
Abstract
The common pheasant is listed as a nationally protected species in Iran because it faces many threats such as habitat destruction, pollution from pesticides and overhunting. The species’ habitat selection remains unknown in Iran; consequently, conservation planning for the species is hampered by this lack of information. In this study we used predictor variables including topographic, anthropogenic, land cover and climate and 122 occurrence points to model the distribution of common pheasant (subspecies P. c. talischensis), in Gilan province, Iran. The results showed that distance to agriculture and orchards as well as distance to plantation forests were the most important variables in predicting species distribution. Furthermore, we found that the total potential suitable habitat for the species in Gilan province is 315,990 hectares. We observed a shift of the species to higher altitudes in the province. We recommend raising awareness about the presence of the species especially among private landowners, creating a network of protected areas on private land, and changing land-use policies at the provincial level as essential measures for the conservation of the species in Gilan province. Our results can be applied to management and conservation of the species in other modified or rapidly changing landscapes.
Introduction
Natural habitats are rapidly changing worldwide; therefore, persistence of biodiversity on this changing globe is a major challenge (Ricketts et al. Citation2004; Lavorel et al. Citation2007; Priess et al. Citation2007; Turner et al. Citation2007a,Citationb). Human transformation of land cover and land use are the primary causes of biodiversity loss (Haines-Young Citation2009). As a consequence, continuous habitats are being divided into discontinuous patches, which affects population recruitment (Robinson et al. Citation1995; Smith & Hellmann Citation2002), survival (Harris Citation1984) and movement (Shirley Citation2006; Tucker et al. Citation2018) of terrestrial animal species (Suárez-Seoane et al. Citation2008; Tucker et al. Citation2018).
Figure 1. Distribution records of Phasianus colchicus talischens in Gilan province (black dots).
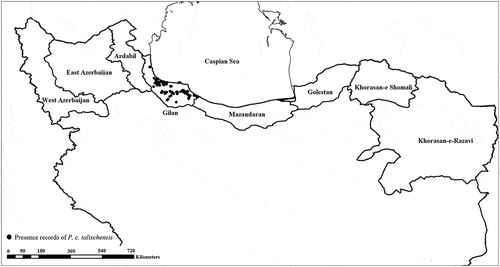
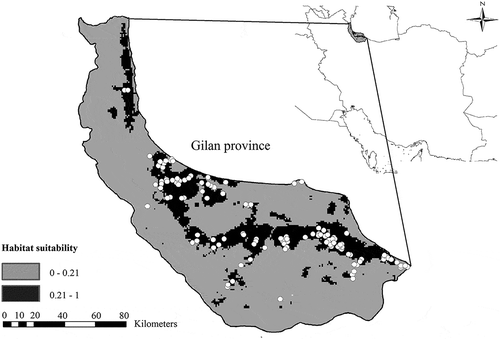
Figure 2. Habitat suitability map of Phasianus colchicus talischens in Gilan province along with distribution records (white dots).
Gilan province is one of the most rapidly changing provinces in terms of land use (forest to agricultural land) in Iran (Nohegar et al. Citation2015). As a result of land-use change, urban growth and land degradation, the distributions of some terrestrial species have changed in recent years (Ashoori Citation2009). To conserve biodiversity at a provincial scale, it is necessary to understand how recent environmental changes may affect the distribution of species.
The common pheasant (Phasianus colchicus) is one of the most widespread ground-dwelling game birds in the Palearctic region, listed as Least Concern in the International Union for Conservation of Nature Red List (IUCN Citation2016). Several studies that have been conducted on the demography of and habitat selection by pheasants in Italy showed that male territories are preferentially placed along natural or spontaneous vegetation strips (wood edges, fences, hedgerows and tree rows) and in areas characterised by high habitat diversity (Meriggi et al. Citation1996; Nelli et al. Citation2012). The presence and abundance of populations are linked to the presence of suitable vegetation for the settlement of territorial males (Nelli et al. Citation2012). Hen density, however, seems to be more linked to suitable sites for nesting, because they choose to nest out of territories defended by males (Hill & Robertson Citation1988; Nelli et al. Citation2012). Baxter et al. (Citation1996) showed that patches of smaller size that are characterised by long and irregular edges are more suitable habitats compared to larger patches with regular edges. However, the study by Nelli et al. (Citation2012) showed that pheasants select small habitat patches but with more regular edges. Sage and Tucker (Citation1998) found that nesting bird density was significantly lower in larger patches of short-rotation forestry, and highlighted the positive effect of ecotone on nesting pheasants (Nelli et al. Citation2012). Pheasants utilise marginal belts that supply them with food and protect them from predators and severe weather, due to the development of herbaceous and bushy cover (Nelli et al. Citation2012). A study by Shuai et al. (Citation2007) revealed that vegetation cover, distance to roads and slope are important variables in habitat and nest site selection by common pheasant at Taihe Nature Reserve in China. In addition, Li et al. (Citation2009) showed that topography, vegetation cover, distance to water sources and human presence are important variables in foraging habitats selection by common pheasants in Huanglong Mountains, China. Climatic variables, especially precipitation, also were recognised to affect the species’ nest survival (Geaumont et al. Citation2017).
The common pheasant is included in the list of Iran’s protected species (Ashoori Citation2009), because it faces many threats such as habitat destruction, pollution from pesticides and overhunting. The common pheasant occurs in deciduous forests, woodlands, coastal scrub, marshes and riverine forests from the Kaleibar Mountains in the north of East Azerbaijan to the valley of the Hari Rud near Sarakhs in north-eastern Khorasan-e Razavi (Scott et al. Citation1975). Phasianus colchicus has four subspecies in Iran – P. c. colchicus, P. c. talischensis, P. c. persicus and P. c. principalis – that can be distinguished from each other based on morphological characteristics (Kayvanfar & Aliabadian Citation2013; Kayvanfar et al. Citation2013, Citation2017). Phasianus c. colchicus occurs in Kalibar Mountains in North of East-Azerbaijan province, P. c. talischensis in Gilan province, P. c. persicus from west of Mazandaran province to Golestan province (Kayvanfar & Aliabadian Citation2013; Mahmoudi et al. Citation2016), and P. c. principalis in north-east Khorasan-e-Razavi province from Quchan County east to Sarakhs County (Kayvanfar & Aliabadian Citation2013; Kayvanfar et al. Citation2017). Despite this species being known in terms of morphology (Kayvanfar et al. Citation2013) and phylogeny (Kayvanfar et al. Citation2017), its habitat selection remains unknown in Iran; as a consequence, conservation planning of the species is hampered by this lack of information.
Our work is the first effort to build a robust habitat suitability model and explore the environmental drivers of common pheasant distribution in Iran. In the present study we examined the population from Gilan province, where the subspecies P. c. talischensis is present. It has been reported that the subspecies talischensis is very rare throughout its range (BirdLife International Citation2016) and has decreased in many parts of the province during recent years due to habitat destruction, land-use changes and overhunting. Because of recent rapid changes of land use in Gilan province (Ashoori Citation2009; Nohegar et al. Citation2015) we hypothesised that anthropogenic factors such as roads and human-modified land use including agricultural lands and orchards are the most important variables in shaping the distribution of P. c. talischensis in Gilan province.
Materials and methods
Study area
Gilan province (36°34ʹN to 38°27ʹN, 48°53ʹE to 50°34ʹE) is located in the north of Iran and south-west of the Caspian Sea. It is the second smallest province in Iran, with an area of 14,042.3 km2. Elevation ranges from 26 m below sea level to over 3000 m above sea level. The population of Gilan is 2,410,523, giving a population density of 171.7 inhabitants per square kilometre. The climate is humid subtropical, with up to 1850 mm annual precipitation (Molavi‐Arabshahi et al. Citation2016). Much of the province was formerly covered in dense Hyrcanian forest with many tree genera such as Persian iron wood (Parrotia persica), Caspian honey locust (Gleditsia caspica), Caucasian wingnut (Pterocaya fraxinifolia) and Silk tree (Albizia julibrissin) surviving from the last ice age (Scharnweber & Rietschel Manthery Citation2007), but only a small portion of this ecoregion remains now. The main forms of land use include livestock rearing, rice farming, tea plantations and fruit farms (Eshaghpour et al. Citation2010; Ashoori & Abdoos Citation2013).
Species distribution data and explanatory variables
In order to obtain occurrence data from the entire distribution range of the species, field observations were carried out throughout the Gilan province () between 3 April and 5 May 2015. This is the best time of year to hear calls from male common pheasants, especially in the early morning (Ashoori Citation2009). Fieldwork was conducted from 05:30 until 07:30 every morning in all 16 counties in the province, and all potential sites for the presence of the species were surveyed. Unpublished reports and old distribution maps of the species in the Department of Environment were used to identify all potential sites for the presence of the common pheasant. To make sure we covered all potential sites, we searched 3–6 stations at each potential site. At least one observer was present at each station. To avoid duplicate recording of one individual the observer did not leave the station at sampling time and could only move at a distance of 100 to 150 m around their established location. At the end of sampling time, we registered coordinates of the site by global positioning system (GPS), if we heard a calling male common pheasant or saw a male or female of the species. Due to the topographic condition of the area, the stations within potential sites were selected between 1000 and 1500 m apart.
Ecogeographical variables including land cover, topography, and anthropogenic and bioclimatic variables () were used for modeling the distribution of common pheasant. These factors are known to influence the species’ distribution (Robertson et al. Citation1993; Robertson Citation1997; Ni et al. Citation2001; Bliss Citation2004; Leif Citation2005; Shuai et al. Citation2007; Li et al. Citation2009; Geaumont et al. Citation2017). Climatic variables were obtained from the WorldClim database (Hijmans et al. Citation2005). Common pheasants are mainly observed in forests, plantation forests, rangelands, agriculture and orchards (Ashoori Citation2009). These habitats provide food and shelter for the species in the study area (Ashoori Citation2009). Land cover data were obtained from the National Land Cover Map of the Forests, Range and Watershed Management Organization of Iran (FRWOI Citation2010). These data were derived from 30 m Landsat 7 Enhanced Thematic Mapper Plus (ETM+) imagery for the conterminous Iran in 2010. The distance to human settlements (urban and rural areas) and distance to roads were included in the model because these anthropogenic variables had an influence on the distribution of birds in the study area (Ashoori Citation2009). The layer of human settlements was extracted from the National Land Cover Map of FRWOI (FRWOI Citation2010). Road data were downloaded from OpenStreetMap. All environmental variables were prepared using ArcGIS 9.3 (ESRI Citation2009).
Table I. Environmental variables used in developing a distribution model for subspecies talischensis in Gilan province.
Data analysis
In order to develop a habitat suitability model for the subspecies talischensis in Gilan, we used Maxent software, version 3.3.3.k (Phillips et al. Citation2006). The Maximum entropy model (Maxent) is one of the universal Java-based learning tools in species distribution models based on the maximum-entropy approach (Phillips et al. Citation2006, Citation2017). By using presence data of species and environmental layers, Maxent calculates the probability of occurrence of a given species in the study area (Phillips et al. Citation2006). This approach has greater predictive power compared to other models such as GARP and BIOCLIM (Elith et al. Citation2006; Pearce & Boyce Citation2006), and produces robust models when the sample size is low (Hernández et al. Citation2006; Pearson et al. Citation2007).
We ran Maxent with maximum iterations of 500, a convergence threshold of 0.0001 and 10,000 background points. We randomly partitioned occurrence records of the species into a training set with 80% of the records and the test set with 20% of the records.
The performance of the model was assessed using the area under the curve (AUC) metric of the receiving operator characteristic (ROC) curve (Phillips et al. Citation2006). The AUC is commonly used as a measure of model performance (Fourcade et al. Citation2013; Kafash et al. Citation2016, Citation2018). Models with AUC > 0.75 are considered adequate and those with AUC > 0.90 are considered excellent (Elith Citation2000). We also obtained alternative estimates of variable importance for our Maxent models by conducting a jackknife test of variable importance (Phillips et al. Citation2006).
Results
Habitat suitability modeling using 11 ecogeographical variables and 122 occurrence points indicated that 22.5% of Gilan province is suitable for the subspecies talischensis. Given that the total area of the province is 1,404,230 hectares, the total potential suitable habitat for the subspecies talischensis in Gilan province is 315,990 hectares ().
The overall predictive ability of the model (AUC = 0.906 for training and 0.842 for test data) showed high discrimination capacity in determining suitable and unsuitable habitats.
We found that agriculture and orchards, with 35% contribution, and plantation forests, with 24.4% contribution, were the most important variables in predicting the distribution of the common pheasant in Gilan province. Results showed that these two variables had a positive effect on the distribution of common pheasant. The probability of the presence of the species decreases with an increase in distance to agriculture and orchards and as well as distance to plantation forests ().
Table II. Relative contribution of each environmental variable in the subspecies talischensis model.
Jackknife analysis
Results of the jackknife test of variable importance showed that the environmental variable with the highest gain, when used in isolation, is the presence of agriculture and orchards. The environmental variable that decreases the gain the most when it is omitted is the presence of plantation forests. This means that the variable ‘plantation forests’ has the most information that is not present in the other variables ().
Table III. The results of the jackknife test of importance of variables. “Without variable” refers to when each variable is excluded in turn and a model created with the remaining variables; “with only variable” refers to when each model is constructed using only one variable.
Discussion
Species distribution models are fundamental tools for conservation and management of rare and poorly documented species (Williams et al. Citation2009; Yousefi et al. Citation2015). In addition, they help ecologists and conservation biologists to better understand the potential distribution of species and determine the most important environmental variables that shape their distribution. Despite the fact that P. c. talischensis has been described as a very rare species throughout its range (BirdLife International Citation2016), we obtained 122 distribution records of the subspecies and were able to build a robust distribution model for it in Gilan province.
Based on the Maxent model and the jackknife test of variable importance, distance to agriculture and orchards and distance to plantation forests were the most important variables in predicting the distribution of common pheasant in Gilan province. We concluded that anthropogenic variables are more important in predicting the distribution of the species than natural environmental variables. These variables (agriculture and orchards and plantation forests) are important because they provide food and shelter for the species. Agricultural lands have been reported as an important habitat requirement for other game birds in Iran, such as the Asian houbara bustard (Chlamydotis macqueenii) (Yousefi et al. Citation2017a) and little bustard (Tetrax tetrax) (Yousefi et al. Citation2017b). Here we report on the importance of orchards and plantation forests as human-modified ecosystems in shaping the distribution of common pheasant in Iran.
We also found that the species prefers forests with canopy cover of 5–25% because these forests are largely covered by shrubs and bushes, which common pheasant use as a refuge. Li et al. (Citation2009) showed that in Huanglong Mountains of China, common pheasant use habitats with less than 30% tree cover. However, habitats with more than 30% (30–50%) cover were occupied by brown-eared pheasant (Crossoptilon mantchuricum) (Li et al. Citation2009). Since the anti-predator behaviour of common pheasant is to hide (Hill & Robertson Citation1988; Robertson Citation1997; Li et al. Citation2009), the species avoids forests with canopy cover greater than 25% because it may not be possible for the species to hide in these forests due to the lack of shrubs and bushes (A. Ashoori pers. obs.). In addition, because in emergency situations the species chooses flight to avoid predators, lower tree density might facilitate take-off (Li et al. Citation2009). In conclusion, common pheasants need both forests and agricultural lands because agricultural lands provide food and forests provide shelter for the species.
Previous studies have found that topographic variables are important in species habitat selection (Shuai et al. Citation2007; Li et al. Citation2009), but in our study topography was not identified as an important predictor in species habitat selection. This may be a result of studying the species habitat selection at a different spatial scale. We studied the species’ habitat selection at a large spatial scale (Gilan province) compared to other studies, which assessed the species’ habitat selection in a relatively smaller area (Shuai et al. Citation2007; Li et al. Citation2009).
Implications for conservation and future research
Conservation of the common pheasant in a rapidly changing province in the north of Iran will be a challenging task. In Gilan, many suitable habitats located in hilly areas with a high density of common pheasants have been converted into orchards and tea plantations, while suitable habitats in the plains have been destroyed as a result of urban development and population growth, except in some private lands (Ashoori Citation2009). These changes have led to shifts of the species to higher altitudes in the province, and now they can be seen at altitudes of more than 1500 m above sea level. This was never observed in the past (Ashoori Citation2009). A similar pattern was reported for the common pheasant in Xiaoshennongjia Mountains, China (Wang et al. Citation2004). Deforestation and replanting on mountains in Xiaoshennongjia have led to a significant increase in the elevational distribution of the species (Wang et al. Citation2004). Now, in Xiaoshennongjia Mountains common pheasant is absent from low-lying areas and occurs at what appears to be an unusually high altitude (Wang et al. Citation2004).
We predict that as natural habitats decrease in Gilan province, more common pheasants will be forced to move into agricultural land and orchards. We suggest raising awareness among local landowners about the presence of common pheasants on their land. In particular, landowners should be made aware that their daily activities may put common pheasant nests and chicks at risk. We also encourage ecologically friendly farming practices for conservation of this species, for example reduced use of pesticides, and leaving rough vegetation around field edges that provides a source of insects and vegetable food in the breeding season. In addition, improving habitat quality (e.g. preserving shrub cover) would increase suitable habitats for the species. Browsing and grazing can be serious threats to germination and may hamper height growth in trees (Nasiri et al. Citation2018). This may eventually lead to an empty forest floor (Goetsch et al. Citation2011). Therefore, limiting grazing in the area is critical for retaining the understory needed for many forest species (Gharehaghaji et al. Citation2012). The creation of a network of protected areas on private land and changes in land-use policy at the provincial level are essential for the conservation of the subspecies talischensis in Gilan province.
Our results show that human-induced land-cover changes may favour the species. As the extent of the natural habitat for the common pheasant in Gilan province is decreasing, the species is shifting into the agricultural land, orchards and plantation forests that are the dominant land-use types in Gilan province. Currently, it is unknown whether moving into a new habitat has any influence on the breeding success and survival rates of the common pheasant. Further studies should therefore be carried out in the area to compare the breeding success and survival rates of common pheasants in their natural habitat and in those habitats into which they have been forced to move.
We interpreted our results in terms of habitat suitability of the subspecies talischensis, which can be reflective of species habitat suitability. Further studies on the other subspecies of the common pheasant in Iran could improve the knowledge of the habitat preferences of the species.
Acknowledgements
We would like to thank Derek Scott, Crawford Prentice, Robert Sheldon, Mauro Fasola and two anonymous reviewers for their useful comments and for improving the English of the manuscript. We also thank personnel of the Department of the Environment of Gilan province for their help during data collection.
Disclosure statement
No potential conflict of interest was reported by the authors.
References
- Ashoori A. 2009. Endangered and protected birds of Gilan province. Iran: Katibeh Gil.
- Ashoori A, Abdoos A. 2013. Important wetland habitats for the waterbirds of Gilan. Iran: Katibeh Gil.
- Baxter DA, Sage RB, Hall DO 1996. A methodology for assessing game bird use of short rotation coppice. Biomass and Bioenergy 10:301–306. DOI: 10.1016/0961-9534(95)00115-8
- BirdLife International. 2016. Species factsheet: Phasianus colchicus. Avialable: http://www.bird life.org Accessed Jul 2016 09.
- Bliss TH 2004. Habitat requirements of ring-necked pheasant hens (Phasianus colchicus) on farmland in Lower Austria during nesting and brood rearing. MSc thesis, University of Georgia.
- Elith J. 2000.Quantitative methods for modeling species habitat: comparative performance and an application to Australian plants. Quantitative methods for conservation biology. New York, USA: Springer.pp. 39–58.
- Elith J, Graham CH, Anderson RP, Dudik M, Ferrier S, Guisan A, Hijmans RJ, Huettmanm LG, Loiselle BA, Manion G, Moritz C, Nakamura M, Nakazawa Y, Overton JM, Peterson AT, Phillips SJ, Richardson K, Scachetti-Pereira R, Schapire RE, Soberon J, Williams S, Wisz MS, Zimmermann NE. 2006. Novel methods improve prediction of species distributions from occurrence data. Ecography 29: 129–151. DOI: 10.1111/j.2006.0906-7590.04596.x.
- Eshaghpour M, Saloomahalle HA, Fatehi A 2010. Hydrogeochemical studies of hot springs in Gilan Province, N-Iran. Proceedings World Geothermal Congress, Bali, Indonesia, 25–29 April 2010. pp. 22–29.
- ESRI. 2009. ArcGis 9.3. Redlands, CA: Environmental Systems Research Institute.
- Fourcade Y, Engler JO, Besnard AG, Rödder D, Secondi J. 2013. Confronting expert-based and modelled distributions for species with uncertain conservation status: A case study from the corncrake (Crex crex). Biological Conservation 167: 161–171. DOI: 10.1016/j.biocon.2013.08.009.
- FRWOI. 2010. National land cover map. Iran: Iranian Forests, Range and Watershed Management Organization.
- Geaumont BA, Sedivec KK, Schauer C. 2017. Ring- necked Pheasant use of post - conservation reserve program lands. Rangeland Ecology and Management 70: 569–575. DOI: 10.1016/j.rama.2017.04.003.
- Gharehaghaji M, Shabani AA, Feghhi J, Danehkar A, Kaboli M, Ashrafi S 2012. Effects of landscape context on bird species abundance of tree fall gaps in a temperate deciduous forest of Northern Iran. Forest Ecology and Management 267:182–189. DOI: 10.1016/j.foreco.2011.12.001
- Goetsch C, Wigg J, Royo AA, Ristau T, Carson WP 2011. Chronic over browsing and biodiversity collapse in a forest understory in Pennsylvania: Results from a 60 year-old deer exclusion plot. Journal of the Torrey Botanical Society 138:220–224. DOI: 10.3159/TORREY-D-11-00013.1
- Haines-Young R 2009. Land use and biodiversity relationships. Land Use Policy 26:178–186. DOI: 10.1016/j.landusepol.2009.08.009
- Harris LD. 1984. The fragmented Forest: Island biogeography theory and the preservation of biotic diversity. Chicago: University of Chicago Press.
- Hernández PA, Graham CH, Master LL, Albert DL. 2006. The effect of sample size and species characteristics on performance of different species distribution modeling methods. Ecography 29: 773–785. DOI: 10.1111/j.0906-7590.2006.04700.x.
- Hijmans RJ, Cameron SE, Parra JL, Jones PG, Jarvis A. 2005. Very high resolution interpolated climate surfaces for global land areas. International Journal of Climatology 25: 1965–1978. DOI: 10.1002/joc.1276.
- Hill DA, Robertson P. 1988. The pheasant: ecology, management and conservation. Oxford, UK: Blackwell Scientific Publication.
- IUCN. 2016. The IUCN Red List of Threatened Species. Version 2016-1. Available: www.iucnredlist.org. Accessed Jul 2016 10.
- Jarvis A, Reuter HI, Nelson A, Guevara E 2008. Hole-filled SRTM for the globe Version 4, available from the CGIAR-CSI SRTM 90m database. Available: http://srtm.csi.cgiar.org. Accessed Jul 2014 01.
- Kafash A, Kaboli M, Köhler G, Yousefi M, Asadi A. 2016. Ensemble distribution modeling of the Mesopotamian spiny-tailed lizard (Saara loricata) in Iran, an insight into the impact of climate change. Turkish Journal of Zoology 40: 262–271. DOI: 10.3906/zoo-1504-10.
- Kafash A, Malakoutikhahh S, Yousefi M, Ataei F, Heidari H, Rastegar-Pouyani E. 2018. Phrynocephalus scutellatus (Olivier, 1807) species complex in Iranian Plateau: The degree of niche overlap depends on the phylogenetic distance. Zoology in the Middle East 64: 47–54. DOI: 10.1080/09397140.2017.1401309.
- Kayvanfar N, Aliabadian M. 2013. Distribution, density and biological breeding of white wing pheasant (Phasianus colchicus principalis, Sclater, 1885) in Northeast of Iran. Taxonomy and Biosystematics. 5: 33–45.
- Kayvanfar N, Aliabadian M, Ghasempouri SM. 2013. Morphometric and morphological differentiation of the subspecies of Phasianus colchicus (Linnaeus, 1758) on the Iranian Plateau (Aves: Galliformes). Zoology in the Middle East. 6: 9–17.
- Kayvanfar N, Aliabadian M, Niu XJ, Zhang ZW, Liu Y. 2017. Phylogeography of the Common Pheasant Phasianus colchinus. Ibis 159: 430–442. DOI: 10.1111/ibi.12455.
- Lavorel S, Flannigan MD, Lambin EF, Scholes MC. 2007. Vulnerability of land system to fire: Interactions among humans, climate, the atmosphere, and ecosystem. Mitigation and Adaptation Strategies for Global Change 12: 1381–2386. DOI: 10.1007/s11027-006-9046-5.
- Leif AP. 2005. Spatial ecology and habitat selection of breeding male pheasants. Wildlife Society Bulletin 33: 130–141. DOI: 10.2193/0091-7648(2005)33[130:SEAHSO]2.0.CO;2.
- Li HQ, Zhen L, Chen CG 2009. Winter foraging habitat selection of brown-eared pheasant (Crossoptilon mantchuricum) and the common pheasant (Phasianus colchicus) in Huanglong mountains, Shaanxi province. Acta Ecologica Sinica 29: 335–340. DOI: 10.1016/j.chnaes.2009.09.013
- Mahmoudi S, Sheykhi Ilanloo S, Keyvanloo Shahrestanaki A, Valizadegan N, Yousefi M. 2016. Effect of human-induced forest edges on the understory bird community in Hyrcanian forests in Iran: Implication for conservation and management. Forest Ecology and Management 382: 120–128. DOI: 10.1016/j.foreco.2016.10.011.
- Meriggi A, Pandini W, Cesaris C. 1996. Demography of the pheasant in relation to habitat characteristics in northern Italy. Journal of Wildlife Research. 1: 15–23.
- Molavi‐Arabshahi M, Arpe K, Leroy SAG. 2016. Precipitation and temperature of the southwest Caspian sea region during the last 55 years: Their trends and teleconnections with large‐scale atmospheric phenomena. International Journal of Climatology 36: 2156–2172. DOI: 10.1002/joc.4483.
- Nasiri N, Mohadjer MRM, Etemad V, Sefidi K, Mohammadi L, Gharehaghaji M. 2018. Natural regeneration of oriental beech (Fagus orientalis Lipsky) trees in canopy gaps and under closed canopy in a forest in northern Iran. Journal of Forestry Research 29: 1075–1081. DOI: 10.1007/s11676-017-0490-8.
- Nelli L, Meriggi A, Vidus-Rosin A 2012. Effects of habitat improvement actions (HIAs) and reforestations on pheasants Phasianus colchicus in northern Italy. Wildlife Biology 18:121–130. DOI: 10.2981/11-022
- Ni XJ, Zheng GM, Zhang ZW, Liu N. 2001. Modelling study on the nesting habitat of ring-necked pheasant (Phasianus colchicus). Acta Ecologica Sinica 21: 969–977.
- Nohegar A, Amiri CHB, Afrakhteh R. 2015. Land use analysis on Gilan central district using landscape ecology approach. Geography and Territorial Spatial Arrangement 15: 197–214.
- Pearce J, Boyce M. 2006. Modelling distribution and abundance with presence-only data. Journal of Applied Ecology 43: 405–412. DOI: 10.1111/j.1365-2664.2005.01112.x.
- Pearson RG, Raxworthy CJ, Nakamura M, Peterson AT. 2007. Predicting species distributions from small numbers of occurrence records: A test case using cryptic geckos in Madagascar. Journal of Biogeography 34: 102–117. DOI: 10.1111/j.1365-2699.2006.01594.x.
- Phillips SJ, Anderson RP, Dudík M, Schapire RE, Blair ME. 2017. Opening the black box: An open-source release of Maxent. Ecography 40: 887–893. DOI: 10.1111/ecog.03049.
- Phillips SJ, Anderson RP, Schapire RE. 2006. Maximum entropy modeling of species geographic distributions. Ecological Modeling 190: 231–259. DOI: 10.1016/j.ecolmodel.2005.03.026.
- Priess JA, Mimler M, Klein AM, Schwarze S, Tscharntke T, Steffan-Dewenter I. 2007. Linking deforestation scenarios to pollination services and economic returns in coffee agroforesty systems. Ecological Applications 17: 407–417. DOI: 10.1890/05-1795.
- Ricketts T, Daily G, Ehrlick P, Michencer C 2004. Economic value of tropical forest to coffee production. Proceedings of the National Academy of Sciences USA 101:12579–12661. DOI: 10.1073/pnas.0405147101
- Robertson P. 1997. A Natural History of the Pheasant. Shrewsbury: Swan Hill Press.
- Robertson PA, Woodburn MIA, Neutel W, Bealey CE, 1993. Effects of land use on breeding pheasant density. Journal of Applied Ecology 30:465–477. DOI: 10.2307/2404187
- Robinson SK, Thomson FR, Donovan TM, Whitehead DR, Faaborg J 1995. Regional forest fragmentation and the nesting success of migratory birds. Science 267:1987–1990. DOI: 10.1126/science.267.5206.1987
- Sage R, Tucker K. 1998. Integrated crop management of SRC plantations to maximise crop value, wildlife benefits and other added value opportunities. Oxford, UK: The Game Conservancy Trust report for DTI. DTI publication ETSUB/W2/00400.
- Scharnweber T, Rietschel Manthery M. 2007. Degradation stages of the Hyrcanian forest in southern Azerbaijan. Archiv Fur Naturshutz Und Landshafts Forschung 46: 133–156.
- Scott DA, Moravvej-Hamadani H, Adhami-Mirhosseyni A. 1975. The Birds of Iran. Tehran: Iran Department of Environment (In Farsi, with Latin, English and French names).
- Shirley S 2006. Movement of forest birds across river and clear cut edges of varying riparian buffer strip width. Forest Ecology and Management 223:190–199. DOI: 10.1016/j.foreco.2005.10.071
- Shuai L, Zhou CQ, Wang WK, Wei W, Hu JC. 2007. The habitat and nest-site selection of common pheasants in spring and summer in Nanchong, China. Zoological Research 28: 249–254.
- Smith JNM, Hellmann JJ. 2002. Population persistence in fragmented landscapes. TREE 17: 397–399.
- Suárez-Seoane S, García De La Morena EL, Prieto MBM, Osborne PE, De Juana E 2008. Maximum entropy niche‐based modelling of seasonal changes in little bustard (Tetrax tetrax) distribution. Ecological Modelling 219:17–29. DOI: 10.1016/j.ecolmodel.2008.07.035
- Tucker MA, Böhning-Gaese K, Fagan WF, Fryxell JM, Van Moorter B, Alberts SC et al. 2018. Moving in the anthropocene: Global reductions in terrestrial mammalian movements. Science 359:466–469. DOI: 10.1126/science.aam9712.
- Turner BL, Lambin EF, Reenberg A 2007b. Land change science has emerged as a fundamental component of global environmental change and sustainability research. Proceedings of the National Academy of Sciences USA 104:20672–20677. DOI:10.1073/pnas.0704119104
- Turner WR, Brandon K, Brooks TM, Costanza R, da Fonseca GAB, Portela R. 2007a. Global conservation of biodiversity and ecosystem services. BioScience 57: 868–873. DOI: 10.1641/B571009.
- Wang N, Zhengwang Z, Guangmei Z, McGowan PJ. 2004. Relative density and habitat use of four pheasant species in Xiaoshennongjia mountains, Hubei Province, China. Bird Conservation International 14: 43–54. DOI: 10.1017/S095927090400005X.
- Williams JN, Seo C, Thorne J, Nelson JK, Erwin S, O’Brien JM, Schwartz MW. 2009. Using species distribution models to predict new occurrences for rare plants. Diversity and Distributions 15: 565–576. DOI: 10.1111/j.1472-4642.2009.00567.x.
- Yousefi M, Ahmadi M, Nourani E, Behrooz R, Rajabizadeh M, Geniez P, Kaboli M. 2015. Upward altitudinal shifts in habitat suitability of Mountain Vipers since the last glacial maximum. PLoS ONE 10: e0138087. DOI: 10.1371/journal.pone.0138087.
- Yousefi M, Ahmadi M, Nourani E, Rezaei A, Kafash A, Khani A, Sehhatisabet ME, Adibi MA, Goudarzi F, Kaboli M. 2017a. Habitat suitability and impacts of climate change on the distribution of wintering population of Asian Houbara Bustard (Chlamydotis macqueenii) in Iran. Bird Conservation International 27: 294–304. DOI: 10.1017/S0959270916000381.
- Yousefi M, Kafash A, Malakoutikhah S, Ashoori A, Khani A, Mehdizade Y, Ataei F, Sheykhi Ilanloo S, Rezaei HR, Silva JP. 2017b. Distance to international border shapes the distribution pattern of the growing little Bustard Tetrax tetrax winter population in Northern Iran. Bird Conservation International: 1–10. DOI: 10.1017/S0959270917000181.