
ABSTRACT
Late Triassic plant macro-remains are extremely rare in an equatorial belt stretching across the North of Gondwana from northern South America to Arabia. Located between the Laurussian floral province to the North and the Gondwana floral province to the South, this large area, is almost entirely lacking palaeobotanical data. Thus, this region is of special interest to our knowledge of Late Triassic floral provincialism and plant distribution. Based on impressions from ferruginous crusts, the first record of identifiable plant macro-remains from the Late Triassic (Norian-Rhaetian) Minjur Formation is described, as belonging to the bennettitalean taxon Zamites aff. persicus. This new finding at least tentatively supports palynological data, which indicated similarities between the Late Triassic (Norian) palynoflora of the Minjur Formation of the Arabian Peninsula on the southern shores of the Tethys and the Norian palynoflora of Iran, during this time located in the northern part of the Tethys.
Introduction
Triassic macrofloras from the northern regions of Gondwana (i.e. the northern parts of South America and Africa, as well as Arabia) are virtually unknown (e.g. Dobruskina Citation1994; Kustatscher et al. Citation2018). Focussing on the eastern part of the region, so far only a few plant macro-remains (mostly consisting of charred and permineralized wood, as well as dispersed cuticles) have been described in some detail from Middle Triassic (Anisian-Ladinian) deposits of Jordan (e.g. Abu Hamad et al. Citation2013, Citation2014, Citation2019). Additionally, the rare occurrence of plant macro-remains (including permineralized wood) has repeatedly been mentioned in studies focusing on other subjects related to the Triassic geology of this region (e.g. Jordan: Bandel and Salameh Citation2013; Saudi Arabia: Powers et al. Citation1966; Issautier et al. Citation2019). Besides these few, more or less detailed, reports on Triassic macrofloras mentioned above, as well as palynological studies dealing with early Triassic deposits (e.g. Eshet and Cousminer Citation1986; Eshet Citation1990; Abu Hamad Citation2004; Powell et al. Citation2016), there is a limited number of studies dealing with middle to late Triassic palynofloras from the Middle East, as well as the Arabian Peninsula, which were mostly conducted for biostratigraphic purposes (e.g. Powers et al. Citation1966; Abu Hamad Citation2004; Issautier et al. Citation2019; and citations therein).
This lack of (macro)floristic data from an equatorial (tropical) belt stretching from northern South America to Arabia across the entire North of Gondwana left a large geographical gap in our knowledge about the worldwide floral zonation during the Late Triassic (Kustatscher et al. Citation2018). This gap lies between the Gondwana floral province (comprising the Onslow and Ipswich subprovinces) and the Laurussian floral province, which can be divided into a number of subprovinces (cf. Dobruskina Citation1994; Kustatscher et al. Citation2018).
Recently, targeted fieldwork, as suggested by Kustatscher et al. (Citation2018), led to the discovery of the first determinable foliage remains from a locality within this large geographic gap. Based on this material occurring in ferruginous crusts the first record of the bennettitalean genus Zamites can be described from the Late Triassic Minjur Formation of Saudi Arabia.
Geological setting and lithostratigraphy
From the Permian to the Early Jurassic, the Arabian plate drifted from south of the equator northwards, reaching the equator by the Early Jurassic (Stampfli and Borel Citation2002; Golonka Citation2007), accompanied by a transition from semi-arid to warm-humid climate (Issautier et al. Citation2019). This climatic change contributed to a lithological variation from carbonate and evaporites to clastics. However, the whole period is marked by tectonic activity related to the pre-breakup of Pangaea (Sharland et al. Citation2001).
After an early Middle Permian rifting phase, Late Permian subsidence was mainly driven by post-rift thermal sag. During the rest of the Permo-Triassic super-cycle, the contribution of sediment loading is thought to have been the dominant mechanism driving subsidence (Bishop Citation1995), despite local structural control.
During the Early Triassic, a period of basin restriction led to a widespread accumulation of shale and evaporite deposits, i.e. the Sudair Shale, in a broad intrashelf basin extending up to the Arabian Gulf. The overlying mid-Late Triassic Jilh Formation consists of sandstone, shales with evaporites, and carbonate, which correspond to periodic clastic input, alternating with three brief marine transgressive cycles over the platform during the Ladinian and the Carnian (Issautier et al. Citation2019). The overlying facies resemble alternating intertidal carbonate flats and siliciclastic littoral deposits that display an aggrading trend. The Triassic-Jurassic boundary in Arabia witnessed a clear discontinuity which reflects a piece of evidence for the sudden lowering in the sea level as resulted of depositing fluvial to continental facies represented by the Upper Triassic Rukhman Formation in northern Saudi Arabia and its equivalent Minjur Sandstone in southern areas (Khalifa Citation2007).
The Minjur Sandstone, which hosts the plants studied here, was named for Khashm al Minjur (lat. 23° 31’ N, long 46° 10') where the lower part of the formation is exposed in surface outcrops (). The term was originally proposed by Bramkamp (unpublished report, 1950) as a replacement for Upper Es Sirr sandstone (discarded) and henceforth raised to formation rank. The type section of the Minjur Sandstone (315 m thick) extends from the top bed of the Jilh Formation (at lat. 23°34ʹ33”N, long. 46° 07’ 15” E) up to the base of the Marrat Formation in the face of Khashm al Khalta (lat. 23° 35’ 24” N, long. 46° 10’ 36” E). The Minjur Formation shows thinning north of the type locality (Powers et al. Citation1966). The boundary between the Minjur Sandstone and Marrat Formation is located at the contact of grey, friable, cross-bedded sandstone of the Minjur Sandstone below and the tan-brown limestone above (Khalifa Citation2007).
Figure 1. Simplified geological map of Saudi Arabia; based on Al-Husseini (Citation2009). Red star refers to the sampling locality.
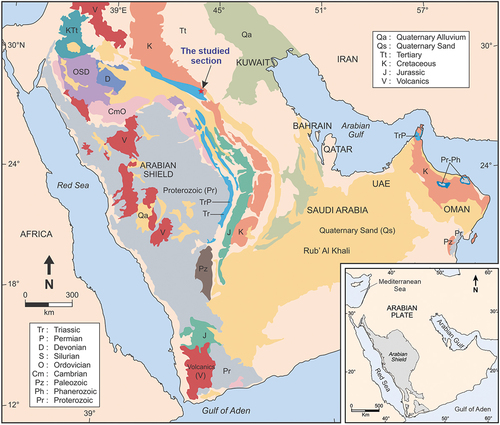
Figure 2. Field photographs of the outcrops of the Minjur Formation in Hunaydhel Village Creek in the vicinity of Hunaydhel Village, north Wadi Al-Rumah, north Al-Qassim region, Saudi Arabia and unidentifiable plant axis. a) Coarse-grained sandstone channel fill, crusted with hard ferruginous fine sandstone overlain by red sandstone, and thick mudstone capped with weathered hematite and palaeosoil. The ferruginous crusts from which the plants described here have been sampled, form a distinct feature of the landscape in this area; b) Side of Hunaydhel Village Creek with cliff-forming, friable coarse sandstones; c) ferruginous crust in situ with plant axes on the surface; d) impression of the surface of a woody log; scalebar = 1 cm; specimen SM.B 22252; e) impression of a plant axes (?) with fine, longitudinal striation; scalebar = 1 cm; specimen SM.B 22253; f) – g) three-dimensionally, but slightly flattened preserved steinkern(?) of undeterminable plant axes; scalebars = 1 cm; specimen SM.B 22254.

Lithologically, the Minjur Formation is a thick continental siliciclastic rock unit, showing minor variation in composition and essentially comprises littoral sandstone, buff, commonly cross-bedded, locally calcareous with multiple zones of red, purple, and blue-grey shale, sandy shale, and shaly sand; black to brown ironstone at several levels forms thin layers and concretions with few conglomeratic lenses that encompass stem impressions, tree trunks and silicified wood of unknown affinity (Powers et al. Citation1966; Sharief Citation1986). Moreover, it was documented from outcrop studies as an interbedded succession of sandstone and shale with minor evaporites and dolomite that were originally assigned to a dominantly fluvial-deltaic system (Le Nindre Citation1987). In surface exposures, unidentifiable fossil plant remains (i.e. axes, stems and foliage remains [the determination of such remains as Calamites foliage by Issautier et al. (Citation2019) is most likely a misidentification]) have been reported from the base and the top of the Minjur Formation (Issautier et al. Citation2019). Based on the stratigraphic position, Steineke et al. (Citation1958) considered the Minjur to be Upper Triassic or Lower Jurassic. In surface exposures, it was dated as Norian near the base by conodonts (Vrielynck et al. Citation1986). However, in subsurface sections, the palynological investigations of Jones and Hooker (Citation2011) assumed a Norian-Rhaetian age. In addition, a fourfold biostratigraphic subdivision extending from the latest Carnian-early Norian to the latest Rhaetian-Pliensbachian (Triassic-Early Jurassic), was recently suggested based on palynological studies of core samples (Issautier et al. Citation2019). Moreover, the Minjur Formation is informally subdivided into a lower and an upper unit, their depositional environments ranging from alluvial to marginal marine with tidally influenced channels (Issautier et al. Citation2019).
The lower member starts with unsorted, gravelly, cross-bedded sandstone containing fairly steep sets and a channeled surface, and displays traces of leached silty palaeosoils and ferruginization. It is topped by medium-grained, well-bedded sandstone with ferruginous crusts at various levels (Al-Husseini Citation2009). The plants analysed in the present study were retrieved from one of these crusts of the lower member of the Minjur Formation (). This fossiliferous crust consists of a lower, (non-fossiliferous) coarse-grained sandstone and an upper, very fine-grained claystone, which exhibit a thick, secondary(?) mineralization with iron minerals. Large parts of the surface of the crust, which was exposed in the outcrop, is covered with a kind of rather glossy “desert varnish”. Those ferruginous crusts are usually interbedded with mustard-yellow, coarse channel-fill sandstone. Here, we follow the dating of Vrielynck et al. (Citation1986) who dated the Minjur Formation as Norian, as this is so far the only available reliable dating for surface exposures as in our case.
Material and methods
The studied material has been collected from an outcrop in the vicinity of Hunaydhel Village (lat. 26° 59’ 62” N, long. 44° 06’ 19” E), north Wadi Al-Rumah, north Al-Qassim region, where the lower part of the Al-Minjur Formation is exposed in the form of scattered hills (). Besides the remains of Zamites the ferruginous crusts contain also abundant impressions and “steinkerns” of unidentifiable plant axes (). The specimens described here are curated in the palaeobotanical collection of the Senckenberg Forschungsinstitut und Naturmuseum Frankfurt, under accession numbers SM.B 22252 – SM.B 22257. Photographic images were taken with a NIKON Z7, with a AF-S Micro Nikkor 60 mm 1:2,8 G ED objective, a Leica M80 stereo microscope equipped with a Leica EC3 camera, as well as a Nikon SMZ 25 microscope equipped with a Nikon DS-Ri2 camera. As the surfaces of the specimens (fossils and rock), which were exposed in the outcrop, are covered by a kind of rather glossy “desert varnish” photographing the specimens proved to be rather difficult. Standard techniques which can often be applied to reduce such reflections (e.g. Kerp and Bomfleur Citation2011) did not work in this particular case: Photographs taken under liquid did not result in a reduction of the reflections on the glossy surface of the “varnish”, and photographs taken under polarized light or after whitening of the fossils, did not show enough details of the fossils to allow for the observation of most of the morphological details.
Systematic palaeontology
Order Bennettitales Engler 1892
Family Williamsoniaceae Carruthers 1870
Genus Zamites Brongniart 1828
Zamites aff. persicus Boureau 1950 ()
Figure 3. Zamites aff. persicus from the Minjur Formation. a) Zamites aff. persicus; scalebar = 1 cm; specimen SM.B 22255; b) Zamites aff. persicus, exhibiting the longest pinnae observed so far in the material from Saudi Arabia; scalebar = 1 cm; specimen SM.B 22256.

Figure 4. Zamites aff. persicus from the Minjur Formation in adaxial view exhibiting constricted pinnae bases. Enlargement from ; scalebar = 1 cm; specimen SM.B 22255.

Figure 5. Zamites aff. persicus from the Minjur Formation in abaxial view, bases of pinnae thus covered partly by the rachis. Enlargement from ; arrow points to basally overlapping pinnae; scalebar = 1 cm; specimen SM.B 22255.

Figure 6. Zamites aff. persicus from the Minjur Formation. a) Overview of a constricted pinna base and the rachis with transverse “wrinkels” (arrow); scalebar = 2 mm; specimen SM.B 22255; b) detail of the basal venation of a pinna, arrow points to a bifurcation; scalebar = 2 mm; specimen SM.B 22255.

Figure 7. Detail of the surface of a pinna of Zamites aff. persicus exhibiting impressions of the epidermal cells, arranged in longitudinal rows, running parallel to the veins (arrows); scalebar = 1 mm; specimen SM.B 22255.

Nomenclatural remark
Boureau (in Boureau et al. Citation1950) described the new Zamites species Z. persica from Triassic-Jurassic strata of the Shemshak region in Iran. According to §62.4 of the International Code of Nomenclature for algae, fungi, and plants (Turland et al. Citation2018), all plant genera ending with – ites are treated as masculine. Thus, the specific epithet of this taxon is here corrected to persicus
Description
Fragments of pinnate fronds, with base and apex of fronds missing (). Pinnae arranged oppositely to sub-oppositely or even alternating (). Bases of pinnae sometimes overlapping (). Insertion angles between rachis and pinnae vary from 41° to 76°, decreasing apically (). Distance between insertion points of pinnae varies from ~6.5 mm to 11 mm (). Base of pinnae more or less rounded (constricted), symmetric, latero-adaxially attached with a relatively small area of attachment (approx. 2.5 mm wide) (). Pinnae apex not preserved. Length of the most complete pinnae ranges from ~4 to >8 cm, width from ~6 to 10.5 mm (). Length to width ratio varies from ~5.8 to 8. Venation parallel, individual veins bifurcating only near the base of the pinnae (). Venation density ~37–42 veins/cm. On the adaxial surfaces of various pinnae, more or less isodiametric epidermal cells are visible, which are arranged in longitudinal rows (). No details of the stomata, etc., could be detected. Rachis with slight longitudinal striation and transverse “wrinkles” (), diameter of the rachis ~1.8–2.2 mm.
Remarks
The affiliation to the genus Zamites is based on the latero-adaxially attachment of the pinnae with a more or less rounded (constricted) and symmetric base (see Watson and Sincock Citation1992; Schweitzer and Kirchner Citation2003). Many Zamites species have been described from numerous Mesozoic, mainly Jurassic, deposits of the northern hemisphere (Zijlstra and van Konijnenburg-van Cittert Citation2020). Only few Zamites taxa are known from the Middle and Late Triassic (e.g. Ash Citation1975; Dobruskina Citation1994; Lucas Citation2001; Schweitzer and Kirchner Citation2003; Kustatscher et al. Citation2018). The best-known Triassic taxa occur in Iran and North America (e.g. Boureau et al. Citation1950; Ash Citation1975; Fakhr Citation1977; Schweitzer and Kirchner Citation2003; Vaez-Javadi Citation2006), whereas for other taxa from Europe and Asia the intra-specific variability is less well known.
Two Late Triassic Zamites species are known from the Norian–Rhaetian Shemshak Formation in Iran (e.g. Boureau et al. Citation1950; Corsin and Stampfli Citation1977; Fakhr Citation1977; Schweitzer and Kirchner Citation2003; Vaez-Javadi Citation2006). These species, Zamites persicus Boureau 1950 and Zamites boureaui (Fakhr Citation1977), are morphologically very similar to the material analysed here. However, Schweitzer and Kirchner (Citation2003) synonymized Zamites boureaui with Zamites persicus based on Iranian specimens that exhibited characters originally used to differentiate between both taxa. Zamites persicus (Zamites boureaui) is characterized by the following characters according to Schweitzer and Kirchner (Citation2003), those characters that agree with the material described here are printed in bold:
1. Rachis with slight longitudinal striation and transverse “wrinkles”.
2. Length to width ratio of pinnae up to 18x.
3. Pinnae apices rounded to slightly acute.
4. Distance between insertion points of pinnae on the rachis ranging from less than 7.5 mm to 10 mm.
5. Insertion angles of pinnae decrease from 70° in the centre of a frond to 45° near the apex.
The pinnae dimensions, which vary considerably between individual specimens of Zamites persicus, are considered unsuitable for taxonomic purposes by Schweitzer and Kirchner (Citation2003), as these may just represent differences between sun- and shade leaves within an individual taxon. Such a differentiation into sun- and shade leaves seems unlikely considering the assumed divaricate growth form or plant architecture and common occurrence in more open habitats of many taxa belonging to Williamsoniaceae (e.g. Pott and McLoughlin Citation2014). In fact, several studies have repeatedly demonstrated that leaf size, as well as a number of other morphological characters like venation density, of many angiosperms but also gymnosperms, are sensitive to a wide variety of environmental parameters that may act as stressors for plants (e.g. Uhl and Walther Citation2000; Uhl and Mosbrugger Citation1999, Citation2002; Uhl et al. Citation2002; Zhang et al. Citation2018; and citations therein). Such environmentally influenced characters should therefore only be used with great care for taxonomic purposes, especially in extinct plant groups with no modern representatives. Additionally, it should be considered that pinnae dimensions could also be related to the age of a plant with leaves of younger plants being smaller than those of older plants, as it was suggested previously for other bennettitalean taxa (Pott et al. Citation2007; Pott and Krings Citation2007; and citations therein).
The material from Saudi Arabia exhibits many similarities to Zamites persicus specimens described from Iran, but due to the fragmentary preservation of the material described here many characters must remain uncertain for the time being (i.e. the form of the pinna apex) and there are even characters which differ from all Zamites persicus specimens described so far in the literature (i.e. the diameter of the rachis, which is smaller in the material described here [~2 mm] than in the type material [~4 mm] as described by Boureau et al. Citation1950). Based on our observations, the Saudi material cannot be assigned to Zamites persicus, although it seems to have a close affinity to this taxon. It seems possible that this material even represents a so far undescribed new species of the genus Zamites, but more complete and better-preserved material is needed to clarify the affiliation of this material.
The also well-known and widely distributed North American “Zamites” powellii differs considerably from the Saudi Arabian material, as this material is characterized by the occurrence of basiscopic and acroscopic auricles at the base of the broadly attached pinnae bases (e.g. Ash Citation1975). This character, which also occurs in several Jurassic taxa from Mexico, was used as one character to establish the genus Laurozamites for such material by Weber and Zamudio-Varela (Citation1995). These authors also included “Zamites” powellii into this new genus, a combination not accepted by all subsequent authors (e.g. Lucas Citation2001).
Preliminary palaeoecological and palaeobiogeographic considerations
Based on the divaricate growth form of many members of the Williamsoniaceae and the ecology of modern taxa exhibiting such a growth form, as well as the depositional environments in which many members of this family have been preserved (i.e. coastal and delta-plain settings, including floodplains), Pott and McLoughlin (Citation2014) tried to reconstruct the habitats of plants belonging to this family. These authors came to the conclusion that members of the Williamsoniaceae may have inhabited preferentially open vegetation types on floodplains in deltaic and/or coastal settings, favouring low-nutrient sites on sandy plains or in peat mires. As the Minjur Formation has been deposited under alluvial to marginal marine conditions including tidally influenced channels (Issautier et al. Citation2019), it seems possible that the habitat(s) of Zamites recovered from this formation were also within the area of an alluvial or coastal plain. However, as stated above for the taxonomic affiliation, more complete and better-preserved material is needed to evaluate such a preliminary palaeoecological interpretation.
So far Late Triassic plant macro-remains are extremely rare in an equatorial belt stretching across the North of Gondwana from northern South America to Arabia (e.g. Dobruskina Citation1994; Kustatscher et al. Citation2018). The here described plant remains come from a region that is located within this vast belt from which so far palaeobotanical and thus palaeobiogeographical data are missing () (Kustatscher et al. Citation2018). Although it is difficult to provide a meaningful palaeobiogeographic interpretation based on a single occurrence we try to provide a preliminary interpretation based not only on this new occurrence of Zamites, but also on the rare, previously published palynological data from the Minjur Formation (i.e. Powers et al. Citation1966; Issautier et al. Citation2019).
Figure 8. Palaeogeographic map showing the floral zonation of the Late Triassic and the position of the sampling locality (red star); redrawn and modified from Kustatscher et al. (Citation2018)
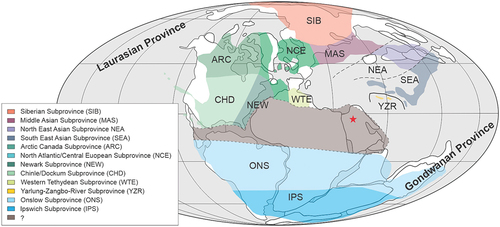
During the Norian, the composition of palynofloras from the Nayband Formation in Iran shows great affinities to Gondwana floras (e.g. Eastern Australia and New Zealand) and after the collision of the Iranian microplate with Eurasia the flora became more similar to Eurasian floras during the Rhaetian, mostly by an enrichment of cosmopolitan taxa (e.g. Cirilli et al. Citation2005). It has been noted by previous authors, that the palynoflora of the Minjur Formation also shows some similarities to the palynoflora of the Shemshak Group in Iran (Powers et al. Citation1966; Issautier et al. Citation2019). Unfortunately, the interpretation of the palynological data from the Minjur Formation is hampered by the fact that so far only selected biostratigraphic data have been published from cores and cuttings drilled by the oil industry (e.g. Powers et al. Citation1966; Issautier et al. Citation2019) and thus only limited data are available for any interpretation that goes beyond biostratigraphy.
Many different species of the genus Zamites have been described from Mesozoic strata of different continents, especially Europe, Asia, and the Americas (e.g. Zijlstra and van Konijnenburg-van Cittert Citation2020). Although the majority of these taxa has so far been described from Jurassic deposits (see Zijlstra and van Konijnenburg-van Cittert Citation2020), a few taxa were already known from Triassic strata (e.g. Dobruskina Citation1994; Kustatscher et al. Citation2018 [and citations therein]). The plants described here from Saudi Arabia exhibits a morphological affinity to Zamites persicus from the Norian-Rhaetian part of the Shemshak Group in Iran (e.g. Schweitzer and Kirchner Citation2003). Although this must be seen with great care, it seems possible that these morphological similarities also imply a more or less close “phylogenetic” relationship between Zamites persicus from the Norian-Rhaetian of Iran and Zamites aff. persicus from Saudi Arabia. This assumed relationship, together with previously published palynological data from the Late Triassic of Iran and Saudi Arabia may point to a certain biogeographic relationship between both regions and the rest of Gondwana. However, more data from plant macro-remains from the Minjur Formation will be necessary to test this preliminary interpretation and we expect that ongoing fieldwork may help to clarify this and other problems related to the Late Triassic flora of the tropics in the eastern part of Gondwana.
Data availability
Non-digital data supporting this study (i.e. fossil specimens) are curated at the palaeobotanical collection of the Senckenberg Forschungsinstitut und Naturmuseum Frankfurt, under accession numbers SM.B 22252 – SM.B 22257.
Acknowledgments
The authors gratefully thank Prof. Abdulaziz Al-Laboun (Saudi Geologists Society) for his support during fieldwork and Michael Ricker as well as Robin Kunz (Senckenberg Frankfurt) for help with taking the photographs of the specimens. H. El Atfy acknowledges financial support from Alexander von Humboldt Foundation (Germany – EGY - 1190326 - GF-P). S. Aba Alkhayl thanks the Saudi Geologists Society for their help and continuous support. The authors thank Johanna H.A. van Konijnenburg-van Cittert and Christian Pott for their constructive reviews of an earlier version of the manuscript, which helped to improve the manuscript.
Disclosure statement
No potential conflict of interest was reported by the authors.
Additional information
Funding
References
- Abu Hamad, AMB. 2004. Palaeobotany and palynostratigraphy of the Permo-Triassic in Jordan. Unpublished PhD Thesis, University of Hamburg, Germany; p. 1–316.
- Abu Hamad, AMB, A Jasper, D Uhl. 2013. Charcoal remains from the Mukheiris Formation of Jordan – the first evidence of palaeowildfire from the Anisian (Middle Triassic) of Gondwana. Jordan J Earth and Environ Sci. 5(1):17–22.
- Abu Hamad, AMB, A Jasper, D Uhl. 2014. Wood remains from the Late Triassic (Carnian) of Jordan and their paleoenvironmental implications. J African Earth Sci. 95:168–174. doi:10.1016/j.jafrearsci.2014.03.011.
- Abu Hamad, AMB, B Al‑Saqarat, CV Gonçalves, R Spiekermann, A Jasper, D Uhl. 2019. The first record of Dicroidium from the Triassic palaeotropics based on dispersed cuticles from the Anisian Mukheiris Formation of Jordan. PalZ. 93(3):487–498. doi:10.1007/s12542-019-00470-1.
- Al-Husseini, M. 2009. Stratigraphic note: update to Late Triassic – Jurassic stratigraphy of Saudi Arabia for the Middle East Geologic Time Scale. GeoArabia. 14(2):145–186. doi:10.2113/geoarabia1402145.
- Ash, SR. 1975. Zamites powelli and its distribution in the Upper Triassic of North America. Palaeontogr B. 149:139–152.
- Bandel, K, E Salameh. 2013. Geologic development of Jordan – evolution of its rocks and life. Amman: University of Jordan Press; p. 1–276.
- Bishop, RS 1995. Maturation history of the lower Palaeozoic of the eastern Arabian platform. In: M I Al-Husseini, editor: Middle East Petroleum Geosciences: Selected Middle East papers from the Middle East Geoscience Conference, GEO ’94, Bahrain: Gulf PetroLink, Bahrain; p. 180–189.
- Boureau, E, R Furon, LF Rosset. 1950. Contribution a I’etude les flores Jurassiques d. Asie Mem Mus Hist Nat Paris. 30:207–242.
- Cirilli, S, N Buratti, B Senowbari-Daryan, FT Fürsich. 2005. Stratigraphy of the Upper Triassic Nayband Formation of East-Central Iran and paleoclimatological implications. Rivista Italiana di Paleontologia e Stratigrafia. 111:259–270.
- Corsin, P, G Stampfli. 1977. La formation de Shemshak dans l’Elburz Oriental (Iran). Flore - Stratigraphie - Paléogéographie. Geobios. 10(4):509–571. doi:10.1016/S0016-6995(77)80037-5.
- Dobruskina, IA. 1994. Triassic Floras of Eurasia. Österr Akad Wissensch, Schriftenreihe Erdwiss Kommiss. 10:1–422.
- Eshet, Y, HL Cousminer. 1986. Palynozonation and correlation of the Permo-Triassic succession in the Negev, Israel. Micropaleontolo. 32(3):193–214. doi:10.2307/1485617.
- Eshet, Y. 1990. The palynostratigraphy of the Permian Triassic boundary in Israel: two approaches to biostratigraphy. Israel J Earth Sci. 39:1–15.
- Fakhr, MS. 1977. Contribution a l’etude de la flore Rhéto-Liasique de la formation de Shemshak de l’Elbourz (Iran). Mémoires de la Section Des Sciences, Paris. 5:1–178.
- Golonka, J. 2007. Late Triassic and Early Jurassic paleogeography of the world: palaeogeography, Palaeoclimatology. Palaeoecology. 244(1–4):297–307. doi:10.1016/j.palaeo.2006.06.041.
- Issautier, B, Y-M Le Nindre, N Hooker, C Reid, A Memesh, S Dini. 2019. Depositional environments, age, and sequence stratigraphy of the Minjur Formation in outcrop and near subsurface—Central Saudi Arabia. In HR AlAnzi, Rahmani, RA, Steel, RJ, Soliman, OM, editors. Siliciclastic reservoirs of the Arabian plate. AAPG Memoir 116. p. 141–184.
- Jones, NS, NP Hooker. 2011. The Upper Triassic Minjur Formation: a new bio- and sequence-stratigraphic framework and its implications for hydrocarbon prospectivity, In: 9th Middle East Geosciences Conference, GEO 2010, Bahrain: GeoArabia, Abstract Part 1, v. 16, no. 1; p. 181–182.
- Kerp, H, B Bomfleur. 2011. Photography of plant fossils – new techniques, old tricks. Rev Palaeobot Palynol. 166(3–4):117–151. doi:10.1016/j.revpalbo.2011.05.001.
- Khalifa, M. 2007. Triassic-Jurassic boundary on the southern margin of Tethys: implications of facies, tectonics and volcanism. In S G Lucas, J A Spielmann, editors. The Global Triassic. New Mexico Museum of Natural History and Science Bulletin, Albuquerque. Vol. 41; p. 110–119.
- Kustatscher, E, SR Ash, E Karasev, C Pott, V Vajda, J Yu, S McLoughlin. 2018. Flora of the Late Triassic. In L Tanner, editor. The Late Triassic world – earth in a time of transition: topics in Geobiology (Vol. 46). New York: Springer; p. 545–622.
- Le Nindre, YM. 1987. Permien supérieur, Trias, Jurassique: Sédimentologie, Livre 3. Doctoral thesis, University of Pierre et Marie Curie, Paris (VI).
- Lucas, SG 2001. The Bennettitalean leaf “Zamites” powellii from the Middle Triassic Moenkopi Formation, east-central New Mexico. New Mexico Geological Society 52nd Annual Fall Field Conference Guidebook; p. 111–114.
- Pott, C, M Krings. 2007. First record of circinate vernation in bennettitalean foliage. Neues Jahrbuch für Geologie und Paläontologie Abh. 245(3):315–321. doi:10.1127/0077-7749/2007/0245-0315.
- Pott, C, JHA van Konijnenburg-van Cittert, H Kerp, M Krings. 2007. Revision of the Pterophyllum species (Cycadophytina: bennettitales) in the Carnian (Late Triassic) flora from Lunz, Lower Austria. Rev Palaeobot Palynol. 147(1–4):3–27. doi:10.1016/j.revpalbo.2007.03.005.
- Pott, C, S McLoughlin. 2014. Divaricate growth habit in Williamsoniaceae (Bennettitales): unravelling the Ecology of a key Mesozoic Plant Group. Palaeobiodiversity and Palaeoenvironments. 94(2):307–325. doi:10.1007/s12549-014-0157-9.
- Powell, JH, MH Stephenson, A Nicora, R Rettori, LM Borlenghi, MC Perri. 2016. The Permian-Triassic boundary, Dead Sea, Jordan: transitional alluvial to marine depositional sequences and biostratigraphy. Rivista Italiana di Paleontologia e Stratigrafia. 122(3):23–40.
- Powers, RW, LF Ramirez, CD Redmond, EL Elberg Jr. 1966. Sedimentary geology of Saudi Arabia in Geology of the Arabian Peninsula. U.S. Geological Survey Professional Paper 560-D, D1–D147.
- Schweitzer, H-J, M Kirchner. 2003. Die rhäto-jurassischen Floren des Iran und Afghanistans 13. Cycadophyta. III Bennettitales. Palaeontogr B. 264(1–6):1–166. doi:10.1127/palb/264/2003/1.
- Sharief, FA. 1986. Depositional environments of the Triassic system in central Saudi Arabia. Geol J. 21(4):403–420. doi:10.1002/gj.3350210406.
- Sharland, PR, R Archer, DM Casey, RB Davies, SH Hall, AP Heward, AD Horbury, MD Simmons. 2001. Arabian plate sequence stratigraphy. GeoArabia Spec Publ. 2:371.
- Stampfli, GM, GD Borel. 2002. A plate tectonic model for the Paleozoic and Mesozoic constrained by dynamic plate boundaries and restored synthetic oceanic isochrones. Earth Planet Sci Lett. 196(1–2):17–33. doi:10.1016/S0012-821X(01)00588-X.
- Steineke, M, RA Bramkamp, NJ Sander. 1958. Stratigraphic relations of Arabian Jurassic oil. In: LG Weeks, editor. Habitat of oil: AAPG Symposium, p. 1294–1329.
- Turland, NJ, JH Wiersema, FR Barrie, W Greuter, DL Hawksworth, PS Herendeen, S Knapp, W-H Kusber, L D-Z, K Marhold, et al., editors. 2018. International code of Nomenclature for algae, fungi, and plants (Shenzhen Code) adopted by the Nineteenth International Botanical Congress Shenzhen, China, July 2017. Regnum Vegetabile 159, Glashütten: Koeltz Botanical Books. doi:10.12705/Code.2018.
- Uhl, D, V Mosbrugger. 1999. Leaf venation density as a climate and environmental proxy - a critical review and new data. Palaeogeography, Palaeoclimatology, Palaeoecology. 149(1–4):15–26. doi:10.1016/S0031-0182(98)00189-8.
- Uhl, D, H Walther. 2000. Sun leaf or shade leaf? Known facts in the light of new data with implications for palaeobotany. Feddes Repertorium. 111(3–4):165–174. doi:10.1002/fedr.20001110308.
- Uhl, D, H Walther, V Mosbrugger. 2002. Leaf venation density in angiosperm leaves as related to climatic and environmental conditions – problems and potential for palaeoclimatology. Neues Jahrbuch für Geolo Paläontologie Abh. 224(1):49–95. doi:10.1127/njgpa/224/2002/49.
- Uhl, D, V Mosbrugger. 2002. Some data on the potential use of leaf venation density as a palaeoclimatic proxy in leaves from the Permocarboniferous. Neues Jahrbuch für Geolo Paläontologie Abh. 223(3):405–432. doi:10.1127/njgpa/223/2002/405.
- Vaez-Javadi, F. 2006. Plant fossil remains from the Rhaetian of Shemshak Formation, Narges-Chal area, Alborz, NE Iran. Rivista Italiana di Paleontologia e Stratigrafia. 112(3):397–416.
- Vrielynck, B, J Manivit, D Vaslet, YM Le Nindre. 1986. Conodont stratigraphy of the Jilh Formation and the base of the Minjur Sandstone: middle and Late Triassic (Central Saudi Arabia)—Identification of depositional diachronism. Saudi Arabia Deputy Ministry Minel Res Open File Rep. BRGM-OF-06-31, 13.
- Watson, J, CA Sincock. 1992. Bennettitales of the English Wealden. Monogr Palaeontogr Soc. 145:1–228.
- Weber, R, G Zamudio-Varela. 1995. Laurozamites, a new genus and new species of bennettitalean leaves from the Late Triassic of North America. Rev Mex de Cienc Geol. 12:68–93.
- Zhang, L, J Yang, Y Huang, Z Jia, Y Fang. 2018. Leaf venation variation and Phenotypic Plasticity in response to Environmental Heterogeneity in Parrotia subaequalis (H. T. Chang) R. M. Hao et H. T. Wei, An Endemic and Endangered Tree Species from China. Forests. 9(5):247. doi:10.3390/f9050247.
- Zijlstra, G, JHA van Konijnenburg-van Cittert. 2020. (2788) Proposal to conserve the name Zamites (fossil Cycadophyta: bennettitales) with a conserved type. Taxon. 69(5):1122–1123. doi:10.1002/tax.12343.