
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.Abstract
The present work aims to evaluate the influence of varieties and malting conditions on the liquefying and sweetening capacities of germinated flours of two new maize varieties (Atp-Y and Coca-sr). Five (5) factors such as soaking time (18–42 h), vegetable salt concentration (0.5–1.2%), soaking temperature (25–41°C), germination time (80–195 h) and maturation time (17.5–42 h) were used to potentiate the liquefying and sweetening capacities of the two flours. The dehulled flours of the two maize varieties were also used as a substrate for the evaluation of these responses. The optimal flours were then subjected to physicochemical analysis. From these analyses, it follows that the responses were affected (p < 0.05) by the malting conditions, the variety of germinated flours and the variety of substrate flours. Overall, the sprouted flours respectively from the Coca-sr variety showed the highest liquefying (190 and 282.50 mm/30 s on the gruels from the Atp-Y and Coca-sr dehulled flours) and sweetening (7193.10 and 8735.63 mg/100 mL on the gruels from the Atp-Y and Coca-sr dehulled flours) potential of the two dehulled flours. A positive correlation was observed between the sweetening and liquefying potential of the germinated flours. The combination of these different flours (malted and dehulled flours) allowed to potentiate the responses up to 3.5 times compared to the previous data. The optimal conditions obtained in this study could be applied to reduce the energy and nutrient deficiency and improve the fluidity of infant formula.
1. Introduction
Malnutrition, and particularly child malnutrition, is a global public health problem as it affects nearly one in three children. It is qualified by international organizations as a child-eating disease and is defined as a nutritional state marked by a deficiency or excess in one or more nutrients (OMS, Citation2016; UNICEF, Citation2013). According to the FAO (Citation2018) report, it mainly affects children of complementary feeding age. Among these forms of malnutrition, protein-energy deficiency is the most prevalent in developing countries with an average prevalence of 11.53% from 2005 to 2018 (FAO, Citation2019) and nearly 220 million affected children under 5 years of age. Cameroon is not left behind in this situation, with nearly 35% of the child population suffering from deficiency of any kind (INS, Citation2018). In this country, the populations mainly affected are housed in the Northern, Eastern and North-Western Regions due to the multi-faceted crises they are experiencing (INS, Citation2018). Another report by the National Institute of Statistics (INS) on this situation reveals that the alarming situation is indeed related to the inadequacy of the supplemental foods that are administered to children (INS, Citation2018). Indeed, the weaning period that corresponds to the intake of exogenous ground foods to children is the most delicate in that an unbalanced diet would lead to deficiencies that could prove irreversible (Klang et al., Citation2019a, Citation2019b).
During this period, mothers most often administer nutritionally and rheologically inadequate complementary foods to children (Ndagire et al., Citation2015; Tambo et al., Citation2019a, Citation2019b). These supplementary foods made by mothers are most often derived from raw materials such as cereals, legumes, roots and tubers in combination or not (Ndagire et al., Citation2015). These different materials undergo transformation processes such as fermentation, boiling, roasting and germination, which, if not controlled, could lead to considerable nutrient loss (Dongmo et al., Citation2020; Fotso et al., Citation2020). Another problem with these foods is the high consistency that makes them almost indigestible. Indeed, during cooking, the starch in these foods (nearly 70%) swells after water absorption, giving the gruel a consistency (less than 100 mm/30 s) that is unacceptable for the child who, because of his small stomach volume, will have difficulty consuming more than 100 mL and more than three meals, thus reducing his energy intake (Elenga et al., Citation2012). The solution would therefore lie in the use of elements that could improve this energy intake as well as the sweetening of gruels.
The use of small quantities of malted cereals such as rice, sorghum, millet and maize (Klang et al., Citation2021; Tsopbeng et al., Citation2018) is proving to be an adequate solution as this methodology is very familiar to farmers. Maize, highly produced in the West Cameroon Region Region, is the one mainly used during malting. It consists of soaking the seeds in a saline solution for 12-24 h, germination usually 72-120 h followed by optional ripening and drying (Kolawole & Kolawole, Citation2015). A basic solution made off of vegetables ashes peels adding during soaking reduces soaking time and lixiviation of antinutrient like phytates (Mallemo et al., Citation2009). It also improves the malting process through activation of many enzymes as a cofactor (Djameh et al., Citation2014).
Malting leads to an improvement in the nutritional value of the cereals, notably the protein content, minerals such as calcium and magnesium, but also to the reduction of anti-nutrients such as phytates, oxalates, saponins and trypsin inhibitors (Kolawole & Kolawole, Citation2015). It also reduces the swelling power of flours by hydrolyzing the starch contained in them into simple sugars of low swelling capacity, making them suitable for use in infant food (Mbithi-Mwikya et al., Citation2000). Moreover, malting also leads to the activation of hydrolytic enzymes such as lipases, proteases and amylases, which have the capacity to improve the digestibility of the nutrients contained in the flours, thus facilitating their absorption by the immature digestive system of children (Kolawole & Kolawole, Citation2015). These can be used in the preparation of supplementary foods to reduce the consistency of gruels with high concentrations of flour dry matter (Hounhouigan et al., Citation2007). In addition to this capacity, the use of these malted cereals would also be a favourable way to improve the natural sweetness of gruels and the limitation of the use of industrial sugars not always healthy.
Nevertheless, the sweetening and liquefying capacity of these malted cereals are impacted by several parameters such as production conditions (Klang et al., Citation2020,b; Klang et al., Citation2021) and the variety of the malted cereal used (Tambo et al., Citation2019a,b; Klang et al., Citation2020). Thus, the present work was initiated to determine the influence of variety and malting conditions on the liquefying and sweetening power of sprouted flours of two new maize varieties (Atp-Y and Coca-sr).
2. Material and methods
2.1. Material
The Atp-Y and Coca-sr maize varieties used in this work were obtained from the multipurpose station of the Institute of Agricultural Research for Development (IRAD) in Dschang (West Cameroon Region) during the dry season. The purchased batches were transported to the laboratory where they were processed into a malted and dehulled maize meal. The vegetable ashes were obtained from plantain peels collected in the city of Bafoussam in the “Toungang village” district.
2.2. Methods
2.2.1. Process for obtaining unripe plantain peel ash
The procedure established by Tambo et al. (Citation2022) was used to produce plantain peel ash. The unripe peel (5 kg) was exposed continuously to the sun for 148 h (average temperature of 28°C) and then incinerated. For this purpose, a “Heraeus” muffle furnace (Model Perkin Elmer, USA) calibrated at 300°C was used for 5 h. The residues, free of all organic matter, were then ground using the ordinary mill (no name brand, Cameroon) and stored in polyethylene bags to prevent any exchange of air and moisture.
2.2.2. Production of dehulled maize flour
In order to evaluate the fluidizing and sweetening capacity of the gruels, maize kernels (1000 g each) of varieties Atp-Y and Coca-sr were processed into dehulled flour following the protocol described by Tambo et al. (Citation2019a,b). In fact, the grains were sorted, dehulled using an ordinary mill (no name brand, Cameroon), ground using a laboratory warring blender (Model POLYMIX, KINETICA, JAPAN) and sieved (Ø = 300 µm). The flour obtained was packaged in polyethylene bags and then stored in a desiccator before use.
2.2.3. Optimization of the production conditions of malted maize flour (Atp-Y and Coca-sr varieties)
2.2.3.1. Production process of malted maize flour
The transformation of maize grains into malt flours was carried out according to the method initially described by Traoré et al. (Citation2003) and modified by Tambo et al. (Citation2022). The key steps of the process and having undergone optimization were soaking time ranging from 18 to 42 h in water in the presence of vegetable salt (plantain peel ash) whose concentration varied from 0.5% to 1.2% and at soaking temperature varying between 25°C and 41°C, germination time ranging between 80 and 195 h in an incubator thermostated at 25°C by sprinkling water (1 sprinkling/day; 15 mL/100 g of kernels) and finally maturation ranging between 17.5 and 42 h. The germination process was stopped by introducing the grains at the end of the maturation in a “Venticell” oven (Heraeus, Germany) at the temperature of 45 ± 0.5°C for 45 h for drying. After drying, the grains were rendered rootless, ground with a polymixer (Model POLYMIX, KINETICA, JAPAN) and then sieved (Ø = 300 µm). The sprouted flours obtained were packed in plastic bags and stored in a desiccator before further use.
2.2.3.2. Choice of the optimization plan, factors, experimental domain and responses
The application of the response surface methodology allowed us to optimize the five selected factors through the centered composite design and with the help of Minitab 18.1 software. Preliminary tests on the limits of the domains of the different factors allowed us to select the upper and lower limits of the constraints and present them in Table .
Table 1. Definition of experimental domain
A non-exhaustive literature review also allowed us to select these factors. For the choice of the response, those retained were the content of reducing sugars and the liquefying capacity of the gruels of the two varieties. The choice of factors and study areas allowed us to generate an experimental matrix of 54 trials (divided into 32 factorial trials, 10 in the summits of the axis and 12 in the center of the area) as shown in Table S2.
2.2.3.3. Mathematical model proposal and validation of models
The second order polynomial equation was chosen to evaluate the impact of factors and their effects on responses. On the performance of the experiments, the flow velocities and sweetening capacities were taken as the experimental response (Y), I the constant, Xi and Xj are the levels of variables, ai and aj are the coefficients of the linear terms, aii are the coefficients of the quadratic terms, aij are the coefficients of the interactions terms and i or j represent the independent variables. The model is presented in EquationEquation 1(1)
(1) :
To predict and validate the responses, some parameters like determination coefficient (R2), Absolute Mean Deviation Analysis (AMDA) and Bias factor (Bf) were determined and calculated (Baranyi et al., Citation1999). For the model to be validated, the value of the determination coefficient (R2) must be greater than 70%; Absolute Mean Deviation Analysis must be equal to 0, desirability greater than 0.5 and Bias factor (Bf) must be between 0.75 and 1.25.
2.2.3.4. Evaluation of the responses
2.2.3.4.1. Gruels preparation process
The incorporation of amylases rich flours (ARF) was done during pre-cooking of the gruels (Elenga et al., Citation2012). To 15 g of dry matter of dehulled maize flour (17.13 g of flour) was added 3 g of flour rich in amylases (FRA) and the whole diluted with 25 mL of distilled water. Subsequently, 75 mL of hot distilled water was added. After 5 min of pre-cooking time, the different containers were placed in a water bath with an initial temperature of 25°C and calibrated at 99 ± 1°C where they were gradually heated. The whole was stirred until the core temperature of the slurries reached 95°C (appearance of bubbles) and then maintained at this temperature for 10 min. After cooking, the temperature of these gruels was lowered to 45 ± 1°C and then the flow velocity and reducing sugar content were evaluated.
2.2.3.4.2. Determination of flow velocity
Gruel consistency was estimated by measuring the Bostwick flow velocity (Elenga et al., Citation2012). When the gruel reached 45 ± 0.5°C, 100 mL were poured into the first compartment of the Bostwick consistometer (model ZXCON-CON3, France), the trigger (guillotine) of the apparatus was operated to release the gruel which then flowed into the second compartment. The parameter used was the distance covered by the front of the slurry in 30 s and expressed in mm/30 s.
2.2.3.4.3. Determination of reducing sugars in gruels
The content of free sugars in the gruels was determined according to the method using 3,5-dinitrosalicylic acid (Chaplin, Citation1987). To 1 mL of gruel collected, 24 mL of distilled water was added (dilution factor 1/25; v/v). Subsequently, 1 mL of the diluted sample was taken and mixed with 3 mL of DNS. The tubes were heated in a boiling water bath for 5 min and then cooled to room temperature. Subsequently, 3 mL of distilled water was added and the absorbance was read at 540 nm against the blank of each test.
2.2.4. Physico-chemical analysis of flours and optimal gruels
2.2.4.1. Determination of the free amino nitrogen content (FAN) of flours and gruels
The standard method using glycine as a reference and described by the European Brewery Convention (Citation1987) is the one used in this quantification. FANs of malt flours were extracted as described by Tambo et al. (Citation2022). For the gruels, 1 mL of each preparation was diluted 25 times before quantification of FAN. The results are calculated as mg FAN/100 g dry mass of malt, which can be obtained from the following EquationEquation 2(2)
(2) :
2.2.4.2. Determination of soluble protein content
The standard method using biuret reagent was used for the quantification of enzymatic proteins. Bovine serum albumin was used as the standard and the soluble protein content was expressed in mg.
2.2.4.3. Determination of amylolytic (diastatic power) and α-amylase activities
The diastatic power and α-amylase activity of the extracts of the optimized flours were evaluated following the procedure described by Bernfeld (Citation1955) and partially modified by Tambo et al. (Citation2022). Regarding α-amylase activity, the enzymes were extracted in CaCl2 (3.30 g/l) followed by the inactivation of β-amylase at 68°C for 15 min. Absorbance was read at 540 nm using UV-light spectrophotometer (MCL-SP-V1100, China) against a blank and enzyme activity defined as the amount of enzyme that allows the formation of 1 µmol of maltose per minute.
2.2.4.4. Proteolytic activity
Enzymatic extracts of the optimal flours obtained according to the protocol described by Tambo et al. (Citation2018) were also used for the evaluation of proteolytic activity. The method described by Garcia de Fernando and Fox (Citation1991) and using azocasein as substrate was used to detect this activity in the extracts. One unit of activity was defined as the amount of activity that gives a change of one unit of absorbance at 440 nm/min/ml of enzyme preparation.
2.2.4.5. Malt yield, malt loss and kolbach index
The method described in Embashu et al. (Citation2019) was adopted to determine the malt yield, malt loss and Kolbach index following the Equations 3, 4 and 5:
2.2.4.6. Determination of germinative capacity
Germinative capacity was determined following the method used by Ebbah et al. (Citation2015). To 90 mm petri dishes, 100 kernels maize were placed with two layers of filter paper (Whatman N° 1) wetted with 4 mL of water. The petri dishes with corn kernels were placed in an incubator at 25°C. The germinated kernels were counted and removed at 24, 48 and 72 h. The germination energy was calculated using the following equation 6.
Where: n24, n48, n72 – numbers of germinated kernels at 24, 48 and 72 h.
2.2.5. Statistical analysis
The results of the analyses carried out in three replicates were expressed as mean ± standard deviations. The analysis of variance (ANOVA) was used to determine the influence of each factor and the degree of significance of each of these effects. The significance of each factor was determined by the Fisher test. The regression equations were also subjected to the Fisher test to determine the coefficient of determination R2. The calculations were carried out using MINITAB 18.0 software (IBM, USA). The accepted confidence level was P < 0.05. Graphical representations of the contour and surface response curves of the postulated models were made using SIGMA PLOT 12.0 software. IBM SPSSTM software version 25.0 and the Duncan test were used to verify the existence of significant differences between the predicted and experimental responses within the two corn varieties. ANOVA (Analysis of variance) test at the 5% probability threshold and the Duncan test were also used to compare the means of the physico-chemical and functional properties of the flours sample using IBM SPSSTM software version 25.0
3. Results and discussion
3.1. Results
3.1.1. Modelization
The effect of malting conditions (soaking time, plant salt concentration, soaking temperature, germination and ripening time) and variety on the flow velocity and reducing sugar content of slurries of Coca-sr and Atp-Y maize varieties was also determined. From Table S2, it was found that the flow velocity and reducing sugar content varied with maize variety and enzyme source. In addition, there was a positive correlation between soluble sugar content and flow velocity regardless of the variety of maize and malt flour. Indeed, according to Yaredi et al. (Citation2016), the variation of amylolytic activity would be at the origin of the modifications observed at the level of the content of reducing sugars and the liquefying capacity which are two very important parameters in the formulation as well as the food intake of the complementary foods by the weaning age children. Similarly, there is a variation in the liquefying capacity and reducing sugar content of gruels with the source of malt flour, substrate flour and malting conditions. Indeed, Farooq et al. (Citation2005)showed that soaking and germination times stimulated enzyme development by breaking dormancy. Similarly, Yaredi et al. (Citation2016)demonstrated that a variation in germination time affected the liquefying activity of germinated corn. The liquefying capacity of the germinated maize variety Atp-Y varied from 0 mm/30 s (trials 8 and 19) to 220 mm/30 s (trial 54) and from 0 mm/30 s (trial 19) to 224 mm/30 s (trial 30) on the dehulled maize variety Coca-sr and Atp-Y respectively. For the Coca-sr sprouted maize variety, it ranged from 0 mm/30 s (trials 11, 19 and 20) to 197 mm/30 s and from 0 mm/30 s (trial 11) to 130 mm/30 s (trial 33) over the Coca-sr and Atp-Y dehulled maize, respectively. The sweetness capacity of Atp-Y sprouted maize variety ranged from 2100.34 mg/100 mL of gruel (trial 11) to 5386.21 mg/100 mL of gruel (trial 12) and from 124.13 mg/100 mL of gruel (trials 8 and 19) to 4455 mg/100 mL of gruel (trial 46) on Atp-Y and Coca-sr dehulled maize, respectively. For Coca-sr sprouted maize, it ranged from 0.00 mg/100 mL of gruel (trial 11) to 6924.14 mg/100 mL of gruel (trial 42) and from −165.52 mg/100 mL of gruel (trial 11) to 8872.41 mg/100 mL of gruel (trial 53) on Atp-Y and Coca-sr dehulled maize, respectively. It further follows from this table that the sweetening and liquefying capacities of the germinated maize variety Atp-Y were higher on the dehulled maize variety Atp-Y. In contrast, the germinated maize variety Coca-sr exhibited better liquefying capacity on dehulled maize from Coca-sr and good reducing sugar producing capacity with dehulled maize from Atp-Y. Similarly, Coca-sr variety exhibited significantly (p < 0.05) higher liquefying and gruel sweetening capacities than Atp-Y. Tambo et al. (Citation2019a,b) reported that genetic diversity in relation to the level of expression of the gene responsible for the synthesis of enzymes responsible for starch digestion would be at the origin of the difference in activity observed between the Coca-sr and Atp-Y varieties. The high sweetening and liquefying capacities of the gruels observed with the Coca-sr variety are in line with the results of Klang et al. (Citation2020) who demonstrated that the liquefying power was higher in the germinated maize flour from the Kassaï variety compared to that from the Atp-Y. Yadang et al. (Citation2013) further explained this difference by the existence of a high saccharogenic potential in the Atp-Y variety in contrast to the Coca-sr, which instead has a high amylolytic potential. The high liquefying potential of Coca-sr malted maize on the dehulled maize from Coca-sr and a good capacity to produce reducing sugars with the dehulled maize variety Atp-Y would be linked to the specificity between the amylolytic enzymes present in the variety Atp-Y and its starch. Similarly, the low liquefying and sweetening capacity observed with Coca-sr dehulled maize would be explained by its biochemical composition, especially its richness in proteins that can form a barrier at the surface of the starch, thus reducing the contact between the enzymes and the substrate (Tambo et al., Citation2019a,b). On the other hand, the phenolic compounds found in much higher proportion in this maize variety would also be responsible for the inhibition of the activity of the amylases of the Atp-Y variety; in addition to their capacity to form insoluble complexes and thus hinder the action of the amylases. In addition, a significant variation (p < 0.05) was noted in some trials between the predicted and experimental values.
3.1.2. Contribution of variables, coefficient, p-value and sums of squares of variables
The influence of the different factors on the reducing sugar content and the liquefying capacity of malt flours of Atp-Y and Coca-sr varieties are presented in Table S3. The proposed mathematical models show that the different factors increase or decrease the different responses evaluated. Irakoze et al. (Citation2010) also demonstrated the influence of soaking and germination temperatures, soaking and germination times on amylolytic activity, consistency, FAN content and extraction rate of sorghum and maize malts. It is apparent that linear effects predominantly influenced the liquefying and sweetening capacities of both maize varieties. The linear effect of germination time contributed the most to the different responses for any variety. Klang et al. (Citation2020,b) furthermore made similar observations when optimizing the liquefying powers of malted flours of two varieties of rice and maize. Moreover, the liquefying capacity of Coca-sr malted flour over dehulled flour of the same variety was significantly (p < 0.05) and positively (see sign in front of the coefficient of the variables) affected by the linear effects of germination and maturation times, the quadratic effect of plant salt concentration, the interactions of plant salt concentration-soaking temperature and soaking temperature-germination time. These observations are contrary to those of Tambo et al. (Citation2022), particularly with respect to the quadratic effect of plant salt concentration. The positive evolution of the flow velocity of Atp-Y dehulled flour was observed with the linear effects of germination and ripening time. Klang et al. (Citation2021) also observed similar effects with optimization of the flow velocity of malted rice flours. The lowering of the liquefying capacity of Coca-sr malted flour over dehulled flour from the same variety was observed with the interaction between soaking time and germination time. This decrease could be explained by a leaching of soluble proteins with amylolytic activity associated with the inactivation of these proteins during germination due to their high water content. Tambo et al. (Citation2022) also observed the same phenomena. Furthermore, Bryce et al. (Citation2010) reported that the inactivation of metabolic phenomena during germination was due to a low dissolved oxygen content in the water as a consequence of a high content. In addition to these observations, it could also be explained by the low content of reducing sugars provided by malted flour due to their uses as metabolic precursors of anabolism. Similarly, the linear and quadratic effects of soaking time and temperature, the quadratic effect of germination time, the interactions of soaking time-plant salt concentration, soaking time-germination time and plant salt concentration-germination time significantly (p < 0.05) and negatively lowered the liquefying capacity of Coca-sr malt with respect to the Atp-Y dehulled flour. As for the capacity to produce reducing sugars, it increased with the soaking time, the interactions plant salt concentration-soaking temperature, germination time-maturing time and finally soaking temperature-germination time in both dehulled flours. On the other hand, this capacity to produce reducing sugars was reduced in the Atp-Y variety with increasing soaking time and temperature, quadratic effects of soaking time, vegetable salt concentration and germination time, interaction between soaking temperature and germination time. It was also reduced in Coca-sr dehulled flour by the linear and quadratic effect of soaking temperature, the quadratic effect of germination time, the vegetable salt concentration-maturing time and germination time-maturing time interactions.
The influence of different factors on the liquefying and reducing sugar-producing capacity of Atp-Y malt showed that germination and ripening time, soaking time-maturing time, salt concentration-soaking temperature and plant salt concentration-maturing time interactions positively affected the liquefying capacity of Atp-Y malt. A reduction of this liquefying capacity was noted with the increase in the soaking temperature, the quadratic effects of the five (5) factors, the soaking time-vegetable salt concentration interactions, the soaking temperature-germination time and the soaking temperature-maturation time. The sweetness of the flour was improved by the germination and maturation times, the soaking time-plant salt concentration and plant salt concentration-germination time combinations. At high ion concentrations, there is an inhibition of amylolytic and saccharifying activities of amylases through a steric hindrance phenomenon (Tambo et al., Citation2018). Gujjaiah and Kumari (Citation2013) reported a nearly six (6) fold decrease in proteolytic activity in some cereals when soaking and germination doubled. The same is true of Nadeem et al. (Citation2017) who noted a decline in germination capacity (from 99% to 87%) of yellow corn beyond 24 h of soaking. Similarly, this parameter was improved on the Coca-sr variety by germination time, plant salt concentration-soaking temperature and plant salt concentration-maturing time combinations. An inhibition of the capacity to produce the reducing sugars of this maize malt on the two dehulled flours was observed with an increase in the soaking time, of the soaking temperature, of the quadratic effects of the five (5) factors, of the combinations soaking time-soaking temperature, soaking temperature-sprouting time, soaking time-sprouting time and finally concentration in vegetable salt-sprouting time. Indeed, a multiplication of the times of germination and maturation by two would involve the use of the reducing sugars produced at the time of germination for the production of energy necessary for anabolism. It would also be linked to the mobilization of these sugars in metabolic processes as well as to their use as precursors in the synthesis of complex carbohydrates (Gernah et al., Citation2011). It also follows from all these results that the difference observed between the two varieties in the variables that activate and inhibit their activities would be related to the pectocellulosic and nutritional composition, the speed of activation of hydrolases at the aleurone layer by gibberellins, a difference in the mechanism of action of the enzymes of the different varieties and a different composition of amino acids at the active site of these (Klang et al., Citation2021). It is also evidence that enzyme composition and class are influenced by variety (Tambo et al., Citation2022). Validation of the mathematical parameters and responses required the determination of several parameters. Coefficients of determination (R2) ranged from 81.40% to 89.03%. The values were all above the threshold value for validation of responses following this metric which is 70% (Klang et al., Citation2021; Sagu et al., Citation2015). The same is true for the adjusted coefficient of determination (R2 adj) which ranged from 70.13% to 82.38% (Goupy & Creighton, Citation2006; Joglekar & May, Citation1987). These results further show that there is a good fit between the experimental values and the values theoretically predicted by the model. It is also evident from Table S3 that the Absolute Average Deviation Analysis (AADM) and the Bias Factor (bf) have values within the range of standard values (0–0.30 for AADM and 0.75–1.25 for bf) for all the responses evaluated regardless of the variety (Baş & Boyaci, Citation2007; Dalgaard & Jørgensen, Citation1998).
3.1.3. Effect of individual factors on flow velocities and reducing sugar contents of gruels
Figure shows the influence of the different factors and variety on the different responses evaluated. From the analysis of the influence of the different variables on these responses within the two varieties, it follows that the effects observed vary from one variety to another. Steeping plays a major role in the malting process. This represents the first step in the activation process of gibberellins, hormones responsible for the lifting of seed dormancy. Steeping showed an overall positive effect on flow velocities regardless of the variety of malt and dehulled flours used. Flow velocities gradually increased with steeping to a maximum at 30 h and then decreased. The increase of the flow velocities of the gruels would be explained by the hydrolysis of the starch contained in the flours by the enzymes brought by the germinated flours. Indeed, these are activated by the oxygen contained in the absorbed soaking water. It would also be linked to a high availability of proteins with amylolytic activity following their dissociation from certain compounds (phenols, tannins, phytates and oxalates) (Djameh et al., Citation2014). The reduction of the liquefying power observed with the soaking of malted flours would be related to a reduction (loss by leaching) of the content of soluble proteins endowed with amylolytic power (Nadeem et al., Citation2017). With respect to the reducing sugar content of the grueles, a different variation was observed depending on the sources of malts and dehulled flours. An overall negative effect was observed with the combinations of Atp-Y malt and dehulled Coca-sr flours and Coca-sr malt and dehulled Atp-Y flours. The reducing sugar content of the gruels of the previous combinations dropped gradually with the soaking time and this from the first hours. This could be explained by leaching of soluble sugars, dilution of reagents, acidification of the medium following fermentation of released low molecular weight sugars (Adesokan et al., Citation2011; Djameh et al., Citation2014; Pelembe et al., Citation2004). On the other hand, the reducing sugar contents of the Atp-Y malt-Atp-Y dehulled flour and Coca-sr malt-Coca-sr dehulled flour combinations evolved gradually with soaking time until a maximum around 25 h followed by a decrease in them. Indeed, Kanensi et al. (Citation2013), showed that the diastatic power and reducing sugar content of amaranth seeds increased up to 15 h of soaking and dropped beyond.
Figure 1. Effect of individual variables on flow velocities (a, b and c, d for Coca-sr and Atp-Y respectively) and reducing sugar contents of gruels (e, f and g, h for Coca-sr and Atp-Y respectively).
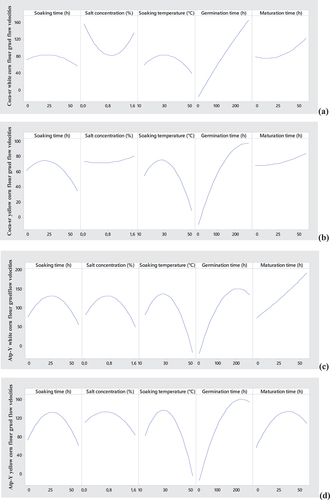
Figure 1. (Continued).
Regarding the concentration of vegetable salt, we note a constant evolution of the fluidizing power up to 0.8% followed by a decrease in the value beyond whatever the combinations. The ions provided would facilitate the fixation of starches at the active site of amylases and consequently their digestibility in low weight molecules. Fotso et al. (Citation2020) reported an improvement in the digestibility of soybean proteins when the latter underwent alkaline soaking. They explained this by the dissociation of the protein complexes, the loss of the three-dimensional structure of these facilitating the access to the bonds and especially the stabilization of the catalytic sites by these ions. This influence on the catalytic activity is concentration dependent and beyond the threshold concentration of the activator, the ions would lead to an inhibition of the activities as well as an increased leaching of the soluble sugars due to the increase in the permeability of the membranes. Similar effects were observed for the reducing sugar contents of the Atp-Y malt-Coca-sr dehulled flour and Coca-sr malt-Atp-Y dehulled flour combinations. However, there was a decrease in the reducing sugar production capacity with increasing plant salt concentration in the Atp-Y malt and Atp-Y dehulled flour gruels. The opposite effects observed in the two maize varieties are thought to be related to the different mechanisms of action of amylases and the compact molecular structure formed between proteins and polysaccharides in each of these varieties (Milala & Addy, Citation2014).
The temperature causes a weakening of the membrane of the epidermis of the seeds facilitating the absorption of water, the absorption of the ions contained in the soaking water necessary to lift the state of dormancy and the activation of soluble proteins. It affects the fluidizing and producing capacity of reducing sugars. The influence of temperature on the liquefying and producing capacity of reducing sugars had a sigmoidal effect on their main effect curves. The response maxima were found to be around 30°C for all malt varieties and combinations. These results are consistent with those of Helland et al. (Citation2002) who showed a causal relationship between steeping temperature and amylase activity. They showed that on both sides of 25°C of soaking temperature, there is a fall of the activity as a consequence of a reduction of the activation energy of the enzymes. The loss of amylase activity and the strong solubilization of reducing sugars with increasing temperature would explain the inflection observed in the different curves (Kotzia et al., Citation2012; Nguemogne et al., Citation2020).
Germination represents the limiting step in the malting process. Also, germination leads to the activation of metabolic and respiratory reactions (Djameh et al., Citation2014). This figure shows a positive correlation between germination time, liquefying and reducing sugar producing capacities. This is explained by the mobilization of reserve enzymes from all plant compartments in the degradation of carbohydrates (Tambo et al., Citation2022). Indeed, the flow velocities of the different combinations as well as the reducing sugar content of the Coca-sr malt-Coca-sr dehulled flour combination gradually evolve with germination time until stabilizing beyond 200 h. These results are similar to those of Gujjaiah and Kumari (Citation2013) who also reported improved soluble protein content, proteolytic activity, and protein digestion products after 192 h of germination. In addition, germination had a sigmoidal effect on reducing sugar contents involving corn malt variety Atp-Y and the combination Coca-sr malt-dehulled flour Atp-Y. The maxima were obtained at germination times of 200 h. The rapid and drastic reduction of the different responses beyond the peaks is in line with the results of Ratnavathi and Ravi (Citation1991) who reported a drastic decrease in the diastatic power of sorghum beyond 4 days of germination.
Maturation marks the transitional stage of malting (Tambo et al., Citation2022). It is located at the gateway to anabolism and is responsible for the mobilization of all reserve enzymes activated during the previous stages. It is shorter than germination (between 6 and 48 h) and does not require imbibition. It is also the stage of overexpression of the enzymes and of almost complete degradation of all the substances of reserve which are proteins, carbohydrates and lipids. With the exception of the reducing sugar content of the gruels made with Atp-Y malt, which showed an inflection point after 50 h of maturation, all other responses (flow velocities with both malts and reducing sugar content with Coca-sr malt) evolved gradually with maturation time. These results indicate a need for maturation of the enzymes to optimally mobilize their activities.
3.1.4. Contour plots defining the optimal zones of reducing sugars contents and gruel flow velocities according to the significant interactions
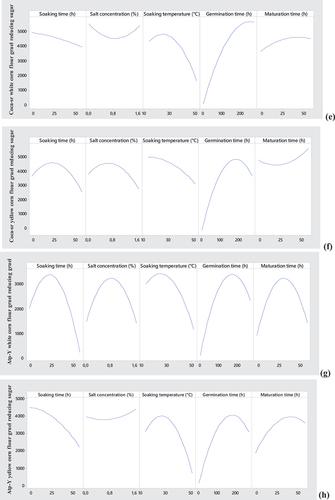
3.1.4.1. Contour plots showing the trade-offs of flow velocity and reducing sugar content of gruels mode from the combination between Atp-Y malt and Atp-Y dehulled maize
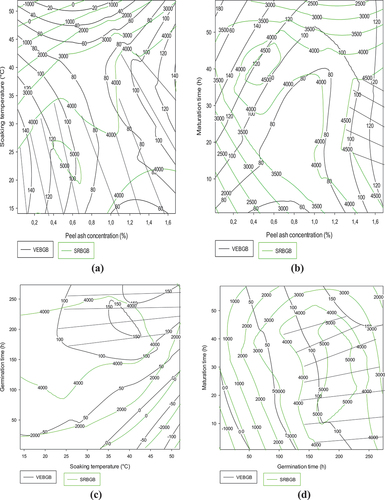
Figure shows the trade-offs between the flow velocity and the reducing sugar content of the gruels of the Atp-Y malt and Atp-Y dehulled maize combination. Both parameters are mainly influenced by the amylolytic capacity of the maize variety and the substrate specificity. In general, these figures show a positive relationship between the liquefying capacity and the reducing sugar content of the gruels. Figure shows that the flow velocity and reducing sugar content decrease with soaking time and plant salt concentration. The optimal zone was observed around 0–0.4% plant salt for less than 10 h of soaking. The significant negative effect of this combination could explain these observations. In addition, we note a positive evolution of these responses with the soaking time, whereas the temperature presents the opposite effect in Figure . A combination of soaking time and temperature allowed us to obtain flow velocities respecting the standard (100–160 mm/30 s) for reducing sugar contents of 4 g/100 mL of gruel. Regarding the combination of soaking time and maturation time, a positive evolution of these two parameters with the maturation time follows, while the optimal zone was located between 10 and 40 h of soaking. The values obtained show that the application of these conditions could allow to obtain gruels respecting the standard. A loss of sugars and proteins endowed with amylolytic activity by leaching would explain the tendency observed with soaking time. The inhibition of amylases by metal ions present in the plant could explain the trends observed with this parameter in Figures . Moreover, Figure shows a reduction of the liquefying capacity and of the reducing sugar content with the soaking temperature contrary to Figure where they increase with the germination time. Figure shows a positive correlation between the evaluated parameters and the germination time but a negative evolution with the increase in the soaking temperature.
Figure 2. Contour plots showing the trade-off areas of significant interactions on flow velocity and reducing sugar content of the Atp-Y dehulled maize Atp-Y malt mixture.
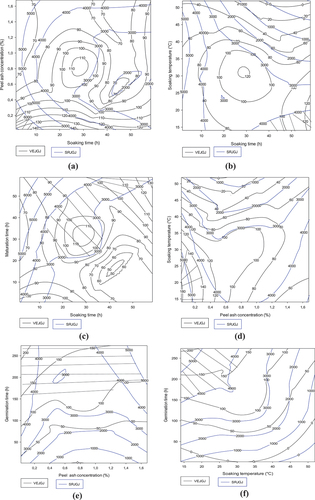
3.1.4.2. Contour plots showing the trade-off zones of flow velocity and reducing sugar content of gruels from the Coca-sr malt-Atp-Y dehulled maize mixture
Figure shows the optimal zones (hatched areas) for the flow velocity and reducing sugar content of the combination of Coca-sr malt and Atp-Y dehulled maize. Both parameters are negatively associated with steeping time but positively associated with plant salt concentration and germination time (Figures ). Indeed, high salt concentration requires short soaking times in order to limit an increased solubilization of enzymes and other molecules. Figures show, respectively, that concentrations of vegetable salt lower than 1.4% must be associated with low soaking temperatures and long germination times in order to activate large quantities of amylases which will be able to degrade the reserve carbohydrates into reducing sugars and molecules of low molecular weight with a reduced water retention capacity. Nevertheless, Figure shows that the loss of enzymes and other soluble molecules by leaching as well as the destruction of proteins under the effect of temperature would explain the need to soak the seeds at temperatures lower than 25°C for germination times of more than 200 h in order to obtain Coca-sr malts very rich in amylases. Germination and maturation are very important parameters in the process of activation and acquisition of hydrolytic power of enzymes. They are positively correlated with the fluidizing and reducing capacity of the malts. The optimal zones were marked at more than 200 h of germination with no real significant effect of ripening time (see Figure ).
Figure 3. Contour plots showing the trade-off areas of significant interactions on flow velocity and reducing sugar content of the Coca-sr-germinated Atp-Y malt gruel mixture.
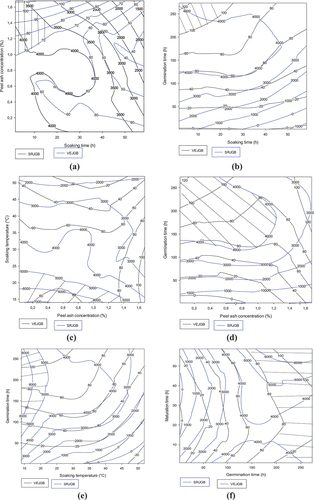
3.1.4.3. Contour plots showing trade-offs in flow velocity and reducing sugar content of slurries from the Atp-Y malt-Coca-sr dehulled maize combination
Figure shows the effect of the combination of Atp-Y malt and Coca-sr dehulled corn showed in general that the values of the optimal zones are lower than those of the Atp-Y malt-Atp-Y dehulled corn combination due to the enzyme-substrate non-specificity. It follows from graphs 4a and 4b, respectively, that the adequate steeping of maize of this variety allowing a good fluidization is between 10 and 40 h for a concentration of vegetable salt between 0.5% and 1% and a steeping at less than 40°C. Moreover, these compromise areas present values recommended by the standard for both parameters, which shows that the combination of the two varieties should be considered in areas where they are abundant in order to diversify the supplementary feeding. Notwithstanding this observation, it appears that the water content of nearly 14% of the Atp-Y variety means that it requires a short soaking for a long germination (more than 200 h) in order to activate the digestive enzymes (see Figure ). Similarly, the combination of plant salt concentration and soaking temperature, germination time and maturation time (Figures ) shows that the effect of plant salt is dependent on the combination. Indeed, at high concentrations of vegetable salt associated with a high soaking temperature (see Figure ), there is an inhibition of the fluidizing capacity of Atp-Y malt on Coca-sr dehulled maize. The opposite effect was noted with germination and maturation times due to the consumption of these metals by the plant for its physiological processes during its processes. Moreover, of these combinations, the best results were obtained with the combination of plant salt concentration and germination time. Figures show that a high soaking temperature inhibits the liquefying capacity of Atp-Y maize. Soaking at a controlled temperature followed by germination and maturation for a long time would not only compensate for the effect of soaking but also provide enzymes with high amylolytic and liquefying potential. The effect of the combination of germination and maturation times (Figure ) is similar to that reported previously (Figure ).
Figure 4. Contour plots showing the trade-off areas of significant interactions on flow velocity and reducing sugar content of the gruel made from mixture between germinated Atp-Y and dehulled Coca-sr.
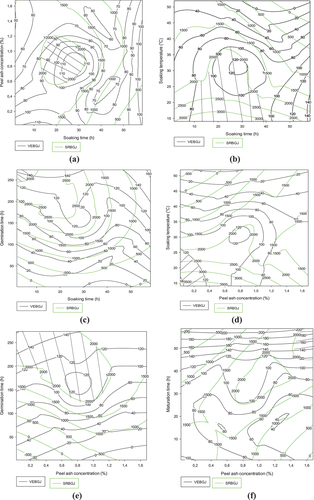
Figure 4. (Continued).
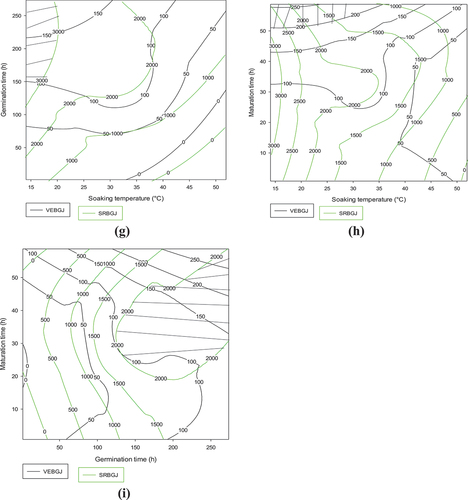
3.1.4.4. Contour plots showing the trade-off areas of flow velocity and reducing sugar content of the Coca-sr malt-Cocoa-sr dehulled maize combination
The results presented in the graphs below show in general that the Coca-sr variety requires a high steeping temperature as well as a high plant salt concentration due to its compact epidermal structure. Indeed, the highest liquefying capacity and reducing sugar content were noted when Coca-sr maize kernels were soaked between 15°C and 40°C in the presence of 0–2% vegetable salt (Figure ). In other words, the concentration of vegetable salt would not influence this ability in this variety. The same is true for graph 5b. Figure shows a gradual evolution of the responses with the soaking temperature and the germination time. The optimum values were 100–160 mm/30 s flow velocities and 4 g/100 mL reducing sugars, respectively. These results suggest that soaking at 20–40°C followed by germination for a minimum of 200 h would improve the amylolytic and fluidizing capacity of Coca-sr maize. Regarding the association of germination time and maturation time, it appears that beyond 180 h of germination for 10–50 h of maturation, the Coca-sr variety would be very capable of producing amylases capable of degrading the reserve carbohydrates into simple sugars.
Figure 5. Contour plots showing the trade-off areas of significant interactions on flow velocity and reducing sugar content of Coca-sr dehulled maize-Coca-sr malt mixture.
3.1.5. Validation trial of optimal conditions
Table shows the optimal conditions for plant salt concentration, soaking temperature, soaking time, germination and maturation of Atp-Y and Coca-sr maize kernels to obtain high flow velocities and high reducing sugar contents. These values were obtained from the manipulations done on Minitab 18.1 software. The predicted optimum values obtained from the software were compared to the experimental optimum values obtained in the laboratory. From these analyses, it appears that a soaking of 30 h, at a temperature of 18.66°C in the presence of 0.37% of vegetable salt followed by a germination during 218.38 h and a maturation of 40 h allowed to maximize the parameters evaluated in the Atp-Y maize malt. For the Coca-sr variety, it was necessary to soak during 3.90 h at a temperature of 20.57°C in the presence of 0.0219% of vegetable salt for a germination of 251.55 h and a maturation of 58.73 h. These optimal conditions show a dependence on the variety as the work of Klang et al. (Citation2021) on the flours of germinated rice seed varieties Nerica 3 and Nerica L56. They further reported a negative correlation between reducing sugars and longer soaking time for the Atp-Y variety. Klang et al. (Citation2020) also reported a strong liquefying and amylolytic capacity of the white variety of corn in contrast to the yellow variety. Desirability represents the percent reproducibility of a system and its robustness. The validation of a model requires a desirability higher than 0.5. From this table, it can be seen that all the responses evaluated within each variety were consistent with the mathematical model, thus demonstrating that the factors, responses and areas of variation for each factor were well defined. The composite desirability of the Coca-sr variety shows that the equations as well as the optimal conditions are 99% reproducible and 86% for the Atp-Y variety. These optimal conditions can be applied in malting plants as well as in the feed formulation industry to improve the quality of finished products. From the response optimization tests presented in Table S3, it appears that with the exception of the flow velocity of the Coca-sr malt/dehulled Atp-Y flour combination, all other experimental responses of the two varieties did not differ significantly (p > 0.05) from those predicted by the system. Similarly, with the exception of the flow velocity of the previous combination, all other responses were significantly (p < 0.05) elevated in the Coca-sr variety. This observation would be related to an overexpression in this variety of the gene responsible for the synthesis of amylolytic proteins especially α-amylase which is more fluidizing in contrast to β-amylase which is more saccharifying (Okoli et al., Citation2010). A difference related to soaking and germination time could further explain these observations. Indeed, Klang et al. (Citation2020,b) demonstrated that the liquefying capacity of germinated cornmeal was entirely dependent on soaking time and was maximal below 40% moisture content. These results are in contrast to those of Tsopbeng et al. (Citation2018) and Tambo et al. (Citation2019a,b) who reported a higher fluidizing capacity with the Atp-Y variety compared to the Kassaï and Coca-sr varieties. This difference could be explained by the localization of the enzymes, the richness of lipids in the Atp-Y variety that could also hinder the action of the amylases and finally the production conditions. The results obtained are also more important than those of many authors (Klang, Citation2015; Klang et al., Citation2019a,b; Klang et al., Citation2020,b; Nguemguo et al., Citation2020; Klang et al., Citation2021). These results suggest the use of the Coca-sr variety as a source of enzymes in the digestion of starches in supplemental feeds for improved acceptability, flow velocity, energy and nutrient density.
Table 2. Validation of the optimum conditions
3.1.6. Physical and chemical analysis of the optimal flours
Table shows some physical and chemical parameters of the grains and malt flours of Atp-Y and Coca-sr maize varieties. It follows from this Table that the ability of malt flours to degrade proteins to FAN during the preparation of slurries is significantly (p < 0.05) influenced by the variety of malt and dehulled maize. Indeed, the values are 315.87 and 376.67 mg/100 g for the combinations of Atp-Y dehulled maize flour and Coca-sr and Atp-Y maize malts, respectively. For the combination of Coca-sr dehulled maize, the values were 262.66 and 303.36 mg/100 g for the combinations with Atp-Y and Coca-sr malts, respectively. Similarly, it follows that the combination of Coca-sr malt and Coca-sr dehulled corn flour presented significantly (p < 0.05) higher values than those with Atp-Y malt. The observations made above would be related to an important contribution of FAN by the optimal germinated flour of the Coca-sr variety associated with an important expression of peptidase genes. Similarly, a sensitivity of the proteins of the different varieties of dehulled flours to the proteases of this variety of germinated maize would also explain the results obtained. Similarly, the values reported in this study are higher than those of Djameh et al. (Citation2014). These results further show that the Atp-Y variety has the least complex and most sensitive proteins to hydrolysis. It is also evident from this Table that maize malts were more able to release nitrogen compounds on the Atp-Y dehulled maize flour. The FAN contents of both flours were not affected (p > 0.05) by the variety and their malting conditions. The soluble protein content was 12.11 mg and 15.14 mg for the optimal Atp-Y and Coca-sr malts, respectively. The observed differences could be explained by the differences in steeping time. Indeed, Tambo et al. (Citation2022) reported that an increase in steeping time would result in the removal of soluble proteins by leaching. Nevertheless, the contents obtained are higher than those of these same authors, and this would be related to the optimal malting conditions. Concerning the amylolytic, alpha-amylolytic and proteolytic activities, the Atp-Y variety presented significantly (p < 0.05) higher values. The reduced soaking time associated with advanced germination and maturation would explain this difference. Indeed, many authors (Narsih & Harijono, Citation2012; Tsopbeng et al., Citation2018; Klang et al., Citation2020,b; Tambo et al., Citation2022) have reported that a reduced soaking time would result in a low imbibition of the grain and consequently a delay in the activation of metabolic processes. Germination capacity represents the percentage of germinated seeds when soaking conditions followed by germination are applied (Tambo et al., Citation2022). This parameter can be used to boost crop production and reduce grain loss. From this study, it was found that 85.33 and 97.33% of maize kernels of Atp-Y and Coca-sr varieties, respectively, germinated. This difference would be related to the destruction of the epidermis due to the high soaking temperature for the Atp-Y variety, the fragile structure of its epidermis and the steric hindrance of digestive enzymes or catabolic reactions in the Atp-Y variety (Ebbah et al., Citation2015). The values obtained are lower for the Atp-Y variety but similar for the Coca-sr variety to those of Ebbah et al. (Citation2015) which ranged from 94% to 96% with red sorghum. Application of the present soaking conditions to the Coca-sr variety could improve field production yields. Malt loss and yield of both varieties were also evaluated. Table shows that the mass loss is 45.49 and 42.66%, respectively, for an observed yield of 54.51 and 57.34% for Atp-Y and Coca-sr. These two parameters were, respectively, influenced by the variety. The difference obtained could be explained by the high soaking temperature of the Atp-Y variety, which made the grain more crumbly, resulting in losses (Irakoze et al., Citation2010). In addition, the losses obtained are close to those of Embashu and Nantanga (Citation2019) which varied between 11.00% and 30.40%. The Kolbach index is positively influenced by proteolytic activity and FAN content (Briggs et al., Citation2004). The variety Atp-Y presented the highest value (0.20%) compared to the 0.16% of the variety Coca-sr. These observations are contrary to those reported by Hassani et al. (Citation2013) who demonstrated that high concentrations of plant salt used during soaking weakened the epidermal membrane of cereals facilitating the loss of soluble compounds such as soluble proteins. Nevertheless, Tambo et al. (Citation2022) found similar values to those in this study. It is also evident from this Table that the moisture content as well as the water-holding capacity of the grains were 38.84% and 20.06% and 33.78% and 16.52% for the varieties Atp-Y and Coca-sr, respectively. The high values of Atp-Y variety would be explained by the high soaking time and the skeleton of its skin. Klang et al. (Citation2020,b) and Tambo et al. (Citation2022) reported an increase in moisture content of maize and rice grains with soaking time and plant salt concentration. Evaluation of the influence of plant ash addition on the pH of soaking water before and after shows that these parameters are significantly (p < 0.05) influenced by plant salt concentration, temperature and soaking time. Nevertheless, pH lowering was not significantly (p > 0.05) affected by maize variety and soaking conditions.
Table 3. Determination of the physicochemical parameters of the two optimal flours
4. Conclusion
At the end of this work, which aims to evaluate the influence of the variety and the malting conditions on the liquefying and sweetening power of the germinated flours of two new varieties of maize (Atp-Y and Coca-sr). It appears from this study that the Coca-sr variety had the best improvement of the liquefying and sweetening power, whatever the variety of flour substrate on which it was applied. Optimal malting conditions for both corn varieties resulted in flow velocities up to two times higher than the norm and reducing sugar contents of more than 3.50 g per 100 mL of gruels. The dehulled flour variety Atp-Y (yellow) exhibited the highest sensitivity to amylases and proteases of the two sprouted maize varieties. The optimal germinated flour variety Atp-Y showed the highest amylolytic and proteolytic capacities. The Atp-Y variety required a longer soaking time and a higher concentration of vegetable salt, while the opposite was observed with soaking temperature, germination and maturation times. The optimal conditions for both varieties were obtained within the range of the study. At the end of this work, it appears that the application of optimal malting conditions on these two maize varieties would boost the fight against child malnutrition in all its forms.
Authors’ contribution
Conceptualization, sample collection, preparation and design of research work (STT, JMK, OAA and HMW); execution of laboratory experiments and data collection (STT, HAKF, JOK, ABTT and MHKK); Analysis of data and Interpretation (all the authors); supervision of the work (HMW); preparation of manuscript (all the authors).
Ethical approval
This article does not contain any studies with humans or animal participants performed by any of the authors.
Originality of work
The authors declare that this work is original and has not been submitted anywhere.
Supplemental Material
Download MS Word (80.9 KB)Acknowledgments
The authors take this opportunity to express their sincere gratitude to all those who participated in this work.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Supplemental data
Supplemental data for this article can be accessed online at https://doi.org/10.1080/23311932.2023.2279724
Data availability statement
The data that supports the findings of the study are available within the manuscript.
Additional information
Funding
References
- Adesokan, I. A., Fawole, A. O., Ekanola, Y. A., Odejayi, D. O., & Olanipekun, O. K. (2011). Nutritional and sensory properties of soybean fortified composite ogi – a Nigerian fermented cereal gruel. African Journal of Microbiological Research, 5(20), 3144–24. https://doi.org/10.5897/AJMR10.907
- Baranyi, J., Pin, C., & Ross, T. (1999). Validating and comparing predictive models. International Journal of Food Microbiology, 48(3), 159–166. https://doi.org/10.1016/S0168-1605(99)00035-5
- Baş, D., & Boyaci, I. H. (2007). Modeling and optimization i: Usability of response surface methodology. Journal of Food Engineering, 78(3), 836–845. https://doi.org/10.1016/j.jfoodeng.2005.11.024
- Bernfeld, P. (1955). Amylase alpha in calomic SP Kaplan NO. Methods in enzymology (Vol. 1). Academic Press.
- Briggs, D. E., Boulton, C. A., Brookes, P. A., & Stevens, R. (2004). Brewing science and practice. Woodhead Publishing Limited.
- Bryce, H., Goodfellow, V., Agu, R., Brosnan, J. T., Bringhurst, A., & Jack, F. (2010). Effect of different steeping conditions on endosperm modification and quality of distilling malt. Journal of the Institute of Brewing, 116(2), 125–133. https://doi.org/10.1002/j.2050-0416.2010.tb00408.x
- Chaplin, M. F. (1987). Monosaccharides. In M. F. Chaplin et & J. F. Kennedy (Eds.), Carbohydrate analysis a practical approach (pp. 1–13). IRL Press.
- Dalgaard, P., & Jørgensen, L. V. (1998). Predicted and observed growth of listeria monocytogenes in seafood challenge tests and in naturally contaminated cold-smoked salmon. International Journal of Food Microbiology, 40(1–2), 105–115. https://doi.org/10.1016/S0168-1605(98)00019-1
- Djameh, C., Saalia, E. K., Sinayobye, E., Budu, A., Essilfie, G., Mensah-Brown, H., & Sefa-Dedeh, S. (2014). Optimization of the sorghum malting process for pito production in Ghana. Journal of Institute of Bewing, 121(1), 106–112. https://doi.org/10.1002/jib.191
- Dongmo, H., Tambo, T. S., Teboukeu, B. G., Mboukap, N. A., Fotso, S. B., Djuidje, T. M. C., & Klang, J. M. (2020). Effect of process and variety on physico-chemical and rheological properties of two new corn flour varieties (Atp and Kassaï). Journal of Agriculture and Food Research, 2, 10075. https://doi.org/10.1016/j.jafr.2020.100075
- Ebbah, L., Laryea, D., Barimah, J., & Djameh, C. (2015). Effect of steeping temperature on the quality of malt and Pito (an indigenous Ghanaian drink). Journal of the Institute of Brewing, 121(4), 518–523. https://doi.org/10.1002/jib.265
- Elenga, M., Massamba, J., & Silou, T. (2012). Effet de l’incorporation de malt de maïs sur la fluidité et la densité énergétique des bouillies de maïs-arachide destinées aux nourrissons et aux jeunes enfants. Journal of Applied Biosciences, 55, 3995–4005.
- Embashu, W., Iileka, O., & Nantanga, K. M. (2019). Namibian opaque beer: A review. Journal of the Institute of Brewing, 125(1), 4–9. https://doi.org/10.1002/jib.533
- Embashu, W., & Nantanga, K. K. M. (2019). Malts: Quality and phenolic content of pearl millet and sorghum varieties for brewing nonalcoholic beverages and opaque beers. Cereal Chemistry, 96(4), 765–774. https://doi.org/10.1002/cche.10178
- European Brewing Convention. (1987). Method 8.8.1. Ninhydrin colorimetric method (International method) (4th ed.). Analytica‐EBC, Brauerei‐und Getranke‐Rundschau.
- FAO. (2018). The future of food and agriculture: Alternative pathways to 2050. Summary version. 64pp. ( also available at http://www.fao.org/3/CA1553EN/ca1553en.pdf).
- FAO. (2019). GLOSIS – GSOCmap (v1.5.0). Global soil organic carbon map. Contributing countries. In [ online]. http://54.229.242.119/GSOCmap/.
- Farooq, M., Basra, S. M. A., Hafeez, K., & Ahmad, N. (2005). Thermal hardening: A new seed vigor enhancement tool in rice. Journal of Integrative Plant Biology, 47(2), 187–193. https://doi.org/10.1111/j.1744-7909.2005.00031.x
- Fotso, S. B., Tambo, T. S., Teboukeu, B. G., & Klang, J. M. (2020). Optimization of the reduction of phytates and trypsin inhibitors of soybeans (Glycine max L.): Effect of soaking and cooking. International Journal of Food and Nutritional Science, 7(1), 70–76. https://doi.org/10.15436/23770619.20.2900
- Garcia de Fernando, G. D., & Fox, P. F. (1991). Study of proteolysis during the processing of a dry fermented pork sausage. Meat Science, 30(4), 367–383. https://doi.org/10.1016/0309-1740(91)90045-R
- Gernah, D. I., Ariahu, C. C., & Ingbian, E. K. (2011). Effects of malting and lactic fermentation on some chemical and functional properties of maize (Zea mays). American Journal of Food Technology, 6(5), 404–412. https://doi.org/10.3923/ajft.2011.404.412
- Goupy, J., & Creighton, L. (2006). Introduction aux plans d’expériences (3rd ed.). Usine Nouvelle, Dunod.
- Gujjaiah, S., & Kumari, C. (2013). Evaluation of changes in α-amylase, β-amylase and protease during germination of cereals. International Journal of Agricultural Science and Research, 3(3), 55–62.
- Hassani, A., Zarnkow, M., & Becker, T. (2013). Fermented beverages (L.) Moench) as a raw material forSorghum bicolorInfluence of malting conditions on sorghum. Food Science and Technology International, 1–11. https://doi.org/10.1177/1082013213490710
- Helland, M., Wicklund, T., & Narvus. (2002). Effect of germination time on alpha-amylase production and viscosity of maize porridge. Food Research International, 35(23), 315–321. https://doi.org/10.1016/S0963-9969(01)00202-2
- Hounhouigan, D. J., Bougouma, B., Mouquet-Rivière, C., Fliedel, G., Mestres, C., Noël, J. M., Triande, E., & Monteiro, G. (2007). Production of good quality sorghum or millet malts for semi-industrial foods production in West Africa. Unpublished.
- INS. (2018). Rapport de l’enquête démographique et de santé (EDSC) 2018.
- Irakoze, P., Haihua, Z., Qin, L., Zhou, K., & Huiming, Z. (2010). Optimization of ultrasonic extraction of polysaccharides from Chinese malted sorghum using response surface methodology. Pakistan Journal Nutrition, 9(4), 336–342. https://doi.org/10.3923/pjn.2010.336.342
- Joglekar, A. M., & May, A. T. (1987). Product excellence through design of experiments. Cereal Foods World, 32, 857–868.
- Kanensi, O. J., Ochola, S., Gikonyo, N. K., & Makokha, A. (2013). Effect of steeping and germination on the diastatic activity and sugar content in amaranth grains and viscosity of porridge. Journal of Agriculture and Food Technology, 3(1), 1–7.
- Klang, J. (2015). Etude comparative des propriétés techno-fonctionnelles des amylases de Abrus precatorius, Burnatia enneandra et Cadaba farinosa : plantes utilisées traditionnellement pour sucrer et fluidifier les bouillies. Thèse de doctorat en Sciences Alimentaires et Nutrition, soutenue à l’ENSAI de l’Université de Ngaoundéré, Ngaoundéré, Cameroun, 223.
- Klang, J. M., Matueno, K. F. E., Tambo, T. S., Teboukeu, B. G., & Womeni, H. M. (2021). Optimization using response surface methodology (RSM) of the soaking and germination time of two rice varieties (Nerica 3 and Nerica L56) grown in the locality of Dschang (West-Cameroon). Journal of Food Science and Technology, 59(5), 1901–1911. https://doi.org/10.1007/s13197-021-05204-3
- Klang, J. M., Tambo, T. S., Doungmo, F. A., Tsopbeng, T. A. B., & Womeni, H. M. (2019a). Application of germinated corn flour on the reduction of flow velocities of the gruels made from corn, soybean, Moringa oleifera Leaf Powder and cassava. Journal of Food Processing & Technology, 10(7), 800. https://doi.org/10.4172/2157-7110.1000800
- Klang, J. M., Tambo, T. S., Matueno, K. F. E., Teboukeu, B. G., & Womeni, H. M. (2020). Optimization using response surface methodology (RSM) of the energy density of flour-based gruels of sweet cassava (Manihot esculenta Crantz) flour: Effect of the addition of two new sprouted rice varieties produced under optimal conditions (Nerica 3 and Nerica L56). NFS Journal, 19, 16–25. https://doi.org/10.1016/j.nfs.2020.04.001
- Klang, J. M., Tambo, T. S., Nguemguo, K. L. G., Teboukeu, B. G., Ndomou, H. S. C., Kohole, F. H. A., & Womeni, H. M. (2019b). Effect of bleaching and variety on the physico-chemical, functional and rheological properties of three new Irish potatoes (Cipira, Pamela and Dosa) flours grown in the locality of Dschang (West region of Cameroon). Heliyon, 5(12), e02982. https://doi.org/10.1016/j.heliyon.2019.e02982
- Klang, J. M., Wouatidem-Nanfack, S. L., Tambo, T. S., Teboukeu, B. G., & Womeni, H. M. (2020). Optimisation of the production of corn amylase flour from corn Atp and Kassaï varieties for the fluidification and energy density increase of cassava gruel. Heliyon, 6(10), e06344. https://doi.org/10.1016/j.heliyon.2020.e05344
- Kolawole, A. N., & Kolawole, A. O. (2015). Diastatic enzymes malting process optimisation of African finger millet for biotechnological uses. African Journal of Biochemistry Research, 9(6), 81–88. https://doi.org/10.5897/AJBR2015.0848
- Kotzia, G. A., Platis, D., Axarli, I. A., Chronopoulou, E. G., Karamitros, C., & Labrou, N. E. (2012). Biocatalysis, enzyme Engineering and Biotechnology. In B. K. Simpson, L. M. L. Nollet, F. Toldŕa, S. Benjakul, G. Paliyath, & Y. H. Hui (Eds.), Food biochemistry and food processing (pp. 125–166). John Wiley & Sons, Inc.
- Mallemo, R. C. J. Ogwok, P. Makokha, V. Mugampoza, E. & Ugen, M. A.(2009). Effect of banana peel-ash-extract on cooking time and acceptability of hard-to-cook beans (phaseolus vulgaris L.) in Uganda. International Journal of Tropical Agriculture and Food Systems, 3(1), 1–6. https://doi.org/10.4314/ijotafs.v3i1.50014
- Mbithi-Mwikya, S., Camp, J. V., & Huyghebaert, A. (2000). Nutrient and antinutrient changes in finger millet (Eleucine coracona) during sprouting. LWT-Food Science and Technology, 33(1), 9–14. https://doi.org/10.1006/fstl.1999.0605
- Milala, M. A., & Addy, O. E. (2014). Hydrolytic enzyme levels in malted cereals. Advances in Biochemistry, 2(5), 76–79. https://doi.org/10.11648/j.ab.20140205.14
- Nadeem, M. K., Qaswar, M., Ahmed, N., Rabnawaz, & Rasool, S. J. (2017). Effect of seed soaking time on germination of maize (Zea mays L.). PSM Biological Research, 2(1), 46–50.
- Narsih, Y., & Harijono, H. (2012). The study of germination and soaking time to improve nutritional quality of sorghum seed. International Food Research Journal, 19(4), 1429–1432.
- Ndagire, C. T., Muyonga, H. J., Manju, R., & Nakimbugwe, D. (2015). Optimized formulation and processing protocol for a supplementary bean- based composite flour. Food Science and Nutrition, 3(6), 527–538. https://doi.org/10.1002/fsn3.244
- Nguemguo, K. L. G., Tambo, T. S., & Klang, J. M. (2020). Effect of blanching, varietal difference of Irish potato flour and sprouted maize flour on energy density of gruels of three Irish potatoes varieties (Cipira, Pamela and Dosa) in dschang, West Cameroon: Optimization using response surface methodology (RSM). Asian Food Science Journal, 17(4), 28–43. https://doi.org/10.9734/afsj/2020/v17i430198
- Nguemogne, A. C., Desobgo, Z. S. C., & Nso, J. E. (2020). Optimisation de l’Extraction de la Dextrinase Limite de la Variété de Sorgho Camerounais Safrari. Journal of Food Stability, 3(2), 9–26. https://doi.org/10.36400/J.Food.Stab.3.2.2020-0016
- Okoli, E. V., Okolo, B. N., Moneke, A. N., & Ire, F. S. (2010). Effect of cultivar and germination time on amylolytic potential, extract yield and wort fermenting properties of malting sorghum. Asian Journal of Biotechnology, 2(1), 14e26. https://doi.org/10.3923/ajbkr.2010.14.26
- OMS. (2016). Rapport sur les résultats de l’OMS: budget programme 2016-2017. Organisation Mondiale de la Santé.
- Pelembe, L. A., Dewar, J., & Taylor, J. R. (2004). Effect of germination moisture and time on pearl millet malt quality–with respect to its opaque and lager beer brewing potential. Journal of the Institute of Brewing, 110(4), 320–325. https://doi.org/10.1002/j.2050-0416.2004.tb00627.x
- Ratnavathi, C., & Ravi, S. (1991). Effect of different durations of steeping and malting on the production of alpha-amylase in sorghum. Journal of Cereal Science, 14(3), 287–296. https://doi.org/10.1016/S0733-5210(09)80047-7
- Sagu, S. T., Nso, E. J., Karmakar, S., & De, S. (2015). Extraction and purification of beta amylase from stems of Abrus precatorius by three phase partitioning. Food Chemistry, 183, 144–153. https://doi.org/10.1016/j.foodchem.2015.03.028
- Tambo, T. S., Klang, J. M., Ndomou, H. S. C., Kohole, F. H. A., & Womeni, H. M. (2019b). Application of amylase rich flours of corn and sweet potato to the reduction of consistency of cassava and corn gruels. Journal of Food Processing and Preservation, 43(9), e14058. https://doi.org/10.1111/jfpp.14058
- Tambo, T. S., Klang, J. M., Ndomou, H. S. G., Teboukeu, B. G., Kohole, F. H. A., & Womeni, H. M. (2018). Characterization of crude extracts amylase flours of corn malt (Kassaï and Atp varieties) and sweet potato (Local and 1112 varieties). International Journal of Advanced Research in Biological Sciences, 5, 230–240. https://doi.org/10.22192/ijarbs.2018.05.05.024
- Tambo, T. S., Klang, J. M., Ndomou, H. S. C., Teboukeu, B. G., & Womeni, H. M. (2019a). Characterisation of corn, cassava and commercial flours: Use of amylases rich flours of germinated corn and sweet potato in the reduction of the consistency of the gruels made from these flours-influence on the nutritional and energy value. Food Science and Nutrition, 7(4), 1190–1206. https://doi.org/10.1002/fsn3.902
- Tambo, T. S., Ndinteh, T. D., Dongmo, J. R., Adebo, O. A., Kewuyemi, Y. O., Kamdem, K. M. H., Obilana, O. A., Klang, J. M., Njobeh, B. P., & Womeni, H. M. (2022). Optimization using response surface methodology of amylolytic capacity of maize Atp-Y and Coca-sr varieties: in vitro digestibility capacity, physico-chemical and functional properties of optimal sample. Journal of Agriculture and Food Research, 9, 100342. https://doi.org/10.1016/j.jafr.2022.100342
- Traoré, T., Icard-Vernière, C., Mouquet, C., Picq, C., Traoré, A., & Trèche, S. (2003). Variation de l’activité α α-amylasique et des teneurs en certains nutriments et facteurs anti-nutritionnels au cours de la préparation de farines de céréales germées par les dolotières de Ouagadougou. Voies alimentaires d’amélioration des situations nutritionnelles. Food-Based Approaches for a Healthy Nutrition, 14, 525–538.
- Tsopbeng, T. A. B., Tambo, T. S., Teboukeu, B. G., Zokou, R., & Klang, J. M. (2018). Effect of germination time on the diastatic power of maize (Coca-sr variety) and paddy rice (Nerica L 56 variety): Application of amylase rich flours and their extracts in the fluidification and improvement of the energy density of fermented maize gruel. Journal of Herbal Medicine Research, 3, 27. https://doi.org/10.28933/jhmr-2018-11-1108
- UNICEF. (2013). Children with disabilities, annual report. 52p.
- Yadang, G., Mbome Lape, I., & Ndjouenkeu, R. (2013). Changes in amylase activity, hot-paste viscosity and carbohydrates during natural fermentation of sweet potato (Ipomea batatas). AGRAR-2013. Proceedings of the 1ère conférence de la recherché africaine sur l’agriculture, l’alimentation et la nutrition, Yamoussokro, Cote d’ivoire, 4-6 juin 2013 (p. 683).
- Yaredi, P., Nabubuya, A., & Msuya, J. (2016). Effect of amylase activity in germinated maize flour on viscosity, energy and nutrient density of complementary porridge. Fifth RUFORUM Biennial Regional Conference, 17-21 October 2016, Cape Town, South Africa, 14(2), 1059. 1065.