
Abstract
Here we describe the substitution of fluorescently labeled ddUTP for dUTP in the TUNEL assay to allow quantification of generated fluorescence signals by epifluorescence microscopy. The capping of DNase type I 3′OH DNA ends using ddTUNEL was further combined with phosphatase treatment for detection of DNase type II 3′PO4 ends in the same sample using a second round of ddTUNEL. Levels of modified DNA bases in tissues and fixed cultured cells could be interrogated in the ddTUNEL assay with the base modification repair enzyme formamidopyrimidine DNA glycosylase. Using rat mammary gland, from days 1 and 7 of involution, we validate the methodology's ability to label apoptotic nuclei and apoptotic inclusion bodies. In addition, we examined the types of DNA damage and modification that occur in human glioblastoma, U87 cells, following exposure to reactive oxygen stressing agents, chemotherapeutic alkylating agents, and a topoisomerase I inhibitor, irinotecan.
Digging a New TUNEL
The terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) assay is a standard method for detecting DNA fragmentation associated with apoptosis. Labeled dUTP is added by terminal deoxynucleotidyl transferase to the 3′OH ends of DNA fragments that are generated by type I DNases during apoptosis. However, this assay is inherently nonquantitative since the added dUTP itself has a 3′OH end, resulting in the formation of deoxyuridine polymers of indeterminate length at each primary 3′OH site. In this issue, Baskin et al. from the Methodist Hospital (Houston, T X) present a modified TUNEL assay they call ddTUNEL, which uses ddUTP instead of dUTP to stoichiometrically label each DNA3′OH end with a sing le dideoxyuridine base that cannot be further extended, thereby permitting quantitation of the number of 3′OH ends by fluorescence microscopy. This capping of the DNase type I–induced 3′OH ends has the added benefit of enabling additional rounds of ddTUNEL to examine 3′OH ends subsequently generated from other forms of DNA damage in the same sample. For instance, the 3′ phosphate ends of DNA fragments generated by type II DNases associated with caspase independent apoptosis can be detected by treatment of the sample from a first round of ddTUNEL with calf intestinal alkaline phosphatase (CIAP). This creates additional 3′OH ends that can be differentially labeled in a second round of ddTUNEL. Modified bases and apurinic/apyrimidinic (AP) sites in DNA can also be independently assayed by using the bacterially derived enzyme formamidopyrimidine-DNA glycosylase, which excises many modified bases to form an AP site that is then hydrolyzed to a nick containing a 3′ phosphate end that can then be converted to a 3′OH end with CIAP. These new ddTUNEL assays were validated in a study of mammary involution, which involves large-scale apoptosis, and by an examination of DNA damage in U87 cells upon exposure to various DNA-modifying agents.
See “Quantification of DNase type I ends, DNase type II ends, and modified bases using fluorescently labeled ddUTP, terminal deoxynucleotidyl transferase, and formamidopyrimidine-DNA glycosylase” on page 505.
Introduction
Terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) is a common method for detecting DNA fragmentation. Originally described in 1992, terminal deoxynucleotidyl transferase (TdT) catalyzes the addition of labeled dUTP onto 3′OH DNA ends (Citation1). As the generation of 3′OH ends is commonly found in apoptosis, the TUNEL assay has become one of the main methods for detecting apoptotic programmed cell death.
One deficiency with the classical TUNEL assay is the use of dUTP as a substrate, which results in formation of (U)n 3′OH polymers at each DNA 3′OH primary site. Formation of indeterminate length polymers renders the assay intrinsically unquantifiable (Citation2,Citation3). Each addition of dUTP creates a new dU 3′OH end, precluding additional rounds of TUNEL. There are a large number of methodologies that can generate 3′OH DNA ends de novo. For example, 3′OH ends are formed when BrdU incorporated into DNA is irradiated with U V light (Citation4). Here we describe a modified TUNEL assay called ddTUNEL, in which each 3′OH end is capped with a TUNEL-negative ddU 3′H end, thus enabling multiple sequential ddTUNEL assays to be performed on the same sample.
Materials and methods
All reagents were obtained from Sigma-Aldrich (St. Louis, MO, USA) unless otherwise specified.
Preparation of fluorescein isothiocyanate–labeled gelatin phantoms
We have developed a methodology for the production of labeled tissue phantoms that allows the quantification of fluorescence in microscopy (Citation5). Gelatin was dissolved in 1 mL 10 mM potassium phosphate, pH 7.0, to a concentration of 3%, at 45°C. A small amount (≤100 µg) of the solid fluorescein isothiocyanate (FITC) probe was added and incubated at 45°C. The gelatin conjugate was precipitated by the addition of ice-cold ethanol, washed, dried, and resuspended in hot water.
Casting and fixing of standard blocks
The labeled 3% gelatin solution was dissolved in 19% gelatin, at 45°C, to arrive at a final concentration of 15%. A concentration gradient of samples of gelatin conjugate was prepared by diluting the stock in 15% gelatin. The spectrum of the samples was taken to determine the concentration of FITC-gelatin, and aliquots were transferred into silicone molds. The molds were cooled to 4°C, and the casts were fixed in 4% paraformaldehyde (PFA). The plasticized blocks were treated in exactly the same manner as any authentic fixed tissue, being dehydrated, impregnated with wax, sliced, and mounted on slides. They were then dewaxed with xylene, rehydrated, and covered with a coverslip and sealed.
ddTUNEL
A TdT reaction buffer was prepared daily diluting a stock solution 1:5 TUNEL buffer (125 mM Tris-HCl, 1 M sodium cacodylate, 1.25 mg/mL BSA, pH 6.6) and a 25 mM cobalt chloride stock solution, 1:25. The sample was washed twice in this reaction buffer, and then ∼50 µL reaction buffer containing 20 U/mL TdT and 250 nM labeled ddUTP were applied to each of the samples, which were then incubated in a humidified box overnight at room temperature or for 2 h at 37°C.
ddTUNEL: biotinylated ddUTP and fluorescently labeled ddUTP
Biotin-16-ddUTP (Roche Diagnostics, Indianapolis, IN, USA) labels were developed using labeled avidin, which we fluorescently labeled ourselves. PromoFluor-594 and PromoFluor-425 ddUTP were obtained from PromoCell GmbH (Heidelberg, Germany).
CIAP-ddTUNEL
Each sample, having previously undergone ddTUNEL, was washed and incubated with NEBuffer 3 (New England BioLabs, Ipswich, MA, USA) for 30 min and then with a ∼50-µL aliquot of the same buffer containing 100 U/mL calf intestinal alkaline phosphatase (CIAP; Sigma-Aldrich) for ≥2 h; the newly generated 3′PO4→3′OH ends were ddTUNEL-labeled.
CIAP-ddTUNEL positive controls
As levels of 3′PO4 were typically very low in all of the samples we investigated, we prepared positive 3′PO4 controls using authentic DNase type II (porcine spleen). We treated fixed, permeabilized, and washed U87 cells with 10 U/mL DNase type II for 30 min at 37°C in 80 mM sodium acetate buffer, 25 mM MgCl2, pH 4.6. 3′OH ends were labeled using ddTUNEL with FITC-avidin/bitotin-ddUTP, and then half of the samples were incubated with CIAP and the other half in buffer without CIAP. The samples underwent a second round of ddTUNEL with newly generated 3′PO4 →3′OH ends labeled with Texas Red-avidin/biotin-ddUTP.
The Fpg-ddTUNEL assay
Following ddTUNEL and CIAP-ddTUNEL, capping all 3′OH/3′PO4 ends, samples were washed twice in 10 mM HEPES, 10 mM NaCl, 2mM EDTA, and 0.1% BSA. Then, a ∼50-µL aliquot of the same buffer containing 100 U/mL formamidopyrimidine-DNA glycosylase (Fpg; USB, Cleveland, OH, USA) was applied to each of the sections, and then incubated in a humidified box for ≥2 h. Each sample was washed twice in 1× PBS (Thermo Fisher Scientific, Rockford, IL, USA), twice in NEBuffer 3, and then a ∼50-µL aliquot of the same buffer containing 100 U/mL CIAP was applied to each section and incubated for ≥2 h; samples then underwent a third round of ddTUNEL.
Reduction of apurinic/apyrimidinic (AP) sites
Fpg-negative deoxyribitol samples were generated by incubation in 25 mM NaBH4/70% methanol for 30 min.
2,4-Dinitrophenylhydrazine derivatization and labeling of AP sites
Fpg-negative dinitrophenylhydrazone samples were prepared in 15 mM 2,4-dinitro-phenylhydrazine (DNP-H)/2.5 M HCl for 30 min. After incubation with 10% horse sera (Invitrogen, Carlsbad, CA, USA) for 1h and washing, dinitrophenyl (DNP) was imaged using a rabbit anti-DNP primary antibody and donkey anti-goat FITC-labeled secondary antibody (Invitrogen).
H2A.X and CD3-∊ labeling
After incubation with 10% horse sera and washing, the presence of Ser139-phosphorylated H2A.X histone was imaged using an FITC-labeled monoclonal antibody (Biolegend, San Diego, CA, USA). Immune cell marker CD3-∊ (Citation6) was imaged using an FITC-labeled monoclonal antibody (Santa Cruz Biotechnologies, Santa Cruz, CA, USA).
Rat mammary gland
Slides containing 5 µm–thick formalin-fixed paraffin-embedded (FFPE) slices of rat mammary gland on day 1 and day 7 of involution [a model system of apoptosis (Citation7–9)] were purchased from Zyagen (San Diego, CA, USA). Sudan Black was used to remove autofluoresence signals, as suggested by Romijn (Citation10); 0.3% Sudan Black/70% EtOH was applied to the slide for 10 min and washed eight times in 1× PBS.
U87 cells
U87 human glioblastoma were grown in MEM with penicillin/streptomycin and 10% FBS. Following 3 weeks of growth, 5 mL cells were plated into slide chambers (Lab-Tek, Rochester, NY, USA) at 2 × 105 cells/mL and grown for 24h. The media was removed, and 2 mL fresh media [which contained 25 µM temozolomide, 2.5 µM carmustine, 250 µM irinotecan (all from AXXORA, San Diego, CA, USA), 1 mM H2O2, and 300 µM paraquat] or an ethanol vehicle control was added. These concentrations were chosen since they represent the median lethal dose (LD50) of these cells measured over a 24-h incubation period. Twenty-four hours later, cells were fixed with 2% PFA and permeabilized in 0.1% Triton X-100.
Epifluorescence microscopy
The signal was acquired using a Nikon Eclipse TE2000-E fluorescent microscope (Nikon, Melville, NY, USA) equipped with a CoolSnap ES digital camera system (Roper-Scientific, Trenton, NJ, USA) containing a charge-coupled device (CCD)-1300-Y/HS 1392 × 1040 imaging array cooled by a Peltier device. Images were recorded using Nikon NIS-Elements software (Nikon).
Microscopic calculations
The pixel dimensions of our microscope/camera have been calibrated by a representative of the manufacturer. At 100× magnification, each pixel element represented an interrogated area of 0.061 µm × 0.061 µm. At 40× magnification, the interrogated area per pixel is 0.162 µm × 0.162 µm. Given that 1000 L = 1 m3, for each 1 µm of sample depth, the pixel volume at 100× is 3.7 × 10−18 L (1 × 10−6 × 6.1 × 10−8 × 6.1 × 10−8 m3) and at 40× is 2.62 × 10−17L (1 × 10−6 × 1.62 × 10−7 × 1.62 × 10−7m3). Therefore, in a 6-µm phantom slice, there are ∼13.4 molecules/µM at 100× magnification, whereas at 40× magnification, each pixel interrogates ∼95 molecules/µM.
Results and discussion
Measuring 3′OH and 3′PO4 ends using ddTUNEL and CIAP-ddTUNEL assays
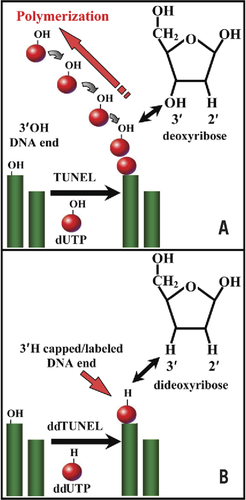
shows the mechanism by which the ddTUNEL assay eliminates polymer formation and stoichiometrically labels each 3′OH end with a ddU 3′H. DNase type II enzymes are associated with caspase-independent calpain/serpin– driven apoptosis (Citation11–13). The 3′PO4 ends that result from DNase type II activity are not substrates for TdT. The sequential usage of two rounds of ddTUNEL, prior to and following incubation with CIAP, allows the determination of both 3′OH and 3′PO4 ends, giving the steady-state levels of both DNase type I and type II DNA damage (Supplementary Figure S1).
(A) The use of 2′ dUTP in the TUNEL assay leads to the unquantifiable polymetric labeling of a single 3′OH present on genomic DNA. Each labeled dUTP added to a 3′OH end acts a substrate for a subsequent dUTP. In contrast, using 2′,3′ ddUTP in the ddTUNEL assay ensures that one and only one labeled ddU is added to each 3′OH DNA end (B). The structures of deoxyribose and dideoxyribose are shown in the appropriate panel, and the arrow indicates the presence of 3′H in ddUTP.
Detection of modified bases and AP sites using Fpg-ddTUNEL assay
DNA is under continuous attack from oxidative/nitrosative stressors, as well as from acylation agents. In living cells, modified DNA bases are repaired by a variety of enzymatic pathways. In Escherichia coli, the enzyme Fpg excises many types of modified DNA bases from double-stranded DNA. Fpg identifies a modified base and, initially, generates an AP site and then hydrolyzes the deoxyribose of each AP site to form a DNA nick flanked by 3′PO4/5′PO4 (Citation14–20). Treatment of a sample with Fpg and CIAP produces a 3′OH at each Fpg-sensitive DNA base and AP site.
While a sample that undergoes ddTUNEL and then CIAP-ddTUNEL leads to capping of all 3′OH/3′PO4 ends, incubation with Fpg/CIAP, followed by ddTUNEL, labels all the newly Fpg-generated 3′OH ends derived from modified bases and AP sites.
AP sites are only hydrolyzed by Fpg if the site contains the aldehyde tautomeric form of deoxyribose (Citation21–24). Reduction with sodium borohydride (NaBH4) (Citation25) or incubation with DNP-H renders AP sites Fpg-negative. AP sites converted into hydrazones by DNP-H can be interrogated independently with a labeled anti-DNP antibody (Supplementary Figure S2).
Quantification of ddTUNEL-labeled fluorescence signals
To quantify signals from different labeled ddUTP probes, we developed a methodology to prepare gelatin-based tissue phantoms containing known amounts of covalently bound fluorophore.
We covalently labeled gelatin with amine-reactive fluorophoric dyes that are cast into molds, fixed using paraformaldehyde, impregnated with wax, sectioned to a known thickness, mounted on slides, dewaxed, rehydrated, and then used to prepare standard curves of fluorescence versus fluorophore concentration.
By combining fluorescently labeled tissue phantoms, we were able to quantify the levels of different types of DNA damage in fixed tissue.
shows typical images obtained from FITC-gelatin phantoms that were then used to prepare a calibration curve (). The 6 µm–thick FITC-gelatin phantom images (; I to VII) were recorded using 40× magnification and an accumulation time of 100 ms. The final image (; VIII) shows background levels of autofluorescence in unconjugated gelatin and was accumulated over 10s.
How FITC-labeled gelatin tissue phantoms are used to calibrate fluorescence signals for the ddTUNEL reaction is shown. (A) The images are of different labeled FITC phantoms. (B) Average fluorescence, n = 3, of these standards and those from two other slides (magnification 100×, accumulation time 100 ms). (C) Fluorescence of the 11.5-µM standard is proportional to exposure time.
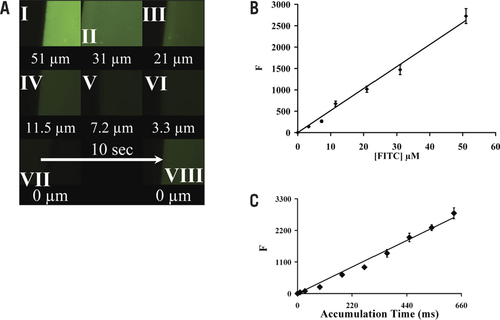
The signal was proportional to concentration, with R2 = 0.99 (). The standard deviation of the FITC signal, at each concentration and using three different slides, averages 7.3% of signal. shows the relationship between signal and accumulation time, using three different 11.5-µM samples measured using 100× magnification. The relationship is linear, with R2 = 0.994 and sd = 5.3% signal mean.
ddTUNEL to study mammary involution
To validate the ddTUNEL assay, we used the assay to characterize the process of mammary involution. Mammary involution is characterized by a massive loss of epithelial cells due to apoptosis, and organ sculpting in this tissue has been widely studied (Citation7–9). The final processing of DNA digestion products, released from epithelial cells in apoptotic bodies, occurs within the macrophages/lymphocytes of the immune system (Citation26–28). , A–D, shows the presence of ddTUNEL-positive nuclei and the formation of apoptotic bodies at different magnifications in breast tissue on day 1; while , E–H, shows day 7 of involution. It can be seen that the apoptotic bodies contain both ddTUNEL- and Fpg-ddTUNEL–positive DNA. Moreover, this DNA is associated with histone γ-H2A.X (, D and H).
Validation of the ddTUNEL (biotin-ddUTP/FITC-avidin) and Fpg-ddTUNEL (PromoFluor-594 ddUTP) assays in rat mammary gland on day 1 and day 7 of involution. (A–C) The presence of ddTUNEL/Fpg-ddTUNEL– positive cells and the formation of apoptotic bodies, at three different magnifications, in breast on day 1 and (E–G) the same labeling of tissue on day 7 of involution. The solid arrows in panel B indicate two cells that are in the early stages of apoptosis, and the cell shown by the dotted arrow is in a later stage of apoptosis. The apoptotic bodies contain ddTUNEL-positive DNA that is associated with histone γ-H2A.X (D and H). Please note that we switched the ddTUNEL probe from green FITC-avidin to red PromoFluor-594 ddUTP to demonstrate that the florescence results from probes and is not due to cytoplasmic lipofuscin pigment. (I and J) CD3-∊–positive immune cells are found near apoptotic cells and apoptotic bodies and contain apoptotic DNA that comes from these cells. (K and L) The presence of ddTUNEL/Fpg-ddTUNEL DNA in apoptotic bodies on days 1 and 7 that is not labeled by DNP-H lacking of AP sites. The ddTUNEL/Fpg-ddTUNEL apoptotic DNA is surrounded by oxidized protein, which is itself surrounded by dying cells. DNA is counterstained by DAPI where indicated.
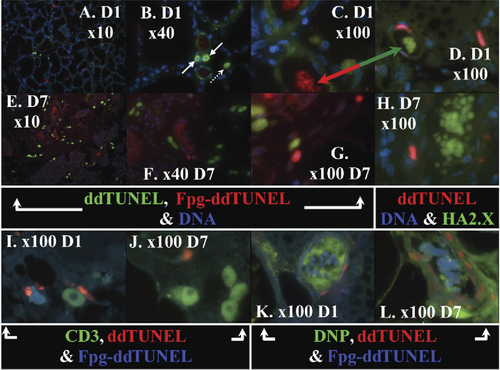
It should be noted that the labeled ddTUNEL probes in , D and H were switched and compared with , A–C and E–G, to demonstrate that the fluorescence of the apoptotic bodies is neither an artifact caused by nonspecific binding of the probes nor due to background fluorescence (Citation28). The red/green arrow in , C and D, shows that the apoptotic bodies have high levels of Fpg-ddTUNEL–positive DNA, and this DNA is co-localized with phosphorylated histone γ-H2A.X.
In , a pair of ddTUNEL-positive cells with a halo of digested oligonucleotides around denuded 4′,6-diamidino-2-phenylindole (DAPI) nuclei can be observed (arrow). In this case, biotin-ddUTP/FITC-avidin was used in the ddTUNEL assay, and the FITC-avidin had a ratio of FITC to protein of 1.03:1. The average signal correlates to an FITC phantom concentration of 24 µM, and we could therefore determine that, in this pair of cells, there was approximately one 3′OH for every 640bp. A cell nearby (dotted arrow) has almost completed its apoptotic death and has a 3′OH for every 105bp. If a cell is digested into the theoretical minimum-sized 180- to 200-bp fragments, there would be a 3′OH for every 95bp, ignoring the contribution of nicks. The average number of cuts/nicks in the cells that do not appear to be apoptotic is in the order of one free 3′OH for every 15,000bp.
The role of immune cells in the cleanup of apoptotic debris is highlighted in , I and J. We labeled the sections with ddTUNEL, Fpg-ddTUNEL, and also with FITC–anti-CD3-∊, a marker of lymphocytes and macrophages. It can be seen that the immune cells are taking up DNA fragments (into the cytosol) and are associated with, but are not part of, the apoptotic bodies.
We measured the relative levels of modified bases and AP sites in the Fpg-ddTUNEL–positive DNA present in the apoptotic bodies. Tissue sections were treated with DNP-H and then labeled using ddTUNEL, Fpg-ddTUNEL, and FITC-labeled anti-DNP antibodies (, K and L). The levels of Fpg-ddTUNEL positive DNA are unchanged, with respect to sections that had not undergone DNP-H treatment, indicating that the DNA in the apoptotic bodies has very few AP sites, but many Fpg-labile bases. We found that there is aldehyde/ketone DNP-H–positive material in the apoptotic bodies, but it is not associated with DNA and is almost certainly oxidized protein.
shows the formation of DNA 3′OH ends and modified bases using ddTUNEL and Fpg-ddTUNEL; along with green DNA (YoPro) (, A–F) or green γ-H2A.X (FITC IgG) (, G–L). Absolute quantification of the levels of the ddTUNEL and Fpg-ddTUNEL, in their respective cellular compartments, is beyond the scope of this publication, but the data from the sample shown in is tabulated in to show the approximate average concentration of the probes, assuming that each section is 6 µm thick, in the cells shown. There are few 3′OH DNA nicks/breaks in the control cells, and they appear green in , but in , the background level of ddTUNEL-positive 3′OH ends is more apparent, as is the presence of γ-H2A.X, which is mostly found in the cytosol.
Use of biotinylated ddUTP in ddTUNEL (fluram-avidin) and Fpg-ddTUNEL (Texas Red–avidin) assays shows DNA damage caused to U87 cells following incubation with ROS-inducing and chemotherapeutic reagents. (A–F) DNA is stained with green YoPro, and (G–L) green γ-H2A.X is shown. The stressors all increase the levels of DNA damage (shown in and discussed in the “Results and discussion” section).
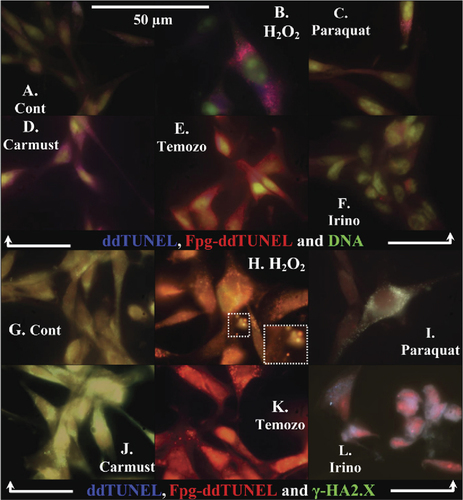
Table 1. Levels of ddTUNEL and Fpg-ddTUNEL in U87 cells following treatment.
Exposure of U87 cells to various modifying agents
H 2 O 2 treatment. Hydrogen peroxide–treated cells (, B and H) show the formation of blebs that are positively stained for both γ-H2A.X and ddTUNEL. ddTUNEL-positive DNA is found at very high levels in the cytosol, mostly within small vesicles, and 3′OH levels are low within the nucleus.
The red Fpg-ddTUNEL is also unevenly distributed; DNA that contains high levels of oxidized bases and AP sites appear to be exported from the nucleus and then concentrated up in the apoptotic vesicles. There appears to be a high degree of heterogeneity in the composition of these apoptotic vesicles, with respect to the levels of γ-H2A.X, ddTUNEL, and Fpg-ddTUNEL–positive DNA. The insert in is an enlargement of the indicated portion of these vesicles and conveys the degree of heterogeneity.
Paraquat treatment. Paraquat, which generates both superoxide and H2O2, has a completely different death signature to H2O2, with no evidence of blebbing. ddTUNEL correlates with γ-H2A.X and is mostly cytosolic and found in the small vesicles (, C and I). It again appears that Fpg-ddTUNEL–positive oxidized DNA is exported out of the nucleus and concentrated into vesicles. This oxidized ddTUNEL-positive DNA is clearly associated with γ-H2A.X, as shown by the presence of the bright white spots ().
Carmustine and temozolomide treatment. Many chemotherapeutic agents, including temozolomide and carmustine, derive their toxicity from the acylation of the N7 position of guanine. The ability of Fpg to directly remove N7 methyl and ethyl guanine lesions is poor. However, both these modified residues can undergo slow ring opening to form formamidopyrimidines, which act as substrates for Fpg. In addition, the formation of bulky adducts in the N7 position, which are generally good Fpg substrates, occurs in vivo upon exposure of mammalian cells to carmustine (Citation29). Bodell and coworkers (Citation29) demonstrated that in mammalian cells, N7-(2-hydroxyethyl)guanine, N7-(2-chloroethyl) guanine, and eight other species were the major adducts following carmustine treatment. In addition, in mammalian cells, N7 methyl and ethyl guanine lesions are processed by the enzyme N-methylpurine DNA glycosylase (MPG), which removes these residues to generate AP sites. In contrast, the lesions 3-methyladenine and O6-methylguanine are primarily species responsible for temozolomide toxicity, and these lesions are also removed by MPG, generating AP sites. Therefore, treatment of cells with chemotherapeutic agents, such as carmustine and temozolomide, generates a variety of Fpg-positive species, which may be measured using Fpg-ddTUNEL.
Treating cells with carmustine (, D and J) induced 6-fold increases in ddTUNEL and Fpg-ddTUNEL. In carmustine-treated cells, ddTUNEL ends are found in a 2:1 ratio in the cytosol compared with the nucleus. The levels of γ-H2A.X, mostly cytoplasmic, are elevated more than 30-fold with respect to the controls.
Temozolomide-treated cells (, E and K) gave the highest Fpg-ddTUNEL–positive result, and it is obvious that this methylated DNA is exported into and concentrated in cytosolic vesicles; these vesicles are also γ-H2A.X– and ddTUNEL-rich. In comparing the ethylation agent, carmustine, to the methylation agent, temozolomide, the striking difference is the 10-fold difference in cytosolic γ-H2A.X levels in carmustine-treated cells and that these histones are not packaged into inclusion bodies. We can only conclude that methylated DNA is treated in a completely different fashion than ethylated/acylated DNA.
Irinotecan treatment. The final pairing (, F and L) shows the effect of the topoisomerase I inhibitor, irinotecan, on cell death. ddTUNEL-rich DNA is intimately associated with both γ-H2A.X and Fpg-ddTUNEL, but strikingly, the apoptotic vesicles are heterogeneous, containing either ddTUNEL with γ-H2 A. X or ddTUNEL with Fpg-ddTUNEL.
We differentiate between DNA base modification and the presence of AP sites using NaBH4 reduction. We expect that H2O2 would increase the levels of oxidized DNA and that carmustine would increase ethylation. We prepared slides that had irinotecan-, H2O2-, and carmustine-treated cells and incubated half of a slide in methanolic NaBH4 to reduce all the AP sites and then ran Fpg-ddTUNEL,
In irinotecan- and carmustine-treated cells, the levels of Fpg-ddTUNEL–sensitive generated 3′OH ends were insensitive to NaBH4 treatment. In striking contrast, in H2O2-treated cells, the Fpg-ddTUNEL signal was abolished by NaBH4 treatment (). Irinotecan increases DNA breaks and does not directly modify DNA bases, carmustine ethylates DNA bases, and H2O2 is a DNA base oxidant.
We found low levels of DNase type II 3′PO4 ends in the both rat mammary tissue and also in the various U87 cell samples. To demonstrate that CIAP-ddTUNEL was able to measure the levels of DNase type II 3′PO4 ends, positive controls were generated by incubating U87 cells with DNase type II. DNase type II–treated cells underwent a first round of biotin-ddUTP-ddTUNEL and were labeled with avidin-FITC. Half the samples were then incubated with CIAP, half were in buffer with the enzyme omitted, and then all samples underwent a second round of ddTUNEL using biotin-ddUTP/avidin–Texas Red. Supplementary Figure S3 shows the visualization of this type of labeling. There is very little (<5%) labeling in the second round of ddTUNEL in the absence of CIAP, demonstrating that it is the presence of CIAP, with its 3′PO4→3′OH activity, that allows labeling in the second round of ddTUNEL.
We have modified the traditional TUNEL assay (Citation1) to allow the absolute levels of TdT-accessible 3′OH to be measured. We have shown that the substitution of ddUTP for dUTP allows the levels of both 3′OH and 3′PO4 DNA ends to be measured in the same sample, using ddTUNEL and CIAP-ddTUNEL, respectively. We have also demonstrated that E. coli enzyme Fpg can be used to excise oxidized/acylated DNA bases and AP sites in vitro using U87 cells in combination with oxidative and acetylating agents.
Fpg-ddTUNEL is very useful for converting AP sites and modified bases into 3′OH-labeled ends. In addition to 8-oxo-7,8-dihydroguanine, a wide range of modified pyrimidines are recognized and excised by Fpg (Citation30). In addition, some modified purines are also substrates for this enzyme (Citation31,Citation32).
The broad substrate specificity of Fpg can also be discerned by the protection it affords mammalian cells that have been transformed with E. coli Fpg. Fpg-transfected cells are much less sensitive to DNA acylating agents [ThioTEPA (Citation14,Citation33), bis(2-chloroethyl)-N-nitrosourea (Citation20), and carmustine (Citation19)], DNA oxidants [potassium bromate, H2O2, and γ-rays (Citation34)], and hyperoxia (Citation18).
We have shown that it is possible to derivativize AP sites with dinitrophenyl hydrazine, which allows one to interrogate the localization of ROS damage within a cell using an anti-DNP antibody. Derivatization of AP sites by fluorophores linked to an aminooxy group (Citation24,Citation35) or using a commercially available biotinylated hydroxylamine (Aldehyde Reactive Probe; Invitrogen, Carlsbad, CA, USA) are neutral pH reactions and may be less harsh alternatives to DNP-H/antibody labeling.
Treatment of DNA with methanolic NaBH4 reduces AP sites, and then Fpg-ddTUNEL allows the discrimination between the levels of modified bases and AP sites. We find that AP sites are the principle product of oxidative stressors, such as H2O2 and paraquat ( and unpublished data). Fenton chemistry is a major cause of oxidative cellular damage (Citation36), and it has previously been postulated that DNA damage caused by Fenton chemistry results in much higher levels of AP sites than oxidized bases, such as oxyguanine (Citation37).
The discrimination between acylated/oxidized bases and AP sites using NaBH reduction. (A–C) ddTUNEL (biotin-ddUTP/FITC-avidin) and Fpg-ddTUNEL (biotin-ddUTP/Texas Red–avidin) sensitive damage to DNA in irinotecan-treated cells, and in cells treated with either H2O2 (which will oxidize DNA) or carmustine (a DNA ethylating agent). (D–F) ddTUNEL and Fpg-ddTUNEL on slides treated with methanolic NaBH4. Only in the H2O2-treated cells is the level of Fpg-ddTUNEL–positive DNA damage altered, which indicates that H2O2 treatment generates a high fraction of AP sites.
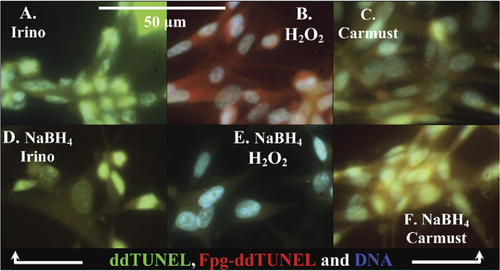
In breast tissue, the Fpg-ddTUNEL– positive DNA concentrated in apoptotic bodies had few AP sites, since Fpg-ddTUNEL was insensitive to either DNP-H or NaBH4; this would suggest that the DNA in the apoptotic bodies is highly methylated.
In vitro, both methyl and ethyl guanine can be converted into formamidopyrimidines, by incubation at high pH (Citation38), and the Fpg-ddTUNEL assay could be modified to include such an alkyl-hydrolysis step. However, similar to Speit and coworkers (Citation17), we find that the products of DNA methylation/ethylation generated within mammalian cells in vivo are Fpg-labile without prealkalinization. Moreover, they found that the changes in pH imposed by an alkalinization/neutralization protocol resulted in a considerable loss in sensitivity in their combined Fpg-Comet assay.
The three techniques—ddTUNEL, CIAP-ddTUNEL, and Fpg-ddTUNEL, introduced and validated herein—can be combined with the use of tissue phantoms for signal calibration, allowing the absolute levels of three different types of DNA damage to be quantified within individual cells for the first time.
Competing interests
The authors declare no competing interests.
Supplementary Material For: Quantification of DNase type I ends, DNase type II ends, and modified bases using fluorescently labeled ddUTP, terminal deoxynucleotidyl transferase, and formamidopyrimidine-DNA glycosylase
Download PDF (1.6 MB)Acknowledgments
We thank our research assistant, Sophie Lopez, for all her assistance in cell growth, sample preparation, and many other duties. We also thank Kalika M. Landua of Nikon Instruments for her aid in the calibration of our microscope and camera. Funding for this research was provided by The Henry J.N. Taub Fund for Neurological Research, The Pauline Sterne Wolff Memorial Foundation, Golfers Against Cancer, and the Methodist Hospital Foundation.
Supplementary data
To view the supplementary data that accompany this paper please visit the journal website at: www.tandfonline.com/doi/suppl/10.2144/000113439
Additional information
Funding
References
- Gavrieli, Y., Y.Sherman, and S.A.Ben-Sasson. 1992. Identification of programmed cell death in situ via specific labeling of nuclear DNA fragmentation. J. Cell Biol.119:493–501.
- Kelly, K.J., R.M.Sandoval, K.W.Dunn, B.A.Molitoris, and P.C.Dagher. 2003. A novel method to determine specificity and sensitivity of the TUNEL reaction in the quantitation of apoptosis. Am. J. Physiol. Cell Physiol.284:C1309–C1318.
- Kanoh, M., G.Takemura, J.Misao, Y.Hayakawa, T.Aoyama, K.Nishigaki, T.Noda, T.Fujiwara, et al.. 1999. Significance of myocytes with positive DNA in situ nick end-labeling (TUNEL) in hearts with dilated cardiomyopathy: not apoptosis but DNA repair. Circulation99:2757–2764.
- Darzynkiewicz, Z., X.Huang, and M.Okafuji. 2006. Detection of DNA strand breaks by flow and laser scanning cytometry in studies of apoptosis and cell proliferation (DNA replication). Methods Mol. Biol.314:81–93.
- Sharpe, M.A., M.A.Widmayer, and D.S.Baskin. 2010. Quantification and calibration of images in fluorescence microscopy. Anal. Biochem. May 28. [Epub ahead of print].
- Wingender, G., B.Schumak, A.Schurich, J.E.Gessner, E.Endl, A.Limmer, and P.A.Knolle. 2006. Rapid and preferential distribution of blood-borne CD3-∊ to the liver is followed by local stimulation of T cells and natural killer T cells. Immunology117:117–126.
- Colitti, M. and M.Farinacci. 2009. Cell turnover and gene activities in sheep mammary glands prior to lambing to involution. Tissue Cell41:326–333.
- Bagheri-Yarmand, R., R.K.Vadlamudi, and R.Kumar. 2003. Activating transcription factor 4 overexpression inhibits proliferation and differentiation of mammary epithelium resulting in impaired lactation and accelerated involution. J. Biol. Chem.278:17421–17429.
- Lacher, M.D., A.Siegenthaler, R.Jager, X.Yan, S.Hett, L.Xuan, S.Saurer, R.R.Lareu, et al.. 2003. Role of DDC-4/sFRP-4, a secreted frizzled-related protein, at the onset of apoptosis in mammary involution. Cell Death Differ.10:528–538.
- Romijn, H.J., J.F.M.Van Uum, I.Breedijk, J.Emmering, I.Radu, and C.W.Pool. 1999. Double immunolabeling of neuropeptides in the human hypothalamus as analyzed by confocal laser scanning fluorescence microscopy. J. Histochem. Cytochem.47:229–235.
- Counis, M.F. and A.Torriglia. 2006. Acid DNases and their interest among apoptotic endonucleases. Biochimie88:1851–1858.
- Reme, C.E., C.Grimm, F.Hafezi, A.Marti, and A.Wenzel. 1998. Apoptotic cell death in retinal degenerations. Prog. Retin. Eye Res.17:443–464.
- Torriglia, A. and C.Lepretre. 2009. LEI/LDNase II: interplay between caspase-dependent and independent pathways. Front. Biosci.14:4836–4847.
- Gill, R.D., C.Cussac, R.L.Souhami, and F.Laval. 1996. Increased resistance to N,N′,N′′-triethylenethiophosphoramide (thiotepa) in cells expressing the Escherichia coli formamidopyrimidine-DNA glycosylase. Cancer Res.56:3721–3724.
- O'Connor, T.R. and J.Laval. 1989. Physical association of the 2,6-diamino-4-hydroxy-5N-formamidopyrimidine-DNA glycosylase of Escherichia coli and an activity nicking DNA at apurinic/apyrimidinic sites. Proc. Natl. Acad. Sci. USA86:5222–5226.
- Ropolo, M., A.Geroldi, P.Degan, V.Andreotti, S.Zupo, A.Poggi, A.Reed, M.R.Kelley, and G.Frosina. 2006. Accelerated repair and reduced mutagenicity of oxidative DNA damage in human bladder cells expressing the E. coli FPG protein. Int. J. Cancer118:1628–1634.
- Speit, G., P.Schitz, I.Bonzheim, K.Trenz, and H.Hoffmann. 2004. Sensitivity of the FPG protein towards alkylation damage in the comet assay. Toxicol. Lett.146:151–158.
- Wu, M., Y.H.He, M.Kobune, Y.Xu, M.R.Kelley, and W.J.MartinII. 2002. Protection of human lung cells against hyperoxia using the DNA base excision repair genes hOgg1 and Fpg. Am. J. Respir. Crit. Care Med.166:192–199.
- Ying-Hui, H.E., Y.I.Xu, M.Kobune, W.U.Min, M.R.Kelley, and W.J.MartinII. 2002. Escherichia coli FPG and human OGG1 reduce DNA damage and cytotoxicity by BCNU in human lung cells. Am. J. Physiol. Lung Cell. Mol. Physiol.282:L50–L55.
- Xu, Y., W.K.Hansen, T.A.Rosenquist, D.A.Williams, M.Limp-Foster, and M.R.Kelley. 2001. Protection of mammalian cells against chemotherapeutic agents thiotepa, 1,3-N,N-bis(2-chloroethyl)-N-nitrosourea, and mafosfamide using the DNA base excision repair genes Fpg and Ogg1: implications for protective gene therapy applications. J. Pharmacol. Exp. Ther.296:825–831.
- Nakamura, J., V.E.Walker, P.B.Upton, S.Y.Chiang, Y.W.Kow, and J.A.Swenberg. 1998. Highly sensitive apurinic/apyrimidinic site assay can detect spontaneous and chemically induced depurination under physiological conditions. Cancer Res.58:222–225.
- Dahlmann, H.A., V.G.Vaidyanathan, and S.J.Sturla. 2009. Investigating the biochemical impact of DNA damage with structure-based probes: a basic sites, photodimers, alkylation adducts, and oxidative lesions. Biochemistry48:9347–9359.
- Chastain, 2nd, P.D., J.Nakamura, J.Swenberg, and D.Kaufman. 2006. Nonrandom AP site distribution in highly proliferative cells. FASEB J.20:2612–2614.
- Boturyn, D., J.F.Constant, E.Defrancq, J.Lhomme, A.Barbin, and C.P.Wild. 1999. A simple and sensitive method for in vitro quantitation of abasic sites in DNA. Chem. Res. Toxicol.12:476–482.
- Smith, M.A., L.M.Sayre, V.E.Anderson, P.L.R.Harris, M.F.Beal, N.Kowall, and G.Perry. 1998. Cytochemical demonstration of oxidative damage in A lzheimer disease by immunochemical enhancement of the carbonyl reaction with 2,4-dinitrophenylhydrazine. J. Histochem. Cytochem.46:731–735.
- Tatarczuch, L., C.Philip, and C.S.Lee. 1997. Involution of the sheep mammary gland. J. Anat.190:405–416.
- Kralj, M. and N.Pipan. 1995. Degradation processes in different functional states of the mouse mammary gland. Period. Biol.97:201–206.
- Walker, N.I., R.E.Bennett, and J.F.R.Kerr. 1989. Cell death by apoptosis during involution of the lactating breast in mice and rats. Am. J. Anat.185:19–32.
- Bodell, W.J., A.P.Bodell, and D.D.Giannini. 2007. Levels and distribution of BCNU in GBM tumors following intratumoral injection of DTI-015 (BCNU-ethanol). Neuro. Oncol.9:12–19.
- Tchou, J., V.Bodepudi, S.Shibutani, I.Antoshechkin, J.Miller, A.P.Grollman, and F.Johnson. 1994. Substrate specificity of Fpg protein. Recognition and cleavage of oxidatively damaged DNA. J. Biol. Chem.269:15318–15324.
- Hatahet, Z., Y.W.Kow, A.A.Purmal, R.P.Cunningham, and S.S.Wallace. 1994. New substrates for old enzymes. 5-Hydroxy-2′-deoxycytidine and 5-hydroxy-2′-deoxyuridine are substrates for Escherichia coli endonuclease III and formamidopyrimidine DNA N-glycosylase, while 5-hydroxy-2′-deoxyuridine is a substrate for uracil DNA N-glycosylase. J. Biol. Chem.269:18814–18820.
- D'Ham, C., A.Romieu, M.Jaquinod, D.Gasparutto, and J.Cadet. 1999. Excision of 5,6-dihydroxy-5,6-dihydrothymine, 5,6-dihydrothymine, and 5-hydroxycytosine from defined sequence oligonucleotides by Escherichia coli endonuclease III and Fpg proteins. Biochemistry38:3335–3344.
- Kobune, M., Y.Xu, C.Baum, M.R.Kelley, and D.A.Williams. 2001. Retrovirus-mediated expression of the base excision repair proteins, formamidopyrimidine DNA glycosylase or human oxoguanine DNA glycosylase, protects hematopoietic cells from N,N′,N′′-triethylenethiophosphoramide (thioTEPA)-induced toxicity in vitro and in vivo. Cancer Res.61:5116–5125.
- Frosina, G. 2006. Prophylaxis of oxidative DNA damage by formamidopyrimidine-DNA glycosylase. Int. J. Cancer119:1–7.
- Boturyn, D., A.Boudali, J.-F.Constant, E.Defrancq, and J.Lhomme. 1997. Synthesis of fluorescent probes for the detection of abasic sites in DNA. Tetrahedron53:5485–5492.
- Sharpe, M.A., S.J.Robb, and J.B.Clark. 2003. Nitric oxide and Fenton/Haber-Weiss chemistry: nitric oxide is a potent antioxidant at physiological concentrations. J. Neurochem.87:386–394.
- Henle, E.S., Y.Luo, W.Gassmann, and S.Linn. 1996. Oxidative damage to DNA constituents by iron-mediated Fenton reactions. J. Biol. Chem.271:21167–21176.
- Tudek, B. 2003. Imidazole ring-opened DNA purines and their biological significance. J. Biochem. Mol. Biol.36:12–19.

