Abstract
The density and diversity of woody plant species were studied within grazing, cultivation and charcoal production land-use areas in a multiple-use savanna woodland, central Uganda, using 75 plots with an area of 0.1 ha (Whittaker plots). Plant density was significantly higher under charcoal production (7131 ± 755 plants/ha) and cultivation (6612 ± 665 plants/ha) compared with the grazing lands (4152 ± 525 plants/ha). At the plot level, species richness and Fisher's alpha diversity (α) were relatively low, ranging 2–31 species and 0.34–6.34, respectively, but both were significantly higher under charcoal production and cultivation compared with grazing. Similarly, cumulative species richness and Fisher's alpha diversity were higher under charcoal production and cultivation compared with grazing. Community species composition differed significantly (Global R ANOSIM = 0.14, p = 0.001; ANOSIM, ANalysis Of SIMilarity) among land uses. However, the distance of sampling plots away from households, the assumed source of human disturbance to woodlands, accounts for a very small fraction (<14.2%) of the variation in woody species diversity within this multiple-use savanna. Beta diversity was highest under grazing, followed by cultivation and charcoal production, suggesting a more heterogeneous spatial distribution of species under grazing. Within each land use, mean similarities were low, ranging 25–31%. Mean pairwise dissimilarities between land uses were relatively high, ranging 73–81%. This suggests that variations in species composition and diversity are to a great extent influenced by land-use and anthropogenic disturbances. The relatively low woody species diversity and richness in this savanna indicates woodland degradation, fragmentation and local species loss resulting from unsustainable harvesting for charcoal, and short interval shifting cultivation.
Introduction
Savanna woodlands make up most of the tropical and subtropical woodland cover in sub-Saharan Africa and provide a wide range of benefits and products to local and national production. They provide the major source of energy for the majority of households (Luoga et al. Citation2000a; Kituyi Citation2004; Millennium Ecosystem Assessment Citation2005a) and are the focus of numerous livelihood activities in marginalized communities (Luoga et al. Citation2000b; Dovie et al. Citation2002, Citation2005; Venter and Witkowski Citation2011). These savanna woodlands have numerous wild plant species that are important in the subsistence sector, both for meeting a wide range of household needs and through increasing commercialization, generating incomes (Braedt and Stranda-Gunda Citation2000; Dovie et al. Citation2005). The southern and eastern African miombo woodlands are economically important for supplying timber, poles, firewood, charcoal, medicines, food, fibre and carvings for both rural and urban populations (Luoga et al. Citation2000a, Citation2000b).
There is, however, a widespread concern that the woodland ecosystems with unique and valuable biodiversity resources are being lost (Rennolls and Laumonier Citation2000; Coetzer et al. Citation2010) as a result of both natural and anthropogenic disturbances and mismanagement (Luoga et al. Citation2004; Mwavu and Witkowski Citation2008a). They are rapidly undergoing severe large-scale changes, through indiscriminate burning or conversion to other land uses and cover types (Rennolls and Laumonier Citation2000; Millennium Ecosystem Assessment Citation2005a, Citation2005b). The development of more sustainable land-use systems necessitates an understanding of the nature, dynamics and production potential of these savanna woodland systems. An understanding of the patterns of woody plant species diversity within the prevailing land uses and responses to changes in the environment is essential for long-term sustainability and to aid decisions on conservation priorities (Folke et al. Citation2004).
Relatively little is known about the woody plant diversity in African savannas outside southern Africa and the miombo ecoregion, and this is particularly the case in Uganda. Most diversity studies in Uganda have focused on tropical rainforests (e.g. Eilu et al. Citation2004; Mwavu and Witkowski Citation2008a). Studies of savanna woodland in Uganda have mainly focused on biomass and bioenergy; management of ranching schemes (National Environment Management Authority Citation2002); and the growth, use and management of woodland resources (e.g. Namaalwa et al. Citation2005). Ugandan woodlands also play a major (often unaccounted) role in the economy by providing fuel (charcoal and firewood), grazing resources for livestock, land for cultivation and numerous other services. The charcoal is transported to major commercial centres such as Kampala. Woodland conversion to farmland, selective harvesting of woody plants for firewood and charcoal production, seasonal fires, livestock grazing and hunting of native herbivores are major mechanisms of woodland degradation, habitat change and biodiversity loss in many parts of Africa (Luoga et al. Citation2005; Banda et al. Citation2006).
This study aims to assess woody plant density and species diversity (richness, alpha diversity and beta diversity) in response to the three main land uses (grazing, cultivation and charcoal production) in the extensive savanna woodlands of Nakasongola District, central Uganda. The increasing human pressure on these tropical ecosystems makes information on their patterns of plant diversity (i.e. species richness, alpha diversity and beta diversity) and distributions more critical today than ever before in order to protect and conserve the remaining species effectively and efficiently (e.g. Cadotte et al. Citation2002; Natta et al. Citation2002; Zapfack et al. Citation2002). Alpha diversity and beta diversity together provide a generally good overall assessment of plant diversity or biotic heterogeneity of an area. Beta diversity is important in at least indicating the degree to which ecosystems have been partitioned by species.
Materials and methods
Study area
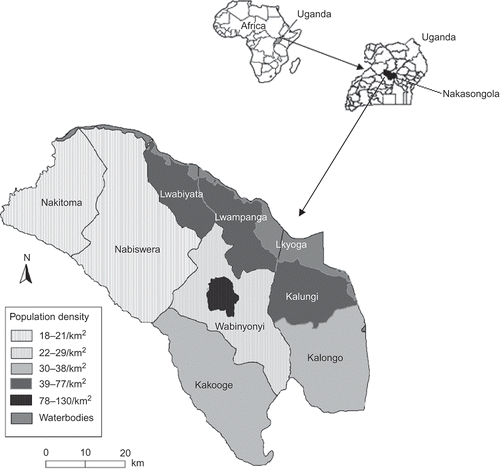
The savanna woodlands of Nakasongola District occur south of Lake Kyoga, central Uganda (0°40′–1°41′ N, 31°57′–32°48′ E). The district covers 3424 km2, 90% of which is woodland and grassy savanna, with 322 km2 (∼10%) of open water and wetlands. It comprises eight subcounties (administrative subdivisions), namely Kakooge, Kalongo, Wabinyonyi, Kalungi, Nabiswera, Nakitoma, Lwampanga and Lwabiyata. Subcounties consist of villages, ranging from 15–77 to 1741–4877 households. By 2001, the district had a total human population of 128,126 (41 people/km2), with 50.2% males and 49.8% females. About 95% of the population are in rural areas (Uganda Bureau of Statistics Citation2002).
Annual rainfall ranges from 500 to 1000 mm and is concentrated within two wet seasons (March–May and August–November). Reliability of rainfall received is higher in the south and declines gradually towards the north. The mean monthly maximum temperature ranges from 25°C to 35°C, and the mean monthly minimum temperature ranges from 18°C to 21°C. The topography of the area undulates from 1036 to 1160 m above sea level. The major geological formations are characterized by the presence of young intrusive rocks, mostly acidic and less commonly basic. The youngest formations date from the Pleistocene era and are represented by sands, quartz and clays of alluvial or lacustrine (i.e. formed at the bottom or along the shores of lakes as geological strata) origin (Parker et al. Citation1967).
The vegetation of the area is classified as Albizia–Combretum woodland (Langdale-Brown et al. Citation1964). This is a natural savanna woodland or woodland of mixed deciduous trees, 3–12 m in height and grasses, 0.3–1.3 m in height at maturity. The cover of the grass layer varies with season, is often patchy and subordinate to the tree layer. Most of the grazing lands occur on hydromorphic grasslands that are only suitable for grazing. There are thicket patches dominated by Acacia hockii, Acacia gerradii, Acacia kirkii ssp. mildbraedii, Acacia senegal and Euphorbia candelabrum established in secondary wooded grasslands as a consequence of anthropogenic disturbances (White Citation1983). The vegetation in Nakasongola District is mainly open woodland in transition to thicket/shrubland, as woodland cover has decreased, while areas under cultivation/settlements have correspondingly greatly increased since 1984 (Kalema Citation2010). In Nakasongola District, 70–80% of households not only engage in charcoal production but also livestock husbandry, and subsistence agriculture, mainly shifting cultivation, provides 85% of the food supply, and more than 50% of monetary income (Nakasongola Local Government Citation2002; ).
Figure 1. Map of Nakasongola District showing the eight constituent subcounties and their human population density. Inset is a map of Africa showing Uganda and a map of Uganda showing Nakasongola District shaded (TC: Town centre; Lkyoga: Lake Kyoga).
Sampling design
There is a sequence of land-use change over time, with most areas used for slash and burn agriculture; charcoal production areas are subsequently used for cultivation (they are burnt and hence have temporarily raised soil nutrient levels suitable for crops), and may then be used for grazing for a short period, allowing the woody plants to recover by basal resprouting from the cut stumps (Luoga et al. Citation2004; Mwavu and Witkowski Citation2008a). The current land use was identified for the purposes of this study. Representative villages (and surroundings) in each of the eight subcounties were selected (with the help of local leaders and officials from the Department of Natural Resources and Environment) during March 2006 for woody vegetation sampling. In each subcounty at least three transects were established. Transects were laid radiating away from the villages/households where subsistence activities are paramount, often resulting in gradients of increasing resource availability or decreasing human disturbance with distance from village (Fisher et al. Citation2011). Each transect represented one of the three major land uses, with the exception of Nabiswera, which had four transects. The additional transect was an attempt to include the most degraded grazing area. Transect locations were selected considering the homogeneous nature of the area for a particular land use and of sufficiently large extent to accommodate at least three 20 × 50 m (0.1 ha) plots, with a minimum separation distance of 200 m. The 20 ×50 m (Whittaker) plot size has been widely used for assessment of shrublands, grasslands, tropical savannas, woodlands and forests (Cowling and Witkowski Citation1994; Luoga et al. Citation2002; Beater et al. Citation2008; Dovie et al. Citation2008; Witkowski and Garner Citation2008). Hence, 24 plots with an area of 0.1 ha were sampled for each of the cultivation and charcoal production land-use types, and 27 plots were sampled for grazing. In total, 25 transects with 75 plots were established ().
Figure 2. Photographs of the major land-use practices: (a) wood stacked in preparation for charcoal production, (b) cattle grazing in a wetland and (c) cultivation fallow in a multiple land-use equatorial African savanna, central Uganda.

Within each 0.1 ha plot, woody plant species, number of individuals and height and stem diameter at reference height (1.3 m, except for seedlings and shrubs) of each individual were recorded. Based on stem diameter and height, woody plants were categorized as trees or shrubs (USDA Forest Service Citation1989). Seedlings had a single stem with stem diameter of ≤0.5 cm, were ≤1 m in height and had not previously resprouted, while saplings or juvenile trees were >0.5–5 cm dbh and >1 m in height. However, the seedling data were not included in this assessment. Species identification in the field was supported by identification guides mainly based on the Flora of Tropical East Africa (Polhill Citation1952 and subsequent volumes), Katende et al. (Citation1995) as well as the assistance of a botanist familiar with the flora of the area. Voucher species were collected and subsequently identified in the Botany Department Herbarium, Makerere University (MHU), Kampala, Uganda.
Data analysis
Alpha diversity
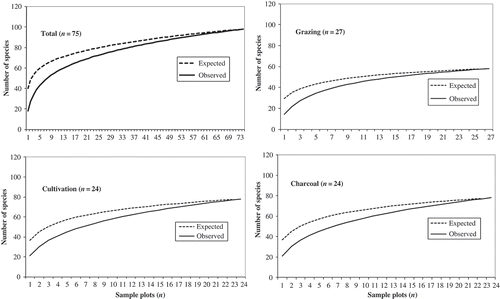
Fisher's alpha (α) and Shannon–Wiener (H′) diversity indices were employed to quantify alpha diversity at the 0.1 ha scale, while rarefaction was used to estimate the number of species expected (E(Sn)) to be present in a random sample (i.e. plot) of individuals taken from any given collection, and provides confidence limits of species richness (Magurran Citation2004). Plotting the rarefaction curves facilitates improved interpretation of species richness from sample plots of varying size and number and from different communities (Gotelli and Colwell Citation2001; Williams et al. Citation2005). The sample plots were randomized 100 times to compute the mean estimator and expected species richness for each sample plot accumulation level using EstimateS (Colwell Citation2006).
Fisher's alpha diversity index is not influenced by the size of the sampling area (i.e. plot) and is less affected by the abundance of the rarest or commonest species compared with other diversity measures (Magurran Citation2004). Fisher's alpha was, however, not calculated separately for both the tree and shrub data due to the high number of singletons in the individual data sets, which resulted in number of individuals (N) divided by number of species (S) being ≤1.44. Fisher's alpha cannot be calculated when N/S ≤ 1.44 (Magurran Citation2004). Diversity indices were calculated using Species Diversity and Richness® version IV (Pisces Conservation Ltd., Lymington, UK) (Seaby and Henderson Citation2007) and EstimateS version 8.0 (Colwell Citation2006). The sample plots were randomized 100 times in EstimateS to compute the cumulative H′ and Fisher's α and to construct the curves. Differences in tree and shrub species richness and diversity between the three land-use types were compared using one-way Analysis of Variance and Tukey's Honestly Significant Difference (HSD) for unequal sample sizes (i.e. different number of plots per land-use type). We also tested for the relationship between woody plant diversity (i.e. species richness and alpha diversity) and the ‘distance of the sampling plot along the transect radiating away from the households’, the assumed source of human disturbance, using regression analysis. The regression analyses were performed separately for (i) total woody (i.e. trees and shrubs combined), (ii) trees and (iii) shrubs at both the whole savanna and land-use type scales.
Beta diversity
Beta diversity at the plot level was assessed using Whittaker's beta diversity index (βW), Wilson and Schmida's index (βT) and the modified Morisita–Horn index of similarity (C MH). The Morisita–Horn index assesses similarity in species composition between plots (Colwell et al. Citation2004; Magurran Citation2004), and is a quantitative similarity index that is not strongly influenced by species richness and sample size. For each land-use type, C MH was calculated by averaging all the plot pairwise values for each land-use type. Morisita–Horn index varies from 1 (complete similarity) to 0 (complete dissimilarity) in species composition.
Plots were also arranged according to land-use type and variations in community species composition were tested by employing ANalysis Of SIMilarity (ANOSIM), a permutation test in CAP® 4 (Pisces Conservation Ltd.) (Seaby et al. Citation2007). ANOSIM computes a test statistic (R ANOSIM) reflecting the observed differences among replicates (i.e. plots) between sites (in this case land-use types), contrasted with differences among replicates within sites. A zero (0) occurs if the high and low similarities are perfectly mixed and bears no relationship to the group (i.e. plots within a land-use type). A value of minus one (−1) indicates that the most similar sample plots are all outside of the group. A value of positive one (+1) indicates that the most similar sample plots are within the same group (Seaby et al. Citation2007). The percentage similarity among land uses was also analysed using SIMilarity PERcentages (SIMPER) in CAP® 4. SIMPER breaks down the contribution of each species to the observed similarity (or dissimilarity) between sample plots. It allows the user to identify the species that are most important in creating the observed pattern of similarity. The method uses the Bray–Curtis measure of similarity, comparing in turn, each sample in Group 1 with each sample in Group 2. The Bray–Curtis method operates at the species level and therefore the mean similarity between groups 1 and 2 can be obtained for each species ().
Table 1. Brief description of the indices and measures applied to the data
Results
Woody plant community composition
A total of 99 woody plant species, from 67 genera within 31 families, were recorded from the 75 plots with an area of 0.1 ha. The most species-rich families were Mimosaceae (13 species), Rubiaceae (9), Moraceae (7), Euphorbiaceae (7), Anacardiaceae (6), Combretaceae (5) and Verbenaceae (5), with 15 others comprising 2–4 species and 9 with one species each (Appendix 1). The most species-rich genera were Acacia (8 species) Combretum (4), Ficus (4), Albizia (3), Pavetta (3) and Tricalysia (3), with the rest each having 1–2 species (Appendix 1). None of the 99 species has a restricted range as they all occur in more than one of the four floral regions of Uganda (Flora of Tropical East Africa (Polhill Citation1952 and more recent volumes). However, Milicia excelsa and Pouteria sp. are near threatened (International Union for Conservation of Nature classification; Pomeroy et al. Citation2002). Calliandra sp., Artocarpus heterophyllus, Senna siamea and Lantana camara are introduced and the latter is also invasive (Appendix 1).
The species accumulation curves for the combined 75 plots with an area of 0.1 ha, and for each land-use type asymptotes are reached, show that sampling was sufficient (). In addition, the species accumulation curve was below the rarefaction curve, suggesting heterogeneity among samples.
Figure 3. Rarefaction (expected) and species accumulation (observed) curves for the total woody species recorded in the 0.1 ha plots throughout the area (total), and for the grazing, cultivation and charcoal production land-use types, within this multiple-use equatorial African savanna, central Uganda.
Community species composition was significantly different among land uses (ANOSIM, Global R ANOSIM = 0.14, p = 0.001). Pairwise comparisons showed significant differences in species composition between cultivation and grazing, as well as grazing and charcoal production, but not between cultivation and charcoal production (). Woody plant density across all land-use types was 5893 ± 399 plants/ha. Density differed significantly (F 2,72 = 6.3, p = 0.003) among land uses, with charcoal production and cultivation significantly higher than grazing ().
Table 2. ANOSIM in community species composition among the different land-use types in a multiple-use equatorial African savanna, central Uganda
Table 3. Plot-level (0.1 ha) woody plant density, and species diversity and richness (means ± SE) in a multiple-use equatorial African savanna, central Uganda
Woody plant species richness and alpha diversity
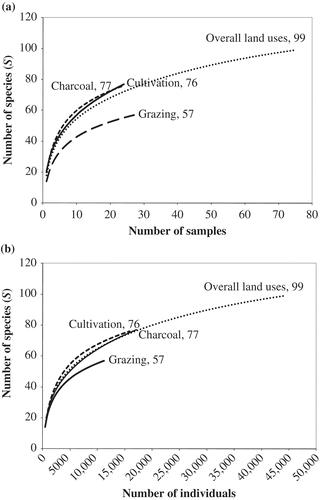
At the plot level, species richness differed (F 2,72 = 10.7, p < 0.0001) among land uses, with charcoal production and cultivation being significantly higher than grazing (). Cumulative species richness was also higher for charcoal production and cultivation than grazing ().
Figure 4. The cumulative species richness curves for the total woody species based on (a) sample plots and (b) number of individuals for all land uses combined (overall), and for the grazing, cultivation and charcoal production land-use types, within this multiple-use equatorial African savanna, central Uganda.
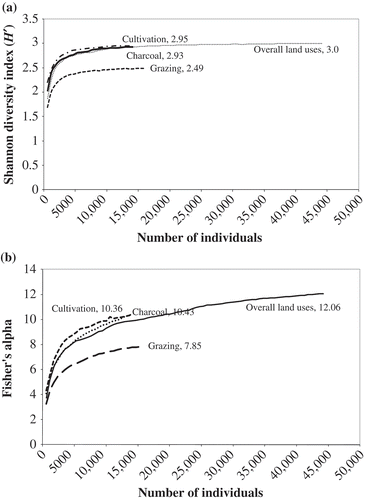
Across all the plots alpha diversity ranged from 0.34 to 6.34 and from 0.14 to 2.64 for Fisher's alpha (α) and Shannon–Wiener index (H′), respectively. However, plot values for both indices did not show a clear and uniform increase with increasing distance along the transect (i.e. first to last plot) radiating away from the households, the assumed source of human disturbance. Both H′ and alpha diversity were significantly different (H′: F 2,72 = 7.31, p < 0.001: α: F 2,72 = 7.29, p < 0.001) among land uses (). H′ was highest under cultivation and charcoal production and significantly lower under grazing. Cumulative H′ followed the same trend, being similar between cultivation (2.95) and charcoal production (2.93), with grazing considerably lower (2.49, ). Fisher's alpha diversity was also higher for cultivation and charcoal production, and significantly lower for grazing (). The cumulative alpha diversity was again higher under charcoal production (10.43) and cultivation (10.36), and lower under grazing (7.85, ). The overall trend is that species diversity was higher in the cultivation and charcoal production and lower in the grazing land use. The regression results showed insignificant weak correlations (with r 2 spanning 0.004–0.095; p > 0.1) between woody species diversity (i.e. richness and alpha diversity) and the ‘distance of the sampling plot along the transect radiating away from the households’ at both the whole savanna and land-use type scales. This suggests that the distance along the transect away from the households, the assumed source of woodland human disturbance, accounts for a very small fraction of the variation in woody species diversity within this multiple-use savanna.
Figure 5. The cumulative diversity curves plotted against number of individuals for (a) Shannon–Wiener diversity index and (b) Fisher's alpha diversity for the total woody plants for all land uses combined (overall), and for the grazing, cultivation and charcoal production land-use types, within this multiple-use equatorial African savanna, central Uganda.
Tree species richness and alpha diversity
Tree species richness differed among the land uses (F 2,72 = 16.6, p < 0.0001), being higher under cultivation and charcoal production and lower under grazing (). The most abundant tree species with high average number of plants per plot were Combretum collinum (147 plants/plot), Capparis fascicularis (120), Combretum molle (97), Combretum capituliflorum (82) and Combretum ghasalense (80), whereas the most frequent were C. collinum (in 63 of 75 plots), C. molle (60), C. ghasalense (59) and A. hockii (49). Acacia seyal, Commiphora africana and Commiphora dawei were only encountered under grazing, Albizia coriaria, Albizia malacophylla, Zanthoxylum chalybeum and Erythrina abyssinica in cultivation, while Pavetta crassipes, Dombeya dawei and Zanthoxylum rubescens were only recorded within charcoal production areas.
Table 4. Plot-level (0.1 ha) tree and shrub species diversity and richness (means ± SE) among the different land-use types in a multiple-use equatorial African savanna, central Uganda
Tree species H′ diversity at the plot level ranged from 0.0 to 2.6, and differed significantly among land uses (F 2,72 = 15.9, p < 0.0001), being higher in both cultivation and charcoal production compared with grazing (). The regression results showed insignificant weak correlations (with r 2 spanning 0.001–0.079; p > 0.18) between tree species diversity (i.e. richness and alpha diversity) and the ‘distance of the sampling plot along the transect radiating away from the source of human disturbance’ at both the whole savanna and land-use type scales. This suggests that the distance of sampling plots away from the households, the source of woodland disturbance, accounts for a very small fraction (<7.9%) of the variation in tree species diversity within this multiple-use savanna.
Shrub species richness and alpha diversity
Shrub species richness differed among land uses (F 2,72 = 8.28, p = 0.0006), being significantly higher under cultivation and charcoal production compared with grazing (). Overall, the most abundantly shared shrub species among land uses were Harrisonia abyssinica (26 plants/plot), Rhus natalensis (21), L. camara (20) and Annona senegalensis (18). The most frequent shrubs were R. natalensis (in 68 of 75 plots), H. abyssinica (65), Grewia mollis (53), Gardenia ternifolia (46) and A. senegalensis (43). Clerodendrum cordifolia, Flueggea virosa, Microglossa pyrifolia and Phyllanthus ovalifolius were recorded only in the grazing areas. Shrub H′ diversity ranged from 0.0 to 2.0, and differed among land uses (F 2,72 = 7.08, p = 0.002), being significantly higher for cultivation and charcoal production relative to grazing (). The regression results showed insignificant weak correlations (with r 2 spanning 0.002–0.141; p > 0.18) between shrub species diversity (i.e. richness and alpha diversity) and the ‘distance of the plot along the transect radiating away from the households’, the assumed source of human disturbance at both the whole savanna and land-use type scales. This suggests that the distance of sampling plots away from the human sources of woodland disturbance accounts for a very small fraction (<14.1%) of the variation in shrub species diversity within this multiple-use savanna.
Beta diversity
Plot-level pairwise Morisita–Horn (C MH) ranged 0–0.979 under grazing, 0.005–0.972 for charcoal production and 0.001–0.961 under cultivation. C MH tended to differ (F 2,900 = 2.43, p = 0.09) between land uses, with charcoal production and grazing being higher than cultivation. βW and βT were highest under grazing, followed by cultivation and lowest for charcoal production (). Hence, the βW, βT and C MH values show an overall higher beta diversity for the grazing land use compared with others. These results are corroborated by the very low within land-use SIMPER similarities, and very high between land-use dissimilarities. The within land-use type similarities were low (ranging 25.3–30.5%), being highest for charcoal production (mean = 30.5%), followed by grazing (25.4%), and then cultivation (25.3%). Furthermore, SIMPER analysis showed relatively high pairwise dissimilarities between land uses, ranging 73.2–81.2%. The highest mean dissimilarity was for cultivation versus grazing (81.2%), followed by charcoal versus grazing (76.7%) and lowest for charcoal versus cultivation (73.2%). These results are corroborated by lower similarity percentages within land uses.
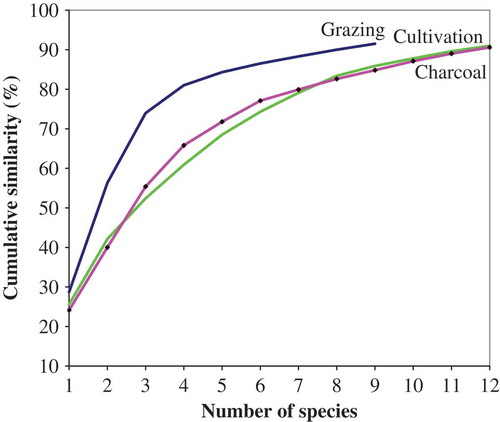
Indeed, SIMPER showed that the number of species making up 90% of the observed similarity per land use was 12 for both cultivation and charcoal production and 9 for grazing (Appendix 3, ). The top two species contributing most to the percentage similarity in community species composition within cultivation and charcoal production were C. collinum and C. molle. However, for grazing, the second species was Piliostigma thonningii (Appendix 3). SIMPER analysis indicated that 24 species between charcoal production and cultivation, 22 species between charcoal and grazing and 23 species between cultivation and grazing contributed most to the dissimilarity between land uses, reflecting the overall differences in community composition (Appendix 3).
Figure 6. Cumulative percentage contribution to overall similarity from ANOSIM within the three land-use types in relation to the number of species contributing to at least 90% similarity in savanna woodland of Nakasongola, central Uganda.
Discussion
Community composition and structure
Generally, C. collinum, C. fascicularis, C. molle, C. capituliflorum, C. ghasalense, G. mollis and A. hockii were the most abundant and frequent species across all land uses. This agrees with Nangendo et al. (Citation2006) that C. molle, G. mollis, A. senegalensis and Grewia bicolor are commonly found in burned areas and in open savanna woodlands in Uganda. Fires are common in these multiple-use savannas, and fire is used as a management tool in both subsistence crop cultivation and livestock grazing. About 20–50% of the grazing areas in the studied multiple-use savannas are burned towards the end of the two dry seasons before the onset of rainfall (in February and again in July) to allow fresh pastures and kill ticks. Similarly, Morris (Citation1995) observed that people in the miombo woodlands of Malawi burn the bush for preparing their gardens for planting, hunting and calling the rains. Since fires occur at more than one time of the year, some specific patches of land may be burnt more than once in a year. These numerous fires result in relatively low fire intensities because fuel does not accumulate to any large extent, and maintains much of the woodlands as a shrub transition. Fire is indeed an important factor for the maintenance of African savanna ecosystems (Bond and Van Wilgen Citation1996).
Over the last 30 years the woodland cover of Nakasongala District has decreased and is now mostly open woodland in transition to thicket/shrubland, while areas under cultivation and settlements have increased (Kalema Citation2010), mirroring patterns seen in other African savannas (Luoga et al. Citation2005; Mwavu and Witkowski Citation2008b; Coetzer et al. Citation2010).
The high density and number of Acacia spp. are probably facilitated by seed dispersal by ruminants such as the abundant cattle that graze in this multiple-use savanna. Certain Acacia spp. also form persistent seed banks (Witkowski and Garner Citation2000), with animal faeces also providing favourable germination beds (Schultka and Cornelius Citation1997). Seasonal bush burning may be a contributory factor to the high abundances of acacias in the studied savanna, since their germination is generally enhanced when subjected to the heat of fire (Mbalo and Witkowski Citation1997; Teketay Citation1997).
Some of the species encountered were restricted to particular land-use types, suggesting the influence of land-use practices in the distribution of woody plants. For example, the higher abundances of P. thonningii and A. seyal in the livestock grazing areas may be attributed to them not being palatable or preferred species by mammalian herbivores (Anderson et al. Citation2007), high resprouting ability and animal seed dispersal. On the other hand, the presence of Acacia mellifera, H. abyssinica, L. camara, R. natalensis and Carissa edulis, which are encroacher species in disturbed vegetation is not surprising (Van Vegten Citation1983). These species are neither browsed by herbivores nor harvested for charcoal production. Indeed, selective predation of preferred seedling species by herbivores has far-reaching consequences for plant community development (Hanley Citation1998).
Anthropogenic factors (charcoal production and shifting cultivation) have similarly been reported to be causative agents of degradation in the central Ethiopian highlands (Yirdaw and Luukkanen Citation2003). However, in this study, the differences among land uses may be the result of the extent (intensity and frequency) and duration of anthropogenic disturbances and particular land-use practices taking place in a given area. Changes in land-use practice may result in continuously or more abruptly deteriorating environmental conditions for some plant species, causing their decline in abundance and distribution. The chopping down of trees for charcoal production on a relatively short rotation has clearly resulted in local species loss. Some of the highly utilized species for charcoal production, firewood and poles for fences and houses are A. hockii, A. polyacantha, A. seyal, Acacia sieberiana, A. coriaria, Albizia zygia, C. capituliflorum, C. collinum, C. ghasalense, C. molle, G. mollis, Hymenocardia acida, Maytenus senegalensis, P. thonningii, Terminalia glaucescens and Vepris nobilis (Namaalwa et al. Citation2005; Kalema Citation2010). However, the most adversely affected species that were decreasing in abundance (Kalema Citation2010) are Albizia spp. and Terminalia spp. This is probably because they are highly preferred for poles, timber and charcoal production, and perhaps also due to their relatively poor regenerative capacities (few resprouting stumps and relatively low recruitment; Kalema Citation2010).
Species richness
Total woody plant species richness differed significantly among land uses with charcoal production and cultivation being higher than grazing. Differences in species richness between land uses may be the result of the different land use practices per se, or because of differences in the number of individuals counted (Gotelli and Colwell Citation2001). Charcoal production and cultivation were equally well sampled (24 plots each) and had an almost equal number of species (77 vs. 76), while grazing with 27 plots had only 57 species. However, the grazing lands include some hydromorphic grasslands, which typically have lower tree densities. Woody species richness in a Tanzanian miombo woodland also differed between shifting cultivation and more permanently cultivated areas (Luoga et al. Citation2005). The patchwork in the landscape after abandonment of the land provides opportunities for different functional groups of species without the disappearance of the original species (Luoga et al. Citation2004). Indeed, richness and diversity may increase with increasing heterogeneity within a range of patch sizes (Cabral et al. Citation2003). However, the ever-expanding shifting cultivation, leading to fragmentation of the remaining natural habitats may result in local extinction of small, isolated populations (Tilman et al. Citation2002). Thus, shifting cultivation can play either a positive or a negative role, depending on how it is managed. Variation in species richness between land uses may also be explained by a variety of other factors that include habitat quality, spatio-temporal dynamics, boundary characteristics and neighbourhood effects (e.g. Waldhardt and Otte Citation2003).
The overall sampled woody species richness of Nakasongola (99 species, 31 families within 75 plots with an area of 0.1 ha) is lower than the forest–woodland–savanna mosaic of northern Budongo Forest Reserve, north-west Uganda (121 species, 38 families within 594 plots with an area of 0.05 ha; Nangendo et al. Citation2006). Although both occur within the same ecological zone, the latter is a reserve, where human disturbances are restricted. Species richness of Nakasongola is also lower than the woodlands of Katavi–Rukwa ecosystem in western Tanzania (229 species, 45 families within 50 plots with an area of 0.1 ha; Banda et al. Citation2008), the Kitulangalo Forest Reserve and surrounding communal lands of Tanzania (133 species, 31 families within 64 plots with an area of 0.1 ha; Luoga et al. Citation2000b) and communal savanna woodlands in three South African provinces (135 species, 42 families within 90 plots with an area of 0.1 ha; Dovie et al. Citation2008). These comparisons suggest loss of species from Nakasongola through habitat degradation. The richness of only 2–31 species per 0.1 ha in Nakasongola is low relative to those reported elsewhere. However, it is similar to 1–28 species per 0.1 ha plot in the woodlands of western Tanzania (Banda et al. Citation2008).
Alpha diversity
Woody plant alpha diversity (H′ and Fisher's) did not generally increase with increasing distance along the transect away from households, the assumed source of disturbance. Indeed, regression analyses revealed that the distance of sampling plots away from households, the assumed source of human disturbance, to woodlands accounted for a very small fraction (<14.2%) of the variation in woody species diversity within this multiple-use savanna. This suggests that there is no clear gradient of increasing resource availability or decreasing human disturbance with increasing distance from homesteads or villages. Indeed, charcoal burning, a major source of disturbance to woody species, is not influenced by the distances from homesteads or villages in this region, but rather the availability of suitable and sizeable woody individuals for charcoal production. However, woody plant alpha diversity was significantly higher under cultivation and charcoal compared with grazing. Similarly, cumulative Fisher's alpha was higher under charcoal and cultivation than under grazing. These results suggest that species composition and distribution of plant communities in this savanna are to a great extent shaped by anthropogenic disturbances. On the other hand, these results are consistent with Hayek and Buzas' (Citation1997) suggestion that the more species that are present, and the more evenly the individuals are spread across the species, the higher the resulting H′. Fisher's alpha is low when the number of species is low. The low species diversity in the grazing lands may be a consequence of greater herbivory and extent of bush fires than in the other land uses. Both fires and herbivory may inhibit establishment of seedlings and recruitment of saplings (Helm et al. Citation2011), consequently affecting the population structure and regeneration status of the species. Tree density at a site may change rapidly with changes in the frequency or intensity of herbivory and fire (Bond and van Wilgen Citation1996). Such changes can be extensive, influencing vegetation dynamics at the landscape scale (Sinclair and Arcese Citation1995). In some places at Nakasongola where livestock grazing and termite damage were heavy, bare patches were common.
Compared with other African savannas, H′ diversity (0.41–2.64) in Nakasongola tended to be higher than southern Malawian miombo woodlands (0.55–1.26; Mwase et al. Citation2007), but lower than the Kitulangalo miombo woodland of Tanzania (2.9–3.13; Malimbwi et al. Citation1994) and South African communal savannas (2.5–3.9; Dovie et al. Citation2008).
Beta diversity
In spite of the low alpha diversity, the multiple-use savannas of Nakasongola have relatively high beta diversity. The relatively high values for all the beta diversity indices and low similarities between plots and land-use types are indicators of gradients in the species composition of these savanna woodlands. The general absence of significant differences in species composition between the charcoal production and cultivated areas also suggests the influence of an environmental gradient rather than differences between the two land-use types. The lack of differences is probably due to the cyclic sequence of land use over time. The high contribution of C. collinum, C. molle and C. ghasalense to the similarity in species composition among plots under the charcoal production land use, despite them being the most utilized species for charcoal production, shows their ability to recover from disturbance and to occupy a wide variety of habitats. These results generally point to influence of an environmental gradient, in addition to anthropogenic disturbance, in the variation of species composition within this multiple-use savanna. Species composition and distribution of plant communities are shaped not only by anthropogenic disturbances but also by environmental conditions, spatial factors and species competition (Cousins and Eriksson Citation2002). Among these is below ground availability of soil nutrients that plays an important role in the assembly of tropical tree communities at local scales (John et al. Citation2007). Other broader scale studies have illustrated how historical or climatic factors may also act to constrain diversity (e.g. Kleidon and Mooney Citation2000; Sarr et al. Citation2005).
Within these savannas, natural and anthropogenic disturbances from subsistence harvesting of poles and commercial charcoal production are scattered in space and time, contributing to habitat heterogeneity. This habitat heterogeneity may favour the coexistence of species with different ecological requirements, thus contributing to the maintenance of community diversity (Mwavu and Witkowski Citation2009) and variation in species composition among the plots along a transect. Anthropogenic land-use change is one of the most important factors contributing to global change (Sala et al. Citation2000), and changes in landscape structure and agricultural land-use intensity are likely to influence plant community composition and similarity. The studied multiple-use savanna is characterized by anthropogenic disturbances which have been patchy and have different influences on community species composition. Such intrinsic disturbances may lead to abrupt changes in habitat, consequently greatly contributing to heterogeneity in this savanna environment and result in reversal or delay of successional states. The high beta diversity under grazing compared with other land uses in this savanna may be attributed to the more heterogeneous spatial distribution, and varying levels and intensities of grazing (stocking rates between villages) that contribute to greater environmental heterogeneity. Livestock grazing impacts are typically not uniform across a landscape because herbivores are highly selective of areas with higher soil fertility (soil type), new grass growth from burnt areas and particular landscape positions (Senft et al. Citation1987; O'Connor et al. Citation2011). Abrupt changes in site conditions that are accompanied by changes in the physical and chemical nature of soil nutrient dynamics and microclimate may influence plant community composition and species diversity patterns, particularly beta diversity (Pitman et al. Citation1999; Grace Citation2001) as was observed in this study. Indeed, beta diversity interacts with species richness gradients and both are the outcome of the assembly of communities through local and regional filters (Soininen et al. Citation2007).
Conclusions
Woody plant density, species richness and diversity were significantly higher under both cultivation and charcoal production relative to grazing. These different land uses have altered natural disturbance patterns, resulting in changes in species abundances and distributions, community composition and ecosystem function. The relatively low woody species alpha diversity and richness in the multiple-use savanna woodlands of Nakasongola indicates woodland degradation, fragmentation and local species loss; a result of unsustainable woody plant harvesting for charcoal production, and possibly the relatively short-term shifting cultivation cycle. Over the long term, charcoal production and grazing (including associated burning practices) have resulted in loss of tree cover, and probably reductions in woody plant species richness and diversity. More sustainable harvesting practices are needed in order to halt and reverse the loss of tree cover and diversity.
Securing the long-term persistence of plant diversity and richness in these savanna woodlands requires appropriate policies to guide changes in land-use practices that accommodate the requirements of land users and conservation efforts. However, current market forces, such as the increasing demand for charcoal and firewood, are likely to result in a rapid attrition of this natural asset if effective management is not implemented. Degraded woodlands in response to these forces are typical of many regions that supply fuelwood to East African urban areas. Together with this understanding of diversity patterns in relation to anthropogenic factors in the woodlands, a detailed study of the influence of other environmental factors, particularly topo-edaphic variability, would provide a more complete understanding of species distribution patterns.
Acknowledgements
This study was financially supported by the Third World Organization for Women in Science (TWOWS), the South African National Research Foundation (NRF 2069152), the Andrew Mellon Postgraduate Mentorship Award and the Post Graduate Merit Award from the University of the Witwatersrand, Johannesburg. We thank Vivienne Williams for guidance with the EstimateS analyses. We thank Edward Mwavu for valuable comments on the manuscript. We thank Nyesiga Grace, Lunkuraite Alex, Mukasa Ivan and Sembatya Tadeo for their assistance with field data collection, our driver Robert, the District Resource, Environment and Livestock Officers in Nakasongola, all chiefs of subcounties and all those in one way or another who helped to make this study a success.
Related Research Data
References
- Anderson , TM , Ritchie , ME and Mc Naughton , SJ. 2007 . Rainfall and soils modify plant community response to grazing in Serengeti National Park . Ecology , 88 ( 5 ) : 1191 – 1201 .
- Banda , T , Mwangulango , N , Meyer , B , Schwartz , MK , Mbago , F , Sungula , M and Caro , T. 2008 . The woodland vegetation of Katavi-Rukwa ecosystem in western Tanzania . For Ecol Manage , 255 ( 8–9 ) : 3382 – 3395 .
- Banda , T , Schwartz , MW and Caro , T. 2006 . Woody vegetation and composition along a protection gradient in a miombo ecosystem of western Tanzania . For Ecol Manage , 230 ( 1–3 ) : 179 – 185 .
- Beater , MMT , Garner , RD and Witkowski , ETF. 2008 . Impacts of clearing invasive alien plants from 1995 to 2005 on vegetation structure, invasion intensity and ground cover in a temperate to subtropical riparian ecosystem . S Afr J Bot , 74 ( 3 ) : 495 – 507 .
- Bond , WJ and van Wilgen , BW. 1996 . Fire and plants , 38 – 42 . London , UK : Chapman and Hall .
- Braedt , O and Stranda-Gunda , W. 2000 . Woodcraft markets in Zimbabwe . Int Tree Crops J , 10 : 367 – 384 .
- Cabral , AC , De Miguel , JM , Rescia , AJ , Schmitz , MF and Pineda , FD. 2003 . Shrub encroachment in Argentinean savannas . J Veg Sci , 14 ( 2 ) : 145 – 152 .
- Cadotte , MW , France , R , Reza , L and Lovett-Doust , J. 2002 . Tree and shrub diversity and abundance in fragmented littoral forest of south-eastern Madagascar . Biodivers Conserv , 11 ( 8 ) : 1417 – 1436 .
- Coetzer , KL , Erasmus , BFN , Witkowski , ETF and Bachoo , AK. 2010 . Land-cover change in the Kruger to Canyons Biosphere Reserve (1993–2006): a first step towards creating a conservation plan for the subregion . S Afr J Sci , 106 ( 7/8 ) : 26 – 35 .
- Colwell RK. 2006. EstimateS: statistical estimation of species richness and shared species from samples, version 8.0 [Internet]. [cited 2008 Jun 15]. purl.oclc.org/estimates (http://purl.oclc.org/estimates)
- Colwell , RK , Mao , CX and Chang , J. 2004 . Interpolating, extrapolating and comparing incidence-based species accumulation curves . Ecology , 85 ( 10 ) : 2717 – 2727 .
- Cousins , SAO and Eriksson , O. 2002 . The influence of management history and habitat on plant species richness in a rural hemiboreal landscape, Sweden . Landscape Ecol , 17 ( 6 ) : 517 – 529 .
- Cowling , RM and Witkowski , ETF. 1994 . Convergence and non-convergence of plant traits in climatically and edaphically matched sites in Mediterranean Australia and South Africa . Aust J Ecol , 19 ( 2 ) : 220 – 232 .
- Dovie , DBK , Shackleton , CM and Witkowski , ETF. 2002 . Direct-use values of woodland resources consumed and traded in a South African village . Int J Sustain Dev World Ecol , 9 ( 3 ) : 269 – 283 .
- Dovie , DBK , Witkowski , ETF and Shackleton , CM. 2005 . Monetary valuation of livelihoods for understanding the composition and complexity of rural households . Agric Human Values , 22 ( 1 ) : 87 – 103 .
- Dovie , DBK , Witkowski , ETF and Shackleton , CM. 2008 . Knowledge of plant resource use based on location, gender and generation . Appl Geogr , 28 ( 4 ) : 311 – 322 .
- Eilu , G , Hafashimana , DLN and Kasenene , JM. 2004 . Density and species diversity of trees in four tropical forest of the Albertine Rift, Western Uganda . Divers Distr , 10 ( 4 ) : 303 – 312 .
- Fisher , JT , Witkowski , ETF , Erasmus , BFN , Van Aardt , J , Asner , GP , Wessels , K and Mathieu , R. 2011 . Human-modified landscapes: patterns of fine-scale woody vegetation structure in communal savannah rangelands . Environ Conserv , 39 ( 1 ) : 72 – 82 .
- Folke , C , Carpenter , S , Walker , B , Scheffer , M , Elmqvist , T , Gunderson , L and Holling , CS. 2004 . Regime shifts, resilience, and biodiversity in ecosystem management . Annu Rev Ecol Syst , 35 : 557 – 581 .
- Gotelli , JN and Colwell , RK. 2001 . Quantifying biodiversity: procedures and pitfalls in the measurement and comparison of species richness . Ecol Lett , 4 ( 4 ) : 379 – 391 .
- Grace , JB. 2001 . The roles of community biomass and species pools in the regulation of plant diversity . Oikos , 92 ( 2 ) : 193 – 207 .
- Hanley , ME. 1998 . Seedling herbivory, community composition and plant life history traits: perspectives in plant ecology . Evol Syst , 1 ( 2 ) : 191 – 205 .
- Hayek , LC and Buzas , MA. 1997 . Surveying natural populations , New York , NY : Columbia University Press .
- Helm , CV , Wilson , G , Midgley , JJ , Kruger , L and Witkowski , ETF. 2011 . Investigating the vulnerability of an African savanna tree (Sclerocarya birrea subsp. caffra) to fire and herbivory . Aust Ecol , 36 ( 8 ) : 964 – 973 .
- John , R , Dalling , JW , Harms , KE , Yavilt , JB , Stallard , RF , Mirabello , M Hubbell , SP . 2007 . Soil nutrients influence spatial distributions of tropical tree species . PNAS , 104 ( 3 ) : 864 – 869 .
- Kalema , VN. 2010 . Diversity, use and resilience of woody species in a multiple land use Equatorial African Savanna, Central Uganda [PhD thesis]. [Johannesburg (South Africa)] , Faculty of Science, University of the Witwatersrand .
- Katende , AB , Birnie , A and Tengnas , B. 1995 . Useful trees and shrubs for Uganda. Identification, propagation and management for agricultural and pastoral communities , Nairobi (Kenya) : Regional Soil Conservation Unit, RSCU/SIDA . Technical Handbook No. 10
- Kituyi , E. 2004 . Towards sustainable production and use of charcoal in Kenya: exploring the potential in life cycle management approach . J Cleaner Prod , 12 ( 8–10 ) : 1047 – 1057 .
- Kleidon , A and Mooney , HA. 2000 . A global distribution of biodiversity inferred from climatic constraints: results from a process-based modelling study . Glob Change Biol , 6 ( 5 ) : 507 – 523 .
- Langdale-Brown , I , Osmaston , HA and Wilson , JG. 1964 . The vegetation of Uganda and its bearing to land-use , Entebbe (Uganda) : Government Printers .
- Luoga , EJ , Witkowski , ETF and Balkwill , K. 2000a . Economic of charcoal production in miombo woodlands of eastern Tanzania: some hidden costs associated with commercialization of the resources . Ecol Econ , 35 ( 2 ) : 243 – 257 .
- Luoga , EJ , Witkowski , ETF and Balkwill , K. 2000b . Differential utilisation and ethnobotany of trees in Kitulanghalo Forest Reserve and surrounding communal lands, eastern Tanzania . Econ Bot , 54 ( 3 ) : 328 – 343 .
- Luoga , EJ , Witkowski , ETF and Balkwill , K. 2002 . Harvested and standing wood stocks in protected and communal miombo woodlands of eastern Tanzania . For Ecol Manage , 164 ( 1–3 ) : 15 – 30 .
- Luoga , EJ , Witkowski , ETF and Balkwill , K. 2004 . Regeneration by coppicing (resprouting) of miombo (African savanna) trees in relation to land-use . For Ecol Manage , 189 ( 1–3 ) : 23 – 35 .
- Luoga , EJ , Witkowski , ETF and Balkwill , K. 2005 . Land-cover and use changes in relation to the institutional framework and tenure of land and resources in eastern Tanzania miombo woodlands . Environ Dev Sustain , 7 ( 1 ) : 71 – 93 .
- Magurran , AE. 2004 . Measuring biological diversity , 1 – 256 . Oxford (UK) : Blackwell Science Ltd .
- Malimbwi , RE , Misana , S , Monela , GC , Jambiya , G and Zahabu , E. 1994 . Impact of charcoal extraction to the forest resources of Tanzania , Sabadell (Spain) : Charcoal Potential in Southern Africa (CHAPOSA) .
- Mbalo , BA and Witkowski , ETF. 1997 . Tolerance to soil temperatures experienced during and after the passage of fire in seed of Acacia karoo, Acacia nilotica and Chromolaena odorata: a laboratory study . S Afr J Bot , 63 ( 6 ) : 421 – 425 .
- Millennium Ecosystem Assessment . 2005a . Ecosystem and human well-being: biodiversity synthesis , Washington , DC : Island Press .
- Millennium Ecosystem Assessment . 2005b . Ecosystem and human well-being: desertification synthesis , Washington (DC) : Island Press .
- Morris , B. 1995 . Woodland and village: reflections on the ‘animal estate’ in rural Malawi . J R Anthropol Inst (NS) , 1 : 301 – 315 .
- Mwase , WF , Bjornstand , A , Bokosi , JM , Kwapata , MB and Stedje , B. 2007 . The role of land tenure in conservation of tree and shrub species diversity in miombo woodlands of southern Malawi . New For , 33 ( 3 ) : 297 – 307 .
- Mwavu , EN and Witkowski , ETF. 2008a . Sprouting of woody species following cutting and tree-fall in a lowland semi-deciduous tropical rainforest, North-Western Uganda . For Ecol Manage , 255 ( 3 ) : 982 – 992 .
- Mwavu , EN and Witkowski , ETF. 2008b . Land-use and cover changes (1988–2002), around Budongo Forest Reserve, NW Uganda: implications for forest and woodland sustainability . Land Degrad Dev , 19 ( 6 ) : 606 – 622 .
- Mwavu , EN and Witkowski , ETF. 2009 . Seedling regeneration, environment and management in a semi-deciduous African tropical rain forest . J Veg Sci , 20 ( 5 ) : 791 – 804 .
- Nakasongola Local Government . 2002 . District environment profile: Nakasongola District Forest Development plan 2003–2014 , 1 – 79 . Nakasongola (Uganda) : District Forest Services .
- Namaalwa , J , Eid , T and Sankhayan , P. 2005 . A multi-species density-dependent matrix growth model for the dry woodlands of Uganda . For Ecol Manage , 213 ( 1–3 ) : 312 – 327 .
- Nangendo , G , Steege , HT and Bongers , F. 2006 . Composition of woody species in a dynamic forest-woodland-savannah mosaic in Uganda: implications for conservation and management . Biodivers Conserv , 15 ( 4 ) : 1467 – 1495 .
- National Environment Management Authority . 2002 . State of environment report for Uganda , Kampala , , Uganda : Ministry of Environment and Natural Resources .
- Natta , AK , Sinsin , B and van Der Maesen , LJG. 2002 . Riparian forests, a unique but endangered ecosystem of Benin . Bot Jahrb Syst , 124 ( 1 ) : 55 – 69 .
- O'Connor , TG , Martindale , G , Morris , CD , Short , A , Witkowski , ETF and Scott-Shaw , R. 2011 . Influence of grazing management on plant diversity of highland Sourveld Grassland, KwaZulu-Natal, South Africa . Rangeland Ecol Manage , 64 ( 2 ) : 196 – 207 .
- Parker , D , Downer , ER and Cole , GED. 1967 . Atlas of Uganda , Entebbe (Uganda) : Department of Lands and Survey, Government Printers .
- Pitman , NCA , Terborgh , J , Silman , MR and Nuñez , PV. 1999 . Tree species distributions in the upper Amazonian forest . Ecology , 80 ( 8 ) : 2651 – 2661 .
- Polhill , RM , ed. 1952 . Flora of tropical east Africa (FTEA) , London (UK) : Royal Botanic Gardens .
- Pomeroy , D , Tushabe , H , Mwima , P and Kasoma , P. 2002 . Uganda ecosystem and protected area characterisation: a contribution to Strategic Criteria for Rural Investments in Productivity (SCRIP). Program of the USAID Uganda Mission , 1 – 55 . Washington (DC) : USAID .
- Rennolls , K and Laumonier , Y. 2000 . Species diversity structure analysis at two sites in tropical rain forest of Sumatra . J Trop Ecol , 16 ( 2 ) : 253 – 270 .
- Sala , OE , Chapin , FS , Armesto , JJ , Berlow , E , Bloomfeld , J , Dirzo , R , Huber-Sanwald , E , Huenneke , LF , Jackson , RB Kinzig , A . 2000 . Global biodiversity scenarios for the year 2100 . Science , 287 ( 5459 ) : 1770 – 1774 .
- Sarr , DA , Hibbs , DE and Huston , MA. 2005 . A hierarchical perspective of plant diversity . Q Rev Biol , 80 ( 2 ) : 187 – 212 .
- Schultka , W and Cornelius , R. 1997 . Vegetation structure of a heavily grazed range in northern Kenya: tree and shrub canopy . J Arid Environ , 36 ( 2 ) : 291 – 306 .
- Seaby , RMH and Henderson , PA. 2007 . Species diversity and Richness® version IV (SDR 4.0) , Lymington (UK) : Pisces Conservation Ltd .
- Seaby , RMH , Henderson , PA and Prendergast , JR. 2007 . Community analysis package® version 4 (CAP 4) , Lymington (UK) : Pisces Conservation Ltd .
- Senft , RL , Coughenour , MB , Bailey , DW , Rittenhouse , LR , Sala , OE and Swift , DM. 1987 . Large herbivore foraging and ecological hierarchies: landscape ecology can enhance traditional foraging theory . Bioscience , 37 ( 11 ) : 789 – 799 .
- Sinclair , ARE and Arcese , P , eds. 1995 . Serengeti II: dynamics, management and conservation of an ecosystem , Chicago (IL) : University of Chicago Press .
- Soininen , J , Lennon , JJ and Hillebrand , H. 2007 . A multivariate analysis of beta-diversity across organisms and environments . Ecology , 88 ( 11 ) : 2830 – 2838 .
- Teketay , D. 1997 . Germination ecology of Acacia negrii, an endemic multipurpose tree from Ethiopia . Trop Ecol , 38 ( 1 ) : 39 – 46 .
- Tilman , D , Cossman , KG , Matson , PA , Naylor , R and Polasky , S. 2002 . Agricultural sustainability and intensive production practices . Nature , 418 : 671 – 677 .
- Uganda Bureau of Statistics . 2002 . Uganda population and housing census , Kampala (Uganda) : Uganda Bureau of Statistics .
- USDA Forest Service . 1989 . Interim resource inventory glossary , Washington (DC) : Department of Agriculture, Forest Service .
- Van Vegten , JA. 1983 . Thornbush invasion in a savanna ecosystem in eastern Botswana . Vegetatio , 56 ( 1 ) : 3 – 7 .
- Venter , SM and Witkowski , ETF. 2011 . Baobab (Adansonia digitata L.) fruit production in communal and conservation land-use types in Southern Africa . For Ecol Manage , 261 ( 3 ) : 630 – 639 .
- Waldhardt , R and Otte , A. 2003 . Indicators of plant species and community diversity in grasslands . Agric Ecosyst Environ , 98 ( 1–3 ) : 339 – 351 .
- White , F. 1983 . The vegetation of Africa. A descriptive memoir to accompany the UNESCO/AETFAET/UNSO vegetation map of Africa , Paris (France) : UNESCO .
- Williams , VL , Witkowski , ETF and Balkwill , K. 2005 . Application of diversity indices to appraise plant availability in the traditional medicinal markets of Johannesburg, South Africa . Biodivers Conserv , 14 ( 12 ) : 2971 – 3001 .
- Witkowski , ETF and Garner , RD. 2000 . Spatial distribution of soil seed banks of three African savanna woody species at contrasting sites . Plant Ecol , 149 ( 1 ) : 91 – 106 .
- Witkowski , ETF and Garner , RD. 2008 . Seed production, seed bank dynamics, resprouting and long-term response to clearing of the alien invasive Solanum mauritianum in a temperate to subtropical riparian ecosystem . S Afr J Bot , 74 ( 3 ) : 476 – 484 .
- Yirdaw , E and Luukkanen , O. 2003 . Indigenous woody species diversity in Eucalyptus globules Labill. ssp globules plantations in the Ethiopian highlands. Biodivers . Conserv , 12 ( 3 ) : 567 – 582 .
- Zapfack , L , Engwald , S , Sonke , B , Achoundong , G and Madong , BA. 2002 . The impact of land conversion on plant biodiversity in the forest zone of Cameroon . Biodivers Conserv , 11 ( 11 ) : 2047 – 2061 .
Appendix 1. Species list arranged alphabetically by families for woody plants recorded in 75 plots (0.1 ha) within the savanna woodland of Nakasongola District, central Uganda
Appendix 2. Contribution of individual species to the overall similarity within land-use types (grazing, cultivation and charcoal production)
Appendix 3. Results of SIMPER analysis highlighting the species contributing most to the dissimilarity between land-use types