
ABSTRACT
Mounting evidence suggests a connection between human parvovirus B19 (B19) and autoimmune diseases, and especially an association between the B19-VP1 unique region (VP1u) and anti-phospholipid syndrome (APS). However, little is known about the antigenicity of B19-VP1u in the induction of APS-like syndrome. To elucidate the antigenicity of B19-VP1u in the induction of APS, N-terminal truncated B19-VP1u (tVP1u) proteins were prepared to immunize Balb/c mice to generate antibodies against B19-tVP1u proteins. The secreted phospholipase A2 (sPLA2) activities and binding specificity of mice anti-B19-tVP1u antibodies with cardiolipin (CL) and beta-2-glycoprotein I (β2GPI) were evaluated by performing immunoblot, ELISA and absorption experiments. A mice model of passively induced APS was adopted. Although sPLA2 activities were identified in all B19-tVP1u proteins, only amino acid residues 61–227 B19-tVP1u exhibited a higher sPLA2 activity. Autoantibodies against CL and β2GPI exhibited binding activities with all B19-tVP1u proteins. IgG that was purified from mice that had been immunized with amino acid residues 21–227 to 121–227 B19-tVP1u proteins exhibited significantly higher binding activity with CL. IgG that was purified from mice that had been immunized with amino acid residues 21–227, 31–227, 82–227 and 91–227 B19-tVP1u proteins exhibited significantly higher binding activity with β2GPI. Accordingly, significantly higher binding inhibition of CL was detected in the presence of amino acid residues 61–227 and 101–227 B19-tVP1u. Significantly higher binding inhibition of β2GPI was detected in the presence of amino acid residues 21–227, 31–227, 82–227 and 91–227 B19-tVP1u. The mice that received amino acid residues 31–227 or 61–227 anti-tB19-VP1u IgG revealed significant thrombocytopenia and those that received amino acid residues 21–227, 31–227, 61–227, 71–227, 82–227, 91–227, 101–227 or 114–227 anti-tB19-VP1u IgG exhibited significantly prolonged aPTT. These findings provide further information concerning the role of B19-VP1u antigenicity in APS-like autoimmunity.
Introduction
The antiphospholipid syndrome (APS) is an autoimmune disorder that is characterized by arterial and venous thromboses, recurrent fetal loss, thrombocytopenia, prolonged coagulation and elevated serum titers of antiphospholipid antibodies (APLs), including anti-beta2-glycoprotein-I (anti-β2GPI) and anti-cardiolipin antibody (aCL).Citation1-4 Human parvovirus B19 (B19) is a human pathogen of erythrovirus in the Parvoviridae family.Citation5-6 Hence, B19 infection has been associated with autoimmune disordersCitation7-8 especially systemic lupus erythematosus (SLE) and the induction of various autoantibodies, including anti-neutrophil cytoplasmic antibody, APL, and aCL.Citation1,9-10
APLs, notably aCL, occur frequently in viral infections, including in HIV, HBV, HCV and B19.Citation11 Evidence has also shown that anti-β2GPI and aCL have been detected in patients with B19-infection and SLE.Citation7,12-15 Notably, a remarkable similarity between the APLs associated with SLE and B19-infection has been reported. Interestingly, aCL from patients with B19-infection increased their binding to antigens in the presence of β2GPI as a cofactor, as does in aCL from SLE patients, but unlike antibodies from patients with other viral infections.Citation9
The B19-VP1-unique region (VP1u), which except for the 227 amino acids at N-terminal end of VP1 protein, identical to VP2, plays a crucial role in autoimmunity.Citation16-18 Previous studies have shown that autoantibodies against CL, β2GPI, and phospholipid (PhL) in sera from patients with B19 infection cross-react with B19-VP1u.Citation16 Similar results have been obtained from animal experiments in which significantly elevated immunoglobulins against B19-VP1u, CL, β2GPI and PhL were detected in mice sera that were immunized with recombinant B19-VP1u protein.Citation17 In a passive transfer mice model, APS-like syndrome, which involves striking thrombocytopenia, prolonged activated partial thromboplastin time (aPTT) and elevated titers of autoantibodies against β2GPI and CL, have been detected in mice that received anti-B19-VP1u IgG.Citation17 These findings strongly associate B19-VP1u with autoimmunity, and notably with APS-like syndrome. Therefore, this study further elucidates the antigenicity of B19-VP1u by the N-terminal truncation of B19-VP1u.
Materials and methods
Preparation of VP1u and truncated B19-VP1u proteins
The full-length B19-VP1u expression vector pET-32a(+)was used as the template for the construction of truncations.Citation16 To synthesize DNA templates for N-terminal deletions, the truncated forms of VP1u cDNA were generated by PCR, named amino acid residues 21–227 (2), 31–227 (3), 51–227 (4), 61–227 (5), 71–227 (6), 82–227 (7), 91–227 (8), 101–227 (9), 114–227 (10), 121–227 (11) and 130–227 (12). All forward and reverse primers have restriction enzyme sites BglII and Sal I that facilitate the cloning of VP1u fragments into the pET-32a(+) expression vector. The ligatant, pET32a-tVP1u, was transformed into Escherichia Coli BL21-DE3 competent cells, which were obtained from Invitrogen (Carlsbad, California, USA). All constructs were confirmed by sequencing. E. coli (BL21-DE3) clones that contained VP1u or tVP1u cDNA in the pET-32a expression vector (Novagene, Cambridge, MA) were grown overnight in one-liter of L-Broth that contained 100 μg/ml ampicillin at 37°C with shaking. When the OD 600 reached 0.7-0.9, protein expression was induced by the addition of IPTG to a concentration of 1 mM for another 3 hr. The cells were harvested by centrifugation at 4000 g for 20 min and resuspended in 20 ml sonication buffer (50 mM NaPO4 pH 8/0.25 mM EDTA). Lysozyme was added to a final concentration of 1 mg/ml and the buffer was kept on ice for 30 min. The cells were sonicated (W385, Heat systems-ultrasonic, INC) for a total of 30 min at 5 min intervals, and then centrifuged at 10,000 g for 30 min. The pellet was dissolved with 10 ml buffer B (8 M urea; 0.1M NaH2PO4; 0.02M Tris-HCl; pH 8.0) for 1 hr at room temperature, and the lysate was centrifuged 10,000 g for 30 min at room temperature to pellet the cellular debris. The supernatant was loaded onto an Ni-NTA spin column (Qiagen, Chatsworth, CA, USA) and PureProteome™ Nickel Magnetic Beads (EMD Millipore, CA, USA), and the recombinant B19-VP1u proteins were purified. Protein concentrations were determined by the RC DC protein assay kit (Bio-Rad, CA, USA) and quantified using a U3000 spectrophotometer (Hitachi, Tokyo, Japan). For endotoxin tests, the E-TOXATE (Limulus Amebocyte Lysate) test kits (Sigma-Aldric, St. Louis, MO, USA) was used. The endotoxin levels were measured and found to be below the detection limit in these recombinant proteins (data not shown).
Immunoblotting
Sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) was performed as described elsewhereCitation19 and the samples of VP1u and tVP1u were electrophoretically transferred to nitrocellulose membranes according to the method of Towbin.Citation20 The membranes were then cut and soaked in 5% nonfat dry milk in PBS. Antibodies were diluted with 5% nonfat dry milk in PBS reacted with the nitrocellulose strips and incubated for 1.5 hr at room temperature with Histidine tag (6xHis) mouse monoclonal antibody (Invitrogen, CA, USA). The strips were then washed and incubated with alkaline phosphatase conjugated goat anti-mice IgG antibodies (Sigma, Saint Louis Mo, USA) at a dilution of 1/1000. The substrate nitroblue tetrazolium/5-bromo-4-chloro-3 indolyl phosphate (NBT/BCIP) (USB Corporation, Ohio, USA) was used to detect antigen-antibody complexes. ACA HRP IgG Calibrator A and anti-β2GPI IgG Calibrator A (INOVA Diagnostics, Inc, CA, USA) were incubated for 3 h with gentle agitation at room temperature. After it had been washed twice with TBS-Tween, secondary antibody conjugated with horseradish peroxidase (HRP) was added and incubated for another hour. Immobilion Western HRP Chemiluminescent Substrate (Millipore, USA) was then used to detect the antigen-antibody complexes.
sPLA2 catalytic activity
VP1u and tVP1u proteins were assayed for sPLA2 activity by a colorimetric assay (sPLA2 Activity Kit; Cayman Chemical), in accordance with the manufacturer's instructions. Dynamic colorimetric measurements of the optical density at 414 nm were made every minute for 14 min.Citation18 They are expressed as micromoles per minute per milliliter. Ten μl bee venom PLA2 control was used as a positive control.
Mice antisera
Fifty-two six-week old female BALB/c mice were purchased from the National Laboratory Animal Center, Taiwan. At an age of eight weeks, they were randomly divided into 13 groups and housed under supervision of the Institutional Animal Care and Use Committee at Chung Shan Medical University, Taichung, Taiwan, which approved all procedures and protocols (Affidavit of Approval of Animal Use Protocol, No. 1130) followed the Guide for the Care and Use of Laboratory Animals published by the United States National Institutes of Health. All animals were kept in a 12-hour light-dark cycle and the ambient temperature was maintained at 25°C. Animals had free access to water and standard laboratory chow (Lab Diet 5001; PMI Nutrition International Inc., Brentwood, MO, USA). To generate antisera against B19-VP1u and B19-tVP1u proteins, 12 groups of four adult female BALB/c mice were immunized with subcutaneous injections of 2 μg purified recombinant B19-VP1u protein in Freund's complete adjuvant (Sigma, Saint Louis Mo, USA), and subsequently injected at two-week intervals with 2 μg B19-VP1u truncated proteins in Freund's incomplete adjuvant (Sigma, Saint Louis Mo, USA). A control group were immunized with Freund's complete adjuvant and then injected with Freund's incomplete adjuvant. The experimental period was eight weeks long and the mice were sacrificed at an age of 16 weeks by asphyxiation using CO2. All sera reacted strongly with B19-VP1u and B19-tVP1u, according to immunoblotting analysis. As described elsewhere,Citation21-23 sera from mice of each group were pooled, divided in 3 aliquots for experimental triplicates, and stored at −80°C until further processing. The mice control, anti-B19-VP1u IgG and anti-B19-tVP1u IgG were purified using a PureProteome™ Protein G Magnetic Bead System (EMD Millipore, CA, USA). The elutant was concentrated and quantified using a U3000 spectrophotometer (Hitachi, Tokyo, Japan).
ELISA
Antigen-specific ELISA kits to detect aCL IgG and anti-β2GPI IgG antibodies (INOVA Diagnostics, Inc. San Diego, CA, USA) as described by the manufacturer. All serum samples (one μg of IgG) were assayed at a dilution of 1:101. After incubation at room temperature for 30 minutes, the liquid was removed from each sample, which was washed with phosphate buffered saline (PBS) and subsequently incubated with horseradish peroxidase (HRP)-conjugated goat anti-mouse IgG (Sigma, Saint Louis Mo, USA) at a dilution of 1/1000. After incubation at room temperature for 30 minutes, the liquid from each sample well was removed and then washed with PBS and subsequently incubated with the color reaction TMB Chromogen (INOVA Diagnostics, Inc. San Diego, CA, USA) in the dark for 30 minutes at room temperature. Finally, 100ul of HRP stop solution (INOVA Diagnostics, Inc. San Diego, CA, USA) was added to each well and the absorbance (OD) of the well at 450 nm was read within one hour of stopping the reaction. For the absorption experiments, the sera were pre-incubated with 500 μM VP1u or tVP1u proteins for one hour at 37°C before ELISA was performed. A cutoff value was determined using sera from CTL mice (mean+2SD) and absorbance above the value was regarded as positive.
Mice and induction of experimental APS by passive transfer
Fifty-two five week-old female BALB/c mice were purchased from the National Laboratory Animal Center, Taiwan. At an age of eight weeks, they were randomly divided into 13 groups and housed under supervision of the Institutional Animal Care and Use Committee at Chung Shan Medical University, Taichung, Taiwan, which approved all procedures and protocols (Affidavit of Approval of Animal Use Protocol, No.1459), which followed the Guide for the Care and Use of Laboratory Animals, which was published by the United States National Institutes of Health. All animals were kept in a 12-hour light-dark cycle and the ambient temperature was maintained at 25°C. Animals had free access to water and standard laboratory chow (Lab Diet 5001; PMI Nutrition International Inc., Brentwood, MO, USA). The induction of experimental APS by passive transfer was performed according to the method of Blank.Citation24 A group of four BALB/c mice (eight weeks of age) were infused with the mouse anti-VP1u or anti-tVP1u IgG antibodies (20 μg per mouse) through their tail veins. Affinity-purified IgG from control mice that were immunized with adjuvant and PBS were used as controls for immunization. After two weeks, a boost injection of 20 μg of the immunoglobulin in PBS into the tail vein was performed. Starting two weeks later, the mice were the same amount of antibodies was injected intradermally in the hind footpads of the mice at one-week intervals. The experimental period was ten weeks long and the mice were sacrificed at an age of 18 weeks by asphyxiation using CO2. APS clinical parameters, including thrombocytopenia and prolonged activated partial thromboplastin time [aPTT], were evaluated in the infused mice. Platelets from individual blood samples were quantified in whole blood using Systemex (KX-21, KOBE, Japan). The amount of lupus anticoagulants present was evaluated by the prolongation of aPTT in mixing tests, one volume of plasma from whole blood was mixed with 0.123 mol/l of sodium citrate in a 9:1 volume ratio with 1 each cephalin, before the mixture was incubated for two minutes at 37°C. Another volume of 0.025M CaCl2 (Sigma, Saint Louis Mo, USA) was added, and the clotting time in seconds was recorded using Coatron M1 (TECO GmbH, Neufahrn NB, Germany).
Statistical analysis
All of the statistical analyses were conducted using GraphPad Prism 5 software (GraphPad Software, CA). They involved one-way analysis of variance (One-way ANOVA), followed by the Tukey multiple-comparisons test. Data were represented as mean ± SEM and verified by at least three independent experiments. A value of P < 0.05 was considered to indicate statistical significance. Significant differences are indicated by symbols in the relevant figures.
Results
Preparation of recombinant B19-tVP1u proteins
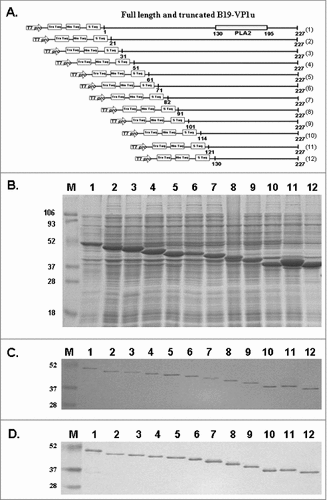
schematically depicts full-length and truncated B19-VP1u with N-terminal deletions. The full-length and 11 N-terminal truncated B19-VP1u recombinant proteins (tVP1u) were purified and analyzed using 12% SDS-PAGE ( and ). Histidine tag (6xHis) mouse monoclonal antibody was used to identify the purified truncated recombinant proteins (). The number 1 indicates the full-length of the B19-VP1u protein, which contains amino acid residues 1 to 227. The numbers 2 to 12 indicated the N-terminal truncated B19-VP1u proteins, which contain amino acid residues 21 to 227 (2), 31 to 227 (3), 51 to 227 (4), 61 to 227 (5), 71 to 227 (6), 82 to 227 (7), 91 to 227 (8), 101 to 227 (9), 114 to 227 (10), 121 to 227 (11) and 130 to 227 (12), respectively. Since the B19-VP1u and all tVP1u are constructed into pET-32a expression vector, the expressed recombinant proteins will contain S-tag and His-tag and exhibit the following molecular weight; (1) 51 kDa, (2) 49 kDa, (3) 48 kDa, (4) 46 kDa, (5) 45 kDa, (6) 44 kDa, (7) 43 kDa, (8) 42 kDa, (9) 40 kDa, (10)39 kDa, (11) 38 kDa, (12) 38 kDa.
Figure 1. Full-length and truncated B19-VP1u (tVP1u) proteins. (A) Schematic representation of full-length (1) and deletions of B19-VP1u. Eleven truncated B19-VP1u recombinant proteins correspond to amino acid sequences 21–227 (2), 31–227 (3), 51–227 (4), 61–227 (5), 71–227 (6), 82–227 (7), 91–227 (8), 101–227 (9), 111–227 (10), 121–227 (11) and 130–227 (12). Separation of (B) total lysate and (C) purified B19-VP1u proteins in a 12.5% SDS-PAGE. (D) Identification of purified recombinant B19-tVP1u proteins with anti-histidine monoclonal antibodies. Lane M indicates pre-stained protein marker; lane 1 indicates full-length B19-VP1u, and lanes 2 to 12 correspond to truncated deletions 2 to 12, respectively.
Secreted phospholipase A2 (sPLA2) activity of truncated B19-VP1u proteins
To determine whether B19-tVP1u proteins exhibit sPLA2 activity, a sPLA2 activity assay was performed. summarizes the sPLA2 activities in full-length, mutated and truncated B19-VP1u proteins. As a positive control, bvPLA2 exhibited sPLA2 activity of 0.528 ± 0.008 µmol/min/mL, whereas B19-VP1uD175A exhibited no detectable sPLA2 activity, as described previously.Citation18 sPLA2 activities were detected in full-length recombinant B19-VP1u and B19-tVP1u proteins with values of 0.124 ± 0.011 (1), 0.006 ± 0.003 (2), 0.022 ± 0.011 (3), 0.025 ± 0.012 (4), 0.172 ± 0.051 (5), 0.030 ± 0.010 (6), 0.011 ± 0.010 (7), 0.021 ± 0.010 (8), 0.017 ± 0.011 (9), 0.027 ± 0.010 (10), 0.034 ± 0.010 (11) and 0.005 ± 0.003 (12) μmol/min/mL (). Notably, higher sPLA2 activity was detected in amino acid residues 61 to 227 (5) B19-tVP1u protein, whereas lower sPLA2 activity was detected in amino acid residues 21 to 227 (2) and 130 to 227 (12) B19-tVP1u proteins ().
Table 1. Secreted phospholipase A2 (sPLA2) activity of recombinant full -length and N-terminal truncated B19-VP1 unique region (B19-tVP1u) proteins.
Antibody against CL and β2GPI cross-reacts with B19-tVP1u proteins
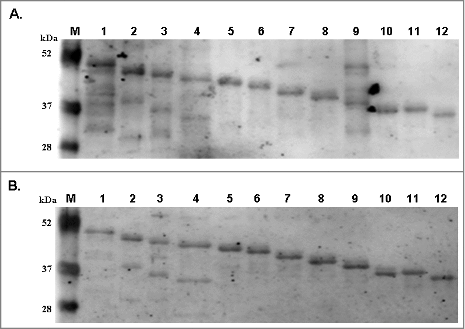
To clarify the association between B19-VP1u and APL, the binding activities of B19-tVP1u proteins with autoantibodies against CL and β2GPI were examined. displays the immunoblot results obtained by probing B19-tVP1u proteins with autoantibodies against CL and β2GPI. The experimental results revealed that autoantibodies against CL () and β2GPI () cross-reacted with all B19-tVP1u proteins.
Figure 2. Reactivity of antibodies against cardiolipin and β2GPI to truncated B19-VP1u (B19-tVP1u) proteins. Full-length B19-VP1u and B19-tVP1u proteins were probed with antibodies against (A) cardiolipin and (B) β2GPI. Lane M indicates pre-stained protein marker; lane 1 indicates the full-length B19-VP1u, and lanes 2 to 12 correspond to the truncated deletions 2 to 12, respectively.
Anti-sera from mouse immunized with B19-tVP1u proteins reveal reactivity against cardiolipin and β2GPI
To analyze the binding activity of antibodies against B19-tVP1u with CL and β2GPI, One μg of IgG against B19-tVP1u proteins was purified from Balb/c mice for ELISA analysis. The normal absorbance values of the controls were 0.191 ± 0.03 (mean ± 2 SD) for CL () and 0.137 ± 0.01 (mean ± 2 SD) for β2GPI (), and these were set as cutoff values for significance. Except for IgG against the amino acid residues 130 to 227 (12) B19-tVP1u protein, IgG against full-length B19-VP1u and the other B19-tVP1u proteins exhibited detectable binding activity with CL. Notably, significantly higher reactivity with CL was observed in IgG against full-length B19-VP1u, and amino acid residues 61 to 227 (5) and 101–227 (9) B19-tVP1u proteins (). IgG against full-length B19-VP1u and amino acid residues 21 to 227 (2), 31 to 227 (3), 82 to 227 (7) and 91 to 227 (8) B19-tVP1u proteins exhibited a significantly higher binding activity with β2GPI (). The binding specificity of purified mice IgG to CL autoantigen was analyzed by absorption assays. Significantly higher binding inhibition of CL was observed with full-length B19-VP1u, and the amino acid residues 61 to 227 (5) and 101 to 227 (9) B19-tVP1u, than the control (). Significantly higher binding inhibition of β2GPI was observed with full-length B19-VP1u, and amino acid residues 21 to 227 (2), 31 to 227 (3), 82 to 227 (7) and 91 to 227 (8) B19-tVP1u, than the control ().
Figure 3. Binding activity of antibodies against B19-tVP1u with CL and β2GPI. IgG against B19-tVP1u was purified from BALB/c mice and reacted with (A) CL and (B) β2GPI. By ELISA analysis, normal absorbance of controls was 0.191 ± 0.03 (mean ± 2 SD) for CL and 0.137 ± 0.01 (mean ± 2 SD) for β2GPI. # indicates significance, P<0.05, relative to the controls (CTL). N = 4 mice per group.
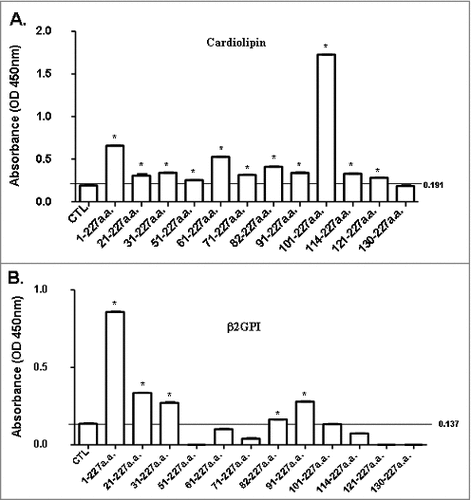
Table 2. Absorption of mouse anti-B19-VP1u or B19-tVP1u antibodies with purified B19-VP1u or B19-tVP1u proteins for binding of cardiolipin autoantigen.
Table 3. Absorption of mouse anti-B19-VP1u or B19-tVP1u antibodies with purified B19-VP1u or B19-tVP1u proteins forβ2GPI autoantigen.
Induction of passive transfer of anti-phospholipid syndrome (APS) by infusion of anti-B19-tVP1u antibody
To clarify whether anti-B19-tVP1u antibodies can induce anti-phospholipid syndrome, a passively induced APS mice model was adopted. Whole blood from Balb/c mice that were intravenously immunized with purified anti-B19-VP1u or anti-B19-VP1u antibodies was collected and hematological and coagulation examinations were performed. No significant difference between the WBC, RBC, Hb, HCT, MCV, MCH, MCHC, lymphocyte or neutrophils of mice that had been immunized with anti-full-length B19-VP1u and control mice was detected (). Significant thrombocytopenia and prolonged aPTT were detected in mice that had received full-length B19-VP1u, relative to the control mice (). The mice that received amino acid residues 31 to 227 (3) or 61 to 227 (5) anti-tB19-VP1u IgG exhibited significant thrombocytopenia, as determined by comparison with the controls (). Notably, the mice had received amino acid residues 21 to 227 (2), 31 to 227 (3), 61 to 227 (5), 71 to 227 (6), 82 to 227 (7), 91 to 227 (8), 101 to 227 (9) or 114 to 227 (10) anti-tB19-VP1u IgG exhibited significantly longer aPTT than the controls ().
Table 4. Mice infused with various B19-tVP1u antibodies.
Discussion
B19 infection has been associated with various autoimmune diseases.Citation1,8 Although many studies have investigated the association between B19-VP1u and APS, the antigenicity of B19-VP1u in induction of APS-like syndrome remains unclear. This study further investigates the regions of B19-VP1u that contribute to the induction of APS-like syndrome by truncating the N-terminal of B19-VP1u and a passively induced APS mice model. The experimental results revealed that the region of amino acids 61 to 227 exhibited high sPLA2 activity. Furthermore, CL and β2GPI autoantibodies exhibited binding activity with all B19-tVP1u proteins. The mice IgG against all B19-tVP1u exhibited significantly higher binding activity with CL and those against amino acid residues 21 to 227 (2), 31 to 227 (3), 82 to 227 (7) and 91 to 227 (8) B19-tVP1u exhibited significantly higher binding activity with β2GPI. Notably, the mice that received IgG against 31 to 227 (3) or 71 to 227 (5) B19-tB19-VP1u revealed significant thrombocytopenia and those mice receiving IgG against amino acid residues 21 to 227 (2), 31 to 227 (3), 61 to 227 (5), 71 to 227 (6), 82 to 227 (7), 91 to 227 (8), 101 to 227 (9) or 114 to 227 (10) anti-tB19-VP1u exhibited significantly prolonged aPTT. These experimental results provide further information about the role of B19-VP1u antigenicity in induction of APS-like autoimmunity.
B19-VP1 is identical to VP2, except for the 227 amino acids at N-terminal end of VP1 protein, the so-called VP1-unique region (VP1u).Citation25 The amino acid sequence of B19-VP1u from 130 to 195 has been identified as having a conserved PLA2-like motif (HDXXY) and, which shares homologies to the Ca2+-binding loop and the catalytic site of secreted PLA2.Citation26-27 Conversely, mutations of certain amino acid residues in the B19 VP1u such as VP1uD175A, greatly reduce PLA2 activity and virus infectivity.Citation28-29 However, the modulatory effects of the region of B19-VP1u at amino acids 1 to 129 on PLA2-like activity remain unknown. Although PLA2 activities were detected in all B19-tVP1u proteins, only the truncated B19-VP1u protein that corresponded to the amino acid sequences 61 to 227 exhibited higher PLA2 activity (). These results reasonably suggest that the B19-VP1u region that corresponded to amino acid sequence from 61 to 71 may have been the domain that activated PLA2 activity whereas the amino acid sequence from 21 to 61 may have contributed to the inhibition of PLA2 activity. However, further investigations are required to clarify the conformational change of recombinant B19-tVP1u that renders these regions accessible and triggers the PLA2 potential.
Human B19-VP1u potent neutralizing epitopes and contributes to a dominant immune response following B19 infection.Citation30-31 Notably, evidence studies have found that the most neutralizing antisera are produced using peptides from the N-terminal part of the VP1u.Citation25,30 One study has shown that the clustering of neutralizing epitopes in the N-terminal 100 amino acids of B19-VP1u suggests the presence of important functional motifs within this region.Citation32 Therefore, the N-terminal amino acids positions 5 to 80 of B19-VP1u, comprising the receptor-binding domain (RBD), are necessary and sufficient for cellular binding, internalization and B19-virus uptake.Citation33 These findings imply that the N-terminal region of B19-VP1u has a crucial role in the induction of autoimmunity. Although previous studies have reported that antibodies against B19-VP1u cross-react with CL and β2GPI,Citation15,17 the critical amino acids within B19-VP1u that are involved in the induction of binding with CL and β2GPI were not elucidated. This study provides further information about the antigenicity of the N-terminal part of B19-VP1u. According to Western blot analysis, the autoantibodies against CL and β2GPI reacted with all N-terminal truncated B19-VP1u proteins, suggesting the presence of epitopes for autoantibodies against CL and β2GPI within amino acids 130 to 227 of B19-VP1u.
The diagnosis of APS is essentially based on not only common clinical manifestations but also the detection of circulating APLs, such as antibodies against CL and β2GPI.Citation34 In the clinic, β2GPI has been identified as the major autoantigen in APS and a specific ELISA for detecting antibodies against β2GPI has been introduced for the diagnosis of APS in the laboratory.Citation34-35 This state anti-β2GPI antibody is a specific index for APS. In this study, purified mice IgG against amino acid residues 21 to 227 (2), 31 to 227 (3), 82 to 227 (7) and 91 to 227 (8) B19-tVP1u proteins exhibited significantly greater binding activity with β2GPI. Consistent therewith, significantly higher binding inhibition of β2GPI was detected in the presence of amino acid residues 21 to 227 (2), 31 to 227 (3), 82 to 227 (7) and 91 to 227 (8) B19-tVP1u. Notably, purified mice IgG against amino acid residues 21 to 227 (2) to 121 to 227 (11) B19-tVP1u exhibited significantly higher binding activity with CL. However, only amino acid residues of 61 to 227 (5) and 101 to 227 (9) B19-tVP1u proteins exhibited a significantly higher binding inhibition to CL in an absorption assay. These experimental results imply that the immunization of B19-tVP1u induces more specific reactivity to β2GPI than to CL. Although further investigations are required, B19-VP1u and β2GPI are reasonably speculated to share more similarities in epitope distribution and conformation than CL.
Finally, there is a concern in the present study should also be noted. We found that anti-sera against B1-tVP1u (a.a. 61–227) has overall low levels of anti-cardiolipin antibodies, but can be efficiently absorbed with recombinant B19-tVP1u (a.a. 61–227). This may be due to the conformational specificity of truncated B19-VP1u proteins and the interference of the truncated B19-VP1u proteins during the competition reaction. Although further investigations are required to verify this deviation, similar tendency and consistency of these truncated B19-VP1u proteins between antibody binding activity assay and absorption assay were still observed.
Evidence is emerging that B19-VP1u has an important role in the development of autoimmunity, and in particular APS-like syndrome, through the direct infection of cells or antibody formation.Citation17,36-38 Indeed, a common pathogenicity of the APS and the autoimmunity features that are observed in B19 infection is supported not only by the close association between the presence of APL and parvovirus B19 infection but also by the similarity between the presented clinical symptoms of patients with parvovirus B19 infection and those with APS.Citation3,15 However, the antigenic regions of B19-VP1u in induction of APS-like syndrome are still obscure. This study reveals that the passive transfer of IgG against amino acid residues 31 to 227 (3) or 61 to 227 (5) B19-tVP1u induces significant thrombocytopenia and those against amino acid residues 21 to 227 (2), 31 to 227 (3), 61 to 227 (5), 71 to 227 (6), 82 to 227 (7), 91 to 227 (8), 101 to 227 (9) or 114 to 227 (10) induces significantly prolonged aPTT in naïve mice. Altogether, this investigation provides advanced information concerning the critical domains of B19-VP1u in the induction of APS-like autoimmunity.
Disclosure and potential conflict of interest
No potential conflicts of interests were disclosed.
Additional information
Funding
References
- Lunardi C, Tiso M, Borgato L, Nanni L, Millo R, De Sandre G, Severi AB, Puccetti A. Chronic parvovirus B19 infection induces the production of anti-virus antibodies with autoantigen binding properties. Eur J Immunol. 1998;28:936–948. doi:10.1002/(SICI)1521-4141(199803)28:03<936::AID-IMMU936>3.3.CO;2-O 10.1002/(SICI)1521-4141(199803)28:03<936::AID-IMMU936>3.0.CO;2-X. PMID:9541589
- Bertolaccini M, Khamashta MA. Laboratory diagnosis and management challenges in the antiphospholipid syndrome. Lupus. 2006;15:172–178. doi:10.1191/0961203306lu2293rr. PMID:16634372
- Von Landenberg P, Lehmann HW, Modrow S. Human parvovirus B19 infection and antiphospholipid antibodies. Autoimmun Rev. 2007;6:278–285. doi:10.1016/j.autrev.2006.09.006. PMID:17412298
- Pavlovic M, Kats A, Cavallo M, Shoenfeld Y. Clinical and molecular evidence for association of SLE with parvovirus B19. Lupus. 2010;19:783–792. doi:10.1177/0961203310365715. PMID:20511275
- Young NS, Brown KE. Parvovirus B19. N Engl J Med. 2004;350:586–97. doi:10.1056/NEJMra030840. PMID:14762186
- Qiu J, Söderlund-Venermo M, Young NS. Human Parvoviruses. Clin Microbiol Rev. 2017;30:43–113. doi:10.1128/CMR.00040-16. PMID:27806994
- Lehmann HW, von Landenberg P, Modrow S. Parvovirus B19 infection and autoimmune disease. Autoimmun Rev. 2003;2:218–223. doi:10.1016/S1568-9972(03)00014-4. PMID:12848949
- Meyer O. Parvovirus B19 and autoimmune diseases. Joint Bone Spine. 2003;70:6–11. doi:10.1016/S1297-319X(02)00004-0. PMID:12639611
- Loizou S, Cazabon JK, Walport MJ, Tait D, So AK. Similarities of specificity and cofactor dependence in serum antiphospholipid antibodies from patients with human parvovirus B19 infection and from those with systemic lupus erythematosus. Arthritis Rheum. 1997;40:103–108. doi:10.1002/art.1780400115. PMID:9008606
- von Landenberg P, Lehmann HW, Knoll A, Dorsch S, Modrow S. Antiphospholipid antibodies in pediatric and adult patients with rheumatic disease are associated with parvovirus B19 infection. Arthritis Rheum. 2003;48:1939–1947. doi:10.1002/art.11038. PMID:12847688
- Sène D, Piette JC, Cacoub P. Antiphospholipid antibodies, antiphospholipid syndrome and infections. Autoimmun Rev. 2008;7:272–277. doi:10.1016/j.autrev.2007.10.001. PMID:18295729
- Hsu TC, Tsay GJ. Human parvovirus B19 infection in patients with systemic lupus erythematosus. Rheumatology (Oxford). 2001;40:152–157. doi:10.1093/rheumatology/40.2.152. PMID:11257151
- Kalt M, Gertner E. Antibodies to beta 2-glycoprotein I and cardiolipin with symptoms suggestive of systemic lupus erythematosus in parvovirus B19 infection. J Rheumatol. 2001;28:2335–2336. PMID:11669178
- Von Landenberg P, Modrow S. Human parvovirus B19 infection and antiphospholipid- syndrome: the two sides of one medal? J Vet Med B Infect Dis Vet Public Health. 2005;52:353–355. doi:10.1111/j.1439-0450.2005.00876.x. PMID:16316400
- von Landenberg P, Döring Y, Modrow S, Lackner KJ. Are antiphospholipid antibodies an essential requirement for an effective immune response to infections? Ann N Y Acad Sci. 2007;1108:578–583. doi:10.1196/annals.1422.060. PMID:17894022
- Tzang BS, Tsay GJ, Lee YJ, Li C, Sun YS, Hsu TC. The association of VP1 unique region protein in acute parvovirus B19 infection and antiphospholipid antibody production. Clin Chim Acta. 2007;378:59–65. doi:10.1016/j.cca.2006.10.016. PMID:17169353
- Tzang BS, Lee YJ, Yang TP, Tsay GJ, Shi JY, Tsai CC, Hsu TC. Induction of antiphospholipid antibodies and antiphospholipid syndrome-like autoimmunity in naive mice with antibody against human parvovirus B19 VP1 unique region protein. Clin Chim Acta. 2007;382:31–36. doi:10.1016/j.cca.2007.03.014. PMID:17451664
- Tzang BS, Chiu CC, Tsai CC, Lee YJ, Lu IJ, Shi JU, Hsu TC. Effects of human parvovirus B19 VP1 unique region protein on macrophage responses. J Biomed Sci. 2009;16:13. doi:10.1186/1423-0127-16-13. PMID:19272185
- Laemmli UK. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature. 1970;227:680–684. doi:10.1038/227680a0. PMID:5432063
- Towbin H, Stachelin T, Gordon J. Electrophoretic transfer of proteins from polyacrylamid gels to nitrocellulose sheet: procedure and some applications. Proc Natl Acad Sci USA. 1979;76:4350–4354. doi:10.1073/pnas.76.9.4350. PMID:388439
- Olszewska W, Obeid OE, Steward MW. Protection against measles virus-induced encephalitis by anti-mimotope antibodies: the role of antibody affinity. Virology. 2000;272:98–105. doi:10.1006/viro.2000.0285. PMID:10873752
- Li J, Zhang WB, Wilson M, Ito A, McManus DP. A novel recombinant antigen for immunodiagnosis of human cystic echinococcosis. J Infect Dis. 2003;188:1951–1960. doi:10.1086/379976. PMID:14673776
- Vignier N, Amor F, Fogel P, Duvallet A, Poupiot J, Charrier S, Arock M, Montus M, Nelson I, Richard I, et al. Distinctive serum miRNA profile in mouse models of striated muscular pathologies. PLoS One. 2013;8:e55281. doi:10.1371/journal.pone.0055281. PMID:23418438
- Blank M, Cohen J, Toder V, Shoenfeld Y. Induction of anti-phospholipid syndrome in naive mice with mouse lupus monoclonal and human polyclonal anti-cardiolipin antibodies. Proc Natl Acad Sci U S A. 1991;88:3069–3073. doi:10.1073/pnas.88.8.3069. PMID:2014226
- Anderson S, Momoeda M, Kawase M, Kajigaya S, Young NS. Peptides derived from the unique region of B19 parvovirus minor capsid protein elicit neutralizing antibodies in rabbits. Virology. 1995;206:626–632. doi:10.1016/S0042-6822(95)80079-4. PMID:7530397
- Zadori Z, Szelei J, Lacoste MC, Li Y, Gariépy S, Raymond P, Allaire M, Nabi IR, Tijssen P. A viral phospholipase A2 is required for parvovirus infectivity. Dev Cell. 2001;1:291–302. doi:10.1016/S1534-5807(01)00031-4. PMID:11702787
- Dorsch S, Liebisch G, Kaufmann B, von Landenberg P, Hoffmann JH, Drobnik W, Modrow S. The VP1 unique region of parvovirus B19 and its constituent phospholipase A2-like activity. J Virol. 2002;76:2014–2018. doi:10.1128/JVI.76.4.2014-2018.2002. PMID:11799199
- Lu J, Zhi N, Wong S, Brown KE. Activation of synoviocytes by the secreted phospholipase A2 motif in the VP1-unique region of parvovirus B19 minor capsid protein. J Infect Dis. 2006;193:582–590. doi:10.1086/499599. PMID:16425138
- Filippone C, Zhi N, Wong S, Lu J, Kajigaya S, Gallinella G, Kakkola L, Söderlund-Venermo M, Young NS, Brown KE. VP1u phospholipase activity is critical for infectivity of full-length parvovirus B19 genomic clones. Virology. 2008;374:444–452. doi:10.1016/j.virol.2008.01.002. PMID:18252260
- Saikawa T, Anderson S, Momoeda M, Kajigaya S, Young NS. Neutralizing linear epitopes of B19 parvovirus cluster in the VP1 unique and VP1-VP2 junction regions. J Virol. 1993;67:3004–3009. PMID:7684458
- Zuffi E, Manaresi E, Gallinella G, Gentilomi GA, Venturoli S, Zerbini M, Musiani M. Identification of an immunodominant peptide in the parvovirus B19 VP1 unique region able to elicit a long-lasting immune response in humans. Viral Immunology. 2001;14:151–158. doi:10.1089/088282401750234529. PMID:11398810
- Leisi R, Ruprecht N, Kempf C, Ros C. Parvovirus B19 uptake is a highly selective process controlled by VP1u: A novel determinant of viral tropism. J Virol. 2013;87:13161–13167. doi:10.1128/JVI.02548-13. PMID:24067971
- Leisi R, Di Tommaso C, Kempf C, Ros C. The Receptor-Binding Domain in the VP1u Region of Parvovirus B19. Viruses. 2016;8:61. doi:10.3390/v8030061. PMID:26927158
- Miyakis S, Lockshin MD, Atsumi T, Branch DW, Brey RL, Cervera R, Derksen RHWM, deGroot PG, Koike T, Meroni PL, et al. International consensus statement on an update of the classification criteria for definite antiphospholipid syndrome (APS). J Thromb Haemost. 2006;4:295–306. doi:10.1111/j.1538-7836.2006.01753.x. PMID:16420554
- Pengo V, Banzato A, Bison E, Denas G, Zoppellaro G, Bracco A, Padayattil Jose S, Hoxha A, Ruffatti A. Laboratory testing for antiphospholipid syndrome. Int J Lab Hematol 2016;38(Suppl 1):27–31. doi:10.1111/ijlh.12507. PMID:27161602
- Tzang BS, Tsai CC, Chiu CC, Shi JY, Hsu TC. Up-regulation of adhesion molecule expression and induction of TNF-alpha on vascular endothelial cells by antibody against human parvovirus B19 VP1 unique region protein. Clin Chim Acta. 2008;395:77–83 doi:10.1016/j.cca.2008.05.012. PMID:18538665
- Hsu TC, Chiu CC, Chang SC, Chan HC, Shi YF, Chen TY, Tzang BS. Human parvovirus B19 VP1u Protein as inflammatory mediators induces liver injury in naïve mice. Virulence. 2016;7:110–118. doi:10.1080/21505594.2015.1122165. PMID:26632342
- Adamson-Small LA, Ruiz-Ordaz BH. Hepatic inflammation resulting from localized, subcutaneous expression of Parvovirus VP1u capsid protein: Importance and implication. Virulence. 2016;7:70–71. doi:10.1080/21505594.2016.1141163. PMID:26786960