C, Shen C, Nayar D, Thavaraj S, Kohli A, Islam A, Mora-Montes H, Challacombe SJ, et al. A biphasic innate immune MAPK response discriminates between the yeast and hyphal forms of Candida albicans in epithelial cells. Cell Host Microbe 2010; 8:225-35; PMID:20833374; http://dx.doi.org/10.1016/j.chom.2010.08.002
- Nather K, Munro CA. Generating cell surface diversity in Candida albicans and other fungal pathogens. FEMS Microbiol Lett 2008; 285:137-45; PMID:18616597; http://dx.doi.org/10.1111/j.1574-6968.2008.01263.x
- Mora-Montes HM, Netea MG, Ferwerda G, Lenardon MD, Brown GD, Mistry AR, Kullberg BJ, O'Callaghan CA, Sheth CC, Odds FC, et al. Recognition and blocking of innate immunity cells by Candida albicans chitin. Infect Immun 2011; 79:1961-70; PMID:21357722; http://dx.doi.org/10.1128/IAI.01282-10
- Medzhitov R, Janeway CA Jr. Innate immunity: the virtues of a nonclonal system of recognition. Cell 1997; 91:295-8; PMID:9363937; http://dx.doi.org/10.1016/S0092-8674(00)80412-2
- Netea MG, Brown GD, Kullberg BJ, Gow NA. An integrated model of the recognition of Candida albicans by the innate immune system. Nat Rev Microbiol 2008; 6:67-78; PMID:18079743; http://dx.doi.org/10.1038/nrmicro1815
- Cheng SC, Joosten LA, Kullberg BJ, Netea MG. Interplay between Candida albicans and the mammalian innate host defense. Infect Immun 2012; 80:1304-13; PMID:22252867; http://dx.doi.org/10.1128/IAI.06146-11
- Jouault T, Ibata-Ombetta S, Takeuchi O, Trinel PA, Sacchetti P, Lefebvre P, Akira S, Poulain D. Candida albicans phospholipomannan is sensed through toll-like receptors. J Infect Dis 2003; 188:165-72; PMID:12825186; http://dx.doi.org/10.1086/375784
- Netea MG, Gow NA, Munro CA, Bates S, Collins C, Ferwerda G, Hobson RP, Bertram G, Hughes HB, Jansen T, et al. Immune sensing of Candida albicans requires cooperative recognition of mannans and glucans by lectin and Toll-like receptors. J Clin Invest 2006; 116:1642-50; PMID:16710478; http://dx.doi.org/10.1172/JCI27114
- Miyazato A, Nakamura K, Yamamoto N, Mora-Montes HM, Tanaka M, Abe Y, Tanno D, Inden K, Gang X, Ishii K, et al. Toll-like receptor 9-dependent activation of myeloid dendritic cells by Deoxynucleic acids from Candida albicans. Infect Immun 2009; 77:3056-64; PMID:19433551; http://dx.doi.org/10.1128/IAI.00840-08
- Biondo C, Signorino G, Costa A, Midiri A, Gerace E, Galbo R, Bellantoni A, Malara A, Beninati C, Teti G, et al. Recognition of yeast nucleic acids triggers a host-protective type I interferon response. Eur J Immunol 2011; 41:1969-79; PMID:21480215; http://dx.doi.org/10.1002/eji.201141490
- Biondo C, Malara A, Costa A, Signorino G, Cardile F, Midiri A, Galbo R, Papasergi S, Domina M, Pugliese M, et al. Recognition of fungal RNA by TLR7 has a nonredundant role in host defense against experimental candidiasis. Eur J Immunol 2012; 42:2632-43; PMID:22777843; http://dx.doi.org/10.1002/eji.201242532
- Gow NA, van de Veerdonk FL, Brown AJ, Netea MG. Candida albicans morphogenesis and host defence: discriminating invasion from colonization. Nat Rev Microbiol 2012; 10:112-22; PMID:22158429.
- Zhu LL, Zhao XQ, Jiang C, You Y, Chen XP, Jiang YY, Jia XM, Lin X. C-type lectin receptors Dectin-3 and Dectin-2 form a heterodimeric pattern-recognition receptor for host defense against fungal infection. Immunity 2013; 39:324-34; PMID:23911656; http://dx.doi.org/10.1016/j.immuni.2013.05.017
- Brouwer N, Dolman KM, van Houdt M, Sta M, Roos D, Kuijpers TW. Mannose-binding lectin (MBL) facilitates opsonophagocytosis of yeasts but not of bacteria despite MBL binding. J Immunol 2008; 180:4124-32; PMID:18322223; http://dx.doi.org/10.4049/jimmunol.180.6.4124
- Brodsky IE, Monack D. NLR-mediated control of inflammasome assembly in the host response against bacterial pathogens. Semin Immunol 2009; 21:199-207; PMID:19539499; http://dx.doi.org/10.1016/j.smim.2009.05.007
- van der Graaf CA, Netea MG, Franke B, Girardin SE, van der Meer JW, Kullberg BJ. Nucleotide oligomerization domain 2 (Nod2) is not involved in the pattern recognition of Candida albicans. Clin Vaccine Immunol 2006; 13:423-5; PMID:16522788; http://dx.doi.org/10.1128/CVI.13.3.423-425.2006
- Vonk AG, Netea MG, van Krieken JH, Iwakura Y, van der Meer JW, Kullberg BJ. Endogenous interleukin (IL)-1 alpha and IL-1 beta are crucial for host defense against disseminated candidiasis. J Infect Dis 2006; 193:1419-26; PMID:16619190; http://dx.doi.org/10.1086/503363
- Gross O, Poeck H, Bscheider M, Dostert C, Hannesschlager N, Endres S, Hartmann G, Tardivel A, Schweighoffer E, Tybulewicz V, et al. Syk kinase signalling couples to the Nlrp3 inflammasome for anti-fungal host defence. Nature 2009; 459:433-6; PMID:19339971; http://dx.doi.org/10.1038/nature07965
- Kumar H, Kumagai Y, Tsuchida T, Koenig PA, Satoh T, Guo Z, Jang MH, Saitoh T, Akira S, Kawai T. Involvement of the NLRP3 inflammasome in innate and humoral adaptive immune responses to fungal beta-glucan. J Immunol 2009; 183:8061-7; PMID:20007575; http://dx.doi.org/10.4049/jimmunol.0902477
- van de Veerdonk FL, Joosten LA, Shaw PJ, Smeekens SP, Malireddi RK, van der Meer JW, Kullberg BJ, Netea MG, Kanneganti TD. The inflammasome drives protective Th1 and Th17 cellular responses in disseminated candidiasis. Eur J Immunol 2011; 41:2260-8; PMID:21681738; http://dx.doi.org/10.1002/eji.201041226
- Hise AG, Tomalka J, Ganesan S, Patel K, Hall BA, Brown GD, Fitzgerald KA. An essential role for the NLRP3 inflammasome in host defense against the human fungal pathogen Candida albicans. Cell Host Microbe 2009; 5:487-97; PMID:19454352; http://dx.doi.org/10.1016/j.chom.2009.05.002
- Joly S, Eisenbarth SC, Olivier AK, Williams A, Kaplan DH, Cassel SL, Flavell RA, Sutterwala FS. Cutting edge: Nlrp10 is essential for protective antifungal adaptive immunity against Candida albicans. J Immunol 2012; 189:4713-7; PMID:23071280; http://dx.doi.org/10.4049/jimmunol.1201715
- Tomalka J, Ganesan S, Azodi E, Patel K, Majmudar P, Hall BA, Fitzgerald KA, Hise AG. A novel role for the NLRC4 inflammasome in mucosal defenses against the fungal pathogen Candida albicans. PLoS Pathog 2011; 7:e1002379; PMID:22174673.
- Hacker H, Vabulas RM, Takeuchi O, Hoshino K, Akira S, Wagner H. Immune cell activation by bacterial CpG-DNA through myeloid differentiation marker 88 and tumor necrosis factor receptor-associated factor (TRAF)6. J Exp Med 2000; 192:595-600; PMID:10952730; http://dx.doi.org/10.1084/jem.192.4.595
- Nagai Y, Akashi S, Nagafuku M, Ogata M, Iwakura Y, Akira S, Kitamura T, Kosugi A, Kimoto M, Miyake K. Essential role of MD-2 in LPS responsiveness and TLR4 distribution. Nat Immunol 2002; 3:667-72; PMID:12055629.
- Vogel SN, Fitzgerald KA, Fenton MJ. TLRs: differential adapter utilization by toll-like receptors mediates TLR-specific patterns of gene expression. Mol Interv 2003; 3:466-77; PMID:14993454; http://dx.doi.org/10.1124/mi.3.8.466
- Medzhitov R, Preston-Hurlburt P, Kopp E, Stadlen A, Chen C, Ghosh S, Janeway CA Jr. MyD88 is an adaptor protein in the hToll/IL-1 receptor family signaling pathways. Mol Cell 1998; 2:253-8; PMID:9734363; http://dx.doi.org/10.1016/S1097-2765(00)80136-7
- Nasu K, Narahara H. Pattern recognition via the toll-like receptor system in the human female genital tract. Mediators Inflamm 2010; 2010:976024; PMID:20396665; http://dx.doi.org/10.1155/2010/976024
- LeibundGut-Landmann S, Gross O, Robinson MJ, Osorio F, Slack EC, Tsoni SV, Schweighoffer E, Tybulewicz V, Brown GD, Ruland J, et al. Syk- and CARD9-dependent coupling of innate immunity to the induction of T helper cells that produce interleukin 17. Nat Immunol 2007; 8:630-8; PMID:17450144; http://dx.doi.org/10.1038/ni1460
- Strasser D, Neumann K, Bergmann H, Marakalala MJ, Guler R, Rojowska A, Hopfner KP, Brombacher F, Urlaub H, Baier G, et al. Syk kinase-coupled C-type lectin receptors engage protein kinase C-sigma to elicit Card9 adaptor-mediated innate immunity. Immunity 2012; 36:32-42; PMID:22265677; http://dx.doi.org/10.1016/j.immuni.2011.11.015
- Sancho D, Reis e Sousa C. Signaling by myeloid C-type lectin receptors in immunity and homeostasis. Annu Rev Immunol 2012; 30:491-529; PMID:22224766; http://dx.doi.org/10.1146/annurev-immunol-031210-101352
- Rogers NC, Slack EC, Edwards AD, Nolte MA, Schulz O, Schweighoffer E, Williams DL, Gordon S, Tybulewicz VL, Brown GD, et al. Syk-dependent cytokine induction by Dectin-1 reveals a novel pattern recognition pathway for C type lectins. Immunity 2005; 22:507-17; PMID:15845454; http://dx.doi.org/10.1016/j.immuni.2005.03.004
- Sato K, Yang XL, Yudate T, Chung JS, Wu J, Luby-Phelps K, Kimberly RP, Underhill D, Cruz PD Jr, Ariizumi K. Dectin-2 is a pattern recognition receptor for fungi that couples with the Fc receptor gamma chain to induce innate immune responses. J Biol Chem 2006; 281:38854-66; PMID:17050534; http://dx.doi.org/10.1074/jbc.M606542200
- Yamasaki S, Ishikawa E, Sakuma M, Hara H, Ogata K, Saito T. Mincle is an ITAM-coupled activating receptor that senses damaged cells. Nat Immunol 2008; 9:1179-88; PMID:18776906; http://dx.doi.org/10.1038/ni.1651
- Saijo S, Ikeda S, Yamabe K, Kakuta S, Ishigame H, Akitsu A, Fujikado N, Kusaka T, Kubo S, Chung SH, et al. Dectin-2 recognition of alpha-mannans and induction of Th17 cell differentiation is essential for host defense against Candida albicans. Immunity 2010; 32:681-91; PMID:20493731; http://dx.doi.org/10.1016/j.immuni.2010.05.001
- Hara H, Ishihara C, Takeuchi A, Imanishi T, Xue L, Morris SW, Inui M, Takai T, Shibuya A, Saijo S, et al. The adaptor protein CARD9 is essential for the activation of myeloid cells through ITAM-associated and Toll-like receptors. Nat Immunol 2007; 8:619-29; PMID:17486093; http://dx.doi.org/10.1038/ni1466
- Gross O, Gewies A, Finger K, Schafer M, Sparwasser T, Peschel C, Forster I, Ruland J. Card9 controls a non-TLR signalling pathway for innate anti-fungal immunity. Nature 2006; 442:651-6; PMID:16862125; http://dx.doi.org/10.1038/nature04926
- Gringhuis SI, den Dunnen J, Litjens M, van der Vlist M, Wevers B, Bruijns SC, Geijtenbeek TB. Dectin-1 directs T helper cell differentiation by controlling noncanonical NF-kappaB activation through Raf-1 and Syk. Nat Immunol 2009; 10:203-13; PMID:19122653; http://dx.doi.org/10.1038/ni.1692
- Gringhuis SI, den Dunnen J, Litjens M, van Het Hof B, van Kooyk Y, Geijtenbeek TB. C-type lectin DC-SIGN modulates Toll-like receptor signaling via Raf-1 kinase-dependent acetylation of transcription factor NF-kappaB. Immunity 2007; 26:605-16; PMID:17462920; http://dx.doi.org/10.1016/j.immuni.2007.03.012
- Yamamoto M, Takeda K. Current views of toll-like receptor signaling pathways. Gastroenterol Res Pract 2010; 2010:240365; PMID:21197425; http://dx.doi.org/10.1155/2010/240365
- Hardison SE, Brown GD. C-type lectin receptors orchestrate antifungal immunity. Nat Immunol 2012; 13:817-22; PMID:22910394; http://dx.doi.org/10.1038/ni.2369
- Wells JM, Rossi O, Meijerink M, van Baarlen P. Epithelial crosstalk at the microbiota-mucosal interface. Proc Natl Acad Sci U S A 2011; 108 Suppl 1:4607-14; PMID:20826446; http://dx.doi.org/10.1073/pnas.1000092107
- Pioli PA, Amiel E, Schaefer TM, Connolly JE, Wira CR, Guyre PM. Differential expression of Toll-like receptors 2 and 4 in tissues of the human female reproductive tract. Infect Immun 2004; 72:5799-806; PMID:15385480; http://dx.doi.org/10.1128/IAI.72.10.5799-5806.2004
- Ghosh M, Schaefer TM, Fahey JV, Wright JA, Wira CR. Antiviral responses of human Fallopian tube epithelial cells to toll-like receptor 3 agonist poly(I:C). Fertil Steril 2008; 89:1497-506; PMID:17669408; http://dx.doi.org/10.1016/j.fertnstert.2007.05.023
- Herbst-Kralovetz MM, Quayle AJ, Ficarra M, Greene S, Rose WA, 2nd, Chesson R, Spagnuolo RA, Pyles RB. Quantification and comparison of toll-like receptor expression and responsiveness in primary and immortalized human female lower genital tract epithelia. Am J Reprod Immunol 2008; 59:212-24; PMID:18201283; http://dx.doi.org/10.1111/j.1600-0897.2007.00566.x
- Hirata T, Osuga Y, Hamasaki K, Hirota Y, Nose E, Morimoto C, Harada M, Takemura Y, Koga K, Yoshino O, et al. Expression of toll-like receptors 2, 3, 4, and 9 genes in the human endometrium during the menstrual cycle. J Reprod Immunol 2007; 74:53-60; PMID:17292969; http://dx.doi.org/10.1016/j.jri.2006.11.004
- Fichorova RN, Cronin AO, Lien E, Anderson DJ, Ingalls RR. Response to Neisseria gonorrhoeae by cervicovaginal epithelial cells occurs in the absence of toll-like receptor 4-mediated signaling. J Immunol 2002; 168:2424-32; PMID:11859134; http://dx.doi.org/10.4049/jimmunol.168.5.2424
- Fazeli A, Bruce C, Anumba DO. Characterization of Toll-like receptors in the female reproductive tract in humans. Hum Reprod 2005; 20:1372-8; PMID:15695310; http://dx.doi.org/10.1093/humrep/deh775
- Furrie E, Macfarlane S, Thomson G, Macfarlane GT. Toll-like receptors-2, -3 and -4 expression patterns on human colon and their regulation by mucosal-associated bacteria. Immunology 2005; 115:565-74; PMID:16011525; http://dx.doi.org/10.1111/j.1365-2567.2005.02200.x
- Cario E, Rosenberg IM, Brandwein SL, Beck PL, Reinecker HC, Podolsky DK. Lipopolysaccharide activates distinct signaling pathways in intestinal epithelial cell lines expressing Toll-like receptors. J Immunol 2000; 164:966-72; PMID:10623846; http://dx.doi.org/10.4049/jimmunol.164.2.966
- Abreu MT, Vora P, Faure E, Thomas LS, Arnold ET, Arditi M. Decreased expression of Toll-like receptor-4 and MD-2 correlates with intestinal epithelial cell protection against dysregulated proinflammatory gene expression in response to bacterial lipopolysaccharide. J Immunol 2001; 167:1609-16; PMID:11466383; http://dx.doi.org/10.4049/jimmunol.167.3.1609
- Otte JM, Podolsky DK. Functional modulation of enterocytes by gram-positive and gram-negative microorganisms. Am J Physiol Gastrointest Liver Physiol 2004; 286:G613-26; PMID:15010363; http://dx.doi.org/10.1152/ajpgi.00341.2003
- Rhee SH, Im E, Riegler M, Kokkotou E, O'Brien M, Pothoulakis C. Pathophysiological role of Toll-like receptor 5 engagement by bacterial flagellin in colonic inflammation. Proc Natl Acad Sci U S A 2005; 102:13610-5; PMID:16157881; http://dx.doi.org/10.1073/pnas.0502174102
- Shaykhiev R, Behr J, Bals R. Microbial patterns signaling via Toll-like receptors 2 and 5 contribute to epithelial repair, growth and survival. PLoS One 2008; 3:e1393; PMID:18167552; http://dx.doi.org/10.1371/journal.pone.0001393
- Williams DW, Jordan RP, Wei XQ, Alves CT, Wise MP, Wilson MJ, Lewis MA. Interactions of Candida albicans with host epithelial surfaces. J Oral Microbiol 2013; 5:22434; PMID:24155995; http://dx.doi.org/10.3402/jom.v5i0.22434
- Rouabhia M, Mukherjee PK, Lattif AA, Curt S, Chandra J, Ghannoum MA. Disruption of sphingolipid biosynthetic gene IPT1 reduces Candida albicans adhesion and prevents activation of human gingival epithelial cell innate immune defense. Med Mycol 2011; 49:458-66; PMID:21091155.
- Weindl G, Naglik JR, Kaesler S, Biedermann T, Hube B, Korting HC, Schaller M. Human epithelial cells establish direct antifungal defense through TLR4-mediated signaling. J Clin Invest 2007; 117:3664-72; PMID:17992260.
- Goodridge HS, Simmons RM, Underhill DM. Dectin-1 stimulation by Candida albicans yeast or zymosan triggers NFAT activation in macrophages and dendritic cells. J Immunol 2007; 178:3107-15; PMID:17312158; http://dx.doi.org/10.4049/jimmunol.178.5.3107
- Weindl G, Wagener J, Schaller M. Epithelial cells and innate antifungal defense. J Dent Res 2010; 89:666-75; PMID:20395411; http://dx.doi.org/10.1177/0022034510368784
- Li C, Zhao GQ, Che CY, Li N, Lin J, Xu Q, Wang Q, Liu Y, Qiu S. Expression of dectin-1 during fungus infection in human corneal epithelial cells. Int J Ophthalmol 2014; 7:34-7; PMID:24634860.
- Jie Z, Wu XY, Yu FS. Activation of Toll-like receptors 2 and 4 in Aspergillus fumigatus keratitis. Innate Immun 2009; 15:155-68; PMID:19474209; http://dx.doi.org/10.1177/1753425908101521
- Aratani Y, Koyama H, Nyui S, Suzuki K, Kura F, Maeda N. Severe impairment in early host defense against Candida albicans in mice deficient in myeloperoxidase. Infect Immun 1999; 67:1828-36; PMID:10085024.
- Li M, Chen Q, Tang R, Shen Y, Liu WD. The expression of beta-defensin-2, 3 and LL-37 induced by Candida albicans phospholipomannan in human keratinocytes. J Dermatol Sci 2011; 61:72-5; PMID:21146963; http://dx.doi.org/10.1016/j.jdermsci.2010.11.009
- Swidergall M, Ernst JF. Interplay between Candida albicans and the antimicrobial peptide armory. Eukaryot Cell 2014; 13:950-7; PMID:24951441; http://dx.doi.org/10.1128/EC.00093-14
- Naglik JR, Moyes D. Epithelial cell innate response to Candida albicans. Adv Dent Res 2011; 23:50-5; PMID:21441481; http://dx.doi.org/10.1177/0022034511399285
- Schaller M, Boeld U, Oberbauer S, Hamm G, Hube B, Korting HC. Polymorphonuclear leukocytes (PMNs) induce protective Th1-type cytokine epithelial responses in an in vitro model of oral candidosis. Microbiology 2004; 150:2807-13; PMID:15347740; http://dx.doi.org/10.1099/mic.0.27169-0
- Villar CC, Kashleva H, Mitchell AP, Dongari-Bagtzoglou A. Invasive phenotype of Candida albicans affects the host proinflammatory response to infection. Infect Immun 2005; 73:4588-95; PMID:16040970; http://dx.doi.org/10.1128/IAI.73.8.4588-4595.2005
- Dongari-Bagtzoglou A, Fidel PL Jr. The host cytokine responses and protective immunity in oropharyngeal candidiasis. J Dent Res 2005; 84:966-77; PMID:16246925; http://dx.doi.org/10.1177/154405910508401101
- Eyerich S, Wagener J, Wenzel V, Scarponi C, Pennino D, Albanesi C, Schaller M, Behrendt H, Ring J, Schmidt-Weber CB, et al. IL-22 and TNF-alpha represent a key cytokine combination for epidermal integrity during infection with Candida albicans. Eur J Immunol 2011; 41:1894-901; PMID:21469124; http://dx.doi.org/10.1002/eji.201041197
- Yano J, Lilly E, Barousse M, Fidel PL Jr. Epithelial cell-derived S100 calcium-binding proteins as key mediators in the hallmark acute neutrophil response during Candida vaginitis. Infect Immun 2010; 78:5126-37; PMID:20823201; http://dx.doi.org/10.1128/IAI.00388-10
- Yano J, Palmer GE, Eberle KE, Peters BM, Vogl T, McKenzie AN, Fidel PL Jr. Vaginal epithelial cell-derived S100 alarmins induced by Candida albicans via pattern recognition receptor interactions are sufficient but not necessary for the acute neutrophil response during experimental vaginal candidiasis. Infect Immun 2014; 82:783-92; PMID:24478092; http://dx.doi.org/10.1128/IAI.00861-13
- Grubb SE, Murdoch C, Sudbery PE, Saville SP, Lopez-Ribot JL, Thornhill MH. Candida albicans-endothelial cell interactions: a key step in the pathogenesis of systemic candidiasis. Infect Immun 2008; 76:4370-7; PMID:18573891; http://dx.doi.org/10.1128/IAI.00332-08
- Faure E, Equils O, Sieling PA, Thomas L, Zhang FX, Kirschning CJ, Polentarutti N, Muzio M, Arditi M. Bacterial lipopolysaccharide activates NF-kappaB through toll-like receptor 4 (TLR-4) in cultured human dermal endothelial cells. Differential expression of TLR-4 and TLR-2 in endothelial cells. J Biol Chem 2000; 275:11058-63; PMID:10753909; http://dx.doi.org/10.1074/jbc.275.15.11058
- Faure E, Thomas L, Xu H, Medvedev A, Equils O, Arditi M. Bacterial lipopolysaccharide and IFN-gamma induce Toll-like receptor 2 and Toll-like receptor 4 expression in human endothelial cells: role of NF-kappa B activation. J Immunol 2001; 166:2018-24; PMID:11160251; http://dx.doi.org/10.4049/jimmunol.166.3.2018
- Tissari J, Siren J, Meri S, Julkunen I, Matikainen S. IFN-alpha enhances TLR3-mediated antiviral cytokine expression in human endothelial and epithelial cells by up-regulating TLR3 expression. J Immunol 2005; 174:4289-94; PMID:15778392; http://dx.doi.org/10.4049/jimmunol.174.7.4289
- El Kebir D, Jozsef L, Pan W, Wang L, Filep JG. Bacterial DNA activates endothelial cells and promotes neutrophil adherence through TLR9 signaling. J Immunol 2009; 182:4386-94; PMID:19299739; http://dx.doi.org/10.4049/jimmunol.0803044
- Li J, Ma Z, Tang ZL, Stevens T, Pitt B, Li S. CpG DNA-mediated immune response in pulmonary endothelial cells. Am J Physiol Lung Cell Mol Physiol 2004; 287:L552-8; PMID:15155271; http://dx.doi.org/10.1152/ajplung.00436.2003
- Groger M, Holnthoner W, Maurer D, Lechleitner S, Wolff K, Mayr BB, Lubitz W, Petzelbauer P. Dermal microvascular endothelial cells express the 180-kDa macrophage mannose receptor in situ and in vitro. J Immunol 2000; 165:5428-34; PMID:11067894; http://dx.doi.org/10.4049/jimmunol.165.10.5428
- Thijssen VL, Poirier F, Baum LG, Griffioen AW. Galectins in the tumor endothelium: opportunities for combined cancer therapy. Blood 2007; 110:2819-27; PMID:17591944; http://dx.doi.org/10.1182/blood-2007-03-077792
- Muller V, Viemann D, Schmidt M, Endres N, Ludwig S, Leverkus M, Roth J, Goebeler M. Candida albicans triggers activation of distinct signaling pathways to establish a proinflammatory gene expression program in primary human endothelial cells. J Immunol 2007; 179:8435-45; PMID:18056390; http://dx.doi.org/10.4049/jimmunol.179.12.8435
- Zhang PP, Xu XY, Zhang F, Shi Y. [The changes of Dectin-1 and TLR2 in human umbilical vein endothelial cells stimulated by Aspergillus fumigatus]. Zhonghua Jie He He Hu Xi Za Zhi 2011; 34:91-4; PMID:21426724.
- Sun JN, Solis NV, Phan QT, Bajwa JS, Kashleva H, Thompson A, Liu Y, Dongari-Bagtzoglou A, Edgerton M, Filler SG. Host cell invasion and virulence mediated by Candida albicans Ssa1. PLoS Pathog 2010; 6:e1001181; PMID:21085601; http://dx.doi.org/10.1371/journal.ppat.1001181
- Phan QT, Myers CL, Fu Y, Sheppard DC, Yeaman MR, Welch WH, Ibrahim AS, Edwards JE Jr, Filler SG. Als3 is a Candida albicans invasin that binds to cadherins and induces endocytosis by host cells. PLoS Biol 2007; 5:e64; PMID:17311474; http://dx.doi.org/10.1371/journal.pbio.0050064
- Moreno-Ruiz E, Galan-Diez M, Zhu W, Fernandez-Ruiz E, d'Enfert C, Filler SG, Cossart P, Veiga E. Candida albicans internalization by host cells is mediated by a clathrin-dependent mechanism. Cell Microbiol 2009; 11:1179-89; PMID:19416270; http://dx.doi.org/10.1111/j.1462-5822.2009.01319.x
- Vonk AG, Netea MG, van der Meer JW, Kullberg BJ. Host defence against disseminated Candida albicans infection and implications for antifungal immunotherapy. Expert Opin Biol Ther 2006; 6:891-903; PMID:16918256; http://dx.doi.org/10.1517/14712598.6.9.891
- Lionakis MS, Lim JK, Lee CC, Murphy PM. Organ-specific innate immune responses in a mouse model of invasive candidiasis. J Innate Immun 2011; 3:180-99; PMID:21063074; http://dx.doi.org/10.1159/000321157
- Lionakis MS, Fischer BG, Lim JK, Swamydas M, Wan W, Richard Lee CC, Cohen JI, Scheinberg P, Gao JL, Murphy PM. Chemokine receptor Ccr1 drives neutrophil-mediated kidney immunopathology and mortality in invasive candidiasis. PLoS Pathog 2012; 8:e1002865; PMID:22916017; http://dx.doi.org/10.1371/journal.ppat.1002865
- Majer O, Bourgeois C, Zwolanek F, Lassnig C, Kerjaschki D, Mack M, Muller M, Kuchler K. Type I interferons promote fatal immunopathology by regulating inflammatory monocytes and neutrophils during Candida infections. PLoS Pathog 2012; 8:e1002811; PMID:22911155; http://dx.doi.org/10.1371/journal.ppat.1002811
- Wuthrich M, Ersland K, Sullivan T, Galles K, Klein BS. Fungi subvert vaccine T cell priming at the respiratory mucosa by preventing chemokine-induced influx of inflammatory monocytes. Immunity 2012; 36:680-92; PMID:22483803; http://dx.doi.org/10.1016/j.immuni.2012.02.015
- Tada H, Nemoto E, Shimauchi H, Watanabe T, Mikami T, Matsumoto T, Ohno N, Tamura H, Shibata K, Akashi S, et al. Saccharomyces cerevisiae- and Candida albicans-derived mannan induced production of tumor necrosis factor alpha by human monocytes in a CD14- and Toll-like receptor 4-dependent manner. Microbiol Immunol 2002; 46:503-12; PMID:12222939; http://dx.doi.org/10.1111/j.1348-0421.2002.tb02727.x
- Yamasaki S, Matsumoto M, Takeuchi O, Matsuzawa T, Ishikawa E, Sakuma M, Tateno H, Uno J, Hirabayashi J, Mikami Y, et al. C-type lectin Mincle is an activating receptor for pathogenic fungus, Malassezia. Proc Natl Acad Sci U S A 2009; 106:1897-902; PMID:19171887; http://dx.doi.org/10.1073/pnas.0805177106
- Jouault T, El Abed-El Behi M, Martinez-Esparza M, Breuilh L, Trinel PA, Chamaillard M, Trottein F, Poulain D. Specific recognition of Candida albicans by macrophages requires galectin-3 to discriminate Saccharomyces cerevisiae and needs association with TLR2 for signaling. J Immunol 2006; 177:4679-87; PMID:16982907; http://dx.doi.org/10.4049/jimmunol.177.7.4679
- Linden JR, Kunkel D, Laforce-Nesbitt SS, Bliss JM. The role of galectin-3 in phagocytosis of Candida albicans and Candida parapsilosis by human neutrophils. Cell Microbiol 2013; 15:1127-42; PMID:23279221; http://dx.doi.org/10.1111/cmi.12103
- Ariizumi K, Shen GL, Shikano S, Ritter R, 3rd, Zukas P, Edelbaum D, Morita A, Takashima A. Cloning of a second dendritic cell-associated C-type lectin (dectin-2) and its alternatively spliced isoforms. J Biol Chem 2000; 275:11957-63; PMID:10766825; http://dx.doi.org/10.1074/jbc.275.16.11957
- Taylor PR, Reid DM, Heinsbroek SE, Brown GD, Gordon S, Wong SY. Dectin-2 is predominantly myeloid restricted and exhibits unique activation-dependent expression on maturing inflammatory monocytes elicited in vivo. Eur J Immunol 2005; 35:2163-74; PMID:15940672; http://dx.doi.org/10.1002/eji.200425785
- McGreal EP, Rosas M, Brown GD, Zamze S, Wong SY, Gordon S, Martinez-Pomares L, Taylor PR. The carbohydrate-recognition domain of Dectin-2 is a C-type lectin with specificity for high mannose. Glycobiology 2006; 16:422-30; PMID:16423983; http://dx.doi.org/10.1093/glycob/cwj077
- Arce I, Martinez-Munoz L, Roda-Navarro P, Fernandez-Ruiz E. The human C-type lectin CLECSF8 is a novel monocyte/macrophage endocytic receptor. Eur J Immunol 2004; 34:210-20; PMID:14971047; http://dx.doi.org/10.1002/eji.200324230
- Wells CA, Salvage-Jones JA, Li X, Hitchens K, Butcher S, Murray RZ, Beckhouse AG, Lo YL, Manzanero S, Cobbold C, et al. The macrophage-inducible C-type lectin, mincle, is an essential component of the innate immune response to Candida albicans. J Immunol 2008; 180:7404-13; PMID:18490740; http://dx.doi.org/10.4049/jimmunol.180.11.7404
- Vijayan D, Radford KJ, Beckhouse AG, Ashman RB, Wells CA. Mincle polarizes human monocyte and neutrophil responses to Candida albicans. Immunol Cell Biol 2012; 90:889-95; PMID:22641025; http://dx.doi.org/10.1038/icb.2012.24
- Forsyth CB, Plow EF, Zhang L. Interaction of the fungal pathogen Candida albicans with integrin CD11b/CD18: recognition by the I domain is modulated by the lectin-like domain and the CD18 subunit. J Immunol 1998; 161:6198-205; PMID:9834106.
- van Bruggen R, Drewniak A, Jansen M, van Houdt M, Roos D, Chapel H, Verhoeven AJ, Kuijpers TW. Complement receptor 3, not Dectin-1, is the major receptor on human neutrophils for beta-glucan-bearing particles. Mol Immunol 2009; 47:575-81; PMID:19811837; http://dx.doi.org/10.1016/j.molimm.2009.09.018
- Gantner BN, Simmons RM, Underhill DM. Dectin-1 mediates macrophage recognition of Candida albicans yeast but not filaments. EMBO J 2005; 24:1277-86; PMID:15729357; http://dx.doi.org/10.1038/sj.emboj.7600594
- Brown GD, Gordon S. Fungal beta-glucans and mammalian immunity. Immunity 2003; 19:311-5; PMID:14499107; http://dx.doi.org/10.1016/S1074-7613(03)00233-4
- McDonald JU, Rosas M, Brown GD, Jones SA, Taylor PR. Differential dependencies of monocytes and neutrophils on dectin-1, dectin-2 and complement for the recognition of fungal particles in inflammation. PLoS One 2012; 7:e45781; PMID:23049859; http://dx.doi.org/10.1371/journal.pone.0045781
- Means TK, Mylonakis E, Tampakakis E, Colvin RA, Seung E, Puckett L, Tai MF, Stewart CR, Pukkila-Worley R, Hickman SE, et al. Evolutionarily conserved recognition and innate immunity to fungal pathogens by the scavenger receptors SCARF1 and CD36. J Exp Med 2009; 206:637-53; PMID:19237602; http://dx.doi.org/10.1084/jem.20082109
- Wagener J, Malireddi RK, Lenardon MD, Koberle M, Vautier S, MacCallum DM, Biedermann T, Schaller M, Netea MG, Kanneganti TD, et al. Fungal chitin dampens inflammation through IL-10 induction mediated by NOD2 and TLR9 activation. PLoS Pathog 2014; 10:e1004050; PMID:24722226; http://dx.doi.org/10.1371/journal.ppat.1004050
- Herre J, Marshall AS, Caron E, Edwards AD, Williams DL, Schweighoffer E, Tybulewicz V, Reis e Sousa C, Gordon S, Brown GD. Dectin-1 uses novel mechanisms for yeast phagocytosis in macrophages. Blood 2004; 104:4038-45; PMID:15304394; http://dx.doi.org/10.1182/blood-2004-03-1140
- Le Cabec V, Emorine LJ, Toesca I, Cougoule C, Maridonneau-Parini I. The human macrophage mannose receptor is not a professional phagocytic receptor. J Leukoc Biol 2005; 77:934-43; PMID:15767290; http://dx.doi.org/10.1189/jlb.1204705
- Mansour MK, Tam JM, Khan NS, Seward M, Davids PJ, Puranam S, Sokolovska A, Sykes DB, Dagher Z, Becker C, et al. Dectin-1 activation controls maturation of beta-1,3-glucan-containing phagosomes. J Biol Chem 2013; 288:16043-54; PMID:23609446; http://dx.doi.org/10.1074/jbc.M113.473223
- Kasperkovitz PV, Khan NS, Tam JM, Mansour MK, Davids PJ, Vyas JM. Toll-like receptor 9 modulates macrophage antifungal effector function during innate recognition of Candida albicans and Saccharomyces cerevisiae. Infect Immun 2011; 79:4858-67; PMID:21947771; http://dx.doi.org/10.1128/IAI.05626-11
- Franchi L, Munoz-Planillo R, Nunez G. Sensing and reacting to microbes through the inflammasomes. Nat Immunol 2012; 13:325-32; PMID:22430785; http://dx.doi.org/10.1038/ni.2231
- Netea MG, Vonk AG, van den Hoven M, Verschueren I, Joosten LA, van Krieken JH, van den Berg WB, Van der Meer JW, Kullberg BJ. Differential role of IL-18 and IL-12 in the host defense against disseminated Candida albicans infection. Eur J Immunol 2003; 33:3409-17; PMID:14635050; http://dx.doi.org/10.1002/eji.200323737
- Pietrella D, Pandey N, Gabrielli E, Pericolini E, Perito S, Kasper L, Bistoni F, Cassone A, Hube B, Vecchiarelli A. Secreted aspartic proteases of Candida albicans activate the NLRP3 inflammasome. Eur J Immunol 2013; 43:679-92; PMID:23280543; http://dx.doi.org/10.1002/eji.201242691
- Wellington M, Koselny K, Sutterwala FS, Krysan DJ. Candida albicans triggers NLRP3-mediated pyroptosis in macrophages. Eukaryot Cell 2014; 13:329-40; PMID:24376002; http://dx.doi.org/10.1128/EC.00336-13
- Kawai T, Akira S. Toll-like receptors and their crosstalk with other innate receptors in infection and immunity. Immunity 2011; 34:637-50; PMID:21616434; http://dx.doi.org/10.1016/j.immuni.2011.05.006
- Dennehy KM, Ferwerda G, Faro-Trindade I, Pyz E, Willment JA, Taylor PR, Kerrigan A, Tsoni SV, Gordon S, Meyer-Wentrup F, et al. Syk kinase is required for collaborative cytokine production induced through Dectin-1 and Toll-like receptors. Eur J Immunol 2008; 38:500-6; PMID:18200499; http://dx.doi.org/10.1002/eji.200737741
- Gerosa F, Baldani-Guerra B, Lyakh LA, Batoni G, Esin S, Winkler-Pickett RT, Consolaro MR, De Marchi M, Giachino D, Robbiano A, et al. Differential regulation of interleukin 12 and interleukin 23 production in human dendritic cells. J Exp Med 2008; 205:1447-61; PMID:18490488; http://dx.doi.org/10.1084/jem.20071450
- van de Veerdonk FL, Marijnissen RJ, Kullberg BJ, Koenen HJ, Cheng SC, Joosten I, van den Berg WB, Williams DL, van der Meer JW, Joosten LA, et al. The macrophage mannose receptor induces IL-17 in response to Candida albicans. Cell Host Microbe 2009; 5:329-40; PMID:19380112; http://dx.doi.org/10.1016/j.chom.2009.02.006
- Esteban A, Popp MW, Vyas VK, Strijbis K, Ploegh HL, Fink GR. Fungal recognition is mediated by the association of dectin-1 and galectin-3 in macrophages. Proc Natl Acad Sci U S A 2011; 108:14270-5; PMID:21825168; http://dx.doi.org/10.1073/pnas.1111415108
- Zhao W, Wang L, Zhang M, Wang P, Zhang L, Yuan C, Qi J, Qiao Y, Kuo PC, Gao C. Peroxisome proliferator-activated receptor gamma negatively regulates IFN-beta production in Toll-like receptor (TLR) 3- and TLR4-stimulated macrophages by preventing interferon regulatory factor 3 binding to the IFN-beta promoter. J Biol Chem 2011; 286:5519-28; PMID:21148557; http://dx.doi.org/10.1074/jbc.M110.149823
- Kawana H, Karaki H, Higashi M, Miyazaki M, Hilberg F, Kitagawa M, Harigaya K. CD44 suppresses TLR-mediated inflammation. J Immunol 2008; 180:4235-45; PMID:18322236; http://dx.doi.org/10.4049/jimmunol.180.6.4235
- Underhill DM, Rossnagle E, Lowell CA, Simmons RM. Dectin-1 activates Syk tyrosine kinase in a dynamic subset of macrophages for reactive oxygen production. Blood 2005; 106:2543-50; PMID:15956283; http://dx.doi.org/10.1182/blood-2005-03-1239
- Vazquez-Torres A, Balish E. Macrophages in resistance to candidiasis. Microbiol Mol Biol Rev 1997; 61:170-92; PMID:9184009.
- Drummond RA, Saijo S, Iwakura Y, Brown GD. The role of Syk/CARD9 coupled C-type lectins in antifungal immunity. Eur J Immunol 2011; 41:276-81; PMID:21267996; http://dx.doi.org/10.1002/eji.201041252
- Suram S, Gangelhoff TA, Taylor PR, Rosas M, Brown GD, Bonventre JV, Akira S, Uematsu S, Williams DL, Murphy RC, et al. Pathways regulating cytosolic phospholipase A2 activation and eicosanoid production in macrophages by Candida albicans. J Biol Chem 2010; 285:30676-85; PMID:20643646; http://dx.doi.org/10.1074/jbc.M110.143800
- Robinson MJ, Osorio F, Rosas M, Freitas RP, Schweighoffer E, Gross O, Verbeek JS, Ruland J, Tybulewicz V, Brown GD, et al. Dectin-2 is a Syk-coupled pattern recognition receptor crucial for Th17 responses to fungal infection. J Exp Med 2009; 206:2037-51; PMID:19703985; http://dx.doi.org/10.1084/jem.20082818
- Barrett NA, Maekawa A, Rahman OM, Austen KF, Kanaoka Y. Dectin-2 recognition of house dust mite triggers cysteinyl leukotriene generation by dendritic cells. J Immunol 2009; 182:1119-28; PMID:19124755; http://dx.doi.org/10.4049/jimmunol.182.2.1119
- Reizis B, Bunin A, Ghosh HS, Lewis KL, Sisirak V. Plasmacytoid dendritic cells: recent progress and open questions. Annu Rev Immunol 2011; 29:163-83; PMID:21219184; http://dx.doi.org/10.1146/annurev-immunol-031210-101345
- Pietrella D, Bistoni G, Corbucci C, Perito S, Vecchiarelli A. Candida albicans mannoprotein influences the biological function of dendritic cells. Cell Microbiol 2006; 8:602-12; PMID:16548886; http://dx.doi.org/10.1111/j.1462-5822.2005.00651.x
- Netea MG, van de Veerdonk F, Verschueren I, van der Meer JW, Kullberg BJ. Role of TLR1 and TLR6 in the host defense against disseminated candidiasis. FEMS Immunol Med Microbiol 2008; 52:118-23; PMID:18036178; http://dx.doi.org/10.1111/j.1574-695X.2007.00353.x
- Bourgeois C, Majer O, Frohner IE, Lesiak-Markowicz I, Hildering KS, Glaser W, Stockinger S, Decker T, Akira S, Muller M, et al. Conventional dendritic cells mount a type I IFN response against Candida spp. requiring novel phagosomal TLR7-mediated IFN-beta signaling. J Immunol 2011; 186:3104-12; PMID:21282509; http://dx.doi.org/10.4049/jimmunol.1002599
- Ariizumi K, Shen GL, Shikano S, Xu S, Ritter R 3rd, Kumamoto T, Edelbaum D, Morita A, Bergstresser PR, Takashima A. Identification of a novel, dendritic cell-associated molecule, dectin-1, by subtractive cDNA cloning. J Biol Chem 2000; 275:20157-67; PMID:10779524; http://dx.doi.org/10.1074/jbc.M909512199
- del Fresno C, Soulat D, Roth S, Blazek K, Udalova I, Sancho D, Ruland J, Ardavin C. Interferon-beta production via Dectin-1-Syk-IRF5 signaling in dendritic cells is crucial for immunity to C. albicans. Immunity 2013; 38:1176-86; PMID:23770228; http://dx.doi.org/10.1016/j.immuni.2013.05.010
- Cambi A, Gijzen K, de Vries l J, Torensma R, Joosten B, Adema GJ, Netea MG, Kullberg BJ, Romani L, Figdor CG. The C-type lectin DC-SIGN (CD209) is an antigen-uptake receptor for Candida albicans on dendritic cells. Eur J Immunol 2003; 33:532-8; PMID:12645952; http://dx.doi.org/10.1002/immu.200310029
- Cambi A, Netea MG, Mora-Montes HM, Gow NA, Hato SV, Lowman DW, Kullberg BJ, Torensma R, Williams DL, Figdor CG. Dendritic cell interaction with Candida albicans critically depends on N-linked mannan. J Biol Chem 2008; 283:20590-9; PMID:18482990; http://dx.doi.org/10.1074/jbc.M709334200
- Fermin Lee A, Chen HY, Wan L, Wu SY, Yu JS, Huang AC, Miaw SC, Hsu DK, Wu-Hsieh BA, Liu FT. Galectin-3 modulates Th17 responses by regulating dendritic cell cytokines. Am J Pathol 2013; 183:1209-22; PMID:23916470; http://dx.doi.org/10.1016/j.ajpath.2013.06.017
- Donini M, Zenaro E, Tamassia N, Dusi S. NADPH oxidase of human dendritic cells: role in Candida albicans killing and regulation by interferons, dectin-1 and CD206. Eur J Immunol 2007; 37:1194-203; PMID:17407098; http://dx.doi.org/10.1002/eji.200636532
- Neumann AK, Jacobson K. A novel pseudopodial component of the dendritic cell anti-fungal response: the fungipod. PLoS Pathog 2010; 6:e1000760; PMID:20169183; http://dx.doi.org/10.1371/journal.ppat.1000760
- Whitney PG, Bar E, Osorio F, Rogers NC, Schraml BU, Deddouche S, LeibundGut-Landmann S, Reis e Sousa C. Syk signaling in dendritic cells orchestrates innate resistance to systemic fungal infection. PLoS Pathog 2014; 10:e1004276; PMID:25033445; http://dx.doi.org/10.1371/journal.ppat.1004276
- Hohl TM, Rivera A, Lipuma L, Gallegos A, Shi C, Mack M, Pamer EG. Inflammatory monocytes facilitate adaptive CD4 T cell responses during respiratory fungal infection. Cell Host Microbe 2009; 6:470-81; PMID:19917501; http://dx.doi.org/10.1016/j.chom.2009.10.007
- Fidan I, Kalkanci A, Yesilyurt E, Erdal B. In vitro effects of Candida albicans and Aspergillus fumigatus on dendritic cells and the role of beta glucan in this effect. Adv Clin Exp Med 2014; 23:17-24; PMID: 24595999; http://dx.doi.org/10.17219/acem/37016
- Steinman RM. Decisions about dendritic cells: past, present, and future. Annu Rev Immunol 2012; 30:1-22; PMID: 22136168; http://dx.doi.org/10.1146/annurev-immunol-100311-102839
- Ramirez-Ortiz ZG, Means TK. The role of dendritic cells in the innate recognition of pathogenic fungi (A. fumigatus, C. neoformans and C. albicans). Virulence 2012; 3:635-46; PMID:23076328; http://dx.doi.org/10.4161/viru.22295
- Beno DW, Stover AG, Mathews HL. Growth inhibition of Candida albicans hyphae by CD8+ lymphocytes. J Immunol 1995; 154:5273-81; PMID:7730631.
- Manel N, Unutmaz D, Littman DR. The differentiation of human T(H)-17 cells requires transforming growth factor-beta and induction of the nuclear receptor RORgammat. Nat Immunol 2008; 9:641-9; PMID:18454151; http://dx.doi.org/10.1038/ni.1610
- Levitz SM. Th17 cells bounce off the fungal wall. Cell Host Microbe 2009; 5:311-3; PMID:19380108; http://dx.doi.org/10.1016/j.chom.2009.04.004
- Acosta-Rodriguez EV, Rivino L, Geginat J, Jarrossay D, Gattorno M, Lanzavecchia A, Sallusto F, Napolitani G. Surface phenotype and antigenic specificity of human interleukin 17-producing T helper memory cells. Nat Immunol 2007; 8:639-46; PMID:17486092; http://dx.doi.org/10.1038/ni1467
- Conti HR, Shen F, Nayyar N, Stocum E, Sun JN, Lindemann MJ, Ho AW, Hai JH, Yu JJ, Jung JW, et al. Th17 cells and IL-17 receptor signaling are essential for mucosal host defense against oral candidiasis. J Exp Med 2009; 206:299-311; PMID:19204111; http://dx.doi.org/10.1084/jem.20081463
- Kagami S, Rizzo HL, Kurtz SE, Miller LS, Blauvelt A. IL-23 and IL-17A, but not IL-12 and IL-22, are required for optimal skin host defense against Candida albicans. J Immunol 2010; 185:5453-62; PMID:20921529; http://dx.doi.org/10.4049/jimmunol.1001153
- Hernandez-Santos N, Huppler AR, Peterson AC, Khader SA, McKenna KC, Gaffen SL. Th17 cells confer long-term adaptive immunity to oral mucosal Candida albicans infections. Mucosal Immunol 2013; 6:900-10; PMID:23250275; http://dx.doi.org/10.1038/mi.2012.128
- Minegishi Y, Saito M, Tsuchiya S, Tsuge I, Takada H, Hara T, Kawamura N, Ariga T, Pasic S, Stojkovic O, et al. Dominant-negative mutations in the DNA-binding domain of STAT3 cause hyper-IgE syndrome. Nature 2007; 448:1058-62; PMID:17676033; http://dx.doi.org/10.1038/nature06096
- Holland SM, DeLeo FR, Elloumi HZ, Hsu AP, Uzel G, Brodsky N, Freeman AF, Demidowich A, Davis J, Turner ML, et al. STAT3 mutations in the hyper-IgE syndrome. N Engl J Med 2007; 357:1608-19; PMID:17881745; http://dx.doi.org/10.1056/NEJMoa073687
- Liu L, Okada S, Kong XF, Kreins AY, Cypowyj S, Abhyankar A, Toubiana J, Itan Y, Audry M, Nitschke P, et al. Gain-of-function human STAT1 mutations impair IL-17 immunity and underlie chronic mucocutaneous candidiasis. J Exp Med 2011; 208:1635-48; PMID:21727188; http://dx.doi.org/10.1084/jem.20110958
- Glocker EO, Hennigs A, Nabavi M, Schaffer AA, Woellner C, Salzer U, Pfeifer D, Veelken H, Warnatz K, Tahami F, et al. A homozygous CARD9 mutation in a family with susceptibility to fungal infections. N Engl J Med 2009; 361:1727-35; PMID: 19864672; http://dx.doi.org/10.1056/NEJMoa0810719
- Drewniak A, Gazendam RP, Tool AT, van Houdt M, Jansen MH, van Hamme JL, van Leeuwen EM, Roos D, Scalais E, de Beaufort C, et al. Invasive fungal infection and impaired neutrophil killing in human CARD9 deficiency. Blood 2013; 121:2385-92; PMID:23335372; http://dx.doi.org/10.1182/blood-2012-08-450551
- Gow NA, Hube B. Importance of the Candida albicans cell wall during commensalism and infection. Curr Opin Microbiol 2012; 15:406-12; PMID:22609181; http://dx.doi.org/10.1016/j.mib.2012.04.005
- Scherwitz C. Ultrastructure of human cutaneous candidosis. J Invest Dermatol 1982; 78:200-5; PMID:7035576; http://dx.doi.org/10.1111/1523-1747.ep12506451
- Ray TL, Payne CD. Scanning electron microscopy of epidermal adherence and cavitation in murine candidiasis: a role for Candida acid proteinase. Infect Immun 1988; 56:1942-9; PMID:3294180.
- Moyes DL, Naglik JR. Mucosal immunity and Candida albicans infection. Clin Dev Immunol 2011; 2011:346307; PMID:21776285; http://dx.doi.org/10.1155/2011/346307
- Lowman DW, Greene RR, Bearden DW, Kruppa MD, Pottier M, Monteiro MA, Soldatov DV, Ensley HE, Cheng SC, Netea MG, et al. Novel structural features in Candida albicans hyphal glucan provide a basis for differential innate immune recognition of hyphae versus yeast. J Biol Chem 2014; 289:3432-43; PMID:24344127; http://dx.doi.org/10.1074/jbc.M113.529131
- Davis SE, Hopke A, Minkin SC Jr, Montedonico AE, Wheeler RT, Reynolds TB. Masking of beta(1-3)-Glucan in the Cell Wall of Candida albicans from Detection by Innate Immune Cells Depends on Phosphatidylserine. Infect Immun 2014; 82:4405-13; PMID:25114110; http://dx.doi.org/10.1128/IAI.01612-14
- Devillers A, Courjol F, Fradin C, Coste A, Poulain D, Pipy B, Bernardes ES, Jouault T. Deficient beta-mannosylation of Candida albicans phospholipomannan affects the proinflammatory response in macrophages. PLoS One 2013; 8:e84771; PMID:24367694; http://dx.doi.org/10.1371/journal.pone.0084771
- Villamon E, Gozalbo D, Roig P, O'Connor JE, Fradelizi D, Gil ML. Toll-like receptor-2 is essential in murine defenses against Candida albicans infections. Microbes Infect 2004; 6:1-7; PMID:14738887; http://dx.doi.org/10.1016/j.micinf.2003.09.020
- Tessarolli V, Gasparoto TH, Lima HR, Figueira EA, Garlet TP, Torres SA, Garlet GP, Da Silva JS, Campanelli AP. Absence of TLR2 influences survival of neutrophils after infection with Candida albicans. Med Mycol 2010; 48:129-40; PMID:19468929; http://dx.doi.org/10.3109/13693780902964339
- Netea MG, Sutmuller R, Hermann C, Van der Graaf CA, Van der Meer JW, van Krieken JH, Hartung T, Adema G, Kullberg BJ. Toll-like receptor 2 suppresses immunity against Candida albicans through induction of IL-10 and regulatory T cells. J Immunol 2004; 172:3712-8; PMID:15004175; http://dx.doi.org/10.4049/jimmunol.172.6.3712
- Bellocchio S, Montagnoli C, Bozza S, Gaziano R, Rossi G, Mambula SS, Vecchi A, Mantovani A, Levitz SM, Romani L. The contribution of the Toll-like/IL-1 receptor superfamily to innate and adaptive immunity to fungal pathogens in vivo. J Immunol 2004; 172:3059-69; PMID:14978111; http://dx.doi.org/10.4049/jimmunol.172.5.3059
- Murciano C, Villamon E, Gozalbo D, Roig P, O'Connor JE, Gil ML. Toll-like receptor 4 defective mice carrying point or null mutations do not show increased susceptibility to Candida albicans in a model of hematogenously disseminated infection. Med Mycol 2006; 44:149-57; PMID:16519018; http://dx.doi.org/10.1080/13693780500294733
- Taylor PR, Tsoni SV, Willment JA, Dennehy KM, Rosas M, Findon H, Haynes K, Steele C, Botto M, Gordon S, et al. Dectin-1 is required for beta-glucan recognition and control of fungal infection. Nat Immunol 2007; 8:31-8; PMID:17159984; http://dx.doi.org/10.1038/ni1408
- Saijo S, Fujikado N, Furuta T, Chung SH, Kotaki H, Seki K, Sudo K, Akira S, Adachi Y, Ohno N, et al. Dectin-1 is required for host defense against Pneumocystis carinii but not against Candida albicans. Nat Immunol 2007; 8:39-46; PMID:17159982; http://dx.doi.org/10.1038/ni1425
- Linden JR, De Paepe ME, Laforce-Nesbitt SS, Bliss JM. Galectin-3 plays an important role in protection against disseminated candidiasis. Med Mycol 2013; 51:641-51; PMID:23488971; http://dx.doi.org/10.3109/13693786.2013.770607
- Joly S, Ma N, Sadler JJ, Soll DR, Cassel SL, Sutterwala FS. Cutting edge: Candida albicans hyphae formation triggers activation of the Nlrp3 inflammasome. J Immunol 2009; 183:3578-81; PMID:19684085; http://dx.doi.org/10.4049/jimmunol.0901323
- Villamon E, Gozalbo D, Roig P, Murciano C, O'Connor JE, Fradelizi D, Gil ML. Myeloid differentiation factor 88 (MyD88) is required for murine resistance to Candida albicans and is critically involved in Candida -induced production of cytokines. Eur Cytokine Netw 2004; 15:263-71; PMID:15542452.
- De Luca A, Montagnoli C, Zelante T, Bonifazi P, Bozza S, Moretti S, D'Angelo C, Vacca C, Boon L, Bistoni F, et al. Functional yet balanced reactivity to Candida albicans requires TRIF, MyD88, and IDO-dependent inhibition of Rorc. J Immunol 2007; 179:5999-6008; PMID:17947673; http://dx.doi.org/10.4049/jimmunol.179.9.5999
- Romani L. Immunity to fungal infections. Nat Rev Immunol 2011; 11:275-88; PMID:21394104; http://dx.doi.org/10.1038/nri2939
- Ozment-Skelton TR, deFluiter EA, Ha T, Li C, Graves BM, Ferguson DA Jr, Schweitzer JB, Preizsner J, Brown GD, Gordon S, et al. Leukocyte Dectin-1 expression is differentially regulated in fungal versus polymicrobial sepsis. Crit Care Med 2009; 37:1038-45; PMID:19237915; http://dx.doi.org/10.1097/CCM.0b013e3181968fa0
- Ozment-Skelton TR, Goldman MP, Gordon S, Brown GD, Williams DL. Prolonged reduction of leukocyte membrane-associated Dectin-1 levels following beta-glucan administration. J Pharmacol Exp Ther 2006; 318:540-6; PMID:16632639; http://dx.doi.org/10.1124/jpet.106.102293
- Cheng SC, van de Veerdonk F, Smeekens S, Joosten LA, van der Meer JW, Kullberg BJ, Netea MG. Candida albicans dampens host defense by downregulating IL-17 production. J Immunol 2010; 185:2450-7; PMID:20624941; http://dx.doi.org/10.4049/jimmunol.1000756
- Rizzo A, Losacco A, Carratelli CR. Lactobacillus crispatus modulates epithelial cell defense against Candida albicans through Toll-like receptors 2 and 4, interleukin 8 and human beta-defensins 2 and 3. Immunol Lett 2013; 156:102-9; PMID:24120511; http://dx.doi.org/10.1016/j.imlet.2013.08.013
- Plantinga TS, Johnson MD, Scott WK, van de Vosse E, Velez Edwards DR, Smith PB, Alexander BD, Yang JC, Kremer D, Laird GM, et al. Toll-like receptor 1 polymorphisms increase susceptibility to candidemia. J Infect Dis 2012; 205:934-43; PMID:22301633; http://dx.doi.org/10.1093/infdis/jir867
- Nahum A, Dadi H, Bates A, Roifman CM. The biological significance of TLR3 variant, L412F, in conferring susceptibility to cutaneous candidiasis, CMV and autoimmunity. Autoimmun Rev 2012; 11:341-7; PMID:22024499; http://dx.doi.org/10.1016/j.autrev.2011.10.007
- Rosentul DC, Delsing CE, Jaeger M, Plantinga TS, Oosting M, Costantini I, Venselaar H, Joosten LA, van der Meer JW, Dupont B, et al. Gene polymorphisms in pattern recognition receptors and susceptibility to idiopathic recurrent vulvovaginal candidiasis. Front Microbiol 2014; 5:483; PMID:25295030; http://dx.doi.org/10.3389/fmicb.2014.00483
- Ferwerda B, Ferwerda G, Plantinga TS, Willment JA, van Spriel AB, Venselaar H, Elbers CC, Johnson MD, Cambi A, Huysamen C, et al. Human dectin-1 deficiency and mucocutaneous fungal infections. N Engl J Med 2009; 361:1760-7; PMID:19864674; http://dx.doi.org/10.1056/NEJMoa0901053
- von Bernuth H, Picard C, Jin Z, Pankla R, Xiao H, Ku CL, Chrabieh M, Mustapha IB, Ghandil P, Camcioglu Y, et al. Pyogenic bacterial infections in humans with MyD88 deficiency. Science 2008; 321:691-6; PMID:18669862; http://dx.doi.org/10.1126/science.1158298