
Abstract
Fertilizer application plays a crucial role in the decomposition of white poplar leaf litter and cycling of nutrients within forest ecosystems. The impact of various fertilizer additions on white poplar leaf litter and nutrient cycling is poorly understood. In this study, seven treatments were conducted at the following levels: Control (CK), no adding mineral fertilizers, N fertilization (+N), N and P fertilization (+NP), N, P, K fertilization (+NPK), P, K (+PK), manure fertilizer (+MF), and bird fertilizers (+BF) in a white poplar plantation in Qadis district, and used the litterbag techniques to measure litter mass remaining. The main objectives of our study were: (1) to explore the response of white poplar leaf litter decomposition to various fertilizers and accelerate the decomposition process; (2) to examine the relationship between C, N, and P concentration and their stoichiometric characteristic in leaf litter and soil. In this investigation, our results showed that white poplar leaf litter was significantly affected by fertilizers, and the decomposition process was greatly accelerated with + MF, +NPK, and + BF. The decay rate constant k (year −1) shows the decomposition rate of white poplar leaf litter as follows: +MF > +NPK > +BF > +PK > +NP > +N > CK (0.56, 0.53, 0.52, 0.51,0.51,0.5, and 0.46). Soil nutrients N, P and K increased significantly during the decomposition time with + MF, +NPK, and + BF, respectively, while C:N, C:P, and N:P ratios were highest in the white poplar leaf litter, and lowest in soil, we observed significant association between nutrients concentrations in soil and white poplar leaf and their stoichiometric. This current study concluded that adding + MF, +NPK and + BF fertilizers might be the preferred management option as they provided potentially beneficial changes in leaf litter decomposition and increased nutrient concentration. The data obtained will be a valuable reference for fertilization management strategies in forest ecosystems.
1. Introduction
Leaf litter decomposition is a critical ecological process that supports nutrient cycling, soil health, carbon sequestration, and ecosystems overall functioning and sustainability (Li et al., Citation2023; Su et al., Citation2023; Wu et al., Citation2023) and is an essential process that regulates the cycling of nutrients and carbon in most terrestrial ecosystems (Allen et al., Citation2020; Cai et al., Citation2021; Liu et al., Citation2022). Accelerating leaf decomposition with fertilizers promotes rapid nutrient cycling in the ecosystem, and leaf litter contains valuable nutrients. Accelerating the decomposition of leaf litter releases nutrients into the soil and helps maintain soil fertility, which is important for the overall health and productivity of the forest (Hu et al., Citation2021; Song et al., Citation2020). The rate of decomposition and patterns of nutrient release depends on abiotic factors such as climate conditions and soil physical and chemical properties and biotic factors like litter quality and microbial community composition (Fanin et al., Citation2020; Liu et al., Citation2023; Shao et al., Citation2023; Teague and Dowhower, Citation2022). At the global scale, the decomposition of plant leaf litter is mostly influenced by both climate and litter quality (Akoto et al., Citation2022; Ouyang et al., Citation2022; Pang et al., Citation2022; Swart et al., Citation2022); on a local scale, it is influenced by the chemical properties of the leaf litter and the soil conditions of the site (Ren et al., Citation2018; Song et al., Citation2016; Xie et al., Citation2016).
Several studies in different ecosystems have been conducted to explore the effects of fertilizers on leaf litter decomposition (Su et al., Citation2021; Zhang et al., Citation2020; Zhu et al., Citation2016). However, the reactions of leaf litter decomposition to fertilizer addition have varied significantly, ranging from positive outcomes (Gurung et al., Citation2023; Wachendorf et al., Citation2020; Wu et al., Citation2023) to neutral (Dong et al., Citation2020; Khalsa et al., Citation2020; Nakatsuka et al., Citation2020) to negative (Cornwell et al., Citation2008; Micks et al., Citation2004; Zhou et al., Citation2017). Several chemical, biological, and biochemical hypotheses have been suggested to elucidate the potential mechanisms behind the diverse impact of N deposition on litter decomposition. Despite these efforts, our understanding of the effects of different fertilizers on leaf litter decomposition remains limited (Carreiro et al., Citation2000; Knorr et al., Citation2005; Liu et al., Citation2010; Sinsabaugh, Citation2010).
In the Qadis district, white poplar is one of the highly-valued species and fast-growing species capable of reaching an age of 35-40 years (Brilli et al., Citation2014). It is distributed widely with broad adaptability, provides a large volume of timber with many applications, and has significant ecological and economic value. In white poplar plantations, management of soil fertility through fertilizer application is needed to inhibit stand retrogression and sustain plantation productivity, but the effect of various fertilizers addition on the leaf litter of white poplar during the decomposition practice has not been evaluated.
In plantations with fast-growing tree species, leaf litter plays an important role in the nutrient cycle, providing a high proportion of mineral nutrients to the soil (González et al., Citation2020). Soil fertility and nutrient cycling is one of the main concerns for sustainable development (Gruhn et al., Citation2000). It is estimated that for common deciduous trees, in mature plantations, about 88% of N, 83% of P, and 78% of K are restored through leaf echoes (Das and Chaturvedi, Citation2005). However, other authors have reported a low reduction rate of 20-40%, suggesting that this ratio is largely dependent on genotype (Salehi et al., Citation2013). Meiresonne et al. (Citation2007) suggested that, in general, nutrient cycling in poplar stands is efficient with no significant nutrient loss, and that the circulars through the leaf circulars are generally rich in basic cations (Stark et al., Citation2015). White poplar communities, usually grown in full sun, tolerate wet and nutrient-poor sites (Griu and Lunguleasa, Citation2016), which are often considered quite sensitive to environmental changes such as nutrient deposition and flooding (Zhang et al., Citation2016). There have been limited reports regarding the influence of fertilizer application on leaf litter decomposition in the white poplar ecosystem. The insufficient investigation of this topic significantly restricts the precise evaluation of soil C sequestration during elevated nutrient deposition, resulting in an inadequate comprehension of nutrient cycling within the white poplar ecosystem. To address these knowledge gaps, we evaluate the effect of various fertilizer additions on white poplar leaf litter during the decomposition of leaf litter in the white poplar plantation; we collected leaf litter and soil samples from fertilizers treatments at four sampling times in white poplar plantation to determine mass remaining and nutrients concentration through the decomposition procedure. The purpose of our study was (1) to explore the response of white poplar leaf litter to various fertilizers and accelerate the decomposition process and (2) to examine the association between C, N, P and K concentration and between their stoichiometric characters under different fertilization treatments. We aimed to assess the following hypotheses: (1) fertilizers may accelerate the decomposition process; (2) carbon, nitrogen, phosphorus and potassium concentration may have a great relationship with their stoichiometric under fertilizers treatments.
2. Materials and methods
2.1. Study area and experimental setup
This research was conducted in a white poplar plantation with 6-years stand age in the Qadis district (34° 43′ 26.4" N, 63° 33′ 12.4" E), which is located in the southern part of Badghis province, Afghanistan (), which has an arid continental climate characterized by cold winters, relatively wet winters (including a peak in spring) and hot summers, with an average annual rainfall around 300 mm, and with an average annual temperature of 24 °C.
Figure 1. Map of the study area showing the location of Qadis district, in Badghis province, Afghanistan.
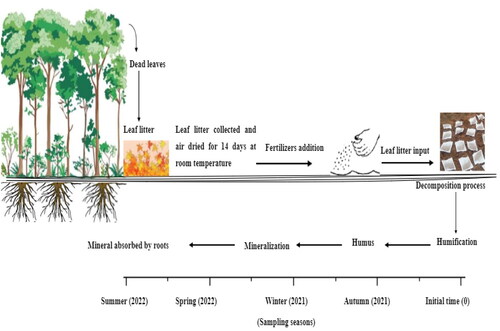
In this experiment, leaf litter that had fallen at the beginning of autumn 2021 was collected from the white poplar plantation; after air drying for 14 days at room temperature, 10 g leaf litter samples were employed in nylon mesh zip bags (200 × 200 mm with 1mm mesh) (). We established seven treatments with three replicate plots (5 × 5 m) at the following levels: Control (CK), with no addition of mineral fertilizers; N fertilization (+N) treatment received 150 kg ha−1 y −1 of urea (CO (NH2)2); N and P fertilization (+NP) treatment received 150 kg ha−1 y −1 of urea (CO (NH2)2), and phosphorus pentoxide (P2O5); N, P, K fertilization (+NPK) treatment received 150 kg ha−1 y −1 of urea (CO (NH2)2), phosphorus pentoxide (P2O5), and potassium oxide (K2O); P, K (+PK) treatment received 150 kg ha−1 y −1 of phosphorus pentoxide (P2O5), and potassium oxide (K2O); manure fertilizer (+MF) treatment received 150 kg ha−1 y −1 of manure, and bird fertilizer (+BF) treatment received 150 kg ha−1 y −1 chicken manure. The fertilizer concentrations used were 46% urea, 46% triple superphosphate, and 50% potassium sulfate, respectively, and the synthetic fertilizer was applied in the summer of 2021. The single litter bags with small pores were then arbitrarily placed on each plot in all litter bags, 252 bags in total (7 treatments × 3 plots × 3 replicates × 4 sampling times). In autumn, winter 2021, spring and summer 2022, leaf litter bags and soil samples were collected from each treatment. At each sampling time three leaf litter bags were randomly removed from each treatment. After removing the roots and residues of other plants from the samples, the samples were dried to a constant mass in an oven at 65 ◦C for 48 h. The dry mass data was recorded and then the litter was crushed according to the requirements for measuring its chemical properties. Soil samples were collected under the decomposition bag, specifically, from the 5-10 cm depth, and transported to the laboratory. In the laboratory soil samples air dried, all soil samples were crushed and sieved (2 mm) to eliminate debris and stone material before analyzing of the soil’s chemical properties.
Figure 2. Diagram showing the experimental process in white poplar forest in Qadis district
2.2. Determination of soil physical and chemical properties and chemical properties of leaf litter
The physical properties of the soil were determined using samples of the undisturbed soil. Kopeck steel cylinders (VS = 100 cm3) were used to take samples from a depth 5-10 cm. Each cylinder (labeled with a unique number) was weighed empty and uncapped and its mass (GV) was noted before sampling. According to (Cai et al., Citation2021), undisturbed soil samples were used to calculate soil moisture (%), soil bulk density (g/cm3), capillary porosity (%) and non-capillary porosity (%). The pH value of the soil was measured in water using a glass electrode (Mettler-Toledo, Greifensee, Switzerland) and soil organic carbon (SOC) and carbon in leaf litter were determined by the heating method (potassium dichromate digestion: (K2Cr2O7). Total nitrogen (TN) and total phosphorus (TP) in soil and leaf litter were determined using an AUTCHEM 1200 (Changchun Xingrui, China) analyzer and total soil, and leaf litter K were determined using a flame emission method (Xiang Yi, Hunan, China)
2.3. Data analysis
A single negative exponential decay model was employed to calculate the decomposition rates of leaf litter (y/y0 = e-kt). One-way analysis of variance (ANOVA) was used to compare the difference between mean chemical properties of soil and leaf litter values as implemented with the (IBM SPSS Statistics for Windows, Version 22. IBM Corp, Armonk, NY, USA), the spearmen correlation function in Origin Pro 2022 was used to examine the possible association between nutrients concentration in leaf litter, soil, and their stoichiometric.
3. Results
3.1. Response of leaf litter decomposition rate to fertilizer applications
At the end of this investigation, there was a significant difference between control plots and the other treatments. The remaining mass, from highest to lowest, was as follows: CK (57.73 ± 0.02) %, +N (52.73 ± 0.01) %, +NP (48.63 ± 0.02) %, +PK (47.74 ± 0.01) %, +BF (46.24 ± 0.05) %, +NPK (44.73 ± 0.01) %, and + MF (40.37 ± 0.03) %, respectively. The results from revealed that organic and inorganic fertilizer applications significantly increased the leaf litter mass loss, except for nitrogen plots (+N). The leaf litter was primarily much more decomposed in + MF, +NPK, and + BF, respectively (Figure3). The mass loss of white poplar leaf litter over time fitted an exponential equation for CK, +N, +NP, +NPK + PK, +MF, and + BF (R2 = 0.9, 0.91,0.94,0.97,0.96,0.98,0.96). The decomposition constant K differed among the CK, +N, +NP, +NPK, +PK, +MF, and + BF, with the highest in + MF and the lowest in the CK. The litter decay rate constant (k) followed the order + MF > + NPK > +BF > +PK > +NP > +N > CK (0.56, 0.53, 0.52, 0.51, 0.51,0.5, and 0.46) per year ().
Figure 3. Effect of various fertilizers on leaf litter decompositions at different sampling times. Values are Mean ± SE.
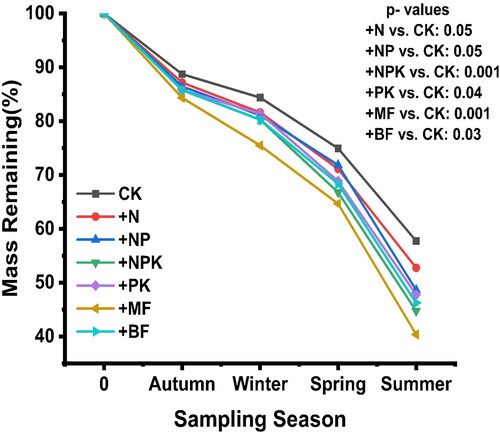
Table 1. Leaf litter decomposition constant, correlation coefficient, at T50% and T95% decomposition of White poplar leaf litter under various fertilizers. All relationship was significant (***p < 0.001)
3.2. Dynamics of C, N, P, and K concentrations in leaf litter under various fertilizer applications
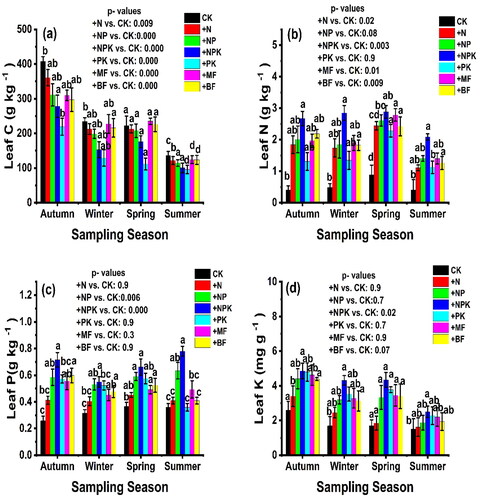
In this study, we examined the dynamics of C, N, P, and K concentrations during the decomposition process. The interaction between fertilizer applications and sampling time resulted in noticeable changes in C, N, P and K concentrations. Our findings indicate a significant decrease in carbon concentration after the first sampling time across all stages of the experiment, with the following order: CK > +N > +NP > +NPK > +PK > +MF > +BF, except for the spring season (p < 0.05) (), The N concentration in white poplar exhibited a gradual increase throughout all stages of the experiments, except for the last sampling stage. However, the concentration varied among the different fertilizer treatments in different seasons. In autumn and winter 2021, the nitrogen concentration in white poplar during the decomposition process followed the order: +NPK > +BF > +NP > +MF > +PK > CK and + NPK > +MF > +BF > +NP > +N > +PK > CK, respectively (). In contrast, the N concentration in white poplar during decomposition was highest in the spring and lowest in the summer of 22, reflecting seasonal variation (). Furthermore, the concentration of P in white poplar leaf litter exhibited an increase in autumn 2021 and spring and summer 2022. The order of P concentration, from highest to lowest, was + NPK > +NP > +MF > +BF > +N > CK > PK. Notably, there was a significant difference in P concentration between + NPK and + BF treatments (). However, the concentration K in white poplar leaf litter significantly decreased throughout all periods of the experiment. Comparatively, the concentration of K in white poplar was higher in + NPK, +PK, +MF, +BF, and + N treatments compared to the control plots ().
Figure 4. The dynamic of Carbon, Nitrogen, Phosphorus, and Potassium concentration in leaf litter for treatments during the decomposition process at four sampling times (means ± SE), p < 0.05.
3.3. Dynamic characteristics of soil physiochemical properties
The utilization of various fertilizers had a significant impact on the physicochemical properties of the soil, as evidenced by the data presented in (). In the initial phase of the experiment, which covered autumn and winter 2021, remarkable increase in soil pH were observed in all experimental treatments (+NPK, +MF, +PK + NP, +N). Fertilizer application had a significant effect on soil physical properties, especially soil moisture, capillary porosity and non-capillary porosity in spring and summer 2022. However, no significant difference was detected in soil bulk density that could be attributed to fertilization. In particular, in summer 2022, +NPK had a higher capillary porosity of 26.76%, +NPK had a higher non-capillary porosity of 13.46% and + MF had a higher soil moisture content of 13.76%, as indicated in (). The assessment of SOC concentration significantly decreased over the year experiment; the concentration of SOC in autumn 2021 in orderly from highest to the lowest (+MF, +NPK, +BF, CK, +NP, +PK, +N), the assessment of SOC in winter 2021 and spring and summer 2022 were respectively from highest to the lowest (+MF, +CK, +NPK, +NP, +PK, and + N) ().
Figure 5. Dynamic characteristics of soil (SOC, TN, TP, and TK) for treatments during the decomposition process at four sampling times (means ± SE), p < 0.05.
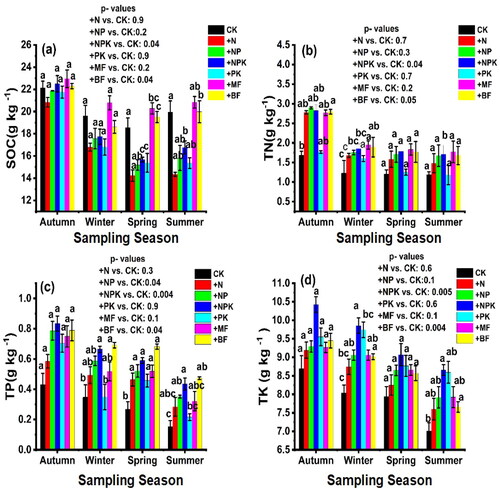
Table 2. Comparison of physicochemical characteristics of soil in different fertilizer treatments at white poplar forest at four sampling times. Different lower- case letters within columns indicate significant difference among means.
The concentration of TN in the soil exhibited an initial decrease during the first sampling time in winter (2021) followed by consistent decreases in spring and summer (2022). In the autumn of 2021, which marked the initial stage of the experiment, the TN content was notably higher in plots treated with + NP, +NPK, +N, +BF, and + MF compared to the control plots. Specifically, in winter 2021, the TN content ranked highest to lowest as + MF, +BF, +NPK, +NP, +N, +PK, and CK. In the spring and autumn of 2022, the TN content followed a similar trend, with + MF, +BF, +NPK, +NP, +N, +PK, and CK showing decreasing concentrations from highest to lowest, as illustrated in (). The response of soil TP and TK different fertilizers were varied at first sampling time, and after first sampling time significantly decreased (p < 0.05), that the content of soil TP was respectively from highest to the lowest + BF, +NPK, +NP, +MF,+N, +PK, CK in winter (2021), +BF, +NPK, +NP,+MF,+N, +PK, CK in spring (2022), +BF, +NPK, +NP, +MF, +N, +PK, CK in summer (2022) (), while the content of soil TK from highest to the lowest were respectively in + NPK, +PK, +MF, +NP, +BF, +N, CK in winter (2021), +NPK, +PK, +MF,+NP, +BF, +N, CK in spring (2022), +NPK, +PK, +NP, +MF, +BF, +N, CK in summer (2022) ().
3.4. Stoichiometric characteristics of C, N, and P response fertilizer applications
As illustrated in , the ratio of C to N in white poplar leaf litter was highest during the initial sampling stages, specifically in the autumn and winter of 2021, as well as in the summer and spring of 2022. The ratio of C to P concentration in white poplar litter under different fertilizers was highest in autumn, winter (2021), and spring and summer (2022), respectively (). The ratio N to P response to different fertilizers shown in () significantly varied, which was highest in spring (2022) and winter (2021) and lowest in summer (2022) and autumn (2021), respectively. The ratio C:N in soil shown in () significantly varied in autumn (2021) and summer (2022) (p<0.05) among the different fertilizer treatments. As shown in (), the ratio of C:P in soil under different fertilizers significantly increased (p<0.05). In contrast, the ratio of N to P concentration in soil varied at different experiment stages in different fertilizer treatments ().
Figure 6. Stoichiometric characteristics of C, N, P, and K in leaf litter and soil for treatments during the decomposition process at four sampling times (means ± SE), p < 0.05, p < 0.01, p < 0.001.
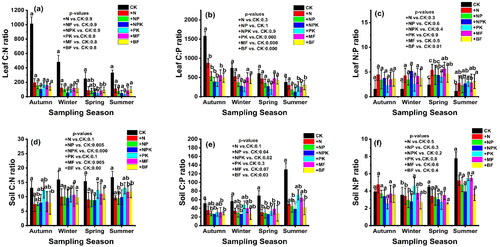
The ratios of C:N in leaf litter under fertilizer treatments at the final stage of the experiment (CK, +N, +NP, +NPK, +PK, +MF, +BF) were 332.58, 109.78, 81.57, 48.18, 35.5, 88. 7, and 98.15, respectively. The C:P ratios in leaf litter under fertilizers at the final stage of the experiment (CK + N, +NP, +NPK, +PK, +MF, +BF) were 379.9, 297.61, 180.67, 128.9, 266.85, 252.07, and 302.25, respectively. The ratios N:P in leaf litter under fertilizers treatments at the final stage of the experiment (CK + N, +NP, +NPK, +PK, +MF, +BF) were 1.14, 2.71, 2.21, 2.67, 3.13, 2.84, and 3.07 respectively. In soil from (CK + N, +NP, +NPK, +PK, +MF, +BF) at the final stage of the experiment, the ratios of C:N were 16.74, 9.68, 9.46, 9.80, 12.9, 11.75, 11.89 respectively. The C:P ratios were 130.16, 50.48, 44.99, 38.57, 71.23, 64.69, 42.17 respectively. The N:P ratios were 7.77, 5.21, 4.75, 3.93, 5.48, 5.5, and 3.54 at the final stage of the experiment, respectively. The differences in white poplar leaf litter C:N, C:P, and N:P ratios, and soil C:N, C:P, and: N:P ratios were positive significant (p<0.05).
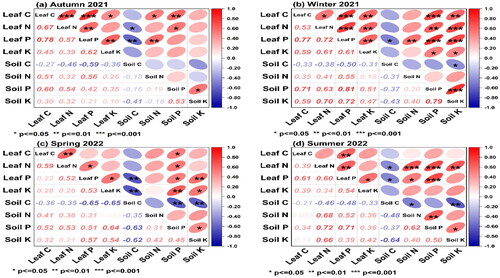
3.5. Relationship between the stoichiometric characteristics of C, N, P, and K as well as their concentrations in various fertilizer applications
The associations between C, N, P, and K content in leaf litter and soil from various fertilizer application treatments at four sampling stages are illustrated in . A significant positive correlation exists between C, N, P and K in leaf litter (p<0.01, p<0.05). At four sampling stages, additionally, we found a negative association between C, N, P, and K concentration in leaf litter and SOC (p<0.01, p<0.05). Positive associations were found between C, N, P, and K concentrations in leaf litter and soil P and K at four sampling times (p<0.01, p<0.05). Under different fertilizer treatments, SOC negatively correlated with soil total N, P, and K (p<0.05, p<0.01) at four seasons.
Figure 7. Correlations between C, N, P and K concentration in leaf litter and soil at four sampling stages (means ± SE), p < 0.05, p < 0.01, p < 0.001.
illustrates the correlation between C, N, P, and K in leaf litter and soils and their stoichiometry at four sampling times in different treatments. The results showed that a significant negative relationship was determined between stoichiometric characters of C, N, P and K concentration in leaf litter at four sampling stages, except for the relationship between C concentration and C:P, N:P were found positive. At four sampling stages, strong negative relationships were found between C, N, P, and K concentrations in leaf litter and soil stoichiometry (p<0.01). Additionally, we found there is a positive connection between leaf C:N and soil C:N, C:N < N:P in the first sampling stages, while the relationship between leaf N:P and soil C:N, C:P, N:P were negative (p<0.05).
Figure 8. Correlations between C, N, P and K concentration and between their stoichiometric characters at four sampling stages (means ± SE), p < 0.05, p < 0.01, p < 0.001.
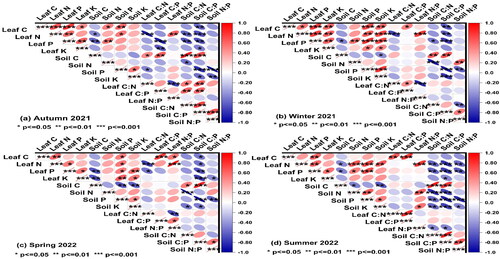
4. Discussion
4.1. Leaf litter decomposition under various fertilizer treatments
Fertilizers afford important nutrients to plants, including N, P, and K potentially influencing the decomposition practice of leaf litter (Chen et al., Citation2013; Cissé et al., Citation2021; Zhang et al., Citation2021). In this investigation, we found that fertilizer application on leaf litter positively impacted the decomposition rate in the white poplar forest. The decomposition process was accelerated mainly in + MF and + NPK at four samplings, while the impact of nitrogen addition (+N) on decomposition was very slow at all stages of the investigation; this was consistent with earlier studies. (Du et al., Citation2020; Zhou et al., Citation2017), which found that the accumulation of organic fertilizers considerably accelerated litter decomposition rates compared to control plots (Liu and Wang, Citation2021; Yu et al., Citation2015). Nitrogen application can lead to a reduction in soil microbial biomass and alter the microbial community composition, which may contribute to the observed decrease in the decomposition rate. In our study, the application of + MF, +NPK, and + BF fertilizers significantly influenced the decomposition rate of white poplar leaf litter. These treatments notably accelerated the decomposition process, which has not been previously studied in the context of white poplar leaf litter. Previous research has mainly focused on the effect of phosphorus addition, which was found to increase the decomposition of coarse woody debris (CWD) (Chen et al., Citation2016).
4.2. Effect of different fertilizers on leaf litter nutrient concentration (C, N, P, and K)
The effect of various fertilizers on leaf litter nutrient concentrations, including N, P, and K, were varied, and Typically, the concentration of C and K decreased during the first stage of the experiment, which occurred during leaf litter decomposition. The results from our one-year decomposition experiment revealed that various fertilizer applications and decomposition stages and their interaction are significant factors in understanding the observed variations. Specifically, the different fertilizer applications accelerated the decomposition of leaf litter. However, our results showed that C concentration after the first sampling stages significantly decreased at all stages of the experiment, respectively, from highest to the lowest (CK > +N > +NP > +NPK > +PK > +MF > +BF) except the spring season (p<0.05) (), The concentration of N in white poplar gradually increased at all stages of the experiments. This can be attributed to the high C:N ratio of the materials used, as their decomposition released N over time. However, it varied among the fertilizer treatments in different seasons (). The concentration of P in white poplar leaf litter exhibited a consistent increase during autumn 2021 and spring and summer 2022, as depicted in . These findings provide support for the widely accepted assumption that the availability of nutrients in the soil and the concentration of nutrients in plant tissues are closely interconnected (Elser et al., Citation2000; W. Zhang et al., Citation2020). The association between TN and TP concentrations in both leaf and soil has been documented at both the community and species levels. At the community level, elevated atmospheric N deposition is linked with higher plant N concentration in urban areas compared to rural areas. Conversely, in rural areas, P fertilization leads to increased soil P and plant P concentration (Elser et al., Citation2007; Sardans and Peñuelas, Citation2015). N fertilization enhanced plant TN concentration at the species level from 18 to 65% in Chinese fir (Xu and Timmer, Citation1999) and 24% in slash pine (Chen et al., Citation2010). Similarly, P fertilization resulted in a 20% increase in tissue in slash pine (Hu et al., Citation2011) and up to 800% in Sitka spruce (Proe and Millard, Citation1995). However, the concentration of K in white poplar leaf litter significantly reduced in all stages of the experiment; the concentration of K in white polar was greater in + NPK, +PK,+MF,+BF, and + N in comparison to the control plots (); this study was in agreement with the previous study (Sardans and Peñuelas, Citation2015).
4.3. Effect of fertilizers on dynamic characteristics of soil (SOC, TN, TP and TK)
The response of soil nutrients to various fertilizers during the breakdown of leaf litter can be complex and is influenced by various factors. The decomposition process involves the breakdown of organic matter in leaf litter, releasing nutrients back into the soil, various fertilizers, when applied to the ground, can interact with this process and affect the availability and cycling of nutrients (Fang et al., Citation2021). In this study, the concentration SOC significantly decreased at + NPK and + BF in comparison to control plots; the concentration of soil TN in white poplar at first sampling time increased and constantly decreased respectively at winter (2021), spring and summer (2022), The response of soil TP and TK different fertilizers were varied at first sampling time, and after first sampling time significantly decreased (p<0.05), in our study, the impact of various fertilizer is evident and had a significant effect on soil fertility a prior study suggested that adding mineral fertilizers is advantageous for the soil health; however, an excessive addition of phosphorus can lead to phosphorus fixation which can further decline soil health. Our finding agrees with a previous study (Chen et al., Citation2015; Liu and Wang, Citation2021) found that nitrogen fertilization significantly increased soil nitrate and ammonium concentrations, indicating increased mineralization of organic nitrogen during leaf litter decomposition, +NP, +NPK, +BF fertilizers had a significant impact on total phosphors in soil, (Keuskamp et al., Citation2015) found that nitrogen fertilization significantly increased soil nitrogen concentrations, while phosphorus and potassium fertilization had no significant effect. Our findings indicate that the response of soil nutrients to fertilizers during leaf litter decomposition may be primarily driven by nitrogen availability, while the influence of other nutrients may be more context-dependent or less significant.
4.4. The patterns of leaf litter and soil C, N, and P stoichiometry of white poplar
The stoichiometric ratios C, N, and P in soil and leaf litter serve not only to unveil the interconnected relationships between these components but also to signify the decomposability of leaf litter and the pace of nutrient cycling (Aerts et al., Citation2012; Ge and Xie, Citation2017; Güsewell and Verhoeven, Citation2006). The N:P ratio in leaves serves as a valuable indicator of nutrient-limiting conditions that impact a plant at various growing stages (Wang and Moore, Citation2014). Tree growth is primarily constrained by N when the N:P ratio below 14, whereas it becomes limited by P when the N:P ratio exceeds 16. Growth is limited by both N and P when the N:P ratio falls within the range of 14 to 16 (Cheng et al., Citation2022; Huang et al., Citation2023; Shen et al., Citation2023). The N:P ratio in leaves from white poplar was <14 at four sampling times; this indicates that N was the constrained factor in the forest ecosystem in the experimental region, potentially attributed to human activities and serious soil erosion. However, there is ongoing judging the determination of critical nutrient dosage using the stoichiometric ratio of elements (Ellison, Citation2006; Güsewell and Verhoeven, Citation2006; Wu et al., Citation2012). The conditions and scale of the study area need to be taken into account when accessing soil nutrients.
The N:P ratio significantly influences litter degradation and nutrient return speed. Faster degradation occurs at lower N: P ratios, while degradation slows when the N: P ratio exceeds 25, indicating conditions conducive to nutrient storage (Chen et al., Citation2020). The leaf litter's N:P ratio in the white poplar area is <25, crucial for enhancing ecosystem functions like soil erosion mitigation and soil moisture retention. Establishing a bush layer is vital to optimize stand structure, as it promotes higher N:P ratios in leaf litter during subsequent arbor and mixed forest formation. The C:P ratio in leaf litter surpasses both the C:N and N:P ratios, indicating lower N content in the white poplar. In this study, the substantial significance of P is apparent in the lower C:P ratios, impacting plant growth. The variations in C:P ratios within soils were significant across different fertilization groups in the experimental area, indicating a consistent impact level of soil phosphorus in this region. Moreover, while soil P availability in the white poplar area surpassed certain thresholds for ecosystem degradation, it still remained below the global average. The soil N:P ratios proved to be a reliable indicator for predicting nutrient limitation. Notable differences in soil N:P ratios among various fertilization treatments in the white poplar area suggest variations in nutrient limitation types among different plant functional groups. The diminished levels of N and P in soils from the studied areas suggest limitations in these nutrients within the examined regions.
4.5. Relationship between C, N, P and K concentration and between their stoichiometric characters in different fertilizer treatments
This study examines the variations in carbon, nitrogen and phosphorus contents across diverse fertilizer treatments along with their relationships with stoichiometric characteristics. Fertilizers are crucial for maintaining the stability of the forest ecosystem. The elements C, N, P, and K are vital for tree growth, playing significant roles in various processes related to tree growth and behavior. Leaf litter and soil, serving as representative components of biotic and plant factors in the forest ecosystem, exhibit sensitivity to environmental changes (Liu et al., Citation2010). The changes are determined by the different functions of soil and plant Yang et al. (Citation2014) and their differing C, N, and P ratios. This investigation identified a significant correlation between N and P in both leaf litter and soil and their stoichiometry (p < 0.01). While prior studies Niu et al. (Citation2017); Liu and Wang (Citation2021) noted a linear correlation between nitrogen and phosphorus contents in plants and soil, our current investigation revealed a notable correlation among C, N, and P contents in both leaf litter and soil. This correlation is consistent with the stoichiometry observed in Tianshan, China (Xie et al., Citation2016). The variations that are seen could result from the effects of different plant species, types of fertilizers, growth, types of soil, ecosystems, or human activity on the components that plants contain (Yu et al., Citation2014).
Similar to this study, Wu et al. (Citation2023) revealed that the C:N and C:P ratios are higher in leaf litter than in soil, with a significant linear relationship (p < 0.01) between the stoichiometry and total nitrogen, total phosphorus and total potassium content of leaves were found (Dong et al., Citation2021). C, N, and P in leaf litter had a significant impact on soil C:N: P ratio stoichiometry characteristics. This emphasizes the contributions of nutrient return within the forest ecosystem as revealed in this study. Strong relationships exist between the carbon, nitrogen, phosphorus and potassium content and their stoichiometry ratios. Reflecting a self-fertilizing aspect within the ecosystem. The white poplar forests we studied are significantly impacted by human activities, including factors like firewood collection, grazing, deforestation and reclamation. However, furthermore, our findings highlight the soil’s influences on the carbon to nitrogen to phosphorus ratio under various fertilizers.
5. Conclusions
In conclusion, mineral fertilizers, manure fertilizers, and bird fertilizers greatly affected leaf litter decomposition, the concentration of nutrients, and their stoichiometry. Our study showed that fertilization generally leads to an increase in nutrient concentrations in leaf litter, reflecting the higher nutrient content and accelerated decomposition rates; the decay rate constant (k) shows the decomposition rate of white poplar leaf litter in and orderly + MF > +NPK > +BF > +PK > +NP > +N > CK (0.56, 0.53, 0.52, 0.51,0.51,0.5, and 0.46) facilitated by fertilizers; the impact of fertilizer applications on leaf litter is primarily driven by increased nutrient availability, which stimulates microbial activity and enhances the decomposition process. This research presents essential insights into understanding the effect of various fertilizers in leaf litter decomposition and nutrient cycling. However, Further research is needed to explore the implications of long-term fertilizer and potential tradeoffs in different ecosystems.
Author Contributions
All authors contributed to the study conception. Material preparation, data collection and analysis were performed by Monira Fayaz and Abdul Baess Keyhani. The first draft of the manuscript was written by Monira Fayaz and Abdul Baess Keyhani and Professor Yang Guang and other authors commented on the manuscript. All authors read and approved the final manuscript.
Acknowledgments
We would like to thank Ustad Hasamuddin for providing access and logistical support for the field fertilization experiment. Special thanks to Mr. Abdul Sabur for field and laboratory work assistance.
Data Availability
Data will be available on request.
Conflicts of Interest
The authors have no conflict of interest in this manuscript.
References
- Aerts, R., van Bodegom, P.M., Cornelissen, J.H.C., 2012. Litter stoichiometric traits of plant species of high-latitude ecosystems show high responsiveness to global change without causing strong variation in litter decomposition. New Phytol. 196, 181–188. doi: 10.1111/j.1469-8137.2012.04256.x.
- Akoto, D.S., Partey, S.T., Abugre, S., Akoto, S., Denich, M., Borgemeister, C., Schmitt, C.B., 2022. Comparative analysis of leaf litter decomposition and nutrient release patterns of bamboo and traditional species in agroforestry system in Ghana. Clean. Mater. 4. doi: 10.1016/j.clema.2022.100068.
- Allen, J., Maunoury-Danger, F., Felten, V., Danger, M., Legout, A., Guérold, F., 2020. Liming of acidified forests changes leaf litter traits but does not improve leaf litter decomposability in forest streams. For. Ecol. Manage. 475. doi: 10.1016/j.foreco.2020.118431.
- Brilli, F., Gioli, B., Zona, D., Pallozzi, E., Zenone, T., Fratini, G., Calfapietra, C., Loreto, F., Janssens, I.A., Ceulemans, R., 2014. Simultaneous leaf- and ecosystem-level fluxes of volatile organic compounds from a poplar-based SRC plantation. Agric. For. Meteorol. 187, 22–35. doi: 10.1016/j.agrformet.2013.11.006.
- Cai, A., Liang, G., Yang, W., Zhu, J., Han, T., Zhang, W., Xu, M., 2021. Patterns and driving factors of litter decomposition across Chinese terrestrial ecosystems. J. Clean. Prod. 278. doi: 10.1016/j.jclepro.2020.123964.
- Carreiro, M.M., Sinsabaugh, R.L., Repert, D.A., Parkhurst, D.F., 2000. Microbial enzyme shifts explain litter decay responses to simulated nitrogen deposition. Ecology 81, 2359–2365. doi: 10.1890/0012-9658(2000)081[2359:MESELD]2.0.CO;2.
- Chen, F. sheng, Fahey, T.J., Yu, M. yuan, Gan, L., 2010. Key nitrogen cycling processes in pine plantations along a short urban-rural gradient in Nanchang, China. For. Ecol. Manage. 259, 477–486. doi: 10.1016/j.foreco.2009.11.003.
- Chen, F.S., Niklas, K.J., Liu, Y., Fang, X.M., Wan, S.Z., Wang, H., 2015. Nitrogen and phosphorus additions alter nutrient dynamics but not resorption efficiencies of Chinese fir leaves and twigs differing in age. Tree Physiol. 35, 1106–1117. doi: 10.1093/treephys/tpv076.
- Chen, H., Dong, S., Liu, L., Ma, C., Zhang, T., Zhu, X., Mo, J., 2013. Effects of experimental nitrogen and phosphorus addition on litter decomposition in an old-growth tropical forest. PLoS One 8. doi: 10.1371/journal.pone.0084101.
- Chen, J., Xiao, W., Zheng, C., Zhu, B., 2020. Nitrogen addition has contrasting effects on particulate and mineral-associated soil organic carbon in a subtropical forest. Soil Biol. Biochem. 142. doi: 10.1016/j.soilbio.2020.107708.
- Chen, Y., Sayer, E.J., Li, Z., Mo, Q., Li, Y., Ding, Y., Wang, J., Lu, X., Tang, J., Wang, F., 2016. Nutrient limitation of woody debris decomposition in a tropical forest: Contrasting effects of N and P addition. Funct. Ecol. 30, 295–304. doi: 10.1111/1365-2435.12471.
- Cheng, X., Xing, W., Xiang, W., 2022. Depth-dependent patterns in the C:N:P stoichiometry of different soil components with reclamation time in coastal poplar plantations. Soil Tillage Res. 223. doi: 10.1016/j.still.2022.105494.
- Cissé, M., Traoré, S., Bationo, B.A., 2021. Decomposition and nutrient release from the mixed leaf litter of three agroforestry species in the Sudanian zone of West Africa. SN Appl. Sci. 3. doi: 10.1007/s42452-021-04242-y.
- Cornwell, W.K., Cornelissen, J.H.C., Amatangelo, K., Dorrepaal, E., Eviner, V.T., Godoy, O., Hobbie, S.E., Hoorens, B., Kurokawa, H., Pérez-Harguindeguy, N., Quested, H.M., Santiago, L.S., Wardle, D.A., Wright, I.J., Aerts, R., Allison, S.D., Van Bodegom, P., Brovkin, V., Chatain, A., Callaghan, T. V., Díaz, S., Garnier, E., Gurvich, D.E., Kazakou, E., Klein, J.A., Read, J., Reich, P.B., Soudzilovskaia, N.A., Vaieretti, M.V., Westoby, M., 2008. Plant species traits are the predominant control on litter decomposition rates within biomes worldwide. Ecol. Lett. 11, 1065–1071. doi: 10.1111/j.1461-0248.2008.01219.x.
- Das, D.K., Chaturvedi, O.P., 2005. Structure and function of Populus deltoides agroforestry systems in eastern India: 2. Nutrient dynamics. Agrofor. Syst. 65, 223–230. doi: 10.1007/s10457-005-1267-1.
- Dong, C., Qiao, Y., Cao, Y., Chen, Y., Wu, X., Xue, W., 2021. Seasonal variations in carbon, nitrogen and phosphorus stoichiometry of a Robinia pseudoacacia plantation on the Loess Hilly Region, China. Forests 12, 1–14. doi: 10.3390/f12020214.
- Dong, L., Berg, B., Sun, T., Wang, Z., Han, X., 2020. Response of fine root decomposition to different forms of N deposition in a temperate grassland. Soil Biol. Biochem. 147. doi: 10.1016/j.soilbio.2020.107845.
- Du, M., Feng, H., Zhang, L., Pei, S., Wu, D., Gao, X., Kong, Q., Xu, Y., Xin, X., Tang, X., 2020. Variations in carbon, nitrogen and phosphorus stoichiometry during a growth season within a platycladus orientalis plantation. Polish J. Environ. Stud. 29, 3549–3560. doi: 10.15244/pjoes/117759.
- Ellison, A.M., 2006. Nutrient limitation and stoichiometry of carnivorous plants, in: Plant Biology. pp. 740–747. doi: 10.1055/s-2006-923956.
- Elser, J.J., Bracken, M.E.S., Cleland, E.E., Gruner, D.S., Harpole, W.S., Hillebrand, H., Ngai, J.T., Seabloom, E.W., Shurin, J.B., Smith, J.E., 2007. Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecol. Lett. 10, 1135–1142. doi: 10.1111/j.1461-0248.2007.01113.x.
- Elser, J.J., Sterner, R.W., Gorokhova, E., Fagan, W.F., Markow, T.A., Cotner, J.B., Harrison, J.F., Hobbie, S.E., Odell, G.M., Weider, L.W., 2000. Biological stoichiometry from genes to ecosystems. Ecol. Lett. doi: 10.1046/j.1461-0248.2000.00185.x.
- Fang, X.M., Zhang, X.L., Zong, Y.Y., Li, W.Q., Li, J.J., Guo, L.P., Wang, H., Chen, F.S., 2021. Responses of leaf litter decomposability to nitrogen and phosphorus additions are associated with cell wall carbohydrate composition in a subtropical plantation. Plant Soil 467, 359–372. doi: 10.1007/s11104-021-05099-1.
- Fanin, N., Alavoine, G., Bertrand, I., 2020. Temporal dynamics of litter quality, soil properties and microbial strategies as main drivers of the priming effect. Geoderma 377. doi: 10.1016/j.geoderma.2020.114576.
- Ge, J., Xie, Z., 2017. Leaf litter carbon, nitrogen, and phosphorus stoichiometric patterns as related to climatic factors and leaf habits across Chinese broad-leaved tree species. Plant Ecol. 218, 1063–1076. doi: 10.1007/s11258-017-0752-8.
- González, I., Sixto, H., Rodríguez-Soalleiro, R., Oliveira, N., 2020. Nutrient contribution of litterfall in a short rotation plantation of pure or mixed plots of populus alba l. And robinia pseudoacacia l. Forests 11, 1–19. doi: 10.3390/f11111133.
- Griu, T., Lunguleasa, A., 2016. The use of the white poplar (Populus alba L.) biomass as fuel. J. For. Res. 27, 719–725. doi: 10.1007/s11676-015-0178-x.
- Gruhn, P., Goletti, F., Yudelman, M., 2000. Integrated nutrient management, soil fertility, and sustainable agriculture: Current issues and future challenges. Food, Agric. Environ. Discuss. Pap. 31.
- Gurung, R., Harada, K., Dahal, N.K., Adhikari, S., Katel, O., 2023. The Transition of Sokshing (Leaf Litter Forest) Property Rights and Management: A Case Study of Punakha and Wangdue District, Bhutan. SSRN Electron. J. doi: 10.2139/ssrn.4341820.
- Güsewell, S., Verhoeven, J.T.A., 2006. Litter N:P ratios indicate whether N or P limits the decomposability of graminoid leaf litter. Plant Soil 287, 131–143. doi: 10.1007/s11104-006-9050-2.
- Hu, D., Wang, M., Zheng, Y., Lv, M., Zhu, G., Zhong, Q., Cheng, D., 2021. Leaf litter phosphorus regulates the soil meso- and micro-faunal contribution to home-field advantage effects on litter decomposition along elevation gradients. Catena 207. doi: 10.1016/j.catena.2021.105673.
- Hu, X.F., Chen, F.S., Nagle, G., Fang, Y.T., Yu, M.Q., 2011. Soil phosphorus fractions and tree phosphorus resorption in pine forests along an urban-to-rural gradient in Nanchang, China. Plant Soil 346, 97–106. doi: 10.1007/s11104-011-0799-6.
- Huang, L., Hu, H., Bao, W., Hu, B., Liu, J., Li, F., 2023. Shifting soil nutrient stoichiometry with soil of variable rock fragment contents and different vegetation types. Catena 220. doi: 10.1016/j.catena.2022.106717.
- Keuskamp, J.A., Hefting, M.M., Dingemans, B.J.J., Verhoeven, J.T.A., Feller, I.C., 2015. Effects of nutrient enrichment on mangrove leaf litter decomposition. Sci. Total Environ. 508, 402–410. doi: 10.1016/j.scitotenv.2014.11.092.
- Khalsa, S.D.S., Smart, D.R., Muhammad, S., Armstrong, C.M., Sanden, B.L., Houlton, B.Z., Brown, P.H., 2020. Intensive fertilizer use increases orchard N cycling and lowers net global warming potential. Sci. Total Environ. 722. doi: 10.1016/j.scitotenv.2020.137889.
- Knorr, M., Frey, S.D., Curtis, P.S., 2005. Nitrogen additions and litter decomposition: A meta-analysis. Ecology 86, 3252–3257. doi: 10.1890/05-0150.
- Li, R., Guo, X., Han, J., Yang, Q., Zhang, W., Yu, X., Han, X., Chen, L., Guan, X., Zeng, Z., Yang, H., Wang, S., 2023. Global pattern and drivers of stable residue size from decomposing leaf litter. Catena 232. doi: 10.1016/j.catena.2023.107390.
- Liu, P., Huang, J., Sun, O.J., Han, X., 2010. Litter decomposition and nutrient release as affected by soil nitrogen availability and litter quality in a semiarid grassland ecosystem. Oecologia 162, 771–780. doi: 10.1007/s00442-009-1506-7.
- Liu, R., Wang, D., 2021. C:N:P stoichiometric characteristics and seasonal dynamics of leaf-root-litter-soil in plantations on the loess plateau. Ecol. Indic. 127. doi: 10.1016/j.ecolind.2021.107772.
- Liu, X., Chen, S., Li, X., Yang, Z., Xiong, D., Xu, C., Wanek, W., Yang, Y., 2022. Soil warming delays leaf litter decomposition but exerts no effect on litter nutrient release in a subtropical natural forest over 450 days. Geoderma 427. doi: 10.1016/j.geoderma.2022.116139.
- Liu, X.-Z., Zhou, G.-Y., Zhang, D.-Q., Liu, S.-Z., Chu, G.-W., Yan, J.-H., 2010. N and P stoichiometry of plant and soil in lower subtropical forest successional series in southern China. Chinese J. Plant Ecol. 34, 64–71.
- Liu, Y., Wang, K., Dong, L., Li, J., Wang, X., Shangguan, Z., Qu, B., Deng, L., 2023. Dynamics of litter decomposition rate and soil organic carbon sequestration following vegetation succession on the Loess Plateau, China. Catena 229. doi: 10.1016/j.catena.2023.107225.
- Meiresonne, L., De Schrijver, A., De Vos, B., 2007. Nutrient cycling in a poplar plantation (Populus trichocarpa x Populus deltoides “Beaupré”) on former agricultural land in northern Belgium. Can. J. For. Res. 37, 141–155. doi: 10.1139/x06-205.
- Micks, P., Downs, M.R., Magill, A.H., Nadelhoffer, K.J., Aber, J.D., 2004. Decomposing litter as a sink for 15N-enriched additions to an oak forest and a red pine plantation. For. Ecol. Manage. 196, 71–87. doi: 10.1016/j.foreco.2004.03.013.
- Nakatsuka, H., Karasawa, T., Ohkura, T., Wagai, R., 2020. Soil faunal effect on plant litter decomposition in mineral soil examined by two in-situ approaches: Sequential density-size fractionation and micromorphology. Geoderma 357. doi: 10.1016/j.geoderma.2019.113910.
- Niu, S., Ren, L., Song, L., Duan, Y., Huang, T., Han, X., Hao, W., 2017. Plant stoichiometry characteristics and relationships with soil nutrients in Robinia pseudoacacia communities of different planting ages. Acta Ecol. Sin. 37, 355–362. doi: 10.1016/j.chnaes.2017.10.003.
- Ouyang, W., Wu, Z., Wang, P., Lin, C., Zhu, W., 2022. Forest Leaf Litter Nutrient Discharge Patterns in Snowmelt Surface Runoff and Watershed Scale Remote Sensed Simulation. SSRN Electron. J. doi: 10.2139/ssrn.4086333.
- Pang, Y., Tian, J., Lv, X., Wang, R., Wang, D., Zhang, F., 2022. Contrasting dynamics and factor controls in leaf compared with different-diameter fine root litter decomposition in secondary forests in the Qinling Mountains after 5 years of whole-tree harvesting. Sci. Total Environ. 838. doi: 10.1016/j.scitotenv.2022.156194.
- Proe, M.F., Millard, P., 1995. Effect of P supply upon seasonal growth and internal cycling of P in Sitka spruce (Picea sitchensis(Bong.)Carr.) seedlings. Plant Soil 168-169, 313–317. doi: 10.1007/BF00029343.
- Ren, H., Qin, J., Yan, B., Alata, Baoyinhexige, Han, G., 2018. Mass loss and nutrient dynamics during litter decomposition in response to warming and nitrogen addition in a desert steppe. Front. Agric. Sci. Eng. 5, 64–70. doi: 10.15302/J-FASE-2017194.
- Salehi, A., Ghorbanzadeh, N., Salehi, M., 2013. Soil nutrient status, nutrient return and retranslocation in poplar species and clones in northern Iran. IForest 6, 336–341. doi: 10.3832/ifor0976-006.
- Sardans, J., Peñuelas, J., 2015. Potassium: A neglected nutrient in global change. Glob. Ecol. Biogeogr. doi: 10.1111/geb.12259.
- Shao, S., Wurzburger, N., Sulman, B., Hicks Pries, C., 2023. Ectomycorrhizal effects on decomposition are highly dependent on fungal traits, climate, and litter properties: A model-based assessment. Soil Biol. Biochem. 184. doi: 10.1016/j.soilbio.2023.109073.
- Shen, F., Liu, N., Shan, C., Ji, L., Wang, M., Wang, Y., Yang, L., 2023. Soil extracellular enzyme stoichiometry reveals the increased P limitation of microbial metabolism after the mixed cultivation of Korean pine and Manchurian walnut in Northeast China. Eur. J. Soil Biol. 118. doi: 10.1016/j.ejsobi.2023.103539.
- Sinsabaugh, R.L., 2010. Phenol oxidase, peroxidase and organic matter dynamics of soil. Soil Biol. Biochem. doi: 10.1016/j.soilbio.2009.10.014.
- Song, X., Wang, Z., Tang, X., Xu, D., Liu, B., Mei, J., Huang, S., Huang, G., 2020. The contributions of soil mesofauna to leaf and root litter decomposition of dominant plant species in grassland. Appl. Soil Ecol. 155. doi: 10.1016/j.apsoil.2020.103651.
- Song, Y.J., Tian, W. Bin, Liu, X.Y., Yin, F., Cheng, J.Y., Zhu, D.N., Ali, A., Yan, E.R., 2016. Associations between litterfall dynamics and micro-climate in forests of Putuoshan Island, Zhejiang, China. Chinese J. Plant Ecol. 40, 1154–1163. doi: 10.17521/cjpe.2016.0157.
- Stark, H., Nothdurft, A., Block, J., Bauhus, J., 2015. Forest restoration with Betula ssp. and Populus ssp. nurse crops increases productivity and soil fertility. For. Ecol. Manage. 339, 57–70. doi: 10.1016/j.foreco.2014.12.003.
- Su, J., Zhao, Y., Bai, Y., 2023. Asymmetric responses of leaf litter decomposition to precipitation changes in global terrestrial ecosystem. J. Clean. Prod. 387. doi: 10.1016/j.jclepro.2023.135898.
- Su, Y., Ma, X., Gong, Y., Li, K., Han, W., Liu, X., 2021. Contrasting effects of nitrogen addition on litter decomposition in forests and grasslands in China. J. Arid Land 13, 717–729. doi: 10.1007/s40333-021-0076-3.
- Swart, R.C., Samways, M.J., Roets, F., 2022. Interspecific green leaf-litter selection by ground detritivore arthropods indicates generalist over specialist detritivore communities. Appl. Soil Ecol. 174. doi: 10.1016/j.apsoil.2022.104439.
- Teague, R., Dowhower, S., 2022. Links of microbial and vegetation communities with soil physical and chemical factors for a broad range of management of tallgrass prairie. Ecol. Indic. 142. doi: 10.1016/j.ecolind.2022.109280.
- Wachendorf, C., Piepho, H.P., Beuschel, R., 2020. Determination of litter derived C and N in litterbags and soil using stable isotopes prevents overestimation of litter decomposition in alley cropping systems. Pedobiologia (Jena). 81–82. doi: 10.1016/j.pedobi.2020.150651.
- Wang, M., Moore, T.R., 2014. Carbon, Nitrogen, Phosphorus, and Potassium Stoichiometry in an Ombrotrophic Peatland Reflects Plant Functional Type. Ecosystems 17, 673–684. doi: 10.1007/s10021-014-9752-x.
- Wu, P., Zhou, H., Cui, Y.C., Zhao, W.J., Hou, Y.J., Zhu, J., Ding, F.J., 2023. Stoichiometric Characteristics of Leaf Nutrients in Karst Plant Species During Natural Restoration in Maolan National Nature Reserve, Guizhou, China. J. Sustain. For. 42, 95–119. doi: 10.1080/10549811.2021.1948868.
- Wu, P.P., Zhang, Z., Li, R., Ji, J.H., Mao, R., 2023. Impact of nitrogen addition on single and mixed tree leaf litter decomposition depends on N forms in subtropical China. Appl. Soil Ecol. 190. doi: 10.1016/j.apsoil.2023.104970.
- Wu, T.G., Yu, M.K., Geoff Wang, G., Dong, Y., Cheng, X.R., 2012. Leaf nitrogen and phosphorus stoichiometry across forty-two woody species in Southeast China. Biochem. Syst. Ecol. doi: 10.1016/j.bse.2012.06.002.
- Xie, J., Chang, S.L., Zhang, Y.T., Wang, H.J., Song, C.C., He, P., Sun, X.J., 2016. Plant and soil ecological stoichiometry with vertical zonality on the northern slope of the middle Tianshan Mountains. Shengtai Xuebao 36, 4363–4372. doi: 10.5846/stxb201506301387.
- Xu, X., Timmer, V.R., 1999. Growth and nitrogen nutrition of Chinese fir seedlings exposed to nutrient loading and fertilization. Plant Soil 216, 83–91. doi: 10.1023/a:1004733714217.
- Yang, J., Zhang, X., Ma, L., Chen, Y., Dang, T., An, S., 杨佳佳, 张向茹, 马露莎, 陈亚南, 党廷辉, 安韶山, 2014. Ecological stoichiometric relationships between components of robinia pseudoacacia forest in loess plateau. Acta Pedol. Sin. 51, 133–142.
- Yu, Y.F., Peng, W.X., Song, T.Q., Zeng, F.P., Wang, K.L., Wen, L., Fan, F.J., 2014. Stoichiometric characteristics of plant and soil C, N and P in different forest types in depressions between karst hills, southwest China. Chinese J. Appl. Ecol. 25, 947–954.
- Yu, Z., Huang, Z., Wang, M., Liu, R., Zheng, L., Wan, X., Hu, Z., Davis, M.R., Lin, T.C., 2015. Nitrogen addition enhances home-field advantage during litter decomposition in subtropical forest plantations. Soil Biol. Biochem. 90, 188–196. doi: 10.1016/j.soilbio.2015.07.026.
- Zhang, J., Li, H., Zhang, H., Zhang, H., Tang, Z., 2021. Responses of Litter Decomposition and Nutrient Dynamics to Nitrogen Addition in Temperate Shrublands of North China. Front. Plant Sci. 11. doi: 10.3389/fpls.2020.618675.
- Zhang, J., Li, J., Fan, Y., Mo, Q., Li, Y., Li, Y., Li, Z., Wang, F., 2020. Effect of nitrogen and phosphorus addition on litter decomposition and nutrients release in a tropical forest. Plant Soil 454, 139–153. doi: 10.1007/s11104-020-04643-9.
- Zhang, W., Chao, L., Yang, Q., Wang, Q., Fang, Y., Wang, S., 2016. Litter quality mediated nitrogen effect on plant litter decomposition regardless of soil fauna presence. Ecology 97, 2834–2843. doi: 10.1002/ecy.1515.
- Zhang, W., Wang, Q., Wu, Q., Zhang, S., Zhu, P., Peng, C., Huang, S., Wang, B., Zhang, H., 2020. The response of soil Olsen-P to the P budgets of three typical cropland soil types under long-term fertilization. PLoS One 15. doi: 10.1371/journal.pone.0230178.
- Zhou, S. xing, Huang, C. de, Han, B. han, Xiao, Y. xiang, Tang, J. dong, Xiang, Y. bin, Luo, C., 2017. Simulated nitrogen deposition significantly suppresses the decomposition of forest litter in a natural evergreen broad-leaved forest in the Rainy Area of Western China. Plant Soil 420, 135–145. doi: 10.1007/s11104-017-3383-x.
- Zhu, X., Chen, H., Zhang, W., Huang, J., Fu, S., Liu, Z., Mo, J., 2016. Effects of nitrogen addition on litter decomposition and nutrient release in two tropical plantations with N2-fixing vs. non-N2-fixing tree species. Plant Soil 399, 61–74. doi: 10.1007/s11104-015-2676-1.