Abstract
Research into the pathogenesis of dengue fever has exploded over the last half-century, with issues that were considered simple becoming more complex as additional data are found. This has led to the development of a number of controversies that are being studied across the globe and debated in the literature. In this paper, the following six controversies are analysed and, where possible, resolved: the 1997 World Health Organization (WHO) case definition of dengue haemorrhagic fever (DHF) is not useful; DHF is not significantly associated with secondary dengue infection; DHF results from infection with a ‘virulent’ dengue virus; DHF is owing to abnormal T-cell responses; DHF results from auto-immune responses; and DHF results from direct infection of endothelial cells.
Background
The world is in the midst of a dengue pandemic and more than 1000 papers are added each year to the literature on dengue. Clinicians and scientists attempting to understand the pathogenesis of severe dengue are confronted by six major controversies: (i) the 1997 World Health Organization (WHO) case definition of dengue haemorrhagic fever (DHF) is not useful; (ii) DHF is not significantly associated with secondary dengue infection; (iii) DHF results from infection with a ‘virulent’ dengue virus; (iv) DHF is caused by abnormal T-cell responses; (v) DHF results from auto-immune responses; and (vi) DHF results from direct infection of endothelial cells.
Each will be considered briefly in this article.
(i) The 1997 WHO Case Definition is Not UsefulWhat is the 1997 WHO case definition of DHF/DSS?
Clinical application of the 1997 WHO case definition of DHF/dengue shock syndrome (DSS) () had several problems. Firstly, the tourniquet test and thrombocytopenia have low positive predictive values,Citation1–Citation4 though the greatest problem was recognising and defining clinically significant vascular permeability. Acute dengue vasculopathy generally lasts for less than 48 hours, presenting physicians with an array of rapidly changing pathophysiological conditions.Citation5 The patient must have multiple haematocrit observations, and a definitive diagnosis depends on the timing of these, making nonsense of the complaint that the WHO case definition requires too many ‘repeated clinical tests’.Citation1
Microhaematocrit testing is critical to establishing a diagnosis of hypervolaemia owing to loss of fluid and to designing and managing fluid and colloid resuscitation.Citation6 In most South-east Asian countries, microhaematocrit centrifuges are on treatment wards and used by ward personnel. However, in the Americas, haematocrit determinations require venepuncture and are performed in central laboratories, resulting in serious reporting delays. Therefore, much of the perceived problem in documenting dengue vascular permeability is owing to the organisation of hospital laboratory services.Citation7,Citation8
The 2009 dengue case definition
The 2009 revised WHO case definition () has, however, created serious difficulties for the clinician and research scientist.Citation9 This two-tiered definition consists of initial ‘warning signs’ and a catch-all category, ‘severe dengue’. Note the failure to supply specific quantitative diagnostic criteria and the reliance on individual clinical judgment. How is one to identify ‘clinical fluid accumulation’, ‘increase in haematocrit’ or ‘severe plasma leakage’? What is ‘narrow pulse pressure’ or ‘high haematocrit’?
Figure 1. WHO 2009 suggested dengue case classification and levels of severityCitation9
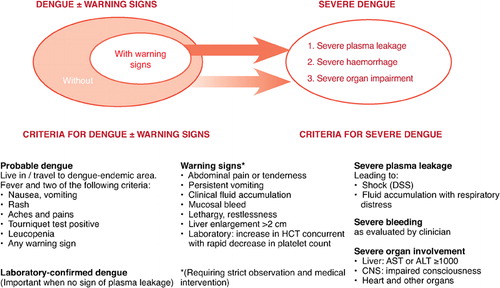
For the clinician, the WHO recommends that all patients with any ‘warning sign’ should be hospitalised.Citation9 Recent experience has demonstrated that compliance with this, particularly in medical communities with little prior experience of DHF, may lead to serious over-hospitalisation. This may then delay triage and recognition of patients requiring life-saving fluid resuscitation, resulting in a high case–fatality rate.Citation10
While the new case definition may be useful for surveillance and reporting, there being no requirement for any laboratory studies, use of the 2009 WHO case definition to define a patient population using the term ‘severe dengue’ will destroy serious research on dengue pathogenesis. ‘Severe dengue’ substitutes a mélange of disease attributes, many end-stage, for the clinically unique and distinct dengue vascular permeability syndrome. Severe organ failure may result from blood loss. It is well documented that dengue fever in adults with peptic ulcer disease may be accompanied by severe focal gastro-intestinal (GI) bleeding, shock and death. Similarly, failure to detect and correct leaky capillaries may result in shock or compensated shock that shunts blood away from the GI tract, resulting in severe bleeding. If uncorrected, these two types of GI bleeding with different causal pathways (and completely different treatment) may result in severe shock, organ failure and encephalopathy. Furthermore, fluid accumulation with respiratory distress (owing to hypervolaemia and pulmonary oedema) is an end-stage outcome possibly resulting from mismanagement of fluid administration, i.e. too much intravenous fluid. To treat end-stage clinical observations as if they derive from a defined clinical syndrome is a serious mistake and has already led to research testing unfeasible pathophysiological hypotheses.Citation11 Therefore, a case definition that discriminates between primary haemorrhage and vascular permeability is necessary for robust pathological research into the spectrum of dengue syndromes.
(ii) The Correlation Between Dengue Vascular Permeability Syndrome (DHF/DSS) and a Secondary Dengue Infection is not Significant
In 1977, Rosen, an early critic of hospital-based observations of an association between a secondary-type antibody response to dengue infection and DHF/DSS, called for field-based studies to clarify the evidence.Citation12 Since then, a number of retrospective sero-epidemiological studies have confirmed that severe dengue disease is associated with secondary dengue infections. This includes data gathered in the ‘favourable environment’ of Cuba, providing unequivocal evidence that individuals circulating dengue 1 antibodies were at risk of DHF during subsequent dengue 2 or dengue 3 infections.Citation13–Citation15
However, there are often problems with data collected in a hospital rather than a study setting, with around 10–30% of hospitalised DHF cases often classified as being caused by primary dengue infection.Citation16 This could be owing to mis-labelling because of the retrospective nature of most of these data where DHF may be diagnosed without evidence of vascular permeability because observers assume that it is indicated by the presence of thrombocytopenia.Citation16 Another explanation is serological misclassification as detection of primary and secondary antibody responses is often based on tests from a single sample of acute-phase serum.Citation16
(iii) DHF/DSS is Caused by Virulent Dengue Viruses
Significant efforts have been directed to finding genetically distinct viruses that cause severe or mild dengue disease. The four dengue virus strains (DENV1–4) vary in terms of pathogenicity and virulence, though the basis for these phenotypic differences is poorly understood. Pathogenicity describes the spectrum of disease syndromes associated with dengue infection. Island epidemics and human volunteer studies provide evidence that different strains within genotypes of dengue viruses vary greatly in intrinsic pathogenicity (i.e. in naive hosts).Citation17,Citation18 The ratio of DHF/DSS to total dengue infections can be measured and is referred to as virulence.
However, the relationship between second infections and dengue vasculopathy is complex. Not all sequential dengue infections result in DHF;Citation19 this can be affected by host factors such as ethnicityCitation20,Citation21 or ageCitation22,Citation23 and viral aspects, including timingCitation24 or sequenceCitation19 of infection, along with heterotypic cross-protection following infection.Citation25,Citation26
In patients at risk of severe disease, the severity, or virulence, of dengue infections is regulated by the antibodies (whether actively or passively acquired). Homologous antibodies can provide complete protection, while heterotypic neutralising antibodies can down-regulate disease. It has also been observed that enhancing antibodies increase the infected cell mass and disease severity. However, it is not understood how this works at the molecular level.Citation27
During the 1997 Santiago de Cuba outbreak caused by DENV2 infection in patients previously exposed to DENV1, the severity of disease increased month by month. The genetic sequences of viruses collected over the course of the epidemic and the serum neutralising antibodies were analysed.Citation13,Citation28 In this way, a single mutation in the non-structural genes of circulating DENV2 viruses might have contributed to viral survival or replication efficiency, thereby enhancing infection in the presence of antibodies.Citation28 This process was described by the researchers as ‘increased viral fitness’, rather than virulence, and might increase the severity of the disease during an outbreak.Citation28
(iv) DHF is caused by Abnormal T-cell Responses
It has been proposed that, in dengue-infected individuals, abnormal and/or accelerated secondary T-cell responses leading to apoptosis contribute to increasing the severity of the immune elimination response.Citation29–Citation33 According to this hypothesis, T-cells from a first infection are inefficient at killing target cells infected with a second virus and would attack infected macrophages, leading to increased cytokine production. These cytokines would affect the vascular endothelium, ultimately causing thrombocytopenia and altered vascular permeability.Citation34
However, in patients with DHF, circulating cytokine levels are similar in infants with primary dengue infections and children of any age with secondary dengue infection.Citation35 DHF/DSS in infants is attributed to antibody-dependent enhancement of dengue infections.Citation36–Citation38 The ability of passively transferred dengue antibodies to enhance dengue viraemia has been demonstrated repeatedly in a monkey model.Citation39,Citation40 Higher levels of dengue plasma viraemia during early disease stages were associated with increased risk of DHF in children with secondary DENV3 infection during a hospital-based prospective study.Citation41
Because primary dengue infections in infants result in authentic DHF, a secondary immune response is not required to produce this syndrome. T-cell researchers need to study infant DHF/DSS to find immunological mechanisms that unify primary- and secondary-infection DHF. Clearly, if T-cell responses contribute directly to vascular permeability, T-cells responding to a first infection must be as efficient as T-cell responses to heterologous infection. However, it might be that T-cells responding to primary infections renders inefficient their response to a secondary infection.Citation33 Speculations that aberrant or abnormal T-cell responses cause DHF/DSS are unwarranted and unnecessary. It has long been noted that individuals with DHF are unusually healthy; surely their immune responses should be normal? Therefore, as two mechanisms cannot be responsible for the same pathology, aberrant or abnormal T-cell responses to dengue infection are not involved.
In the future, when time and effort are invested in studying the pathogenesis and immunology of infant DHF/DSS, it can be expected that a unified explanation will emerge. Meanwhile, it is important to remember that dengue infection in the presence of enhancing antibodies must produce an expanded infected-cell mass. T-cell responses, whether primary or secondary, should be proportional to this antigenic load.Citation42
(v) DHF/DSS Results from an Auto-immune Process
Currently, several mechanisms are proposed to explain auto-immune responses to viral infections, including molecular mimicry.Citation43–Citation45 Similarities have been observed between structural envelope and internal non-structural protein 1 (NS1) of dengue viruses and human proteins.Citation46Citation,47 Furthermore, antibodies to dengue NS1 proteins have been shown to react with plasminogen and integrin,Citation47 plateletsCitation48,Citation49 and endothelial cells.Citation49
However, the hypothesis that this observed structural mimicry is involved in the development of severe disease is inconsistent with the epidemiology and evolution of DHF. For example, in infant DHF, antibodies to envelope or NS1 DENV proteins are unlikely to appear earlier than the fifth day after onset of fever. However, thrombocytopenia in these infants is regularly detected on the second or third day after onset of fever, while vascular permeability occurs around day five. Crucially, thrombocytopenia and vascular permeability ease just as quickly as they begin. It is impossible to understand how NS1 antibodies can produce transient thrombocytopenia and endothelial damage as a result of an antibody response that lasts for many years. If auto-immune responses are mediated by antibodies, why does this not produce chronic vascular permeability and thrombocytopenia?Citation50
(vi) DHF Results from Direct Infection of Endothelial Cells
If the hypothesis that dengue viruses replicate in endothelial cells is correct, it should be possible to observe viral antigens or virions within infected endothelial cells, as has been shown with other infections.Citation51 As yet, however, no unequivocal evidence of DENV infection of endothelial cells in vivo has been shown. Initial findings apparently showing evidence of dengue in endothelial cells was revealed to be dengue antigens on the surface of cells labelled as endothelial cells, and secondary probes showed no evidence of infection within the cells.Citation52
During secondary dengue infection, DENV replication has only been observed within human hepatocytes, monocytes and macrophages.Citation53 Infection peaks after defervescence, with enhanced virus production resulting in a large cell mass. This attracts a massive T-cell response, leading to DSS.Citation54
Conclusions
Dengue infection and the associated spectrum of syndromes are associated with a number of controversies, some of which have been empirically resolved while others require further study. In particular, a clinically and physiologically applicable case classification that will allow robust pathological research into the different levels of disease severity is a major priority.
Box 1 WHO 1997 case definitions for DF, DHF and DSSCitation55
Editorial support was provided by Eleanor Steele of Interlace Global and funded by Sanofi Pasteur.
Related Research Data
References
- Rigau-Perez JG. Severe dengue: the need for new case definitions. Lancet Infect Dis. 2006;6:297–302.
- Bandyopadhyay S, Lum LC, Kroeger A. Classifying dengue: a review of the difficulties in using the WHO case classification for dengue haemorrhagic fever. Trop Med Int Health. 2006;11:1238–55.
- Deen JL, Harris E, Wills B, Balmaseda A, Hammond SN, Rocha C, et al.. The WHO dengue classification and case definitions: time for a reassessment. Lancet. 2006;368:170–3.
- Balmaseda A, Hammond SN, Perez MA, Cuadra R, Solano S, Rocha J, et al.. Short report: assessment of the World Health Organization scheme for classification of dengue severity in Nicaragua. Am J Trop Med Hyg. 2005;73:1059–62.
- Wills BA, Oragui EE, Dung NM, Loan HT, Chau NV, Farrar JJ, et al.. Size and charge characteristics of the protein leak in dengue shock syndrome. J Infect Dis. 2004;190:810–18.
- Halstead SB. The dengue case definition dilemma: a commentary. Pediatr Infect Dis J. 2007;26:291–2.
- Halstead SB. Con: The World Health Organization definition of dengue haemorrhagic fever is inadequate for clinical and epidemiological purposes. In: Halstead SB, ed. Dengue. London: Imperial College Press, 2008; 431–6.
- Rigau-Perez JG. Pro: The World Health Organization definition of dengue haemorrhagic fever is inadequate for clinical and epidemiological purposes. In: Halstead SB, ed. Dengue. London: Imperial College Press, 2008; 427–31.
- World Health Organization. Dengue: Guidelines for Diagnosis, Treatment, Prevention and Control. Geneva, Switzerland: WHO, 2009.
- Srikiatkhachorn A, Rothman AL, Gibbons RV, Sittisombut N, Malasit P, Ennis FA, et al.. Dengue – how best to classify it? Clin Infect Dis. 2011;53:653–7.
- Falconar AK, Martinez F. The NS1 glycoprotein can generate dramatic antibody-enhanced dengue viral replication in normal out-bred mice resulting in lethal multi-organ disease. PLoS ONE. 2011;6:e21024.
- Rosen L. The emperor's new clothes revisited, or reflections on the pathogenesis of dengue hemorrhagic fever. Am J Trop Med Hyg. 1977;26:337–43.
- Guzmán MG, Kouri G, Valdes L, Bravo J, Alvarez M, Vazques S, et al.. Epidemiologic studies on dengue in Santiago de Cuba, 1997. Am J Epidemiol. 2000;159:793–9.
- Guzman MG, Kouri GP, Bravo J, Soler M, Vazquez S, Morier L. Dengue hemorrhagic fever in Cuba, 1981: a retrospective seroepidemiologic study. Am J Trop Med Hyg. 1990;42:179–84.
- Alvarez M, Rodriguez R, Bernardo L, Vasquez S, Morier L, Gonzalez D, et al.. Dengue hemorrhagic fever caused by sequential dengue 1–3 infections at a long interval: Havana epidemic, 2001–2002. Am J Trop Med Hyg. 2006;75:1113–17.
- Anuradha S, Singh NP, Rizvi SN, Agarwal SK, Gur R, Mathur MD. The 1996 outbreak of dengue hemorrhagic fever in Delhi, India. Southeast Asian J Trop Med Public Health. 1998;29:503–6.
- Gubler DJ. Dengue and dengue hemorrhagic fever: its history and resurgence as a global public health problem. In: , Gubler D J, Kuno G, ed Dengue and Dengue Hemorrhagic Fever. New York: CAB International, 1997.
- Gubler DJ, Reed D, Rosen L, Hitchcock JC. Epidemiological, clinical, and virologic observations on dengue in The Kingdom of Tonga. Am J Trop Med Hyg. 1978;27:581–9.
- Watts DM, Porter KR, Putvatana P, Vasquez B, Calampa C, Hayes CG, et al.. Failure of secondary infection with American genotype dengue 2 to cause dengue haemorrhagic fever. Lancet. 1999;354:1431–4.
- Kouri GP, Guzman MG, Bravo JR, Triana C. Dengue haemorrhagic fever/dengue shock syndrome: lessons from the Cuban epidemic, 1981. Bull WHO. 1989;67:375–80.
- Halstead SB, Streit TG, Lafontant JG, Putvatana R, Russell K, Sun W, et al.. Haiti: absence of dengue hemorrhagic fever despite hyperendemic dengue virus transmission. Am J Trop Med Hyg. 2001;65:180–3.
- Guzman MG, Kouri G, Bravo J, Valdes L, Vazquez S, Halstead SB. Effect of age on outcome of secondary dengue 2 infections. Int J Infect Dis. 2002;6:118–24.
- Guzman MG, Kouri G, Valdes L, Bravo J, Vazquez S, Halstead SB. Enhanced severity of secondary dengue 2 infections occurring at an interval of 20 compared with 4 years after dengue 1 infection. PAHO J Epidemiol. 2002;81:223–7.
- Sabin AB. Research on dengue during World War II. Am J Trop Med Hyg. 1952;1:30–50.
- Kochel TJ, Watts DM, Halstead SB, Hayes CG, Espinosa A, Felices V, et al.. Effect of dengue-1 antibodies on American dengue-2 viral infection and dengue haemorrhagic fever. Lancet. 2002;360:310–12.
- Kliks SC, Nisalak A, Brandt WE, Wahl L, Burke DS. Antibody-dependent enhancement of dengue virus growth in human monocytes as a risk factor for dengue hemorrhagic fever. Am J Trop Med Hyg. 1989;40:444–51.
- Halstead SB. Antibodies determine virulence in dengue. Ann NY Acad Sci. 2009;1171 (suppl 1):E48–56.
- Rodriguez-Roche R, Sanchez L, Burgher Y, Rosario D, Alvarez M, Kouri G, et al.. Virus role during intraepidemic increase in dengue disease severity. Vector Borne Zoonotic Dis. 2011;11:675–81.
- Mangada MM, Rothman AL. Altered cytokine responses of dengue-specific CD4+ T cells to heterologous serotypes. J Immunol. 2005;175:2676–83.
- Mongkolsapaya J, Dejnirattisai W, Xu XN. Original antigenic sin and apoptosis in the pathogenesis of dengue hemorrhagic fever. Nat Med. 2003;9:921–7.
- Mongkolsapaya J, Duangchinda T, Dejnirattisai W, Vasanawathana S, Avirutnan P, Jairungsri A, et al.. T cell responses in dengue hemorrhagic fever: are cross-reactive T cells suboptimal? J Immunol. 2006;176:3821–9.
- Midgley CM, Bajwa-Joseph M, Vasanawathana S, Limpitikul W, Wills B, Flanagan A, et al.. An in-depth analysis of original antigenic sin in dengue virus infection. J Virol. 2011;85:410–21.
- Rothman AL. Immunology and immunopathogenesis of dengue disease. Adv Virus Res. 2003;60:397–419.
- Rothman AL, Kanesa-thasan N, West K, Janus J, Saluzzo JF, Ennis FA. Induction of T lymphocyte responses to dengue virus by a candidate tetravalent live attenuated dengue virus vaccine. Vaccine. 2001;19:4694–9.
- Hung NT, Lei HY, Lan NT, Lin YS, Huang KJ, Lien LB, et al.. Dengue hemorrhagic fever in infants: a study of clinical and cytokine profiles. J Infect Dis. 2004;189:221–32.
- Kliks SC, Nimmannitya S, Nisalak A, Burke DS. Evidence that maternal dengue antibodies are important in the development of dengue hemorrhagic fever in infants. Am J Trop Med Hyg. 1988;38:411–19.
- Halstead SB. Neutralization and antibody-dependent enhancement of dengue viruses. In: , Chambers T J, Monath T P, ed, eds. The Flaviviruses: Pathogenesis and Immunity. New York: Elsevier Academic Press, 2003.
- Halstead SB. Immunological parameters of Togavirus disease syndromes. In: Schlesinger RW, ed. Immunological Parameters of Togavirus Disease Syndromes. New York: Academic Press, 1980; 107–73.
- Halstead SB. In vivo enhancement of dengue virus infection in Rhesus monkeys by passively transferred antibody. J Infect Dis. 1979;140:527–33.
- Goncalvez AP, Engle RE, Claire StM, Purcell RH, Lai CJ. Monoclonal antibody-mediated enhancement of dengue virus infection in vitro and in vivo and strategies for prevention. Proc Natl Acad Sci USA. 2007;104:9422–7.
- Libraty DH, Endy TP, Houng HS, Green S, Kalayanarooj S, Suntayakorn S, et al.. Differing influences of virus burden and immune activation on disease severity in secondary dengue-3 virus infections. J Infect Dis. 2002;185:1213–21.
- Halstead SB. Immunological parameters of Togavirus disease syndromes. In: , Schlesinger R W, ed, ed. The Togaviruses, Biology, Structure, Replication. New York: Academic Press, 1980.
- Fujinami RS, von Herrath MG, Christen U. Molecular mimicry, bystander activation, or viral persistence: infections and autoimmune disease. Clin Microbiol Rev. 2006;19:80–94.
- Kim B, Kaistha SD, Rouse BT. Viruses and autoimmunity. Autoimmunity. 2006;39:71–7.
- Rouse BT, Deshpande S. Viruses and autoimmunity: an affair but not a marriage contract. Rev Med Virol. 2002;12:107–13.
- Markoff LJ, Innis BL, Houghten R, Henchal LS. Development of cross-reactive antibodies to plasminogen during the immune response to dengue virus infection. J Infect Dis. 1991;164:294–301.
- Falconar AK. The dengue virus nonstructural-1 protein (NS1) generates antibodies to common epitopes on human blood clotting, integrin/adhesin proteins and binds to human endothelial cells: potential implications in haemorrhagic fever pathogenesis. Arch Virol. 1997;142:897–916.
- Huang KJ, Li SY, Chen SC, Liu HS, Lin YS, Yeh TM, et al.. Manifestation of thrombocytopenia in dengue-2-virus-infected mice. J Gen Virol. 2000;81:2177–82.
- Lin CF, Lei HY, Shiau AL, Liu CC, Liu HS, Yeh TM, et al.. Antibodies from dengue patient sera cross-react with endothelial cells and induce damage. J Med Virol. 2003;69:82–90.
- Halstead SB. Con: Dengue hemorrhagic fever is caused by autoimmune phenomena triggered by a dengue viral infection. In: , Halstead S B, ed, ed. Dengue. London: Imperial College Press, 2008.
- Zaki SR, Greer PW, Coffield LM, Goldsmith CS, Nolte KB, Foucar K, et al.. Hantavirus pulmonary syndrome. Pathogenesis of an emerging infectious disease. Am J Pathol. 1995;146:552–79.
- Jessie K, Fong MY, Devi S, Lam SK, Wong KT. Localization of dengue virus in naturally infected human tissues, by immunohistochemistry and in site hybridization. J Infect Dis. 2004;189:1411–18.
- Balsitis SJ, Coloma J, Castro G, Alava A, Flores D, McKerrow JH, et al.. Tropism of dengue virus in mice and humans defined by viral nonstructural protein 3-specific immunostaining. Am J Trop Med Hyg. 2009;80:416–24.
- Halstead SB, Mahalingam S, Marovich MA, Ubol S, Mosser DM. Intrinsic antibody-dependent enhancement of microbial infection in macrophages: disease regulation by immune complexes. Lancet Infect Dis. 2010;10:712–22.
- World Health Organization. Dengue Haemorrhagic Fever: Diagnosis, Treatment, Prevention and Control, 2nd edn. Geneva, Switzerland: WHO, 1997.