Abstract
Background
Human schistosomiasis is a freshwater snail-transmitted disease caused by parasitic flatworms of the Schistosoma genus. Schistosoma haematobium, Schistosoma mansoni, and Schistosoma japonicum are the three major species infecting humans. These parasites undergo a complex developmental life cycle, in which they encounter a plethora of environmental signals. The presence of genes encoding the universal stress protein (USP) domain in the genomes of Schistosoma spp. suggests these flatworms are equipped to respond to unfavorable conditions. Though data on gene expression is available for USP genes, their biochemical and environmental regulation are incompletely understood. The identification of additional regulatory molecules for Schistosoma. USPs, which may be present in the human, snail, or water environments, could also be useful for schistosomiasis interventions.
Methods
We developed a protocol that includes a visual analytics stage to facilitate integration, visualization, and decision making, from the results of sequence analyses and data collection on a set of 13 USPs from S. mansoni and S. japonicum.
Results
Multiple sequence alignment identified conserved sites that could be key residues regulating the function of USPs of the Schistosoma spp. Based on the consistency and completeness of sequence annotation, we prioritized for further research the gene for a 184-amino-acid-long USP that is present in the genomes of the three human-infecting Schistosoma spp. Calcium, zinc, and magnesium ions were predicted to interact with the protein product of the gene.
Conclusion
Given that the initial effects of praziquantel on schistosomes include the influx of calcium ions, additional investigations are required to (1) functionally characterize the interactions of calcium ions with the amino acid residues of Schistosoma USPs; and (2) determine the transcriptional response of Schistosoma. USP genes to praziquantel. The data sets produced, and the visual analytics views that were developed, can be easily reused to develop new hypotheses.
Introduction
Human schistosomiasis is a freshwater snail-transmitted disease caused by parasitic flatworms of the Schistosoma genus.Citation1Schistosoma haematobium, Schistosoma mansoni, and Schistosoma japonicum are the three major species infecting humans. Schistosomiasis has been designated as one of the “neglected tropical diseases” of poverty and is the second most significant tropical disease, after malaria, in public health significance.Citation2 The large majority of human schistosomiasis and most of the severest disease states are now concentrated in the relatively resource-poor countries of sub- Saharan Africa, contributing to approximately 280,000 deaths per annum.Citation3 Schistosomiasis is also among the severest parasitic diseases targeted, in terms of morbidity and mortality, and has been highlighted for control by the World Health Organization (WHO), with the urinary form highly associated with increased risks for bladder cancer.Citation2,Citation4 The drug of choice for treatment of schistosomiasis is praziquantel (PZQ), but there is great concern regarding effective treatment in affected communities, due to the potential for parasite resistance to PZQ.Citation4–Citation6
The genomes of S. haematobium, S. mansoni, and S. japonicum encode proteins with the universal stress protein (USP) domain (Pfam Identifier: PF00582).Citation7–Citation9 The USPs are known to function during unfavorable environmental conditions, including the life cycle developmental stages in Schistosoma spp.Citation10–Citation12 Though data on gene expression is available for genes encoding USPs (USP genes), their biochemical and environmental regulation are incompletely understood.Citation10–Citation19 The identification of additional regulatory molecules for Schistosoma USPs, which may be present in the human, snail, or water environments, could also be useful for schistosomiasis interventions. Our hypothesis is that the USPs of Schistosoma spp. have shared protein sequence features that can help us infer their biochemical and environmental regulation. Further, in the context of an infectious disease caused by geographically dispersed species of the same genus, proteins that have shared features could be targets for intervention.
The USPs are found in a diverse group of organisms, including archaea, bacteria, yeast, fungi, and plants, and encompass a conserved group of proteins whose expressions are triggered by a variety of environmental insults, including toxic chemicals, drought, and extreme temperature.Citation20 Genes encoding USPs have not been identified in the human genome, thus making them attractive as drug targets.Citation21 In a previous report,Citation12 we analyzed the developmental expression of eight USP genes predicted from the S. mansoni genome. In the case of S. japonicum, multiple research investigations have detected developmental stage expression of USP genes.Citation10,Citation11,Citation16
The Schistosoma spp. undergo a complex developmental life cycle that includes multiple morphological stages and transition between hosts. The life cycle developmental forms of Schistosoma spp. include egg, miracidium, sporocyst, cercaria, schistosomulum, and adult (male and female). These stages must survive diverse stress conditions. The eggs, miracidia, and cercariae are found outside the human and snail hosts and are thus exposed to the stress conditions associated with the freshwater environment of the snail vectors. The cercariae and sporocysts are found in the snail host and must respond to toxic substances, causing oxidative and nitrosative stresses in the snail hemocytes.Citation22,Citation23 In the human host, to develop to the adult form, the schistosomula migrate through multiple organs, including lungs, heart, and liver. These organ systems have defense mechanisms, including production of nitric oxide and hydrogen peroxide, designed to kill the parasite stages.Citation24,Citation25 In summary, all the developmental stages of the Schistosoma spp. are exposed to various stresses. A common aspect of these environment-inducing stresses is that they result in proteins with nonnative conformations.Citation26 Inducible stress tolerance has increasingly been understood to result from numerous molecular mechanisms, such as heat shock proteins (Hsps) and USPs.Citation12,Citation27–Citation31
Adenosine triphosphate (ATP) binding is a biochemical mechanism that regulates the function of USPs through phosphorylation.Citation32 Members of the USP family can be categorized into two groups based on the presence or absence of the ATP-binding motif G-2x-G-9x-G(S/T) in their amino acid sequence.Citation20,Citation33 USPs can be phosphorylated on serine and threonine residues by phosphate donors ATP and guanosine triphosphate (GTP), in the absence of other proteins, coupled with an upregulation response to stressors,Citation34 This observation indicates that in addition to environmental-stressor-mediated regulation, other cellular factors could modulate the activity of USPs by controlling their phosphorylated state.Citation35 Given the broad range of resistance functions conferred by the USPs, it is not surprising that they are encoded in the genomes of a variety of both pathogens and nonpathogens.Citation20,Citation36–Citation38
We have determined protein sequence length, protein domain length, ligand-binding sites, biologically relevant chemical ligands, enzymatic regulation, developmental regulation, and subcellular localization for 13 Schistosoma USP sequences (five S. mansoni and eight S. japonicum sequences). Multiple sequence alignment and phylogenetic analysis provided the evolutionary groupings to allow inferences on biochemical and environmental regulation of the proteins. The results from the analyses were integrated and visualized with visual analytics software. Multiple sequence alignment identified conserved sites of aspartate (Asp), glycine (Gly), histidine (His), leucine (Leu), and proline (Pro) residues in all the sequences. These residues could be key residues regulating the function of the USPs of Schistosoma spp. We prioritized a group of two 184-amino-acid-long USP sequences (Q86DW2 [S. japonicum] and G4LZI3 [S. mansoni]) because they had identical values for multiple annotation features. Data visualization revealed the two proteins have identical values for subcellular localization, ligand-binding sites, chemical ligands, and enzymatic regulation. Specifically, calcium, zinc, and magnesium ions were predicted to interact with the two proteins. Given that the initial effects of PZQ on schistosomes include the influx of calcium ions,Citation13,Citation39–Citation41 additional investigations are required to (1) functionally characterize the interactions of calcium ions with the amino acid residues of S. USPs; and to (2) determine the transcriptional response of Schistosoma USP genes to PZQ.
Methods
Overview of bioinformatics and visual analytics methods
A variety of limitations, including costs, preclude the functional characterization of all predicted proteins from a genome sequencing project. The selection of proteins for further research is a decision-making process by a researcher or research team. Thus, we developed a protocol that integrates the visual analytics stages, to facilitate the interaction with the results from sequence analysis and data collection, on a set of USPs from S. mansoni and S. japonicum. Visual analytics is an iterative process conducted via visual interfaces that involves collecting information, data preprocessing, knowledge representation, interaction, and decision making.Citation42–Citation46
The overview of the bioinformatics and visual analytics methods is summarized in . The protocol consists of five stages that start with the protein sequences to be investigated (Stage 1). Two sets of bioinformatics analyses are performed (Stage 2 and Stage 3). Stage 2 consists of analyses done on each sequence, while, in Stage 3, all the sequences are used for multiple sequence alignment and to construct phylogenetic trees. The bioinformatics sequence analyses in Stage 2 determine (1) the protein sequence length; (2) the protein domain length; (3) the ligand-binding sites; (4) chemical ligand binding; (5) kinase binding; and (6) subcellular localization. These analyses can be particularly useful for prioritizing sequences for research on the biochemical and environmental regulation of proteins. Information on the developmental expression of gene transcripts can assist in deciding the choice of life cycle parasite form to investigate. The data on the developmental expression of gene transcripts were obtained from publications. Multiple sequence alignment and phylogenetic trees provided the statistically and evolutionary support for groupings of the sequences.
Figure 1 Overview of a set of bioinformatics and visual analytics methods used to prioritize protein sequences for further research.
The prioritization process was done in Stage 4, with the criterion determined by the researchers. In this report, the criterion was to identify pairs of protein sequences (one from S. mansoni and another from S. japonicum) that share identical annotations, from the following analyses: protein sequence length, ligand-binding sites (amino acid type and amino acid position), and chemical ligands that are predicted to bind. Stage 5 was the product of the prioritization process. In this report, we expect (i) orthologous pairs of protein sequences and (ii) a visualization that provides an integrated view of the shared annotations. We used a visual analytics software packageCitation45 (Tableau 7.0, Tableau Software Inc, Seattle, WA, USA) to perform several visual analytics tasks, including interaction, computing, analysis, integration, and visualization.
Retrieval of protein sequences
Proteins annotated with the USP domain (PF00582) from the S. mansoni and S. japonicum genomes were identified in Universal Protein Resource (UniProt release 2011_11: http://www.uniprot.org/).Citation47 Predicted protein sequences were retrieved. The final list of protein sequences for comparative sequence analysis was determined by the revision history of the sequence in the UniProt as well as by entries in GeneDBCitation48 and SchistoDB.Citation49
Conserved domain search for functional sites
The search for amino acid residues that are functionally important was performed using two public servers. A single-sequence input server (http://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)Citation50 and a multiple-sequence input or batch server (http://www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi)Citation50 were used in finding conserved domains for the sequences. The ATP-binding motif residues and other ligand-binding residues were identified and documented for all the USP sequences, including their domain architecture. To facilitate comparison of the functional sites, we constructed a functional site signature for each sequence. The signature is a string of the amino acid letters. In a case where no site is predicted in the 12-letter signature, the position was assigned “X.” Therefore, for protein sequence MJ0577, the template for the ligand-binding sites, the signature is PTDVMGHGGSVT. This approach of constructing functional sites has been implemented in previous research on functional sites.Citation51
Prediction of chemical ligand and enzymatic regulation
The three-dimensional (3D) chemical ligands were predicted using the 3DLigandSite server (http://www.sbg.bio.ic.ac.uk/3dligandsite).Citation52 These biologically relevant chemical ligands are potential regulators of the function of the USPs. The 3DLigandSite is a top-performing web server for chemical ligand prediction and provides structural models for unsolved proteins, using protein-structure prediction. The specific kinases were predicted using the NetPhosK 1.0 tool (http://www.cbs.dtu.dk/services/NetPhosK),Citation53 with a stringent threshold value set at 0.65. The kinase with the highest threshold value was selected.
Prediction of subcellular location
The subcellular locations of all the genes were retrieved from literature, databases or predicted if possible using the server Euk-mPLoc 2.0 (http://www.csbio.sjtu.edu.cn/bioinf/euk-multi-2/).Citation54 The prediction by Euk-mPLoc2.0 is based on integrating information from gene ontology, functional domain, and evolutionary relationships.
Compilation of developmental expression of genes
The developmental stage expression profiles of the selected Schistosoma USP genes were extracted from our previous publicationCitation12 for S. mansoni. In the case of S. japonicum, expression profiles were from the SjTPdb, an integrated transcriptome and proteome database and analysis platform for S. japonicum.Citation55 The database contains expressed sequence tags (ESTs), EST clusters, and the proteomic dataset for S. japonicum.
Prediction of evolutionary relatedness of sequences
Evolutionary relatedness of the selected USPs, from both S. mansoni and S. japonicum, was determined to ascertain their functional and evolutionary relationship. The sequences were aligned using the ClustalW tool (http://www.ch.embnet.org/software/ClustalW.html),Citation56 applying the default settings. The evolutionary relationship of the sequences was inferred using the maximum likelihood method,Citation57 based on the JTT matrix-based modelCitation58 at 1000 bootstrap, with MEGA software version 5 (http://www.megasoftware.net/ [Center for Evolutionary Medicine and Informatics, Tempe, AZ, USA]).Citation59 The bootstrap test indicated above, at 1000 replications, was used to determine the percentage of the replicate trees, in which the genes clustered together.Citation60,Citation61
Visual analytics of datasets
A purpose of Stage 4 of the protocol was to provide an integration and visualization portal for the results of the bioinformatics analysis in Stage 2 and Stage 3 (). The visual analytics tasks to be performed on the data sets can be influenced by how the data is organized in the data records (rows) and data fields (columns) in the data source (eg, spreadsheet file and comma delimited file). For Stage 2, each data record had the following data fields: (1) organism; (2) locus tag; (3) UniProt ID; (4) feature; and (5) feature value. The feature field had the following types: protein domain length, protein length, protein domain start position, protein domain end position, ATP-binding motif, kinase type, kinase type score, 3D chemical ligand, ligand-binding amino acid, amino acid and sequence position, developmental expression, and subcellular localization. In the case of the functional site signature data set, each record consisted of the UniProt ID and 12 fields for each of the 12-letter signatures for the USP ligand-binding sites. For the Stage 3 data set (phylogenetic tree groupings), each data record consisted of data fields for (1) organism; (2) UniProt ID; and (3) phylogenetic group. The data sources (in this case, spreadsheet files) were loaded to the visual analytics software for the visual analytics tasks, including the design of the data integration and visualization.
Results
Data set for visual analytics
The data set analyzed consisted of twelve annotation features for 13 USP sequences (). All the sequences analyzed contained the ATP-binding motif G-2x-G-9x-G(S/T). We were particularly interested in shared annotations that can help us infer the joint biochemical and environmental regulation of the USPs from the two pathogenic Schistosoma spp. The protein sequences consisted of five S. mansoni and eight S. japonicum sequences. Seven of the annotation features required only one feature value per protein sequence. These features were protein domain length, protein length, protein domain start position, protein domain end position, ATP-binding motif, kinase type, and kinase type score. The five annotation features with variable frequency per protein were 3D chemical ligand, ligand-binding amino acid, amino acid and sequence position, developmental expression, and subcellular localization. This dataset was formatted for visualization and analysis in a visual analytics resource available at http://public.tableausoftware.com/views/schisto_features_usp/feature_per_usp.
Table 1 Annotation features for universal stress proteins of Schistosoma mansoni and Schistosoma japonicum
The sequences were first grouped by protein length, protein domain length, and ligand-binding sites. Since we intend to conduct additional research on the Schistosoma USPs, the primary purpose of these groups was to prioritize protein sequences that have relatively complete and consistent annotation. The additional bioinformatics predictions from the sequences helped to confirm the sequence level observations. The comparison of the other annotation features was done in the context of the evolutionary relatedness predicted by multiple sequence alignment.
Grouping of Schistosoma USPs by sequence length
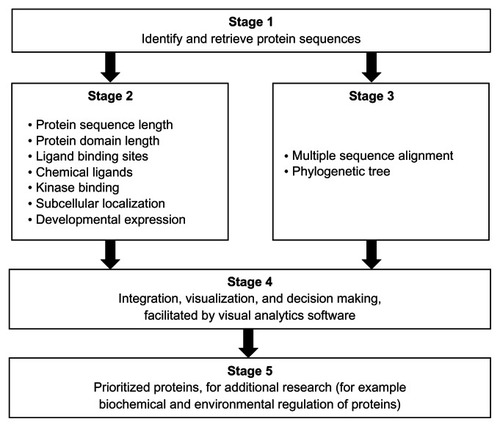
The 13 Schistosoma protein sequences were grouped by protein length and domain length (). Seven distinct domain lengths (122, 125, 136, 146, 147, 148, and 149 amino acids [aa]) were observed in the sequences compared. Ten of the 13 sequences had domain lengths from 146 aa to 149 aa. The groups with domain length types of 148 aa and 149 aa had sequences from the two species. For the protein sequence length grouping, eight distinct types (129, 133, 155, 159, 160, 172, 174, and 184 aa) were observed. Three groups (159 aa, 160 aa, and 184 aa) had members from both species. There were no groups of sequences from both species that had identical members, by protein sequence and protein domain lengths. However, we observed protein sequences from both species that shared protein sequence length with the difference in domain length of 1 aa, as in the 184 aa USPs (Q86DW2 and G4 LZI3), or 2 aa, as in the 159 aa USPs (Q5DGI9 and C1M0Q2).
Figure 2 Grouping of 13 Schistosoma USPs by sequence length.
Abbreviations: aa, amino acid; UniProt, Universal Protein Resource (Apweiler et al);Citation47 USP, universal stress protein.
Functional site signatures of Schistosoma USP sequences
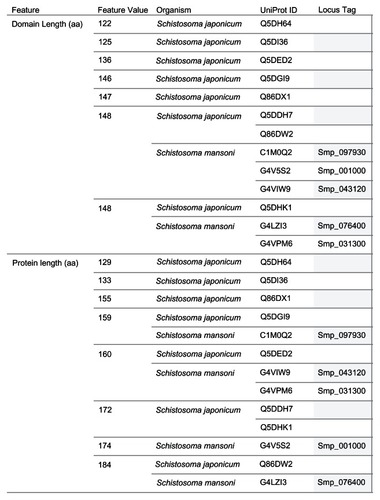
The Conserved Domain Search tool at the National Center for Biotechnology Information (NCBI) website (http://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)Citation50 uses the fragment A (UniProt Identifiers: Q57997; Y577_METJA) of the USP MJ0577 of Methanocaldococcus jannaschii as the template to align the Schistosoma USPs for the conserved protein domain search. The search reports the ligand-binding sites (functional sites) including the ATP-binding motif [G-2X-G-9X- G(S/T)] present in the query sequence. A total of 12 ligand-binding sites are predicted for MJ0577. The amino acids and their position are Pro11, Tyr12, Asp13, Val41, Met126, Gly127, His129, Gly130, Gly140, Ser141, Val142, and Thr143. The Asp13 and Val41 are binding sites for adenosine nucleoside. Citation33 The amino acid sequences from met 126 to Thr143 contain the motif G2xG9xG(S/T), which includes binding sites for the phosphoyrl and ribosyl groups of ATP. Among the 13 Schistosoma sequences with ATP-binding motif, eight functional site signatures (AIDAIGRGGSVS, AIDAVGRGGSVS, PIDVIGRGGSVS, PIDVMGRGGSVS, PVDIIGRGGSVS, PVDSMGRGGSVS, PVDVIGRGGSVS, and XXX-VMGRGGSVS) were observed (). The last seven letters (GRGGSVS) of the signature, which corresponds to the ATP-binding motif, were identical for all the signatures of the Schistosoma USPs. Three shared signatures were observed for the two species. However, only one signature (PVDIIGRGGSVS) had the same members as in the protein length grouping (184 aa: Q86DW2, G4LZI3).
Figure 3 Grouping of 13 Schistosoma universal stress proteins by functional site signature.
Abbreviations: ATP, adenosine triphosphate; UniProt, Universal Protein Resource; USP, universal stress protein.
Grouping of Schistosoma USP sequences by alignment
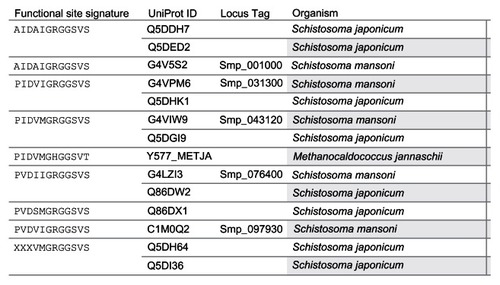
A multiple sequence alignment of the 13 sequences was generated by ClustalW (). The ligand-binding sites (functional sites) annotated in the Conserved Domain Database are labeled with hashes (#) in . The alignment revealed gap positions where amino acid residues were missing. These gaps could explain the differences in lengths reported for the sequences. In the USP functional site signatures, the first three letters were not predicted for sequences Q5DI36 and Q5DH64. The multiple sequence alignment showed where series of gaps were inserted by ClustalW to align the 13 sequences. In addition, using Smp_076400 from S. mansoni as the reference sequence, there are conserved sites (denoted by ^) of aspartate (Asp; D), leucine (Leu; L), glycine (Gly; G), histidine (His; H) and proline (Pro; P) residues at positions 57, 101, 127, 166 and 176 (). The relationship of the sequences was visualized as a phylogenetic tree (). Five groups (A to E) of sequences were observed with each of the five S. mansoni sequences assigned to a group. Groups A and E contained multiple S. japonicum sequences. The bootstrap statistical support value for the branch of Group C (Smp_076400 [G4 LZI3] and Q86DW2) was 100%. The grouping of sequences by the maximum parsimony model was in agreement with the maximum likelihood model ().
Figure 4 Multiple sequence alignment of the sequences of selected universal stress proteins of Schistosoma mansoni and Schistosoma japonicum.
Abbreviations: ATP, adenosine triphosphate; D, aspartate; G, glycine; H, histidine; L, leucine; P, proline; Uniprot, Universal Protein Resource; USP, universal stress protein.
Figure 5 Grouping of 13 Schistosoma universal stress protein sequences. The phylogenetic tree was generated with MEGA5 (Tamura et al),Citation59 using the maximum likelihood method. The 13 Schistosoma universal stress protein sequences were clustered in five groups (A–E). The numbers near the clades are the statistics from the 1000 bootstrap that support the phylogeny recovery of the clades. A visual analytics resource that can be used to view the image, with other associated data, is available at http://public.tableausoftware.com/views/schisto_features_usp/phylotrees. Sequences of S. mansoni have “Smp” in the sequence identifier.
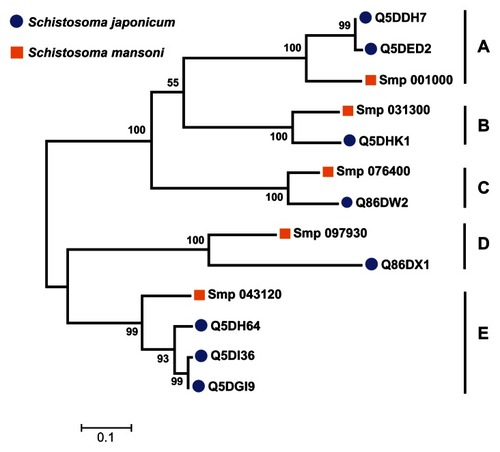
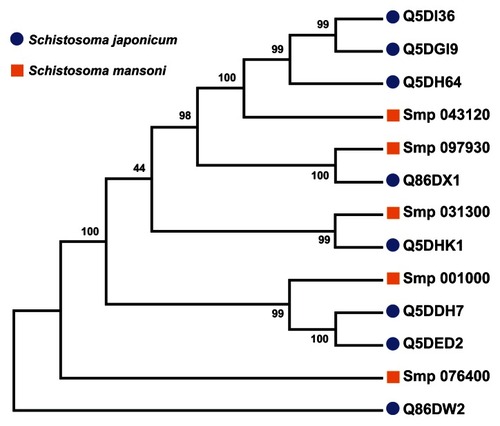
Figure 6 Parsimony test for the phylogeny tree reconstructed for Schistosoma universal stress proteins with the maximum likelihood method.
Dynamic integration of annotation features for Schistosoma USPs
The groupings described in the previous sections were based on the primary amino acid sequences of the USPs. To facilitate dynamic integration and updates of the data sources, we developed a web-based visual analytics resource. As mentioned previously, the five annotation features with variable frequency per protein were 3D chemical ligand, ligand-binding amino acid, amino acid and sequence position, developmental expression, and subcellular localization. To help guide further research and hypotheses generation, we checked for the phylogenetic groups in which the ligand amino acids were identical in amino acid type and the position of the amino acids. Other features based on the protein sequences that were considered were chemical ligands, kinase type, kinase score, and subcellular localization. The developmental expression feature was not considered in the decision-making process for prioritizing the sequences. This feature is based on extracted data from multiple peer reviewed reports and is incomplete. Nonetheless, the information can assist in directing new research.
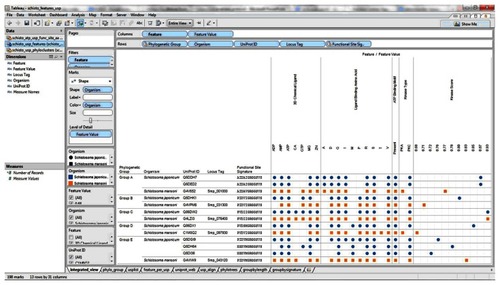
Our visual analytics-supported decision-making process prioritized for discussion Group C, which included sequence Q86DW2 from S. japonicum and sequence G4LZI3 (Smp_076400) from S. mansoni (). An integration view for Group D (Q86DX1 and C1M0Q2 [Smp_097930]) is presented to show the differences in patterns of identical annotations when compared with Group C (). The biologically relevant chemical ligands predicted to bind to the Group C proteins include four phosphate-containing ligands (adenosine diphosphate [ADP], adenosine monophosphate [AMP], adenosine triphosphate [ATP] and guanosine triphosphate [GTP]) and three metallic ion ligands (calcium [Ca2+], magnesium [Mg2+], and zinc [Zn2+]). The two proteins in the group were also predicted to be (1) localized in the cytoplasm and (2) capable of phosphorylation by phosphokinase C, with a value of 0.93. In the Group C USP genes, there was evidence of gene expression in all the stages by at least one of the genes. The schistomulum stage had the only identical annotation for the developmental gene expression for the two Group C USP genes. A screenshot showing a design that provides an integrated view of the chemical ligands, ligand-binding sites, functional site signature, the presence of ATP-binding motif, kinase type, and kinase score is presented in .
Figure 7 Integration and visualization of the data on the sequence features, evolutionary relatedness, and developmental expression of Schistosoma universal stress proteins (Q86DW2 and G4LZI3).
Abbreviations: ADP, adenosine diphosphate; AMP, adenosine monophosphate; ATP, adenosine triphosphate; CA, calcium; D, aspartate; G, glycine; GTP, guanosine triphosphate; I, isoleucine; Mg, magnesium; PKC, protein kinase C; P, proline; R, arginine; S, serine; UniProt, Universal Protein Resource (Apweiler et al);Citation47 V, valine; Zn, Zinc.
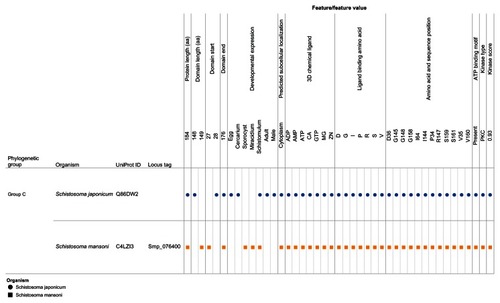
Figure 8 Integration and visualization of the data on the sequence features, evolutionary relatedness, and developmental expression of Schistosoma universal stress proteins (Q86DX1 and C1M0Q2).
Abbreviations: ADP, adenosine diphosphate; AMP, adenosine monophosphate; ATP, adenosine triphosphate; D, aspartate; G, glycine; GTP, guanosine triphosphate; I, isoleucine; M, methionine; Mg, magnesium; PKA, protein kinase A; PKC, protein kinase C; P, proline; R, arginine; S, serine; T, threonine; UniProt, Universal Protein Resource (Apweiler et al);Citation47 V, valine; Zn, Zinc.
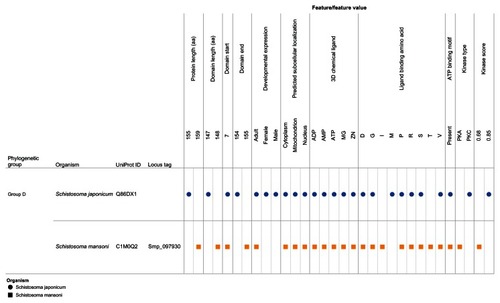
Figure 9 Design layout and visualization of data sets from the sequence analysis, evolutionary relatedness, and developmental expression of 13 Schistosoma universal stress proteins.
Abbreviations: ADP, adenosine diphosphate; AMP, adenosine monophosphate; ATP, adenosine triphosphate; CA, calcium; D, aspartate; G, glycine; GTP, guanosine triphosphate; I, isoleucine; M, methionine; Mg, Magnesium; PKA, protein kinase A; PKC, protein kinase C; P, proline; R, arginine; S, serine; T, threonine; UniProt, Universal Protein Resource (Apweiler et al);Citation47 V, valine; Zn, Zinc.
Discussion
S. haematobium, S. japonicum, and S. mansoni are the major human schistosomiasis parasites. These parasites undergo a complex developmental life cycle, in which they encounter a plethora of environmental stressors, such as transition from aerobic to anaerobic environment during the cercarial penetration of the human skin.Citation62 The presence of genes encoding the USP domain in the genomes of Schistosoma spp. suggests these flatworms are equipped to respond to unfavorable conditions that induce USP function.Citation12,Citation23,Citation25,Citation38 The bioinformatics-based predictions generated a variety of data types, including amino acid functional site, multiple sequence alignment, prediction score, protein domain organization, phylogenetic tree, and sequence length. We used a visual analytics approach to integrate these data types and to identify orthologous pairs of protein sequences with a protein length of 184 aa USP in S. mansoni (Smp_076400) and S. japonicum (Locus Tag: Sjp_0058490; UniProt ID: Q86DW2). Gene synteny, obtained from SchistoDBCitation49 and evolutionary genomics analysis called the S. mansoni phylome,Citation63 indicated that an ortholog (Sha_107834) is encoded in the S. haematobium genome. Thus, the genomes of the three major human schistosomiasis parasites encode the 184 aa USP.
Since inferences on chemical and environmental regulation are our interest, we focus the discussion of the results on the findings on the five conserved residues and the chemical ligands predicted to bind to the prioritized protein. All the 13 protein sequences have conserved sites for Asp, Leu Gly, His, and Pro at positions 57, 101, 127, 166, and 176, using Smp_076400 from S. mansoni as a reference sequence (). These conserved residues did not coincide with any of the predicted ligand-binding sites and could be common functional sites for regulating Schistosoma USPs.
The predicted 3D chemical ligands for Smp_076400 and Sjp_0058490 (Q86DW2) included three metal ions Ca2+, Mg2+, and Zn2+ (). Metal ions are involved in many diverse biochemical reactions,Citation64 including cellular cofactors for phosphorylation. The UspA protein of Escherichia coli undergoes phosphorylation in vitro with its phosphate donors ATP and/or GTP, in the absence of other proteins.Citation65 The ATP molecule and metallic chemical ligands, such as Mg2+ ion, might bind together at the Mg-ATP-binding groove during phosphorylation or ATP-dependent stress-response mechanism. Citation66,Citation67 The presence of Mg2+ ion suggests that it can be an integral and critical component in the reaction.Citation68–Citation71 This result could be affected if there is any structural conformation in the binding site residues that prevents the Mg2+ ion from binding to the ATP molecule at the active groove. The resultant effect might be translated to compromised functional efficiency in binding ATP during phosphorylation and also in keeping the metallic Mg ions unstable in the active groove while it is in contact with the ATP molecule.
Ca2+ was predicted to bind to proteins in Group C of the phylogenetic tree ( and ). In S. mansoni, Ca2+ is considered vital for regulated motor-related activitiesCitation72 and also critical for the egg hatching process in fresh water.Citation73,Citation74 In the tegument fraction of S. mansoni, Ca2+ simulated the activity of ATPase in the absence of Mg2+.Citation75 Further, cyclic adenosine monophosphate and Ca2+ work in synergy to regulate the transformation of miracidial to sporocysts.Citation76 The protein kinase C and Ca2+ metabolism regulate the induction of proteolytic enzyme from cercariae, which is vital for modulating the musculature activity of the schistosome.Citation77,Citation78 A key mechanism for the action of PZQ has been proposed to be the disruption of the Ca2+ homeostasis in schistosomes, leading to the large, rapid influx of Ca2+ ions into the worm and quick muscular contractions.Citation41,Citation79–Citation81 Microarray-based transcriptome analysis of the response of the S. mansoni PR-1 strain to PZQ has identified genes for cytosolic Ca2+ regulation.Citation82
Conclusion
S. haematobium, S. mansoni, and S. japonicum are human parasites that undergo a complex developmental life cycle, in which they encounter a plethora of environmental stressors. Though there are multiple research reports on the developmental regulation of genes encoding USPs in Schistosoma spp., knowledge of their biochemical and environmental regulation is still limited. The draft status of the genome sequences of Schistosoma spp. also provides possibilities that future revisions could be made to gene prediction and protein annotations. We have used a decision-making strategy, facilitated by visual analytics, to identify USPs in two Schistosoma species with shared sequence features and when compared with the other sequences they have relatively complete and consistent annotations. These findings further enabled us to make inferences about the biochemical and environmental regulation of Schistosoma USPs. Future research directions could (1) functionally characterize the interactions of Ca2+ ions with the amino acid residues of Schistosoma USPs; and (2) determine the transcriptional response of Schistosoma USP genes to PZQ. The data sets produced, and the visual analytics views developed, can be easily reused to develop new hypotheses.
Acknowledgments
National Institutes of Health: Research Centers in Minority Institutions (RCMI) – Center for Environmental Health at Jackson State University (NIH-NCRR 2G12RR013459); Mississippi IDeA Network for Biomedical Research Excellence (NIH-NCRR-P20RR016476 and NIH-NIGMS- 8P20GM103476); Arkansas IDeA Network for Biomedical Research Excellence (NIH-NCRR-P20RR016460); Bioinformatics Programs in Minority Institutions (1T36GM095335); and National Center for Integrative Biomedical Informatics (U54DA021519). National Science Foundation: Mississippi NSF-EPSCoR Grant Awards (EPS- 0903787 and EPS 1006883); Undergraduate Research and Mentoring Program (DBI-0958179); and Visual Analytics in Biology Curriculum Network (DBI-1062057). US Department of Homeland Security Science and Technology Directorate (2009-ST-062-000014; 2011-ST-062-000048). The views and conclusions contained in this document are those of the authors and should not be interpreted as necessarily representing the official policies, either expressed or implied, of the funding agencies.
Disclosure
The authors report no conflicts of interest in this work.
References
- NahumLAMourãoMMOliveiraGNew frontiers in Schistosomagenomics and transcriptomicsJ Parasitol Res Epub November 21, 2012
- BorchMKiernanMRustKSchistosomiasis: a case studyUrol Nurs2009291262919331272
- van der WerfMJde VlasSJBrookerSQuantification of clinical morbidity associated with schistosome infection in sub-Saharan AfricaActa Trop2003862–312513912745133
- HanZGBrindleyPJWangSYChenZSchistosoma genomics: new perspectives on schistosome biology and host-parasite interactionAnnu Rev Genomics Hum Genet20091021124019630560
- DoenhoffMHaganPCioliDPraziquantel: its use in control of schistosomiasis in sub-Saharan Africa and current research needsParasitology2009136131825183519281637
- MelmanSDSteinauerMLCunninghamCReduced susceptibility to praziquantel among naturally occurring Kenyan isolates of Schistosoma mansoniPLoS Negl Trop Dis200938e50419688043
- BerrimanMHaasBJLoVerdePTThe genome of the blood fluke Schistosoma mansoniNature2009460725335235819606141
- Schistosoma japonicum Genome Sequencing and Functional Analysis ConsortiumThe Schistosoma japonicum genome reveals features of host-parasite interplayNature2009460725334535119606140
- YoungNDJexARLiBWhole-genome sequence of Schistosoma haematobiumNat Genet201244222122522246508
- GobertGNMcManusDPNawaratnaSMoertelLMulvennaJJonesMKTissue specific profiling of females of Schistosoma japonicum by integrated laser microdissection microscopy and microarray analysisPLoS Negl Trop Dis200936e46919564906
- GobertGNMoertelLBrindleyPJMcManusDPDevelopmental gene expression profiles of the human pathogen Schistosoma japonicumBMC Genomics20091012819320991
- IsokpehiRDMahmudOMbahANDevelopmental regulation of genes encoding universal stress proteins in Schistosoma mansoniGene Regul Syst Bio201156174
- AragonADImaniRABlackburnVRCunninghamCMicroarray based analysis of temperature and oxidative stress induced messenger RNA in Schistosoma mansoniMol Biochem Parasitol2008162213414118775750
- MoertelLMcManusDPPivaTJYoungLMcInnesRLGobertGNOligonucleotide microarray analysis of strain- and gender-associated gene expression in the human blood fluke, Schistosoma japonicumMol Cell Probes200620528028916647836
- WilliamsDLSayedAABernierJProfiling Schistosoma mansoni development using serial analysis of gene expression (SAGE)Exp Parasitol2007117324625817577588
- LiuFChenPCuiSJWangZQHanZGSjTPdb: integrated transcriptome and proteome database and analysis platform for Schistosoma japonicumBMC Genomics2008930418578888
- ProtasioAVTsaiIJBabbageAA systematically improved high quality genome and transcriptome of the human blood fluke Schistosoma mansoniPLoS Negl Trop Dis201261e145522253936
- TaftASVermeireJJBernierJTranscriptome analysis of Schistosoma mansoni larval development using serial analysis of gene expression (SAGE)Parasitology2009136546948519265565
- GobertGNMcInnesRMoertelLTranscriptomics tool for the human Schistosoma blood flukes using microarray gene expression profilingExp Parasitol2006114316017216631746
- NachinLNannmarkUNyströmTDifferential roles of the universal stress proteins of Escherichia coli in oxidative stress resistance, adhesion, and motilityJ Bacteriol2005187186265627216159758
- Hingley-WilsonSMLougheedKEFergusonKLeivaSWilliamsHDIndividual Mycobacterium tuberculosis universal stress protein homologues are dispensable in vitroTuberculosis (Edinb)201090423624420541977
- RaghavanNMillerANGardnerMComparative gene analysis of Biomphalaria glabrata hemocytes pre-and post-exposure to miracidia of Schistosoma mansoniMol Biochem Parasitol2003126218119112615317
- Negrão-CorrêaDMattosACPereiraCAMartins-SouzaRLCoelhoPMInteraction of Schistosoma mansoniSporocysts and Hemocytes of BiomphalariaJ Parasitol Res Epub June 28, 2012
- OswaldIPEltoumIWynnTAEndothelial cells are activated by cytokine treatment to kill an intravascular parasite, Schistosoma mansoni, through the production of nitric oxideProc Natl Acad Sci U S A199491399910037508126
- LoverdePTDo antioxidants play a role in schistosome host-parasite interactions?Parasitology Today199814728428917040785
- SomeroGNProteins and temperatureAnnu Rev Physiol19955743687778874
- IsokpehiRDSimmonsSSCohlyHHEkunweSIBegoniaGBAyensuWKIdentification of drought-responsive universal stress proteins in viridiplantaeBioinform Biol Insights20115415821423406
- KimHGooEKangYKimJHwangIRegulation of universal stress protein genes by quorum sensing and RpoS in Burkholderia glumaeJ Bacteriol2012194598299222178971
- PinedaMCTuronXLópez-LegentilSStress levels over time in the introduced ascidian Styela plicata: the effects of temperature and salinity variations on hsp70 gene expressionCell Stress Chaperones201217443544422249790
- WangHLeiZLiXOettingRDRapid cold hardening and expression of heat shock protein genes in the B-biotype Bemisia tabaciEnviron Entomol201140113213922182622
- ZhangYYYangJYinXXYangSPZhuYGArsenate toxicity and stress responses in the freshwater ciliate Tetrahymena pyriformisEur J Protistol201248322723622342134
- FlorczykMAMcCueLAStackRFHauerCRMcDonoughKAIdentification and characterization of mycobacterial proteins differentially expressed under standing and shaking culture conditions, including Rv2623 from a novel class of putative ATP-binding proteinsInfect Immun20016995777578511500455
- ZarembinskiTIHungLWMueller-DieckmannHJStructure-based assignment of the biochemical function of a hypothetical protein: a test case of structural genomicsProc Natl Acad Sci U S A1998952615189151939860944
- FreestonePNyströmTTrineiMNorrisVThe universal stress protein, UspA, of Escherichia coli is phosphorylated in response to stasisJ Mol Biol199727433183249405142
- FreestonePTrineiMClarkeSCNyströmTNorrisVTyrosine phosphorylation in Escherichia coliJ Mol Biol19982795104510519642082
- LiuWTKaravolosMHBulmerDMRole of the universal stress protein UspA of Salmonella in growth arrest, stress and virulenceMicrob Pathog200742121017081727
- RaspoetRGantoisIDevlooRSalmonella enteritidis universal stress protein (usp) gene expression is stimulated by egg white and supports oviduct colonization and egg contamination in laying hensVet Microbiol20111531–218619021715109
- Seifart GomesCIzarBPazanFUniversal stress proteins are important for oxidative and acid stress resistance and growth of Listeria monocytogenes EGD-e in vitro and in vivoPLoS One201169e2496521980369
- Salvador-RecatalàVGreenbergRMCalcium channels of schistosomes: unresolved questions and unexpected answersWiley Interdiscip Rev Membr Transp Signal201211859322347719
- GreenbergRMAre Ca2+ channels targets of praziquantel action?Int J Parasitol20053511915619510
- ColesGCThe effect of praziquantel on Schistosoma mansoniJ Helminthol19795313133458128
- ChabotCDemystifying visual analyticsIEEE Comput Graph Appl2009292848719462638
- ChangRZiemkiewiczCGreenTMRibarskyWDefining insight for visual analyticsIEEE Comput Graph Appl2009292141719462629
- JohnsonMOCohlyHHIsokpehiRDAwofoluORThe case for visual analytics of arsenic concentrations in foodsInt J Environ Res Public Health2010751970198320623005
- MackinlayJDHanrahanPStolteCShow me: automatic presentation for visual analysisIEEE Trans Vis Comput Graph20071361137114417968057
- ThomasJJCookKAA visual analytics agendaIEEE Comput Graph Appl2006261101316463473
- UniProt ConsortiumOngoing and future developments at the Universal Protein ResourceNucleic Acids Res201139D214D21921051339
- Hertz-FowlerCPeacockCSWoodVGeneDB: a resource for prokaryotic and eukaryotic organismsNucleic Acids Res200432D339D34314681429
- ZerlotiniAHeigesMWangHSchistoDB: a Schistosoma mansoni genome resourceNucleic Acids Res200937D579D58218842636
- Marchler-BauerALuSAndersonJBCDD: a Conserved Domain Database for the functional annotation of proteinsNucleic Acids Res201139D225D22921109532
- SalsburyFRJrKnutsonSTPooleLBFetrowJSFunctional site profiling and electrostatic analysis of cysteines modifiable to cysteine sulfenic acidProtein Sci200817229931218227433
- WassMNKelleyLASternbergMJ3DLigand Site: predicting ligand-binding sites using similar structuresNucleic Acids Res201038W469W47320513649
- BlomNSicheritz-PonténTGuptaRGammeltoftSBrunakSPrediction of post-translational glycosylation and phosphorylation of proteins from the amino acid sequenceProteomics2004461633164915174133
- ChouKCShenHBA new method for predicting the subcellular localization of eukaryotic proteins with both single and multiple sites: Euk-mPLoc 2.0PLoS One201054e993120368981
- LiuFLuJHuWWangSYNew perspectives on host-parasite interplay by comparative transcriptomic and proteomic analyses of Schistosoma japonicumPLoS Pathogens200624e2916617374
- LarkinMBlackshieldsGBrownNPClustal W and Clustal X version 2.0Bioinformatics200723212947294817846036
- TamuraKNeiMEstimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzeesMol Biol Evol19931035125268336541
- JonesDTTaylorWRThortonJMThe rapid generation of mutation data matrices from protein sequencesComput Appl Biosci1992832752821633570
- TamuraKPetersonDPetersonNStecherGNeiMKumarSMEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methodsMol Biol Evol201128102731273921546353
- FelsensteinJConfidence limits on phylogenies: an approach using the bootstrapEvolution1985394783791
- SteelMLockhartPJPennyDA frequency-dependent significance test for parsimonyMol Phylogenet Evol19954164717620636
- JollyERChinCSMillerSGene expression patterns during adaptation of a helminth parasite to different environmental nichesGenome Biol200784R6517456242
- SilvaLLMarcet-HoubenMNahumLAZerlotiniAGabaldónTOliveiraGThe Schistosoma mansoni phylome: using evolutionary genomics to gain insight into a parasite’s biologyBMC Genomics20121361723148687
- LuCHLinYFLinJJYuCSPrediction of metal ion-binding sites in proteins using the fragment transformation methodPLoS One201276e3925222723976
- FreestonePNyströmTTrineiMNorrisVThe universal stress protein, UspA, of Escherichia coli is phosphorylated in response to stasisJ Mol Biol199727433183249405142
- BuchachenkoALKouznetsovDAArkhangelskySEOrlovaMAMarkarianAASpin biochemistry: magnetic 24 Mg-25 Mg-26 Mg isotope effect in mitochondrial ADP phosphorylationCell Biochem Biophys200543224325116049349
- BuchachenkoALKouznetsovDABreslavskayaNNOrlovaMAMagnesium isotope effects in enzymatic phosphorylationJ Phys Chem B200811282548255618247604
- LuoRZhouCLinJYangDShiYChengGIdentification of in vivo protein phosphorylation sites in human pathogen Schistosoma japonicum by a phosphoproteomic approachJ Proteomics201275386887722036931
- NeblTPrietoJHKappEQuantitative in vivo analyses reveal calcium-dependent phosphorylation sites and identifies a novel component of the Toxoplasma invasion motor complexPLoS Pathog201179e100222221980283
- SzaboALangerTSchröderHFlanaganJBukauBHartlFUThe ATP hydrolysis-dependent reaction cycle of the Escherichia coli Hsp70 system DnaK, DnaJ, and GrpEProc Natl Acad Sci U S A1994912210345103497937953
- LangerTLuCEcholsHFlanaganJHayerMKHartlFUSuccessive action of DnaK, DnaJ and GroEL along the pathway of chaperone-mediated protein foldingNature199235663716836891349157
- Soares de MouraRRozentalRClaudio-da-SilvaTSSchistosoma mansoni: effects of bromolysergic acid diethylamide, verapamil, and Ca2+-free solution on the motor activity of the isolated male worm induced by electrical stimulation and oxamniquineExp Parasitol19876321731792883021
- KatsumataTKohnoSYamaguchiKHaraKAokiYHatching of Schistosoma mansoni eggs is a Ca2+/calmodulin-dependent processParasitol Res198976190912622901
- KatsumataTShimadaMSatoKAokiYPossible involvement of calcium ions in the hatching of Schistosoma mansoni eggs in waterJ Parasitol1988746104010412461448
- CunhaVNoëlFA Mg2+-independent Ca2+-stimulated ATPase activity in the tegument of Schistosoma mansoniBraz J Med Biol Res19882134494512976283
- KawamotoFShozawaAKumadaNKojimaKPossible roles of cAMP and Ca2+ in the regulation of miracidial transformation in Schistosoma mansoniParasitol Res19897553683742542928
- BlairKLBennettJLPaxRASchistosoma mansoni: evidence for protein kinase-C-like modulation of muscle activityExp Parasitol19886622432523165068
- MatsumuraKMitsuiYSatoKSakamotoMAokiYSchistosoma mansoni: possible involvement of protein kinase C in linoleic acid-induced proteolytic enzyme release from cercariaeExp Parasitol19917233113202015870
- DoenhoffMJCioliDUtzingerJPraziquantel: mechanisms of action, resistance and new derivatives for schistosomiasisCurr Opin Infect Dis200821665966718978535
- GreenbergRMCa2+ signalling, voltage-gated Ca2+ channels and praziquantel in flatworm neuromusculatureParasitology2005131 SupplS97S10816569296
- Pica-MattocciaLOrsiniTBassoASchistosoma mansoni: lack of correlation between praziquantel-induced intra-worm calcium influx and parasite deathExp Parasitol2008119333233518456260
- AragonADImaniRABlackburnVRTowards an understanding of the mechanism of action of praziquantelMol Biochem Parasitol20091641576519100294