
Enteroaggregative Escherichia coli (EAEC) comprise of a large, diverse group of diarrheagenic E. coli defined by their characteristically ‘stacked brick’ pattern on HEp-2 cells.Citation1 EAEC was first associated with children's diarrhea in developing countries but studies have since shown that EAEC is the cause of both acute and persistent diarrhea in all ages worldwide.Citation2–6 Moreover, EAEC has been implicated in numerous outbreaks, most notably the large outbreak in Germany in 2011 with an EAEC strain lysogenized with a prophage harboring a stx2a-converting phage, resulting in 855 cases of hemolytic-uremic syndrome and 54 deaths.Citation7–11
The pathogenesis of EAEC is not yet fully understood due to the heterogeneity among strains. EAEC are recognized as a diarrheal pathogen but are also isolated from healthy individuals stressing the need to be able to distinguish pathogenic from nonpathogenic EAEC strains. A variety of putative virulence factors have been identified, although none of these have been present only in the strains isolated from symptomatic patients.Citation12,13
Even though the pathogenesis of EAEC is unclear, the 3 following stages have been suggested to occur upon infection; 1) initial adherence to the intestinal mucosa, possible by the aggregative adherence fimbriae (AAFs),Citation14,15 2) biofilm formationCitation16 and 3) induction of an inflammatory response and the release of toxins.Citation16,17
Understanding the complex relationship between the host and the bacterium is a crucial step for revealing the pathogenicity of a certain strain. Although many different animal models have been proposed for this pathogen, none have been able to show all of the clinical manifestation of disease.Citation18–21 Thus, there is still an urgent need for a reproducible animal model that is able to show all aspects of EAEC pathogenesis. Recently, larvae of the greater wax moth Galleria mellonella were established as an acceptable model to study bacterial infections caused by several pathogens including Klebsiella pneumoniae, Listeria monocytogenes, Pseudomonas aeruginosa and extra-intestinal E. coli.Citation22–25 The model has shown many advantages such as decreasing rearing costs, convenient feasibility, ability to carry out experiments at 37 °C and most importantly, correlation was observed between the G. mellonella model and well established vertebrate models.Citation26–28
In this study, we sought to determine whether G. mellonella would be a suitable model for studying the virulence of EAEC infections.
The G. mellonella assays were performed as previously described by Morgan et al.Citation29 Briefly, bacterial overnight cultures were pelleted by centrifugation (4000 × g) and washed twice in PBS and finally re-suspended in PBS containing 10 % glucose. Groups of 10 larvae (200–250 mg) were infected with 10 μl aliquots of serially diluted bacterial suspensions (from 102 to 107 bacterial cells per larvae) by injection with a Hamilton syringe (26 gauge) via the last right proleg. Larvae were incubated at 37 °C after infection and survival was monitored for 96 hours.
Firstly, we investigated the virulence of 6 well characterized EAEC strains in the larvae model. One of the major challenges with EAEC is that the strains are highly heterogeneous with respect to genomic background, phylogroup, serotype, as well as their virulence genes. All strains tested harbored the 4 classical EAEC virulence factors: the transcriptional factor AggR, the aii island encoding a type VI secretion system, as well as dispersin and the dispersin transporter. Infection of G. mellonella with the EAEC strains resulted in rapid killing of the larvae and mortality was shown to be dependent on the number of bacteria injected (). For all of the EAEC strains tested, 100 % of the larvae were killed after 24 hours with an inoculum of 106 CFU/larvae, whereas no mortality was observed when infected with 102 CFU/larvae (data not shown).
Figure 1. EAEC infection of Galleria mellonella larvae. Larvae were infected with 6 different EAEC strains and injected with different doses of bacteria; shown in the figures are injections of A) 103 CFU of EAEC injected into the larvae, where strain 55989 showed to be significantly different compared to the other 5 strains (P < 0.001) and B) 104 CFU of EAEC injected into the larvae. All results represents means of at least 3 independent experiments with 10 larvae per treatment. Survival curves were plotted using the Kaplan-Meier method and statistical analysis were performed using the log rank test for multiple comparisons (GraphPad Software, San Diego, CA)
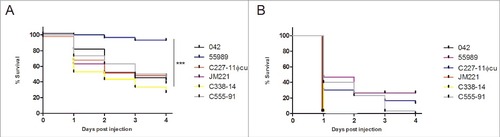
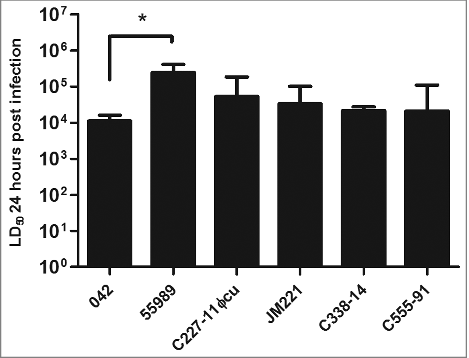
We next determined the LD50s after 24 hours of the strains as previously described.Citation30 Strain 042, the prototype EAEC strain which has also been shown experimentally to elicit diarrhea in human volunteers,Citation31 had the lowest LD50 of 1.11 × 104 CFU after 24 hours post infection. Compared to 042, the 5 other EAEC strains had similar LD50 values, except for strain 55989, which had a significantly higher LD50 compared to strain 042 (P < 0.05) ().
Figure 2. The LD50 of the strains were calculated using probit regression model (SPSS v. 20) for each isolate 24 h after G. mellonella larvae were inoculated with the bacteria. All strains showed to have similar LD50 value except for strain 55989, which had a significant higher LD50 than the other 5 strains tested (P < 0.05). Statistical significance between LD50 values were tested by performing one-way ANOVA with Dunnet post-test in GraphPad and the error bars displayed represent the 95 % confidence intervals
It has previously been shown that nonpathogenic E. coli strain DH5α is not lethal to G. mellonella at inocula up to 107 CFU/larvae.Citation32 We repeated these experiments with 3 commensal E. coli strains: MG1655Citation33, HS Citation34 and F-18,Citation35 which confirmed previous data shown with DH5α. None of these commensal E. coli strains tested were lethal in the larvae up to 107 CFU (data not shown), indicating that it is the presence of EAEC virulence factors mediating the killing of the larvae. A recent study with Caenorhabditis elegans and infection with commensal E. coli strains have shown the O-antigens play an essential role in virulence in this model.Citation36 Thus, whereas the O-rough MG1655 wildtype showed a low killing of C. elegans, restoration of O-antigen expression enabled the strain to kill C. elegans at rates similar to EAEC strain 042. Furthermore, the commensal E. coli strain HS expressing the O9 antigen was as virulent as virulent in the C. elegans model as EAEC stain 042. In contrast, we found in the G. mellonella model that commensal strain HS was avirulent. Thus, whereas O-antigen expression may likely influence virulence in the G. mellonella model, high virulence in this model is not merely related to O-antigen expression.
This was also confirmed by injecting the larvae with the 6 heat-killed EAEC strains (107 CFU/mL). No killing was observed for any of the strains, suggesting that it is alive EAEC and not the LPS mediating the killing of the larvae (data not shown).
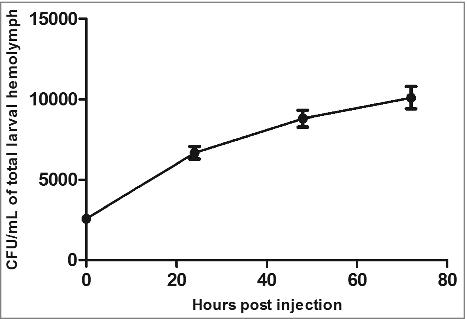
Next, we wanted to address whether the mortality of G. mellonellla is associated with the bacterial load of EAEC in the infected larvae. We therefore infected the larvae with 103 CFU of EAEC strain 042 and determined the bacterial load by enumeration of viable bacteria present at various time point. Each larvae were decapitated and 30 μl of hemolymph were collected using a sterile 1.5 ml Eppendorf tube as previously described.Citation37 The hemolymph was serially diluted and plated on selective MacConkey agar plates containing chloramphenicol.Citation38 Infection with strain 042 resulted in an increase of bacteria over time in the hemolymph, demonstrating that 042 replicates inside the larvae ().
Figure 3. EAEC strain 042 grows inside the larvae. The larvae were infected with 103 CFU and at the indicated time points the hemolymph of 3 larvae were collected and CFU determined by plating on selective agar plates. Error bars at each indicated time point represents standard error of the mean
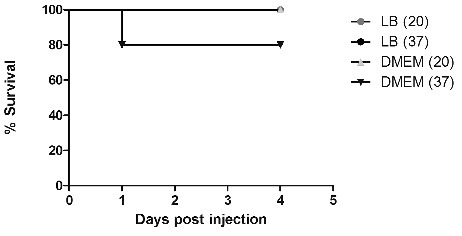
To evaluate whether a bacterial secretory product was involved in host death, a culture filtrate of strain 042 grown to stationary-phase at 20 and 37 °C in LB-broth or the cell medium DMEM/0.5% glucose which have previously shown to upregulate EAEC virulence genes.Citation39 The supernatants were concentrated 10 times using a 10 kDa amicon filter (Merck Millipore, Kenilworth, NJ) and inoculated into the larvae. The results showed that a small fraction of larvae were killed when grown in DMEM/0.5 % glucose at 37 °C, whereas no killing was observed at 20 °C in DMEM/0.5 % glucose or when grown in LB-broth ().
Figure 4. Supernatant of EAEC strain 042 has little effect on mortality. Strain 042 was grown stationary overnight in either LB-broth or DMEM/0.5 % glucose at 20 and 37 °C. The supernatants were then sterile filtered and concentrated 10 times using an 10 kDa amicon filter (Merck Millipore, Kenilworth, NJ), and 10 μl were injected into the larvae. The results represents the average of 3 experiments, repeated with 3 different batches of larvae. Statistical difference was calculated using the log rank test for multiple comparisons in GraphPad, however no statistical significance was observed
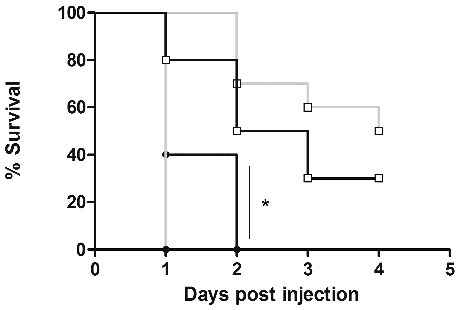
Lastly, we investigated whether the larvae incubation temperature changed the killing rate of larvae by EAEC strain 042. Larvae were injected as previously described and incubated at either 22 °C or 37 °C and mortality was recorded for up to 4 d From these experiments, we could conclude that the lowered temperature attenuated the killing of the larvae (). For example, at day 1 40 % of the larvae were still alive at room temperature with a dose of 104 CFU/larva whereas none of the larvae incubated at 37 °C were alive (P < 0.05). These data suggest that 042 has temperature sensitive virulence traits. This could be speculated to be related to a lower expression of the key regulator of EAEC virulence factors, AggR, which has been shown to be optimally expressed at 37 °C.Citation39
Figure 5. Strain 042 shows temperature dependent mortality. G. mellonella strains were infected with strain 042 (□103 and •104) and incubated at either 22 °C (blue) or 37 °C (black) for 4 d Statistical analysis were performed using the log rank test for multiple comparisons (GraphPad), and a significant difference was observed between larvae inoculated with 104 CFU and the 2 temperatures used (P < 0.05)
In conclusion, we report that EAEC is able to infect and kill G. mellonella in a dose and time dependent manner, and that the model is able to distinguish clearly between virulent wildtype strains of EAEC and non-pathogenic E. coli. Moreover, we also see that EAEC is able to survive and replicate in the larvae, and that viable EAEC are needed to cause mortality. Previous infection studies using ExPEC in the G. mellonella model showed that with doses below 5 × 106 CFU, less than 30 % of larvae were killed after 4 d whereas when using a dose above 5 × 106 CFU, over 80 % of the larvae were killed within 24 h.Citation40 We here show that a dose of 1 × 106 is sufficient for 100 % mortality within 24 h with the EAEC strain 042, suggesting that EAEC could be an even more virulent pathotype than ExPEC. Interestingly, a study investigating ExPEC and virulence factors found the Afa/Dr adhesins to be associated with significantly higher mortality, suggesting that the fimbriae is important in ExPEC pathogenicity.Citation41 The Afa/Dr adhesins are very homologous to the AAF fimbriae encoded by EAEC, and it could be speculated that AAF fimbriae plays a role in the high virulence of EAEC in the larvae.
However, many questions remains open, for example which virulence factors are mediating the killing of the larvae and is it one or a combination of multiple? To address these questions, it will be necessary to investigate a wide array of mutants from various strains, since EAEC is so heterogeneous and one virulence factor may be important in one strain but less important in another. That the G. mellonella model can be a valuable tool to future studies of EAEC pathogenicity is supported by the successful use of this model to study enteropathogenic E. coli (EPEC) virulence.Citation42 However, the model does not replace well-established mammalian models, but it is an inexpensive and reliable model providing the ability to study the difference between virulent and non-virulent EAEC strains, identification of putative virulence markers, and possible novel molecular targets for antimicrobial therapy and vaccine development.
Disclosure of potential conflicts of interest
The authors have no conflict of interest to declare
Acknowledgments
The authors wish to thank Kurt Fuursted for introducing us to the Galleria mellonella model.
Funding
We also want to thank Region Sjælland for financial support.
Related Research Data
References
- Nataro JP, Kaper JB, Robins-Browne R, Prado V, Vial P, Levine MM. Patterns of adherence of diarrheagenic Escherichia coli to HEp-2 cells. The Pediatr Infect Dis J 1987; 6:829-31; PMID:3313248; https://doi.org/https://doi.org/10.1097/00006454-198709000-00008
- Okhuysen PC, Dupont HL. Enteroaggregative Escherichia coli (EAEC): a cause of acute and persistent diarrhea of worldwide importance. J Infect Dis 2010; 202:503-5; PMID:20594108; https://doi.org/https://doi.org/10.1086/654895
- Pabalan N, Singian E, Jarjanazi H, Steiner TS. Enteroaggregative Escherichia coli and acute diarrhea in children: a meta-analysis of South Asian populations. Eur J Clin Microbiol Infect Dis 2013; 32:597-607; PMID:23179250; https://doi.org/https://doi.org/10.1007/s10096-012-1779-3
- Huang DB, Nataro JP, DuPont HL, Kamat PP, Mhatre AD, Okhuysen PC, Chiang T. Enteroaggregative Escherichia coli is a cause of acute diarrheal illness: a meta-analysis. Clin Infect Dis 2006; 43:556-63; PMID:16886146; https://doi.org/https://doi.org/10.1086/505869
- Fang GD, Lima AA, Martins CV, Nataro JP, Guerrant RL. Etiology and epidemiology of persistent diarrhea in northeastern Brazil: a hospital-based, prospective, case-control study. J Pediatr Gastroenterol Nutr 1995; 21:137-44; PMID:7472898; https://doi.org/https://doi.org/10.1097/00005176-199508000-00003
- Lima AA, Guerrant RL. Persistent diarrhea in children: epidemiology, risk factors, pathophysiology, nutritional impact, and management. Epidemiol Rev 1992; 14:222-42.
- Scheutz F, Nielsen EM, Frimodt-Moller J, Boisen N, Morabito S, Tozzoli R, Nataro JP, Caprioli A. Characteristics of the enteroaggregative Shiga toxin/verotoxin-producing Escherichia coli O104:H4 strain causing the outbreak of haemolytic uraemic syndrome in Germany, May to June 2011. Euro Surveill 2011; 16(24).
- Shin J, Oh SS, Oh KH, Park JH, Jang EJ, Chung GT, Yoo CK, Bae GR, Cho SH. An outbreak of foodborne illness caused by enteroaggregative escherichia coli in a high school in South Korea. Japn J Infect Dis 2015; 68:514-9; PMID:25971323; https://doi.org/https://doi.org/10.7883/yoken.JJID.2014.460
- Dallman TJ, Chattaway MA, Cowley LA, Doumith M, Tewolde R, Wooldridge DJ, Underwood A, Ready D, Wain J, Foster K, et al. An investigation of the diversity of strains of enteroaggregative Escherichia coli isolated from cases associated with a large multi-pathogen foodborne outbreak in the UK. PloS one 2014; 9:e98103; PMID:24844597; https://doi.org/https://doi.org/10.1371/journal.pone.0098103
- Cobeljic M, Miljkovic-Selimovic B, Paunovic-Todosijevic D, Velickovic Z, Lepsanovic Z, Zec N, Savić D, Ilić R, Konstantinović S, Jovanović B, et al. Enteroaggregative Escherichia coli associated with an outbreak of diarrhoea in a neonatal nursery ward. Epidemiol Infect 1996; 117:11-6; PMID:8760945; https://doi.org/https://doi.org/10.1017/S0950268800001072
- Harada T, Hiroi M, Kawamori F, Furusawa A, Ohata K, Sugiyama K, Masuda T. A food poisoning diarrhea outbreak caused by enteroaggregative Escherichia coli serogroup O126:H27 in Shizuoka, Japan. Japn J Infect Dis 2007; 60:154-5; PMID:17515660
- Boisen N, Scheutz F, Rasko DA, Redman JC, Persson S, Simon J, Kotloff KL, Levine MM, Sow S, Tamboura B, et al. Genomic characterization of enteroaggregative Escherichia coli from children in Mali. J Infect Dis 2012; 205:431-44; PMID:22184729; https://doi.org/https://doi.org/10.1093/infdis/jir757
- Lima IF, Boisen N, Quetz Jda S, Havt A, de Carvalho EB, Soares AM, Lima NL, Mota RM, Nataro JP, Guerrant RL, et al. Prevalence of enteroaggregative Escherichia coli and its virulence-related genes in a case-control study among children from north-eastern Brazil. J Med Microbiol 2013; 62:683-93; PMID:23429698; https://doi.org/https://doi.org/10.1099/jmm.0.054262-0
- Czeczulin JR, Balepur S, Hicks S, Phillips A, Hall R, Kothary MH, Navarro-Garcia F, Nataro JP. Aggregative adherence fimbria II, a second fimbrial antigen mediating aggregative adherence in enteroaggregative Escherichia coli. Infect Immun 1997; 65:4135-45; PMID:9317019
- Nataro JP, Deng Y, Maneval DR, German AL, Martin WC, Levine MM. Aggregative adherence fimbriae I of enteroaggregative Escherichia coli mediate adherence to HEp-2 cells and hemagglutination of human erythrocytes. Infect Immun 1992; 60:2297-304; PMID:1350273
- Hicks S, Candy DC, Phillips AD. Adhesion of enteroaggregative Escherichia coli to pediatric intestinal mucosa in vitro. Infect Immun 1996; 64:4751-60; PMID:8890236
- Harrington SM, Strauman MC, Abe CM, Nataro JP. Aggregative adherence fimbriae contribute to the inflammatory response of epithelial cells infected with enteroaggregative Escherichia coli. Cell Microbiol 2005; 7:1565-78; PMID:16207244; https://doi.org/https://doi.org/10.1111/j.1462-5822.2005.00588.x
- Zangari T, Melton-Celsa AR, Panda A, Boisen N, Smith MA, Tatarov I, De Tolla LJ, Nataro JP, O'Brien AD. Virulence of the Shiga toxin type 2-expressing Escherichia coli O104:H4 German outbreak isolate in two animal models. Infect Immun 2013; 81:1562-74; PMID:23439303; https://doi.org/https://doi.org/10.1128/IAI.01310-12
- Philipson CW, Bassaganya-Riera J, Hontecillas R. Animal models of enteroaggregative Escherichia coli infection. Gut Microbes 2013; 4:281-91; PMID:23680797; https://doi.org/https://doi.org/10.4161/gmic.24826
- Munera D, Ritchie JM, Hatzios SK, Bronson R, Fang G, Schadt EE, Davis BM, Waldor MK. Autotransporters but not pAA are critical for rabbit colonization by Shiga toxin-producing Escherichia coli O104:H4. Nat Commun 2014; 5:3080; PMID:24445323; https://doi.org/https://doi.org/10.1038/ncomms4080
- Roche JK, Cabel A, Sevilleja J, Nataro J, Guerrant RL. Enteroaggregative Escherichia coli (EAEC) impairs growth while malnutrition worsens EAEC infection: a novel murine model of the infection malnutrition cycle. J Infect Dis 2010; 202:506-14; PMID:20594107; https://doi.org/https://doi.org/10.1086/654894
- Insua JL, Llobet E, Moranta D, Perez-Gutierrez C, Tomas A, Garmendia J, Bengoechea JA. Modeling Klebsiella pneumoniae pathogenesis by infection of the wax moth Galleria mellonella. Infect Immun 2013; 81:3552-65; PMID:23836821; https://doi.org/https://doi.org/10.1128/IAI.00391-13
- Mukherjee K, Altincicek B, Hain T, Domann E, Vilcinskas A, Chakraborty T. Galleria mellonella as a model system for studying Listeria pathogenesis. Appl Environ Microbiol 2010; 76:310-7; PMID:19897755; https://doi.org/https://doi.org/10.1128/AEM.01301-09
- Miyata S, Casey M, Frank DW, Ausubel FM, Drenkard E. Use of the Galleria mellonella caterpillar as a model host to study the role of the type III secretion system in Pseudomonas aeruginosa pathogenesis. Infect Immun 2003; 71:2404-13; PMID:12704110; https://doi.org/https://doi.org/10.1128/IAI.71.5.2404-2413.2003
- Nathan S. New to Galleria mellonella: modeling an ExPEC infection. Virulence 2014; 5:371-4; PMID:24569500; https://doi.org/https://doi.org/10.4161/viru.28338
- Gao W, Chua K, Davies JK, Newton HJ, Seemann T, Harrison PF, Holmes NE, Rhee HW, Hong JI, Hartland EL, et al. Two novel point mutations in clinical Staphylococcus aureus reduce linezolid susceptibility and switch on the stringent response to promote persistent infection. PLoS Pathog 2010; 6:e1000944; PMID:20548948; https://doi.org/https://doi.org/10.1371/journal.ppat.1000944
- Jander G, Rahme LG, Ausubel FM. Positive correlation between virulence of Pseudomonas aeruginosa mutants in mice and insects. J Bacteriol 2000; 182:3843-5; PMID:10851003; https://doi.org/https://doi.org/10.1128/JB.182.13.3843-3845.2000
- Insua JL, Llobet E, Moranta D, Perez-Gutierrez C, Tomas A, Garmendia J, Bengoechea JA. Modeling Klebsiella pneumoniae Pathogenesis by infection of the wax moth galleria mellonella. Infect Immun 2013; 81:3552-65; PMID:23836821; https://doi.org/https://doi.org/10.1128/IAI.00391-13
- Morgan JK, Ortiz JA, Riordan JT. The role for TolA in enterohemorrhagic Escherichia coli pathogenesis and virulence gene transcription. Microb Pathog 2014; 77:42-52; https://doi.org/https://doi.org/10.1016/j.micpath.2014.10.010
- Giannouli M, Antunes LC, Marchetti V, Triassi M, Visca P, Zarrilli R. Virulence-related traits of epidemic Acinetobacter baumannii strains belonging to the international clonal lineages I-III and to the emerging genotypes ST25 and ST78. BMC Infect Dis 2013; 13:282; PMID:23786621; https://doi.org/https://doi.org/10.1186/1471-2334-13-282
- Nataro JP, Deng Y, Cookson S, Cravioto A, Savarino SJ, Guers LD, Levine MM, Tacket CO. Heterogeneity of enteroaggregative Escherichia coli virulence demonstrated in volunteers. J Infect Dis 1995; 171:465-8; PMID:7844392; https://doi.org/https://doi.org/10.1093/infdis/171.2.465
- Alghoribi MF, Gibreel TM, Dodgson AR, Beatson SA, Upton M. Galleria mellonella infection model demonstrates high lethality of ST69 and ST127 uropathogenic E. coli. PloS one 2014; 9:e101547; PMID:25061819
- Scheutz F, Nielsen EM, Frimodt-Møller J, Boisen N, Morabito S, Tozzoli R, Nataro JP, Caprioli A. Blattner FR, Plunkett G rd, et al. The complete genome sequence of Escherichia coli K-12. Science 1997; 5;277(5331):1453-62; https://doi.org/https://doi.org/10.1126/science.277.5331.1453
- Levine MM, Bergquist EJ, Nalin DR, Waterman DH, Hornick RB, Young CR, Sotman S. Escherichia coli strains that cause diarrhoea but do not produce heat-labile or heat-stable enterotoxins and are non-invasive. Lancet 1978; 1:1119-22; PMID:77415; https://doi.org/https://doi.org/10.1016/S0140-6736(78)90299-4
- Sweeney NJ, Laux DC, Cohen PS. Escherichia coli F-18 and E. coli K-12 eda mutants do not colonize the streptomycin-treated mouse large intestine. Infect Immun 1996; 64:3504-11; PMID:8751891
- Browning DF, Wells TJ, Franca FL, Morris FC, Sevastsyanovich YR, Bryant JA, Johnson MD, Lund PA, Cunningham AF, Hobman JL, et al. Laboratory adapted Escherichia coli K-12 becomes a pathogen of Caenorhabditis elegans upon restoration of O antigen biosynthesis. Mol Microbiol 2013; 87:939-50; PMID:23350972; https://doi.org/https://doi.org/10.1111/mmi.12144
- Harding CR, Schroeder GN, Collins JW, Frankel G. Use of Galleria mellonella as a model organism to study Legionella pneumophila infection. J Vis Exp 2013; (81):e50964; https://doi.org/https://doi.org/10.3791/50964.
- Chaudhuri RR, Sebaihia M, Hobman JL, Webber MA, Leyton DL, Goldberg MD, Cunningham AF, Scott-Tucker A, Ferguson PR, Thomas CM, et al. Complete genome sequence and comparative metabolic profiling of the prototypical enteroaggregative Escherichia coli strain 042. PloS One 2010; 5:e8801; https://doi.org/https://doi.org/10.1371/journal.pone.0008801
- Morin N, Tirling C, Ivison SM, Kaur AP, Nataro JP, Steiner TS. Autoactivation of the AggR regulator of enteroaggregative Escherichia coli in vitro and in vivo. FEMS Immunol Med Microbiol 2010; 58:344-55; PMID:20132305; https://doi.org/https://doi.org/10.1111/j.1574-695X.2009.00645.x
- Williamson DA, Mills G, Johnson JR, Porter S, Wiles S. In vivo correlates of molecularly inferred virulence among extraintestinal pathogenic Escherichia coli (ExPEC) in the wax moth Galleria mellonella model system. Virulence 2014; 5:388-93; PMID:24518442; https://doi.org/https://doi.org/10.4161/viru.27912
- Ciesielczuk H, Betts J, Phee L, Doumith M, Hope R, Woodford N, Wareham DW. Comparative virulence of urinary and bloodstream isolates of extra-intestinal pathogenic Escherichia coli in a Galleria mellonella model. Virulence 2015; 6:145-51; PMID: 25853733; https://doi.org/https://doi.org/10.4161/21505594.2014.988095
- Leuko S, Raivio TL. Mutations that impact the enteropathogenic Escherichia coli Cpx envelope stress response attenuate virulence in Galleria mellonella. Infect Immun 2012; 80:3077-85; https://doi.org/https://doi.org/10.1128/IAI.00081-12