
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.ABSTRACT
Tomato (Solanum lycopersicum L.) yield is severely affected by Fusarium fungal disease. To improve the resistance of tomato against Fusarium oxysporum f. sp. lycopersici (FOL), Escherichia coli katE gene was transformed into two tomato cultivars, namely Castle Rock and Super strain B, via Agrobacterium tumefaciens; the transformation efficiency was 5.6% and 3.5%, respectively. The integration of the katE gene into T0, T1, and T2 transgenic tomato lines was confirmed using PCR. In addition, DNA dot blot technique confirmed the integration of the katE gene into T2 transgenic tomato lines. The RT-PCR analysis confirmed that the katE gene could be expressed normally in the T2 modified lines. Under artificial infection with FOL, the non-modified plants exhibited more severe fungal disease symptoms than those observed in katE overexpression (OE) lines. Our analysis showed that the levels of three defense enzymes, namely superoxide dismutase (SOD), catalase (CAT), and peroxidase (POD), were increased during transgenic T2 generation pre-treated with FOL. The bioassay of modified lines revealed that an average of 52.56% of the modified Castle Rock cultivar and 50.28% of the modified Super Strain B cultivar showed resistance under Fusarium infection. These results clearly indicate that the modified tomato plants, in which the katE gene was overexpressed, became more resistant to the infection by FOL than the wild-type plants. Our study has proven that the overexpression of the E. coli katE gene in the OE lines could be utilized to develop and improve the resistance against fungal diseases in the modified crops.
Introduction
Tomato, Solanum lycopersicum L., is one of the most important vegetable crops globally due to its high nutritional value and the availability of nutrients such as vitamin C, flavonoids, beta-carotene, and lycopene.Citation1 As tomatoes contain lycopene, which has anti-cancer and antioxidant activities, the production and consumption of tomatoes have increased considerably in the past few years.Citation1,Citation2 It is noticeable that tomatoes are sold in the form of juice, concentrates, sauce, and soup, in addition to being sold as fresh vegetables. Globally, tomato production was predicted to be at about 180 million tons. Africa and Egypt produced 12% and 3.73% of the total global production of tomatoes, respectively.Citation3
The tomato crop can undergo severe losses as a result of exposure to biotic stresses, such as infection with insects and fungi, which can cause a decrease in the production, irrespective of whether grown in open fields or greenhouses.Citation4 Tomatoes are infected with several fungal pathogens such as Phytophthora, Alternaria, Rhizoctonia, and Fusarium, which lead to many devastating diseases such as late blights, early blights, root rots, and wilt. Wilt diseases were considered as one of the most severe diseases that affect tomato production.Citation5 One of the most dangerous and widespread diseases is Fusarium wilt disease caused by the soil-borne fungus Fusarium oxysporum f. sp. lycopersici (FOL).Citation6,Citation7 FOL causes a decrease in the crop yield by approximately 30–40% and this percentage may increase to about 80% under optimum environmental conditions.Citation8 The loss depends on the plant growth stage and environmental conditions.Citation9 The optimum conditions for the pathogen growth are acidic soil, dry weather, and warm temperature of about 28°C.Citation10 The causal agent, FOL, infects the plant roots, passes from the cortex to the stele, enters the xylem vessels, causes vascular wilts, and finally leads to the death of the plant.Citation11 Symptoms of the disease include yellowing of leaves, plant wilting, and a decrease in the plant productivity. The pathogen can survive in the soil for as many as 10 years.Citation12
To overcome Fusarium wilt disease in tomatoes, many methods such as the use of Agricultural practices (such as crop rotation and soil solarization), synthetic fungicides, resistant cultivars, bio-control agents, and modified lines, which are produced by the modern methods of gene transfer in the plant, are employed.Citation13 The fungicides harm humans and the surrounding environment. In addition, resistant strains of the causal agent may develop due to continuous use of fungicides.Citation14 Breeding programs using resistant varieties are a reliable method to control Fusarium wilt disease; however, this type of resistance was not found to be durable.Citation15 Further, bio-control agents have been applied for disease control; nonetheless, these agents alone cannot fully control the disease due to changes in the pH and temperature, which affect the efficiency of the biocontrol agents.Citation14 In the early 1980s, with advances in molecular plant biology and a better understanding of infection caused by some pathogens, many complex plant pathways were discovered; the immune response genes and various relevant pathways were also identified in the plants.Citation16 The gene transfer methods in plants were evaluated for the possibility of incorporating resistant genes from different species to render the plant disease resistant against infection by fungi and bacteria.Citation17–19 In order to overcome Fusarium wilt, Medicago sativa defensin (MsDef1) geneCitation18 and three other genes for pathogenesis-related proteins (glucanase, chitinase, and PR1)Citation20 were transformed into tomato plants to generate modified lines. Moreover, to confer resistance against FOL, the tomato I-3 resistance gene,Citation21 rice chitinase (RCG3) gene,Citation22 and Agrobacterium rhizogenes rolA geneCitation23 have been inserted into tomato plants to produce the modified lines.
The overexpression of anti-oxidative enzymes in plant cells was carried out to improve the plant tolerance against different biotic and abiotic stresses.Citation24 E. coli possess two types of catalases, a bifunctional catalase peroxidase (HPI) encoded by the katG gene and a monofunctional catalase (HPII) encoded by the katE gene.Citation25 The catalase HPII is composed of 732 amino acids and shows higher activity of H2O2 conversion than the plant catalase.Citation26 The katE gene has been used to provide oxidative stress,Citation26 salt,Citation27 and drought tolerance.Citation28 The katE gene was introduced into different plant species including tobacco,Citation26 rice,Citation27 jute,Citation29 and tomatoCitation30 for improving their defense system. However, the effectiveness of the katE gene in improving plant tolerance against fungal diseases, especially wilt diseases, has not yet been elucidated. Therefore, this study attempted to improve the fungal resistance in tomatoes by overexpressing the bacterial katE gene using Agrobacterium-mediated technique.
Results
Overexpression of kat-E Gene in Tomato
Two tomato (Solanum lycopersicum L.) cultivars, Castle Rock and Super Strain B, were selected for transformation experiments. The hypocotyls (the tomato explants) were co-cultivated with A. tumefaciens LBA-4404 harboring the binary vector pBI121-katE. The putative transgenic calli were transferred within 3 to 4 weeks to the shoot induction medium. The putative shoots for both the cultivars appeared within 10–15 days. The plantlets that could survive in a medium containing kanamycin sulfate were selected, transferred to plastic pots, and were kept under growth chamber conditions (). The T0 and T1 seeds were collected, screened by PCR using katE specific primers, and planted under suitable conditions; the T2 seeds obtained were used for further experiments (supplemented 1).
Figure 1. Recovery of fertile modified tomato plants expressing the bacterial katE gene. (a) The hypocotyl. (b and c) The callus induction. (d and e) The shoot induction. (f) The modified plants transferred to soil

Molecular Analysis of Putative katE Overexpression (OE) Lines
To examine the stable integration of the T-DNA in the putative modified plantlet genomes, the genomic DNA of the T0 plantlet was isolated and analyzed by PCR reaction using specific primers to screen the bacterial catalase (katE) and nptII genes. Clear bands with the expected molecular size of the katE and nptII genes (457 and 254 bp, respectively) were detected only in the modified plants; no such bands were seen in the non-modified (control) plants under identical conditions (). Moreover, the transformation efficiency of the two tested tomato cultivars was comparable. A total of 1000 explants were used for each cultivar. For the cultivar Castle Rock, 39 modified plantlets out of 700 regenerated plantlets showed positive results with the PCR analysis and survived on kanamycin sulfate containing medium, thus representing a transformation efficiency of 5.6%. For Super Strain B cultivar, 22 modified plantlets out of 630 regenerated plantlets exhibited positive PCR results, representing a transformation efficiency of 3.5% ().
Table 1. Transformation frequencies of tomato hypocotyl after Agrobacterium-mediated transformation using the pBI121-katE vector
Figure 2. Detection of katE gene by PCR in the T0 of putative modified plantlets. M: DNA Ladder (100 bp DNA Ladder RTU GeneDirex & TriDye™ 100 bp DNA Ladder BioLabs), Nc: negative control (wild type or non-modified plantlets), Pc: positive control (pBI121-katE vector). (a) katE gene detection in Castle Rock cultivar; lanes 1 and 4–7: modified plantlets; lanes 2 and 3: non-modified plantlets. (b) katE gene detection in Super Strain B cultivar; lanes 1, 3–6, and 8: modified plantlets; lanes 2 and 7: non-modified plantlets
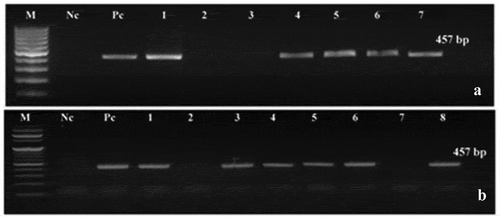
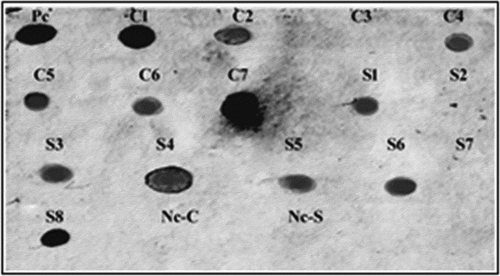
The PCR was used for the screening of T1 modified plants using katE specific primers. Furthermore, the stable inheritance of the katE gene in randomly selected T2 modified tomato plants was confirmed using the dot blot technique. Several modified plants from Castle Rock (four lines: 4, 5, 6, and 7) and Super Strain B (four lines: 3, 4, 5, and 6) cultivars exhibited positive results that confirmed that the plants were harboring katE gene (). The T2 seeds from three positive lines of each cultivar were selected and grown under greenhouse conditions and then were subjected to fungal stress to test for improvement in their fungal resistance.
Figure 3. Dot blot analysis with katE gene-specific probe for confirming the inheritance of the katE gene into T2 tomato lines. Nc-C: negative control (non-modified plant) of Castle Rock cultivar, Nc-S: negative control (non-modified plant) of Super Strain B cultivar, Pc: positive control (pBI121-KatE vector), C1, C2, and C4-C7: modified lines of Castle Rock cultivar, C3: non-modified line of Castle Rock cultivar, S1, S3-S6, and S8: modified lines of Super Strain B cultivar, S2 and S7: non-modified lines of Super Strain B cultivar
Evaluation of the resistance of modified tomato plants against Fusarium disease
The virulent FOL isolate was used to infect modified tomato lines and their corresponding wild type (control) plants. To confirm the infection, the extracted DNA from tomato (wild-type and modified) plants post 1 week of infection with FOL was analyzed by PCR using ITS primers. The data indicated that the PCR product (~550 bp) corroborating with the expected size of the Fusarium ITS region was detected only in tomato plants that were previously infected by FOL and this band was not detected in the non-infected plants (data not shown). After 45 days post inoculation, the non-modified (wild type or control) plants exhibited more severe disease symptoms than those observed in the katE OE lines ().
Figure 4. Plant growth of non-modified (wild-type) and katE overexpression (modified) plants under artificial infestation with Fusarium oxysporum f. sp. lycopersici.

The non-modified infected plants showed typical disease symptoms including stunting, yellowing, and premature loss of cotyledons and lower leaves (). Advanced symptoms like pronounced brown lesions that girdle the hypocotyl (root/shoot junction), root rot, wilting, and death were also observed in the non-modified plants, whereas, the katE OE lines showed normal growth ().
Figure 5. Effect of FOL infection on modified line leaves. (a) non-modified leaves. (b) and roots. (c) of Castle Rock cultivar

Detection of mRNA of katE in T2 modified tomato plants
RT-PCR analysis was performed using RNA samples isolated from the T2 modified lines produced from the selected T0 modified lines no. 4, 5, 6, and 7 of Castle Rock cultivar; these lines showed a positive result upon PCR and dot blot analyses, and the total RNA of the non-modified plant was also isolated. The RT-PCR analysis of the tested lines showed that the mRNA of the katE gene in the modified tomato lines 4, 5, and 7 was overexpressed; however, the katE gene mRNA in line 6 was not found to be expressed. Furthermore, the mRNA was not detected in the non-modified plants upon RT-PCR analysis (). In addition, the total RNA was also isolated from transgenic line numbers 3, 4, 5, and 6 of Super Strain B cultivar that showed positive results upon PCR and dot blot analyses and was subjected to the RT-PCR analysis. Only three lines, line numbers 3, 4, and 5, gave positive results, while line number 6 did not show any expected band.
Figure 6. RT-PCR analysis of the katE gene mRNA in T2 modified tomato lines. C4, C5, and C7: modified lines no. 4–7; Nc: non-modified plant; M: 100 bp DNA ladder

Changes in plant defense enzyme activities under fungal infection
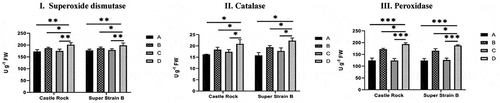
The data showed an increase in all enzyme levels under Fusarium infection for both the cultivars when compared with the non-infected plant (wild type and modified, ). Likewise, the modified plants from each cultivar pre-treated with Fusarium oxysporum f. sp. Lycopersici showed the highest levels for the three defense enzymes, i.e., superoxide dismutase (SOD), catalase (CAT), and peroxidase (POD). Under natural conditions, all non-infected plants displayed the same pattern in both Castle Rock and Super Strain B cultivars. However, statistical analysis showed that the enzyme activities of SOD, CAT, and POD were highly elevated in infected-modified plants; an increase in the activity of about 1.1-, 1.3-, and 1.5-fold was identified, respectively (). These results indicate that the expression of the katE gene in the modified tomatoes leads to an increase in the concentrations of the three defense-related enzymes that could help in improving the plant’s resistance toward fungal disease.
Table 2. Combined analysis of variance of a split-plot design for superoxide dismutase (SOD), catalase (CAT), and peroxidase (POD) enzymes
Figure 7. Defense enzyme activities in Castle Rock and Super Strain B cultivars. I. SOD activity, II. Catalase activity, III. Peroxidase activity, A: wild type (non-modified) plants, B: wild-type plants infected with Fusarium oxysporum f. sp. lycopersici, C: T2 modified plants, D: T2 modified plants infected with Fusarium oxysporum f. sp. lycopersici. Statistical analysis was performed using GraphPad Prism 8, using a two-way ANOVA with Dunnett’s multiple-comparison posttest; * = P < .05, ** = P < .01, *** = P < .001, ns = non-significant
Bioassay for modified plants
The T1 seeds of katE- modified lines were grown until the T2 seeds were obtained, which were then planted to assess the effect of fungal infection. For each cultivar, three individual lines were selected according to PCR, dot blot, and RT-PCR analyses to carry out the bioassay (seeds produced from T0 modified lines no. 4, 5, and 7 for Castle Rock cultivar and line numbers 3, 4, and 5 for Super Strain B cultivar). The modified and non-modified plants (26 seedlings) were infested with Fusarium oxysporum f. sp. lycopersici. In Castle Rock and Super Strain B cultivars, the data revealed that the average number of symptomless (resistance) of all modified line plants under Fusarium infection was 52.56% and 50.28%, respectively (). In both the cultivars, the control (non-modified) plants showed severe wilt symptoms. However, the modified plants that were resistant to Fusarium showed normal morphology, fertility, and ability to produce natural seeds. These results indicate that the expression of the katE gene in the modified tomato plants improves their resistance toward fungal disease.
Table 3. Percentage of susceptible plants, plants showing mild symptoms, and resistant plants in modified and non-modified tomato lines after the Fusarium oxysporum f. sp. lycopersici infection
Discussion
Throughout their life cycle, plants are subjected to different biotic stresses including insect, bacterial, and fungal infections. Fusarium oxysporum f. sp. lycopersici (Sacc.) is the main causative agent of tomato wilts disease. It is considered one of the most destructive tomato diseases and has caused major losses in tomato production worldwide. The tomato cultivars vary in their resistance toward Fusarium according to their genetic makeup. Agrobacterium transformation with katE gene was utilized in this study to increase the resistance of two tomato cultivars against Fusarium infection. Many reports have demonstrated the wide use of transformation techniques to improve the resistance of tomato cultivars against Fusarium. The Castle Rock cultivar of tomato was developed by MsDef1 gene overexpression to improve the FOL resistance.Citation18 In their study, the authors reported that the modified lines became more resistant to the FOL pathogen. The wheat chitinase gene (chi194) encoding a 33-kDa chitinase protein was overexpressed in tomato plants (cv. Pusa Ruby). The transgenic tomato lines with high chitinase activity were found to be highly resistant to the fungal pathogen Fusarium oxysporum f. sp. lycopersici.Citation31 The pathogen resistance was also enhanced in tomato plants transformed by the Agrobacterium-mediated technique using PBI121-ChiGluPRl plasmid containing pathogenesis-related protein genes.Citation20 The resistance genes I, I-2, and I-3 have been incorporated into cultivated tomato plants (Solanum lycopersicum) from wild tomato species to confer resistance against Fusarium oxysporum f. sp. lycoperici races 1, 2 and 3, respectively.Citation21 Besides, the expression of tobacco osmotin, bean chitinase,Citation32 and rice chitinaseCitation33 genes was utilized to produce tomato transgenic lines resistant against Fusarium oxysporum f. sp. lycopersici. Bettini et al.Citation23 transformed the tomato plants with Agrobacterium rhizogenes rolA gene to evaluate the role of this gene in the defense response of plants to FOL. Bacterial katE gene has been introduced into many different plants to evaluate its role in improving plant response under several stress conditions; examples include transgenic canola (Brassica napus L) for resistance against the airborne pathogenic fungi,Citation34 transgenic tobacco for drought and heavy light toleranceCitation28 and transgenic rice for salt tolerance improvement.Citation27,Citation35
Limones et al.Citation36 indicated the involvement of ROS in chickpea fusarium wilt caused by Fusarium oxysporum f. sp. ciceris. Their results indicated that infection by Fusarium oxysporum f. sp. ciceris led to substantial changes in the antioxidant status of chickpea. They concluded that the induction of antioxidant enzymes during chickpea fusarium wilt suggests that changes in oxidative metabolism may be a common plant defense response. Catalase acts as an effective ROS scavenger to avoid oxidative damage. Su et al.Citation37 have suggested that catalase activity may have a positive correlation with smut resistance in sugarcane. They reported that the enzymatic activity of catalase was higher in Yacheng 05–179 (resistant) variety than in Liucheng 03–182 (susceptible) variety. Catalase of E. coli cells (katE) is one of the scavenging enzymes that responds to H2O2 degradation and is involved in the resistance to oxidative stress that is induced under different stresses in many organisms.Citation27,Citation29 It is well known that H2O2 is considered as one of the most important ROS formed and is known to inhibit the plant growth. It also acts as a diffusible signal that induces downstream defense proteins.Citation38 In the present study, modified tomato plants expressing the bacterial katE gene were obtained. Modified and wild-type tomato plants were artificially infested with Fusarium oxysporum. The results showed that overexpression of the katE gene significantly improved the antifungal resistance in the modified lines. Moreover, the activity of SOD, CAT, and POD enzymes showed a significant increase in the modified tomato lines as compared to that in the non-modified plants (control) under fungal infestation. Manikandan and RaguchanderCitation39 increased the Fusarium defensive response in tomatoes using a liquid formulation of Pseudomonas fluorescens (Pf1). This led to an increase in defense mechanism of enzymes such as SOD, CAT and POD. This was in agreement with our results, which showed that an increase in the activities of SOD, CAT, and POD enzymes could lead to enhanced defensive responses in plants against Fusarium. Furthermore, Mandal et al.Citation40 have investigated the responses of tomato plants infected with Fusarium oxysporum f. sp. lycoperici by estimating the activity of the antioxidant enzymes. They found that the activities of the antioxidative enzymes, such as SOD, CAT, and POD, increased in response to pathogen inoculation. In addition, El-Awady et al.,Citation34 using chemical analyses, found high levels of catalase and peroxidase enzymes in the modified canola plants. These results agree with the present data indicating that Fusarium-infected plants tend to accumulate a much higher concentration of the detoxifying enzymes, especially CAT, to resist the fungal infection.
As a result of the expression of the bacterial katE gene in tomato plants, the resistance toward Fusarium may improve via the plant’s enhanced ability to remove H2O2. Although H2O2 is essential for signaling related to pathogen invasion and defense, the accumulation of excessive H2O2 results in oxidative stress that can damage the plant cells.Citation41 The breakdown of H2O2 prevents the formation of the highly toxic hydroxyl radical (ׄ.OH) in the plant cells.Citation42 Therefore, we envisage that the expression of the katE gene in tomato plant cells might have an important role in the enzymatic H2O2 scavenging mechanism. This enzymatic defense mechanism involves SOD and CAT enzymes.Citation43 The main role of SOD is to rapidly convert .OH to H2O2 and the produced H2O2 is then converted to oxygen and water by CAT and POD.Citation44,Citation45 Finally, catalases and other antioxidant enzymes play a key role in defending plants from the harm caused by ROS.Citation46 They maintain the normal balance of ROS in the cells; enhanced ROS production leads to gene mutations, lipid peroxidation, and disruption of molecule building and indirectly influences almost every cell activity, ultimately leading to plant cell death.Citation47,Citation48
Conclusion
The integration of the katE gene into the tomato genome was confirmed using various molecular methods. The modified tomato plants with the katE gene showed resistance to FOL by increasing the cellular activity of defense enzymes such as CAT, SOD, and POD. Their increased cellular levels improved plant resistance against Fusarium wilt by eliminating damage caused by excessive H2O2. Based on these results, it can be suggested that the expression of the katE gene may protect other plants from oxidative damage under fungal infection as well. The results of this study have immense importance in the field of agricultural production as they would facilitate the production of modified tomato cultivars that are more resistant to Fusarium wilt disease, which causes huge losses in the yield. These resistant cultivars would also be included in various breeding programs to improve tomato production.
Materials and methods
Plant materials
Two tomato (Solanum lycopersicum L.) cultivars, Super Strain B and Castle Rock, were used in all treatments. The cultivars were kindly provided by Vegetable Crops Department, Faculty of Agriculture, Cairo University, Egypt. The seeds and seedlings were prepared and maintained as described by Manikandan and Raguchander.Citation39
Primers
For Fusarium detection and identification on infected plants, the internal transcribed spacer (ITS) region was amplified using primers ITS1 5ʹ-TCCGTAGGTGAACCTGCGG-3ʹ and ITS4 5ʹ-TCCTCCGCTTATTGATATGC-3ʹ.Citation49 The katE gene was detected in modified tomato plants using two sets of oligonucleotide primers specific for the nptII and katE genes ().
Table 4. The nucleotide sequence of the katE and nptII primers used for PCR analysis
Fungal pathogen preparation
The fungal artificial infection was done by highly virulent isolate of FOL. Potato Dextrose Agar (PDA) medium was used to maintain the FOL that was received from the Plant Pathology Department, Faculty of Agriculture, Cairo University. Fusarium suspensions were prepared as described by Manzo et al. Citation51 Tomato seedlings were exposed to fungal infection as explained by Manikandan and Raguchander.Citation39
Transformation and regeneration conditions
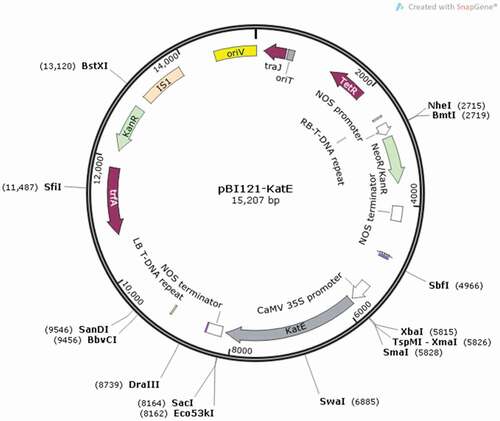
To produce genetically modified tomato plants, Agrobacterium tumefaciens LBA4404 was used in the process of plant genetic transformation using the pBI121-katE constructed binary vector, which was kindly provided by Prof. Shigeru Shigeoka, Kinki University, Japan. The T-DNA region contains the right border and expression cassettes including the neomycin phosphotransferase II (NPTII) selection marker under regulatory of nopaline synthase (nos) promoter and nopaline synthase (nosT) terminator, E. coli catalase gene (katE) under control of cauliflower mosaic virus (CaMV) 35S promoter and nopaline synthase (nosT) terminator, and the left border ().
Figure 8. Schematic illustration of the pBI121-KatE constructed binary vector; Abbreviations: RB: right border; nosP: nopaline synthase (NOS) promoter; NPT-II (KanR): neomycin phosphotransferase-II gene; nosT: nopaline synthase (NOS) terminator; 35SP: cauliflower mosaic virus (CaMV) 35S promoter; katE: E. coli catalase gene; LB: left border
Free hormones MS medium was utilized for Castle Rock and Super strain B seeds germinated. After germination, the hypocotyl was used as an explant for transformation procedure. The pBI121-katE vector was transformed into A. tumefaciens strains LBA4404 using the heat shock method as described previously.Citation52 T0 plantlets were transferred to pots and grown to maturity in a greenhouse. T1 seeds were collected and germinated in an MS medium containing 50 mg/l kanamycin. The germinated seedlings were transferred to a greenhouse and even used to get T2 seeds that were planted and used for further analysis.Citation52
Extraction and purification of genomic DNA
The mass of FOL mycelium was used to extract the fungal genomic DNA according the manufacturer's protocol of DNeasy® Plant Mini Kit (QIAGEN, cat. No. 69104, USA). Total plant DNA was extracted from modified and wild-type tomato plants according to the method of Rogers and Bendich.Citation53
PCR analysis
The ITS and katE PCRs were done using the OnePCRTm Kit (GeneDirex, cat. No. MB203-0100, Taiwan) in a 20 μl total volume consisting of template DNA (5 ng), each primer (0.4 μM), 10 μl of OnePCR pre-mixed solution (2X) and ddH2O to 20 μl total volume. The reaction program was achieved as follows: 94°C for 5 min, followed by 35 cycles of 95°C for 45 s, 52°C for 1 min for ITS region and 56°C for 1 min for katE gene, 72°C for 90 s and finally 72°C of 10 min.
DNA Dot blot analysis
DNA was extracted from both modified and non-modified plants as described by Rogers and Bendich.Citation53 The DNA was denatured from both lines at 95°C for 10 min. DNA was then spotted on a nylon membrane according to the standard method.Citation54 KatE PCR product was labeled and used as a probe. Biotin Chromogenic Detection kit was utilized for both hybridization and detection according to the manufacturer’s instructions (Ferments Life Sciences, cat. No. K0661, K0662, USA).
Reverse Transcription PCR (RT-PCR)
RT-PCR analysis was carried out to evaluate the katE gene expression in modified tomato plants. For the modified and non-modified (control or wild-type) infected plants, total RNA was purified from leaf tissue samples using Total RNA Isolation Kit (GeneDirex, Cat. No. SN020-0100, Taiwan). For cDNA synthesis, the reaction was performed using RevertAid™ First-Strand cDNA Synthesis Kit (Ferments Life Sciences, USA). The specific primer for the katE gene that listed in () was applied to the RT-PCR reaction. The final RT-PCR product was separated in 1% agarose gels and examined.
Estimation of enzyme activity
To estimate the enzyme activity as a result of fusarium infection, three different enzymes were estimated in the plant samples which are superoxide dismutase (SOD), catalase (CAT), and peroxidase (POD). Therefore, different plant leaf tissue samples were prepared as described by Chakraborty et al.Citation55 CAT was determined spectrophotometrically by measuring the decrease in absorbance at 240 nm resulting from the decomposition of H2O2 according to Aebi.Citation56 The reaction mixture was 100 mM sodium phosphate buffer (pH 7.0), 30 mM H2O2 and 100 µL of crude extract in a total volume of 1 mL. The method described by Onsa et al.Citation57 was used to determine POD activity. The reaction mixture contained 0.1 ml H2O2, 1.0 ml of 0.03 M guaiacol as a substrate, 1.8 ml of 0.1 M acetate buffer (pH 4.5), and 0.1 ml of enzyme solution. An increase in optical density at 420 nm was recorded to assay the activity. SOD enzyme activity was measured according to the method of Kumar et al.Citation58 The reaction mixture contained 10 mM EDTA, 50 mM nitroblue tetrazolium (NBT), 10 mM riboflavin, 50 mM sodium phosphate buffer (pH 7.6), 50 mM sodium carbonate, 12 mM L-methionine, and crude extract in a final volume of 1 mL. The reaction was started by adding riboflavin, which started the light-mediated reaction. Tubes were incubated at room temperature for 25 min under white light. Next, the reaction was measured at 560 nm using a spectrophotometer. One unit of enzyme activity between CAT and POD is defined as the amount of the enzyme required for reducing
of the substrate per min while, in SOD, one unit is defined as the amount of the enzyme that causing 50% inhibition of the substrate. The enzyme activities were done on four different levels (four treatments); (1) wild type (non-modified) plants, (2) wild-type plants infected with FOL, (3) T2 transgenic plants, and (4) T2 modified plants infected with the fungus.
Disease resistance analysis
The analysis was carried out as described by Abdallah et al.Citation18 The resistance against Fusarium fungus was scored in the wild type (non-modified or control), T2 of modified plants. The severity of the wilt symptoms was examined by calculating the percentage of symptomless (resistant), mild symptoms (tolerant) and susceptible plants. The symptoms and infection severity assayed 45 days after the infestation.
Statistical analysis
The test of normality distribution was done according to Shapiro and WilkCitation59 method by using SPSS v. 17.0 (2008) computer package. A randomized complete block design with two factors was used to analyze all data with three replications for each parameter. The one-way analysis of variance (ANOVA) was carried out using Graph Pad Prism 8 for Windows 10 computer software package on one factor (treatments) Dunnett’s multiple comparisons test at α = 0.05.
Acknowledgments
The authors appreciated Taif University Researchers Supporting Project number (TURSP- 2020/39), Taif University, Taif, Saudi Arabia.
Disclosure statement
The authors declare that they have no conflict of interest.
References
- Raiola A, Rigano MM, Calafiore R, Frusciante L, Barone A. Enhancing the health-promoting effects of tomato fruit for biofortified food. Mediators of Inflammation. 2014;12(3):345–350. doi:https://doi.org/10.1155/2014/139873.
- Wu X, Zhu W, Zhang H, Ding H, Zhang HJ. Exogenous nitric oxide protects against salt-induced oxidative stress in the leaves from two genotypes of tomato (Lycopersicom esculentum Mill.). Acta Physiologiae Plantarum. 2011;33(4):1199–1209. doi:https://doi.org/10.1007/s11738-010-0648-x.
- FAOSTAT. Food and Agriculture Organization of the United Nations. Rome: FAO; 2019.
- Alatar AA, Faisal M, Abdel-Salam EM, Canto T, Saquib Q, Javed SB, El-Sheikh MA, Al-Khedhairy AA. Efficient and reproducible in vitro regeneration of Solanum lycopersicum and assessment genetic uniformity using flow cytometry and SPAR methods. Saudi J Biol Sci. 2017;24(6):1430–1436. doi:https://doi.org/10.1016/j.sjbs.2017.03.008.
- Gleason ML, Edmunds BA. Tomato diseases and disorders. Iowa (USA): Iowa State University, Ames; 2006.
- Reis A, Costa H, Boiteux LS, Lopes CA. First report of Fusarium oxysporum f. sp. lycopersici race 3 on tomato in Brazil. Fitopatologia Brasileira. 2005;30(4):426–428. doi:https://doi.org/10.1590/S0100-41582005000400017.
- Sudhamoy M, Mallick N, Mitra A. Salicylic acid-induced resistance to Fusarium oxysporum f. sp. lycopersici in tomato. Plant Physiology and Biochemistry. 2009;47(7):642–649. doi:https://doi.org/10.1016/j.plaphy.2009.03.001.
- Nirmaladevi D, Venkataramana M, Srivastava RK, Uppalapati SR, Gupta VK, Yli-Mattila T, Tsui KMC, Srinivas C, Niranjana SR, Chandra NS. Molecular phylogeny, pathogenicity and toxigenicity of Fusarium oxysporum f. sp. lycopersici. Sci Rep. 2016;6:21367. doi:https://doi.org/10.1038/srep21367.
- Srinivas C, Devi DN, Murthy KN, Mohan CD, Lakshmeesha TR, Singh B, Kalagatur NK, Niranjana SR, Hashem A, Alqarawi AA, et al. Fusarium oxysporum f. sp. lycopersici causal agent of vascular wilt disease of tomato: biology to diversity– a review. Saudi Journal of Biological Sciences. 2019;26(7):1315–1324. doi:https://doi.org/10.1016/j.sjbs.2019.06.002.
- Debbi A, Boureghda H, Monte E, Hermosa R. Distribution and genetic variability of Fusarium oxysporum associated with tomato diseases in Algeria and a biocontrol strategy with indigenous Trichoderma spp. Front. Microbiol. 2018;9:282. doi:https://doi.org/10.3389/fmicb.2018.00282.
- Bowers JH, Locke JC. Effect of botanical extracts on the population density of Fusarium oxysporum in soil and control of Fusarium wilt in the greenhouse. Plant Dis. 2000;84(3):300–305. doi:https://doi.org/10.1094/PDIS.2000.84.3.300.
- Akbar A, Hussain S, Ullah K, Fahim M, Ali GS. Detection, virulence and genetic diversity of Fusarium species infecting tomato in Northern Pakistan. PLoS ONE. 2018;13(9):e0203613. doi:https://doi.org/10.1371/journal.pone.0203613.
- Hassan HA. Biology and integrated control of tomato wilt caused by Fusarium oxysporum lycopersici: a comprehensive review under the light of recent advancements. J Bot Res. 2020;3(1):84–99. doi:https://doi.org/10.36959/771/564.
- Singh R, Biswas SK, Nagar D, Singh J, Singh M, Mishra YK. Sustainable integrated approach for management of Fusarium wilt of tomato caused by Fusarium oxysporum f. sp. lycopersici (Sacc.) Synder and Hansen. Sustainable Agriculture Research. 2015;4(1):138–147. doi:https://doi.org/10.5539/sar.v4n1p138.
- Jones JW, Dayan E, Allen LH, Van Keulen H, Challa H. A dynamic tomato growth and yield model (TOMGRO). Transactions of the ASAE. 1991;34(2):663–0672. doi:https://doi.org/10.13031/2013.31715.
- Rommens CM, Kishore GM. Exploiting the full potential of disease-resistance genes for agricultural use. Curr Opin Biotechnol. 2000;11(2):120–125. doi:https://doi.org/10.1016/s0958-1669(00)00083-5.
- Dessoky ES, Ismail RM, Elarabi NI, Abdelhadi AA, Abdallah NA. Improvement of sugarcane for borer resistance using Agrobacterium mediated transformation of cry1Ac gene. GM Crops & Food. 2021;12(1):47–56. doi:https://doi.org/10.1080/21645698.2020.1809318.
- Abdallah NA, Shah D, Abbas D, Madkour M. Stable integration and expression of a plant defensin in tomato confers resistance to fusarium wilt. GM Crops. 2010;1(5):344–350. doi:https://doi.org/10.4161/gmcr.1.5.15091.
- Yoshimura S, Yamanouchi U, Katayose Y, Toki S, Wang ZX, Kono I, Kurata N, Yano M, Iwata N, Sasaki T. Expression of Xa1, a bacterial blight resistance gene in rice, is induced by bacterial inoculation. Proc Nat Acad Sci. 1998;95:1663–1668. doi:https://doi.org/10.1073/pnas.95.4.1663.
- Dolatabadi B, Ranjbar G, Tohidfar M, Dehestani A. Genetic transformation of tomato with three pathogenesis-related protein genes for increased resistance to Fusarium oxysporum f.sp. lycopersici. J Plant Mol Breed. 2014;2(1):1–11. doi:https://doi.org/10.22058/JPMB.2014.8424.
- Catanzariti A, Lim GTT, Jones DA. The tomato I-3 gene: a novel gene for resistance to Fusarium wilt disease. New Phytologist. 2015;207(1):106–118. doi:https://doi.org/10.1111/nph.13348.
- Jabeen N, Chaudhary Z, Gulfraz M, Rashid H, Mirza B. Expression of rice chitinase gene in genetically engineered tomato confers enhanced resistance to fusarium wilt and early blight. Plant Pathol J. 2015;31(3):252–258. doi:https://doi.org/10.5423/PPJ.OA.03.2015.0026.
- Bettini PP, Santangelo E, Baraldi R, Rapparini F, Mosconi P, Crinò P, Mauro ML. Agrobacterium rhizogenes rolA gene promotes tolerance to Fusarium oxysporum f. sp. lycopersici in transgenic tomato plants (Solanum lycopersicum L.). Journal of Plant Biochemistry and Biotechnology. 2016;25(3):225–233. doi:https://doi.org/10.1007/s13562-015-0328-4.
- Polidoros AN, Mylona PV, Scandalios JG. Transgenic tobacco plants expressing the maize Cat2 gene have altered catalase levels that affect plant-pathogen interactions and resistance to oxidative stress. Transgenic Research. 2001;10(6):555–569. doi:https://doi.org/10.1023/A1013027920444.
- Ossowski IV, Mulvey MR, Leco PA, Borys A, Loewen PC. Nucleotide sequence of Escherichia coli katE, which encodes catalase HPII. J Bacteriol. 1991;173(2):514–20. doi:https://doi.org/10.1128/jb.173.2.514-520.1991.
- Shikanai T, Takeda T, Yamauchi H, Sano S, Tomizawa K, Yokota A, Shigeoka S. Inhibition of ascorbate peroxidase under oxidative stress in tobacco having bacterial catalase in chloroplasts. FEBS Lett. 1998;428(1–2):47–51. doi:https://doi.org/10.1016/S0014-5793(98)00483-9.
- Nagamiya K, Motohashi T, Nakao K, Prodhan SH, Hattori E, Hirose S, Ozawa K, Ohkawa Y, Takabe T, Takabe T, et al. Enhancement of salt tolerance in transgenic rice expressing an Escherichia coli catalase gene, katE. Plant Biotechnol Rep. 2007;1:49–55. doi:https://doi.org/10.1007/s11816-007-0007-6.
- Miyagawa Y, Tamoi M, Shigeoka S. Evaluation of the defense system in chloroplasts to photooxidative stress caused by paraquat using transgenic tobacco plants expressing catalase from Escherichia coli. Plant Cell Physiol. 2000;41(3):311–320. doi:https://doi.org/10.1093/pcp/41.3.311.
- Islam MS, Azam MS, Sharmin S, Sajib A, Alam MM, Reza MS, Ahmed R, Khan H. Improved salt tolerance of jute plants expressing the katE gene from Escherichia coli. Turk J Biol. 2013;37:206–211. doi:https://doi.org/10.3906/biy-1205-52.
- Mohamed EA, Iwaki T, Munir I, Tamoi M, Shigeoka S, Wadano A. Overexpression of bacterial catalase in tomato leaf chloroplasts enhances photo-oxidative stress tolerance. Plant Cell and Environment. 2003;26:2037–2046. doi:https://doi.org/10.1046/j.0016-8025.2003.01121.x.
- Girhepuje PV, Shinde GB. Transgenic tomato plants expressing a wheat endochitinase gene demonstrate enhanced resistance to Fusarium oxysporum f. sp. lycopersici. Plant Cell Tissue Organ Cult. 2011;105(2):243–251. doi:https://doi.org/10.1007/s11240-010-9859-5.
- Ouyang B, Chen YH, Li HX, Qian CJ, Huang SL, Ye ZB. Transformation of tomatoes with osmotin and chitinase genes and their resistance to Fusarium wilt. J Hortic Sci Biotechnol. 2005;80(5):517–522. doi:https://doi.org/10.1080/14620316.2005.11511971.
- Jabeen N, Chaudhary Z, Gulfraz M, Rashid H, Mirza B. Expression of rice chitinase gene in genetically engineered tomato confers enhanced resistance to fusarium wilt and early blight. Plant Pathol J. 2015;31(3):252–258. doi:https://doi.org/10.5423/PPJ.OA.03.2015.0026.
- El-Awady M, Moghaieb REA, Haggag W, Youssef SS, El-Sharkawy AM. Transgenic canola plants over-expressing bacterial catalase exhibit enhanced resistance to Peronospora parasitica and Erysiphe polygoni. Arab J Biotechnol. 2008;11:71–84.
- Moriwaki T, Yamamoto Y, Aida T, Funahashi T, Shishido T, Asada M, Prodhan SH, Komamine A, Motohashi T. Overexpression of the Escherichia coli catalase gene, katE, enhances tolerance to salinity stress in the transgenic indica rice cultivar, BR5. Plant Biotechnol Rep. 2008;2:41–46. doi:https://doi.org/10.1007/s11816-008-0046-7.
- Limones C, Hervás A, Navas-cortés JA, Jiménez-Díaz RM, Tena M. Induction of an antioxidant enzyme system and other oxidative stress markers associated with compatible and incompatible interactions between chickpea (Cicer arietinum L.) and Fusarium oxysporum f. sp. ciceris.. Physiol Mol Plant Pathol. 2002;61:325–337. doi:https://doi.org/10.1006/pmpp.2003.0445.
- Su Y, Guo J, Ling H, Chen S, Wang S, Xu L, Que Y. Isolation of a novel peroxisomal catalase gene from sugarcane, which is responsive to biotic and abiotic stresses. PloS One. 2014;9(1):e84426. doi:https://doi.org/10.1371/journal.pone.0084426.
- Lamb C, Dixon RA. The oxidative burst in plant disease resistance. Annu Rev Plant Physiol Plant Mol Biol. 1997;48:251–275. doi:https://doi.org/10.1146/annurev.arplant.48.1.251.
- Manikandan R, Raguchander T. Fusarium oxysporum f. sp. lycopersici retardation through induction of defensive response in tomato plants using a liquid formulation of Pseudomonas fluorescens (Pf1). Eur J Plant Pathol. 2014;140:469–480. doi:https://doi.org/10.1007/s10658-014-0481-y.
- Mandal S, Mitra A, Mallick N. Biochemical characterization of oxidative burst during interaction between Solanum lycopersicum and Fusarium oxysporum f. sp. lycopersici. Physiol Mol Plant Pathol. 2008;72:56–61. doi:https://doi.org/10.1016/j.pmpp.2008.04.002.
- Laloi C, Apel K, Danon A. Reactive oxygen signalling: the latest news. Curr Opin Plant Biol. 2004;7(3):323–328. doi:https://doi.org/10.1016/j.pbi.2004.03.005.
- Huang H, Ullah F, Zhou D, Yi M, Zhao Y. Mechanisms of ROS regulation of plant development and stress responses. Front Plant Sci. 2019;10:800. doi:https://doi.org/10.3389/fpls.2019.00800.
- Apel K, Hir H. Reactive oxygen species: metabolism, oxidative stress, and signal transduction. Annu Rev Plant Biol. 2004;55:373–399. doi:https://doi.org/10.1146/annurev.arplant.55.031903.141701.
- Gechev TS, Van Breusegem F, Stone JM, Denev I, Laloi C. Reactive oxygen species as signals that modulate plant stress responses and programmed cell death. Bioessays. 2006;28:1091–1101. doi:https://doi.org/10.1002/bies.20493.
- Mittler R. ROS are good. Trends Plant Sci. 2017;22:11–19. doi:https://doi.org/10.1016/j.tplants.2016.08.002.
- Foyer CH, Noctor G. Stress-triggered redox signalling: what’s in pROSpect? Plant Cell Environ. 2016;39(5):951–964. doi:https://doi.org/10.1111/pce.12621.
- Mittler R, Vanderauwera S, Gollery M, Van Breusegem F. Reactive oxygen gene network of plants. Trends Plant Sci. 2004;9(10):490–498. doi:https://doi.org/10.1016/j.tplants.2004.08.009.
- Vellosillo T, Vicente J, Kulasekaran S, Hamberg M, Castresana C. Emerging complexity in reactive oxygen species production and signaling during the response of plants to pathogens. Plant Physiol. 2010;154(2):444–448. doi:https://doi.org/10.1104/pp.110.161273.
- White TJ, Bruns T, Lee S, Taylor J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In: Innis MA, Gelfand DH, Sninsky JJ, White TJ editors. PCR protocols: a guide to methods and applications. Vol. 38. San Diego (CA (USA)): Academic Press; 1990. p. 315–322.
- Al-Taweel K, Iwaki T, Yabuta Y, Shigeoka S, Murata N, Wadano A. A bacterial transgene for catalase protects translation of D1 protein during exposure of salt-stressed tobacco leaves to strong light. Plant Physiol. 2007. 145(1):258–265. doi:https://doi.org/10.1104/pp.107.101733
- Manzo D, Ferriello F, Puopolo G, Zoina A, D’Esposito D, Tardella L, Ferrarini A, Ercolano MR. Fusarium oxysporum f. sp. radicis-lycopersici induces distinct transcriptome reprogramming in resistant and susceptible isogenic tomato lines. BMC Plant Biol. 2016;16:53. doi:https://doi.org/10.1186/s12870-016-0740-5.
- Moghaieb REA, Ahmed DS, Abdelhadi AA, Sharaf AN. An efficient and reproducible regeneration and transformation protocol in tomato (Solanum lycopersicum L.). Australian Journal of Basic and Applied Sciences. 2015;9:411–416.
- Rogers SO, Bendich AJ. Extraction of DNA from milligram amounts of fresh, herbarium and mummified plant tissues. Plant Mol Biol. 1985;5(2):69–76. doi:https://doi.org/10.1007/BF00020088.
- Sambrook J, Fritschi EF, Maniatis T. Molecular cloning: a laboratory manual. New York (USA): Cold Spring Harbor Laboratory Press; 1989.
- Chakraborty N, Chandra S, Acharya K. Biochemical basis of improvement of defense in tomato plant against Fusarium wilt by CaCl2. Physiol Mol Biol Plants. 2017;23(3):581–596. doi:https://doi.org/10.1007/s12298-017-0450-y.
- Aebi HE. Catalase. Bergmeyer HU editor. Methods of enzymatic analysis. Weinhem (Germany): verlag Chemie. 1983. 273–286.
- Onsa GH, Bin Saari N, Selamat J, Bakar J. Purification and characterization of membrane-bound peroxidases from Metroxylon sagu. Food Chem. 2004;85(3):365–376. doi:https://doi.org/10.1016/j.foodchem.2003.07.013.
- Kumar A, Dutt S, Bagler G, Ahuja PS, Kumar S. Engineering a thermo-stable superoxide dismutase functional at subzero to >50°C, which also tolerates autoclaving. Sci Rep. 2012;2:387. doi:https://doi.org/10.1038/srep00387.
- Shapiro SS, Wilk MB. Analysis of variance test for normality (complete samples). Biometrika. 1965;52(3/4):591–611. doi:https://doi.org/10.2307/2333709.