Abstract
The phosphoinositide 3-kinase (PI3-K) signaling pathway plays an important role in a wide variety of fundamental cellular processes, largely mediated via protein kinase B/v-akt murine thymoma viral oncogene homolog (PKB/AKT) signaling. Given the crucial role of PI3-K/AKT signaling in regulating processes such as cell growth, proliferation, and survival, it is not surprising that components of this pathway are frequently dysregulated in cancer, making the AKT kinase family members important therapeutic targets. The large number of clinical trials currently evaluating PI3-K pathway inhibitors as a therapeutic strategy further emphasizes this. The serum- and glucocorticoid-inducible protein kinase (SGK) family is made up of three isoforms, SGK1, 2, and 3, that are PI3-K-dependent, serine/threonine kinases, with similar substrate specificity to AKT. Consequently, the SGK family also regulates similar cell processes to the AKT kinases, including cell proliferation and survival. Importantly, there is emerging evidence demonstrating that SGK3 plays a critical role in AKT-independent oncogenic signaling. This review will focus on the role of SGK3 as a key effector of AKT-independent PI3-K oncogenic signaling.
Keywords:
Introduction
The phosphoinositide 3-kinase (PI3-K) pathway integrates signals from growth factors, insulin, nutrients, and oxygen to initiate a plethora of downstream responses. This pathway is frequently dysregulated in human cancer and regulates many of the hallmarks of cancer,Citation1 including cell growth, proliferation, survival, migration, metabolism, and angiogenesis, as shown in ; thus, it is a pivotal target for cancer therapy.Citation2–Citation4 To date, much of the evidence gathered supporting PI3-K as a critical modulator of tumor formation and progression has revealed the main downstream effector to be the v-akt murine thymoma viral oncogene homolog (AKT) family of kinases (AKT1, 2, and 3), with all three AKT isoforms playing both overlapping and distinct roles in cell transformation and tumorigenesis.Citation5 However, despite this paradigm for PI3-K-dependent transformation via AKT, there are multiple alternative oncogenic pathways, such as mitogen-activated protein kinase (MAPK) cascade, liver kinase B1 (LKB1) and v-myc myelocytomatosis viral oncogene (MYC), that interact with the core PI3-K signaling module both upstream and downstream of AKT.Citation6–Citation9 The importance of the PI3-K pathway in a number of cancer types has been well established,Citation10–Citation12 with genetic defects leading to the hyperactivation of the PI3-K cascade occurring in many cancers.
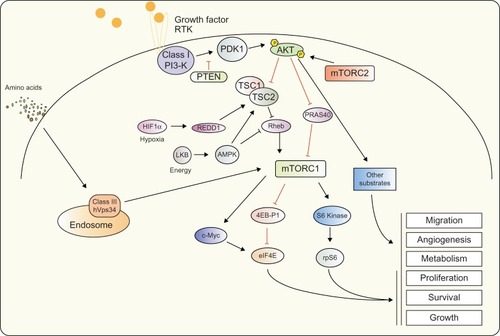
Figure 1 PI3-K signaling.
Notes: Activation of class I PI3-K via RTK phosphorylates and activates downstream target PDK1, in turn phosphorylating AKT kinases at the threonine site. Full activation of AKT requires phosphorylation at the serine residue by the mTORC2. Activated AKT mediates a plethora of effects through phosphorylation of a number of downstream targets including TSC2 and PRAS40. Additional factors such as HIF1α, LKB, c-MYC and the RAS signaling pathway are also able to link into PI3-K signaling at multiple levels and contribute to protein synthesis and cell growth signaling. At the endosome, class III PI3-K hVps34 is able to mediate amino acid signaling to mTORC1.
Abbreviations: AKT, v-akt murine thymoma viral oncogene homolog; AMPK, 5′ AMP-activated protein kinase; HIF1α, hypoxia-inducible factor 1 alpha; hVps34, class III PI3-K human vacuolar sorting protein 34; LKB, liver kinase B1; mTORC1, mammalian target of rapamycin complex 1; mTORC2, mammalian target of rapamycin complex 2; PDK1, 3-phosphoinositide-dependent kinase 1; PI3-K, phosphoinositide 3-kinase; PRAS40, proline-rich AKT substrate of 40 kDa; PTEN, phosphatase and tensin homolog; RTK, receptor tyrosine kinases; TSC1, tuberous sclerosis factor 1; TSC2, tuberous sclerosis factor 2; Rheb, Ras homolog enriched in brain; REDD1, DNA damage inducible transcript 4; eIF4E, eukaryotic translation initiation factor 4E; 4EB-P1, eukaryotic translation initiation factor 4E binding protein 1; rpS6, ribosomal protein S6.
Despite AKT signaling being a major downstream effector of PI3-K signaling, there is mounting evidence to suggest the importance of other signaling factors downstream of PI3-K that act independently of AKT to mediate important cell processes that are involved in malignant transformation. Recent studies focusing on 3-phosphoinositide-dependent kinase 1 (PDK1), an immediate downstream effector of PI3-K (as illustrated in ), have demonstrated that in some tumor models, following knockdown of PDK1, the overexpression of activated AKT is not enough to restore malignant phenotypes, suggesting a subset of tumors that are PI3-K/PDK1-dependent but AKT-independent.Citation13 Further, many of these studies have also demonstrated that in malignancy driven by AKT-independent factors, expression of activated AKT is quite low and correlates poorly with PIK3CA oncogenic mutations.Citation14,Citation15 Together, these studies indicate the presence of a PI3-K/PDK1 downstream target capable of driving oncogenic signaling independently of the AKT kinases, and active in a subset of tumors.
PI3-K activation of PDK1 can induce the activity of several protein kinases, including protein kinase C zeta and the serum- and glucocorticoid-inducible protein kinase (SGK) isoforms, all of which have the potential to contribute to the tumorigenic phenotype.Citation16 The PI3-K pathway leads to activation of the SGK isoforms, in a PDK1-dependent manner, promoting SGK isoform expression and activity.Citation17 All three isoforms of SGK share high structural and functional similarities to the AKT kinases, with many studies demonstrating the SGK isoforms have important roles in PI3-K signaling, both in normal cell physiology and pathophysiology.Citation18,Citation19 Recently, there have been a number of reports that implicate SGK3 as a critical mediator of malignant transformation independently of AKT.Citation20,Citation21 Thus, this review will focus on SGK3 as an alternate signaling effector of PI3-K in tumorigenesis.
PI3-K signaling
The PI3-K family is integral in a variety of cellular processes, coordinating the localization and activity of a multitude of important downstream effector proteins, including the SGK and AKT families of kinases. The PI3-K family is made up of three distinct classes (class I, II, and III), all of which are grouped according to structure, function, and lipid substrate preference.Citation3 The class I PI3-K family can be further divided into class IA and IB, both of which are heterodimers made up of a catalytic subunit and a regulatory subunit. Class 1A PI3-K are made up of one of the three different p110 catalytic subunits (p110α, p110β, and p110δ), all products of separate genes (PIK3CA, PIK3CB, and PIK3CD, respectively), and one of the five p85 regulatory subunits (including p85α, p55α, and p50α), all of which are splice variants of a single gene (PIK3R1), with the remaining two p85 variants (p85β and p55γ) both products of separate genes (PIK3R2 and PIK3R3), respectively.Citation22
The class IB PI3-K family can be distinguished from IA, as the catalytic subunit p110γ, encoded by gene PIK3CG, does not bind p85 regulatory subunit but instead binds regulatory subunits p101 or p87, encoded by genes PIK3R5 and PIK3R6, respectively. The inability to bind the p85 regulatory subunit has upstream signaling consequences in that p85 regulatory subunit contains Src homology 2 domains (SH2), which are able to bind phosphorylated tyrosine, correlating with its ability to be activated through receptor tyrosine kinases (RTKs), therefore enabling class IA to be activated by RTKs, and class 1B to be activated by G protein–coupled receptors (GPCR). Both class IA and 1B can be activated indirectly by RAS.Citation23
Canonical signaling via the class I PI3-K is activated by growth factor RTKs in addition to G-protein-coupled receptors. Once activated PI3-K phosphorylates phosphoinositides, generating the lipid products phosphatidylinositol-3-4-bisphosphate (PI(3,4)P2) and phosphatidylinositol-3,4,5-triphosphate (PI(3,4,5)P3), allowing PI(3,4,5)P3 to recruit Pleckstrin Homology (PH) domain–containing proteins to the plasma membrane, such as PDK1 and AKT, for activation via phosphorylation at key residues.Citation24–Citation26 Once at the plasma membrane, PDK1 is able to phosphorylate and fully activate AKT at threonine 308 following phosphorylation of AKT at the serine 473 site by mTORC2 (mammalian target of rapamycin complex 2).Citation27 Activated AKT has many downstream targets, implicating PI3-K/AKT signaling in processes such as cell growth, proliferation, survival, metabolism, and angiogenesis.
Additionally, a number of other important pathways involved in malignant transformation have also been reported to intersect and cooperate with PI3-K signaling downstream of AKT, including energy sensing via LKB1,Citation28 mitogens via MAPK, hypoxia via regulated in development and DNA damage responses 1 (REDD1), and c-Myc, as shown in .Citation1,Citation9 Further, reports demonstrate that nutrient signaling via the endosomally localized class III PI3-K human vacuolar sorting protein 34 (hVps34) is involved in mTOR regulation downstream of AKT While both the SGK and AKT kinases have shown to be phosphorylated and activated in a class I PI3-K-dependent manner, the SGK3 kinase through its N-terminal phox homology (PX) domain is localized to the endosome. Consequently, investigation into a possible interaction between hVps34 and SGK3 at the endosome may link SGK3 with mediating nutrient signaling via mTOR complex 1 (mTORC1).
Class I PI3-K
The class IA PI3-K signaling cascade is a crucial modulator linking the activation of multiple receptor classes to many core cell processes, including cell cycle, cell survival, protein synthesis, growth, metabolism, motility, and angiogenesis, via key signaling intermediates, including the SGK and AKT families.Citation22,Citation29 Numerous reports have demonstrated an active role for this pathway in many types of human cancers, with one or more of its signaling components exhibiting constitutive activation due to a genetic aberration, which ultimately leads to a malignant phenotype.Citation2,Citation24 Dysregulation of several components of the PI3-K pathway, including PI3-K itself, PTEN (phosphatase and tensin homolog), and AKT, have been demonstrated in human cancer, with probably the most prevalent affecting the PIK3CA gene and tumor suppressor PTEN.Citation30 Furthermore, receptor tyrosine kinases such as epidermal growth factor receptor (EGFR), human epidermal growth factor receptor 2 (HER2), and platelet-derived growth factor receptor (PDGFR) are often up-regulated in human cancer and engage the PI3-K pathway via interaction with the p85 regulatory subunit.Citation31–Citation33
The PIK3CA gene, located on chromosome 3, encodes for the p110α subunit of class IA PI3-K, and is either mutated or amplified in a number of different cancer types.Citation34–Citation36 PIK3CA knockout mouse embryonic fibroblasts are deficient in cellular signaling in response to various growth factors, and resistant to oncogenic transformation induced by RTKs,Citation37 together demonstrating that PI3-K is involved in growth factor signaling and fundamental to tumorigenesis. The somatic missense mutations affecting the PIK3CA gene have been mapped to hotspot regions, exon 9, which encodes the helical domain of p110α, and exon 20, which encodes the catalytic domain of p110α.Citation38,Citation39 These mutations constitutively activate AKT through increased production of PI(3,4,5)P3 and induce oncogenic transformation both in vitro and in vivo.Citation30,Citation40–Citation45 In addition to the frequent hot spot mutations, almost 100 rare mutations have also been identified in PIK3CA.Citation46,Citation47 The PIK3R1, which encodes for the p85 subunit of PI3-K, also exhibits mutations in colorectal and ovarian cancers, which result in overactivity of PI3-K signaling through loss of p85 subunit inhibition of the p110 catalytic subunit of PI3-K.Citation40,Citation48 Class II PI3-K comprises three catalytic isoforms (C2α, C2β, and C2γ) which generate both phosphatidylinositol 3-phosphate (PI(3)P) and PI(3,4) P2,Citation49 and while PI(3)P localizes SGK3 to the endosome, it is the class I PI3-K that have shown to be associated with SGK3 activation. Further, the class III PI3-K hVps34, has shown to be localized at the endosome in addition to producing PI(3)-P only, making it a potential candidate for mediating SGK3 function at the endosome. Thus, both class I and class III PI3-K are likely to be the most relevant of the PI3-K classes in SGK3 activation and function.
Class III PI3-K
The class III PI3-K family consists of only one catalytic subunit, hVps34, localized at the early endosome, which was originally identified in a screen for genes involved in endosomal sorting in Saccharomyces cerevisiae.Citation50 hVps34 forms a constitutive heterodimer with Vps15, and has shown a limited substrate specificity of only inositol-containing lipids (PtdIns), thereby producing a single lipid product phosphatidylinositol 3-phosphate (PI(3)P), allowing it to function in the recruitment of proteins containing PI(3)P binding domains (PX domains) to intracellular membranes.Citation50–Citation52 Many studies have demonstrated an important role for hVps34 in vesicular trafficking in the mammalian endosomal system,Citation53 with stable hVps34 knockdown blocking the formation of multivesicular body formation, and slowing receptor degradation. However, more recent studies in mammalian systems have also recognized that hVps34 is involved in autophagy through association with Beclin-1, and nutrient sensing through signaling to mTOR.Citation54–Citation57 hVps34 has shown involvement in the regulation of the mTOR pathway through studies involving hVps34 knockdown, which demonstrated a block in insulin-stimulated phosphorylation of both S6 kinase 1 (S6K1) and eukaryotic initiating factor 4E binding protein 1 (4EB-P1), both key downstream effectors in the mTORC1 growth signaling pathway and readouts of mTORC1 activity.Citation50 Further, overexpression of hVps34 activates S6K1 in the absence of insulin stimulation; conversely, hVps34 knockdown blocks amino acid stimulation of S6K1.
Growth factor regulated pathways leading to the activation of mTORC1 via AKT have been extensively characterized, while the mechanisms by which nutrients are able to activate mTORC1 remains ill-defined.Citation57 Earlier studies have demonstrated that amino acid-dependent activation of mTORC1 requires the Rag guanosine triphosphate (GTP) ases,Citation58,Citation59 while additional studies have implicated other proteins, including MAP4K3 (mitogen-activated protein kinase kinase kinase kinase),Citation60 and inositol polyphosphate monokinase (IMPK);Citation61 however, how these molecules interact to mediate nutrient signaling requires further investigation. The class III PI3-K hVps34 has also been implicated in nutrient signaling to mTORC1; this regulation is dependent on the associated kinase hVps15 and independent of TSC (tuberous sclerosis complex).Citation54,Citation55 The ability of SGK3 to selectively bind PI(3)P, targeting it to the early endosomes where it is fully activated, suggests a pool of endosomally localized upstream signaling factors such as class I PI3-K and PDK1 may be available for SGK3 activation.Citation19 The class III PI3-K hVps34 has not been shown to be directly involved in SGK3 signaling; however, endosomally localized hVps34 mediates nutrient signaling to mTOR and specifically generates the lipid product PI(3)P, while SGK3 binds PI(3)P, allowing it to be localized to the endosome, where it is activated and can signal to growth via mTORC1. Thus, it is plausible that a growth signaling connection may exist between hVps34 and SGK3, contributing to oncogenic cell growth during cell transformation and tumorigenesis. If so, this would represent an important new aspect to understanding AKT-independent regulation of nutrient signaling.
AKT as an established effector of PI3-K signaling
The PI3-K/AKT pathway has been identified as a crucial node of growth and proliferation through the ability of AKT to regulate mTORC1, which mediates the coordinate growth factor and nutrient signaling. mTORC1, through convergence on downstream targets S6K and 4EB-P1, regulates core growth processes, including ribosome biogenesis, transcription, translation initiation, and protein degradation.Citation62–Citation65 Many studies have identified AKT as an important modulator of mTORC1, and thus cell growth and proliferation. As shown in , AKT phosphorylates the tumor suppressor tuberous sclerosis factor 2 (TSC2), a crucial negative regulator of mTORC1, at two distinct sites (serine 939 and threonine 1462), thereby inhibiting TSC2 function and promoting mTORC1 activation.Citation4,Citation66,Citation67 Furthermore, AKT has also been shown to phosphorylate a proline-rich AKT substrate of 40 kDa (PRAS40), a protein associated with mTORC1. Phosphorylation of PRAS40 at threonine (Thr)246 by AKT prompts its dissociation from mTORC1 and subsequently indirectly activates mTORC1 signaling.Citation68,Citation69 In addition, many reports demonstrate a role for AKT in cell proliferation through the regulation of cyclin dependent kinase (CDK) inhibitors and glycogen synthase kinase β (GSK3β) via PI3-K signaling,Citation70 in addition to cell survival through regulation of forkhead transcription factor 3a (FOXO3a),Citation71 Bcl-2 associated death promoter (BAD),Citation72 murine double minute 2 (MDM2),Citation73 and the nuclear factor κB (NF-κB) pathway.Citation74 AKT can also directly modulate ribosome biogenesis independent of TOR, thus promoting growth and proliferation.Citation75
Much of the aberrant regulation through the PI3-K pathway observed in tumorigenesis is associated with hyperactivation of AKT. Although dysregulation of upstream signaling stimulates AKT activity, the akt1 gene has also found to be amplified, in head and neck, gastric, pancreatic, and ovarian tumors.Citation76–Citation78 Furthermore, a missense mutation identified in the pleckstrin homology domain of akt1 has been described at low frequency in breast, colorectal, and ovarian cancers,Citation79 which leads to targeting of AKT1 to the plasma membrane, constitutive activation of the kinase and enhanced downstream signaling. Genetic aberrations associated with akt2 and akt3 have also been reported, with akt2 frequently amplified in ovarian and breast cancer,Citation77 along with an activation of AKT2 kinase activity in approximately 36% of ovarian tumors.Citation80 An increase in akt3 copy number has also been observed in approximately 70% of sporadic melanomas,Citation81 and AKT3 has shown to be overexpressed in 19 of 92 primary ovarian tumors, showing up to tenfold higher specific activity than AKT1, potentially amplifying any effect of AKT3 overexpression.Citation82 Further, an analysis of frequency for which 316 advanced-stage high-grade serous ovarian cancers harbored one or more mutations, copy number changes or changes in gene expression in the PI3-K/rat sarcoma viral oncogene homolog (RAS) pathway were shown to be deregulated in 45% of cases,Citation83 demonstrating the importance of this pathway in oncogenic pathophysiology.
AKT-independent PI3-K signaling to cancer
While AKT is considered to be the key downstream effector of PI3-K oncogenic signaling, there have been a number of recent studies demonstrating that in many cases there is an AKT-independent signaling node that also contributes to malignant transformation. A recent study to investigate the role of PDK1 in tumor progression using breast cancer cell lines harboring either PIK3CA or KRAS gain of function mutations demonstrated that PDK1 knockdown led to increased anoikis, reduced anchorage independent growth, and apoptosis in breast tumors. Interestingly, the expression of activated AKT was unable to rescue the PDK1-dependent, anchorage-independent growth phenotype, suggesting a PDK1-dependent, AKT-independent signaling node in breast cancer.Citation13 Furthermore, a model of human ovarian endometrioid adenocarcinoma, based on somatic defects in the wingless-related MMTV integration site (Wnt)/β-Catenin and PI3-K/PTEN signaling pathways,Citation84 demonstrated equivalent pPDK1 and phospho ribosomal protein S6 (pRPS6) levels but relatively low levels of pAKT,Citation14 suggesting that these mutations may drive tumor formation via an AKT-independent mechanism. Similarly, prostate-specific loss of PTEN in a murine model resulted in tumors with elevated AKT and mTORC1 activity. However, surprisingly, the inhibition of AKT resulted in little effect on tumor growth, implying that PI3-K-dependent but AKT-independent signaling was driving tumorigenesis.Citation85 Furthermore, in both breast and ovarian cancers, AKT activity has been shown to correlate poorly with PIK3CA mutations,Citation15 suggesting alternative PI3-K-dependent mediators of tumorigenesis driven by mutant PIK3CA.
It has been proposed that the SGK protein isoforms are likely candidates for PI3-K signaling to tumorigenesis, independent of AKT, given that they play roles in cell survival, proliferation, and growth, and share many of the same substrates with AKT.Citation18 The most convincing data to support this hypothesis comes from studies performed using a lentiviral short hairpin RNA (shRNA) library targeting >1000 kinases, phosphatases, and other cancer genes. Using this library to screen PIK3CA mutant tumor cells with minimal AKT activation, SGK3 was identified as a PDK1 substrate that conferred increased cell viability. Furthermore, these cells had a critical reliance on SGK3 for anchorage independent growth,Citation21 thus indicating that in the absence of AKT signaling SGK3 is able to drive certain aspects of malignant cell transformation. Another study examining SGK3 expression in estrogen receptor–positive breast tumors identified a positive correlation between SGK3 level and patient survival and prognosis, where previous analyses had not found a correlation between AKT messenger (m)RNA expression and tumor prognosis.Citation86,Citation87 These observations suggest that SGK3 may be an important downstream effector for many breast and ovarian cancers harboring PIK3CA mutations and reduced AKT signaling, and thus a potential alternative therapeutic target for the treatment of these malignancies. Furthermore, they also raise the possibility that SGK activation is a mechanism of resistance to AKT inhibitors. Indeed, recent studies in breast cancer cell lines show cells that express high levels of SGK1 were resistant to AKT inhibition.Citation88
SGK3 has also been identified as a crucial effector of PI3-K/AKT-independent signaling in the pathogenesis of hepatocellular carcinoma (HCC). Amplification and overexpression of SGK3 is more common than AKT in HCC, suggesting it may have a greater functional significance in the biology of this cancer. For example, HCC tumor tissue demonstrated a significant increase in SGK3 transcript expression when compared with paired nontumor tissue. Moreover, in vitro functional assays demonstrated that enforced expression of SGK3 was able to increase cell growth, colony formation, and anchorage-independent growth in HCC cells, while SGK3 knockdown was able to significantly decrease these processes.Citation20 Furthermore, xenograft models overexpressing SGK3 in a human HCC cell line demonstrated tumor formation in four out of the five mice injected, while mice injected with empty vector cell lines exhibited no tumor growth in any mice. Finally, overexpression of SGK3 significantly correlates with poor overall survival of HCC patients (P = 0.028).Citation20 However, in contrast to these studies demonstrating a role for SGK3 in AKT-independent oncogenic signaling, a recent report failed to demonstrate a role for SGK3 in mediating aberrant PI3-K activity in a panel of ovarian tumor samples exhibiting low AKT activation.Citation89 Specifically, tumors presenting with low phosphorylated AKT but with high PIK3CA, SGK3 activation was detected in only 36% of ovarian tumors, and SGK3 phosphorylation did not correlate with phosphorylated PIK3CA overexpression or AKT activation. Further, activated SGK3 was detected in only three out of the nine ovarian tumors that were positive for phosphorylated PIK3CA and negative for phosphorylated AKT, suggesting that while SGK3 is likely not implicated in all aberrant PI3-K oncogenic signaling, it is consistent with SGK3 playing a role in a subset of tumors. Clearly, further studies in larger patient cohorts are required to more definitively delineate the role of SGK3 in aberrant PI3-K oncogenic signaling.
SGK3 – a unique member of the SGK family
Studies using murine interleukin-3 (IL-3)-dependent 32D cells identified the mouse homolog of human SGK3, known as cytokine independent survival kinase (CISK), in a genetic screen to identify factors that mediate IL-3-dependent survival of hematopoietic cells.Citation90 Several splice variants for sgk3 have also been identified. The human gene encoding sgk3 (also referred to as sgk-l (serum/glucocorticoid regulated kinase-like)) is localized to chromosome 8q12.2;Citation91 it is ubiquitously expressed at the mRNA level, although mRNA abundance can vary considerably from tissue to tissue (www.genecards.org). Although constitutively expressed, sgk3 has estrogen receptor–binding regions and can be transcriptionally induced with estrogen.Citation92
SGK3 is unique within the SGK family as it contains an N-terminal PX domain, as shown in , initially shown to be important for targeting SGK3 to vesicle-like structures.Citation90 The PX domain in many proteins acts as a specific phosphoinositide-binding module, which has varying lipid-binding specificities. The most common binding specificity for the PX domain appears to be for PI(3)P; hence, several PX domain-containing proteins localize to PI(3)P-rich endosomal and vacuolar structures through this domain.Citation93 SGK3 binds strongly and selectively to PI(3)P through its PX domain, which is required for targeting SGK3 to the endosomal compartment.Citation94 Mutation of the SGK3 PX domain at the phospholipid binding pocket diminishes phospholipid binding, endosomal localization, and SGK3 activity.Citation95 SGK3 localization and activation at the endosome is also discussed in the Class III PI3-K section.
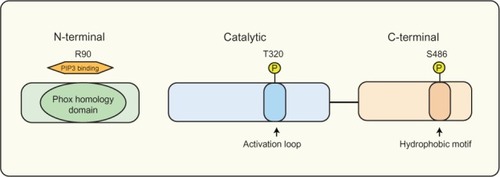
Figure 2 The SGK3 protein domain structure.
Notes: SGK3 variants containing a PX domain in the N-terminal region between amino acids 12–120, allowing SGK3 to bind to PI(3)P, and localize to the early endosomes. SGK3 has two key regulatory sites, consisting of Serine 486 in the C-terminal hydrophobic motif and Threonine 320 in the activation loop of the catalytic domain, both of which require phosphorylation for complete activation.
Abbreviations: PI(3)P, phosphatidylinositol 3-phosphate; PX, phox homology; SGK3, serum and glucocorticoid inducible kinase 3; T, threonine; S, serine.
The catalytic domain of SGK3 shares significant amino acid identity with the AKT kinases,Citation96 and importantly contains a functional serine/threonine protein kinase domain, which includes lysine 191 in the adenosine triphosphate (ATP) binding site and threonine 320 in the activation loop. Both of these sites require phosphorylation for full catalytic activity.Citation97 The C-terminal hydrophobic domain of SGK3 contains a second phosphorylation site, serine 486, which is required for complete kinase activation.Citation96 There is evidence that mTOR complex 2 (mTORC2) controls the phosphorylation of SGK1’s hydrophobic motif and thus its activation.Citation98–Citation100 While the kinase responsible for phosphorylation of SGK3 at the homologous serine (Ser) residue within the hydrophobic motif remains undefined, the strong sequence conservation in this domain among the SGK isoforms indicates it is also likely to be mTORC2. The development of reliable phospho-specific SGK3 antibodies would assist in further characterizing the role of mTORC2 in SGK3 regulation. All SGK isoforms are enzymatically activated via phosphorylation in a PI3-K-dependent manner.Citation101,Citation102 SGK3 phosphorylation and activation has shown to be stimulated by oxidation, insulin and insulin growth factor 1 (IGF-I),Citation94,Citation103 and specifically by IL-3Citation90 and estrogen.Citation92
While studies have demonstrated that SGK3 is activated in a class I PI3-K-dependent manner via PDK1, to date there have been no reports demonstrating that the class III PI3-K family directly or indirectly interacts with SGK3. However, the unique localization of SGK3 at the early endosomes, where the class III PI3-K family catalytic subunit hVps34 resides, raises the possibility that SGK3 may potentially modulate nutrient signaling via interaction with hVps34, in a manner independent of AKT. In support of this, increases in intracellular amino acid levels such as leucine have shown an increase in phosphorylation of mTORC1 effectors, S6K1 and 4EB-P1, independent of AKT.Citation104 Furthermore, overexpression of hVps34 activates S6K1 in the absence of insulin stimulation, and conversely hVps34 knockdown blocks amino acid stimulation of S6K.Citation50 In the endosome, hVps34 is able to produce PI(3)P, thereby recruiting proteins containing PI(3)P-binding domains, such as Fab1/metallo-dependent hydrolase (YOTB)/2K632.12/Vac1/early endosomal antigen 1 (EEA1) and PX domains, many of which are involved in vesicular trafficking and receptor sorting, as discussed in the Class III PI3-K section earlier. Indeed SGK3 has shown to be involved in receptor sorting at the endosome through regulating the degradation of the E3 ligase atrophin-1 interacting protein 4 (AIP4), important for degradation of the chemokine (C-X-C motif) receptor 4 (CXCR4).Citation105 Thus, while it is plausible that SGK3 may also play a role in mediating hVps34-dependent regulation of protein synthesis via mTOR/S6K1, further studies are required to elucidate this connection.
The substrate specificities for the SGK family have been determined through a panel of synthetic peptides, and demonstrate that they preferentially phosphorylate serine and threonine residues within the Arg-Xaa-Arg-Xaa-Xaa-Ser/Thr-φ motifs, (where Xaa stands for any amino acid),Citation101,Citation106–Citation108 similar to the substrate specificity of the AKT familyCitation96 Characterization of SGK3 substrate specificity has shown that it tolerates the presence of lysine instead of arginine at position n-3.Citation96 This difference is consistent with the ability of SGK3 to target substrates such as AIP4Citation105 and flightless-I (FLI-I),Citation109 which are not SGK1 or SGK2 substrates.
A more comprehensive investigation into the role of SGK3 has been achieved through the generation of various sgk gene knockout mice. Characterization of sgk3−/− mice demonstrated a distinct defect in hair follicle morphogenesis, producing a wavy hair phenotype. Further analysis revealed a defect in proliferation and nuclear accumulation of β-catenin in hair-bulb keratinocytes; however, these mice exhibited normal sodium and glucose handling.Citation110,Citation111 Interestingly, a profound proliferation defect has also been reported in pik3cadel/del embryos, which show to die between E9.5 and E10.5.Citation112 A double sgk1−/−/sgk3−/− mouse has also been generated and exhibited the combined phenotype of sgk1−/− and sgk3−/− mice, displaying a wavy hair phenotype and impairment of renal Na+ retention on a low-salt diet.Citation113 These studies using both single and double knockout animals have assisted in determining possible functional redundancies within the SGK family, with both sgk1−/− and sgk3−/− single knockout mice exhibiting quite different phenotypes.
The combined knockout of both sgk1 and sgk3 did not produce a more severe phenotype, suggesting that these two isoforms most likely do not compensate for each other. However, it is possible that the phenotype of the sgk1−/−/sgk3−/− mouse is not more severe as SGK2 may be able to compensate and maintain some level of homeostasis, despite no detectable increase of SGK2 transcript levels in these mice.Citation113 Characterization of an akt2−/−/sgk3−/− mouse found that the defect in hair growth is markedly worse in the double knockout mice than in sgk3−/− mice onlyCitation114 and that they have a markedly greater impairment of glucose homeostasis than Akt2−/− mice.Citation115 Akt2−/− mice also displayed insulin resistance, hyperinsulinemia and increased β-cell proliferation and mass.Citation116 These studies demonstrate that these proteins have both unique and common cellular functions, and in some cases work in parallel to augment the effect.
SGK3 as a key effector of PI3-K signaling
The dysregulation of many SGK3 downstream targets has been associated with important processes such as cell proliferation, growth, survival, and migration, all of which contribute to malignant transformation, as illustrated in . Furthermore, while SGK3 and AKT kinases exhibit very similar substrate specificities, they can also target distinct residues on individual substrates that affect these processes. For example, phosphorylation of FOXO3a, a member of the forkhead transcription factor family involved in the induction of cell cycle arrest and apoptosis, is phosphorylated by both AKT and SGK3 on different sites, and this results in a synergistic response.Citation90,Citation110,Citation117 This example, in addition to the evidence demonstrating clear differences in cellular localization between these kinase families, indicates the potential for SGK and AKT to have complementary roles as downstream effectors of PI3-K. Additionally, the akt2−/−/sgk3−/− double knockout studies show a level of functional redundancy between SGK3 and AKT2, indicating that these kinases may be able to compensate for each other where required. Further studies using additional akt and sgk3 double knockout models will assist in further delineating similarities between these kinase families.
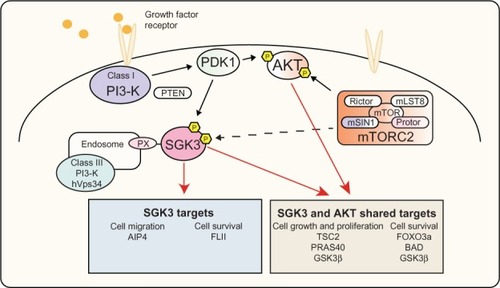
Figure 3 PI3-K signaling via SGK3 and AKT.
Notes: Activation of PI3-K by growth factor receptors leads to phosphorylation of PDK1, subsequently leading to phosphorylation and activation of AKT and SGK3. Following activation these kinases have shown to regulate TSC2 and PRAS40, leading to activation of mTORC1, an important node in signaling to protein synthesis and cell growth. In addition, AKT and SGK3 regulate FOXO3a, BAD, and GSK3β, allowing regulation of cell survival. SGK3 is also able to regulate AIP4 and FLI-I, affecting cell migration and cell survival, respectively.
Abbreviations: AIP4, atrophin-1 interacting protein 4; AKT, v-akt murine thymoma viral oncogene homolog; BAD, Bcl-2 associated death promoter; FLI-I, flightless-I; FOXO3a, forkhead transcription factor 3a; GSK3β, glycogen synthase kinase β; hVps34, class III PI3-K human vacuolar sorting protein 34; mTOR, mammalian target of rapamycin; mTORC1, mammalian target of rapamycin complex 1; mammalian target of rapamycin complex 2; PDK1, 3-phosphoinositide-dependent kinase 1; PI3-K, phosphoinositide 3-kinase; PRAS40, proline-rich AKT substrate of 40 kDa; PX, phox homology; SGK3, serum and glucocorticoid inducible kinase 3; TSC2, tuberous sclerosis factor 2; PTEN, phosphatase and tensin homolog; mLST8, MTOR associated protein LST8 homolog; mSIN1, mitogen-activated protein kinase associated protein 1.
Hallmark of cancer – cell proliferation
SGK3 potentially plays an important role in cell proliferation, through its ability to indirectly regulate the CDK inhibitor p27Kip1 via modulation of forkhead transcription factors. The regulation of FOXO3a by SGK3 occurs via SGK-dependent phosphorylation of FOXO3a at multiple sites, and ultimately prevents FOXO3a from localizing to the nucleus to affect its targets. In addition to transcriptional regulation of p27Kip1, the FOXO proteins are also involved in the regulation of other cell cycle machinery, including CDK4, cyclin D1, and retinoblastoma members p107 and p130;Citation118 thus, SGK3 regulation of FOXO3a is likely to impact cell cycle regulation at multiple levels. SGK3 also modulates GSK3β. GSK3β is involved in the regulation of numerous physiological processes, including the phosphorylation of cyclin D1, important in cell cycle transition. Following phosphorylation by GSK3β, cyclin D1 is marked for degradation by the proteasome,Citation11 and similar to AKT, SGK3 can phosphorylate and inactivate GSK3β, allowing cyclin D1 to continue its role in the cell cycle.Citation103,Citation119
Hallmark of cancer – cell growth
The AKT kinases regulate cell growth at multiple levels; the best characterized pathway is via signaling through mTORC1Citation75,Citation120,Citation121 by its phosphorylation and inhibition of TSC2 and PRAS40. While few studies have definitively demonstrated a role for SGK3 in the control of cell growth, recent studies in our laboratory have shown a role for SGK3 in growth signaling through increasing phosphorylated TSC2, PRAS40, ribosomal protein S6 (rpS6), and 4EB-P1 in normal cell physiology and malignant transformation.Citation18
Hallmark of cancer – cell survival
There have been a number of studies highlighting the role of SGK3 in cell survival. In addition to mediating IL-3-dependent survival of 32D and BAF3 hematopoietic cells,Citation90 SGK3 as well as SGK1 negatively regulate activity of the proapoptotic FOXO transcription factor FOXO3a, and can increase the level of BAD and thus attenuate cell death via B-cell CLL/lymphoma 2 (BCL-2).Citation90 SGK3 is also involved in cell survival signaling in estrogen receptor–positive breast cancer cells,Citation92 potentially via FLI-I a downstream target of SGK3,Citation109 which acts as a coactivator for the estrogen receptor, enhancing receptor activity, and promoting proliferation and survival. In addition, SGK3 is transcriptionally regulated by estrogen receptor;Citation92 thus, a positive feedback loop between SGK3 and the estrogen receptor potentially exits, which may play an important role in estrogen signaling in estrogen receptor–positive breast cancer, highlighting a crucial role for SGK3 in cell survival signaling.
Hallmark of cancer – cell migration
There are a limited number of studies that have addressed the role of SGK3 in cell migration. The phosphorylation and subsequent inactivation of GSK3β by SGK3 has been implicated with alteration of β-catenin dynamics, leading to the formation of adherens junctions and tight junction sealing in mammary epithelial cells,Citation122 raising the possibility that SGK3 may be involved in cell polarity and migration. Further, characterization of the sgk3−/− mouse exhibited a wavy hair phenotype, with further analysis revealing disorganization of hair follicles and cells in the outer root sheath, suggesting dysregulation of cell polarity.Citation110,Citation111 SGK3 also negatively regulates the lysosomal degradation of the CXCR4, whose signaling is strongly associated with the promotion of cell invasion, migration, and adhesion during metastasis in breast cancers.Citation123,Citation124 SGK3 was also shown to be able to colocalize, interact, and phosphorylate the E3 ubiquitin ligase AIP4 in the early endosomes, thereby specifically attenuating the ubiquitin-dependent degradation of CXCR4.Citation105 Together, these studies indicate a connection between SGK3 and cell migration and polarity; however, further studies are required to more specifically characterize the role of SGK3 in these processes.
Conclusion and future directions
The PI3-K pathway is regarded as one of the most crucial for cancer development and maintenance, with the ubiquitous nature of PI3-K pathway activation making both upstream and downstream components of the PI3-K signaling pathway attractive therapeutic targets. Currently, in clinical trials, there are around 30 small molecule and other inhibitors that target this pathway. The recent reports of functional dependency of PI3-K signaling on SGK3 in cancer highlights the ability of SGK3 to act as an alternate, AKT-independent signaling pathway capable of transducing critical cell proliferation and survival signals, and indicates that SGK3 may offer another avenue for targeted therapy. Further investigation into SGK3 signaling in both normal cell physiology and pathophysiology will require studies using inducible small interfering RNA systems, along with the development of specific small molecule inhibitors to further delineate the role of SGK3 signaling in malignant transformation. Currently, two small molecule inhibitors have been designed to target SGK1, suggesting that inhibitors for other members of this kinase family may also be in development.Citation125 Additionally, development of commercially available phospho-specific SGK3 antibodies for all key residues will be essential screening tools for both preclinical and clinical studies. Together, these studies paint an emerging picture of SGK3 as an important mediator of oncogenic signaling, and emphasize the critical importance of further studies focused on elucidating the signaling mechanisms associated with this kinase in both normal and malignant backgrounds.
Disclosure
The authors report no conflicts of interest in this work.
References
- SheppardKKinrossKMSolomonBPearsonRBPhillipsWATargeting PI3 kinase/AKT/mTOR signaling in cancerCrit Rev Oncog2012171699522471665
- De LucaAMaielloMRD’AlessioAPergamenoMNormannoNThe RAS/RAF/MEK/ERK and the PI3K/AKT signalling pathways: role in cancer pathogenesis and implications for therapeutic approachesExpert Opin Ther Targets201216Suppl 2S17S2722443084
- VanhaesebroeckBStephensLHawkinsPPI3K signalling: the path to discovery and understandingNat Rev Mol Cell Biol201213319520322358332
- EngelmanJATargeting PI3K signalling in cancer: opportunities, challenges and limitationsNat Rev Cancer20099855056219629070
- ManningBDCantleyLCAKT/PKB signaling: navigating downstreamCell29200712971261127417604717
- ShawRJBardeesyNManningBDThe LKB1 tumor suppressor negatively regulates mTOR signalingCancer Cell200461919915261145
- WandziochEEdlingCEPalmerRHCarlssonLHallbergBActivation of the MAP kinase pathway by c-Kit is PI-3 kinase dependent in hematopoietic progenitor/stem cell linesBlood20041041515714996702
- MaLChenZErdjument-BromageHTempstPPandolfiPPPhosphorylation and functional inactivation of TSC2 by Erk implications for tuberous sclerosis and cancer pathogenesisCell2005121217919315851026
- SolomonBPearsonRBClass IA phosphatidylinositol 3-kinase signaling in non-small cell lung cancerJ Thorac Oncol20094778779119550242
- LuYWangHMillsGBTargeting PI3K-AKT pathway for cancer therapyRev Clin Exp Hematol20037220522814763163
- VivancoISawyersCLThe phosphatidylinositol 3-Kinase AKT pathway in human cancerNat Rev Cancer20022748950112094235
- MillsGBLuYFangXThe role of genetic abnormalities of PTEN and the phosphatidylinositol 3-kinase pathway in breast and ovarian tumorigenesis, prognosis, and therapySemin Oncol2001285 Suppl 1612514111706404
- GagliardiPAdi BlasioLOrsoF3-phosphoinositide-dependent kinase 1 controls breast tumor growth in a kinase-dependent but Akt-independent mannerNeoplasia201214871973122952425
- KinrossKMMontgomeryKGKleinschmidtMAn activating Pik3ca mutation coupled with Pten loss is sufficient to initiate ovarian tumorigenesis in miceJ Clin Invest2012122255355722214849
- Stemke-HaleKGonzalez-AnguloAMLluchAAn integrative genomic and proteomic analysis of PIK3CA, PTEN, and AKT mutations in breast cancerCancer Res200868156084609118676830
- ChouMMHouWJohnsonJRegulation of protein kinase C zeta by PI 3-kinase and PDK-1Curr Biol1998819106910779768361
- MizunoHNishidaEThe ERK MAP kinase pathway mediates induction of SGK (serum- and glucocorticoid-inducible kinase) by growth factorsGenes Cells20016326126811260269
- BruhnMAPearsonRBHannanRDSheppardKESecond AKT: the rise of SGK in cancer signallingGrowth Factors201028639440820919962
- TessierMWoodgettJRSerum and glucocorticoid-regulated protein kinases: variations on a themeJ Cell Biochem20069861391140716619268
- LiuMChenLLChanTHSerum and glucocorticoid kinase 3 at 8q13.1 promotes cell proliferation and survival in hepatocellular carcinomaHepatology20125561754176522262416
- VasudevanKMBarbieDADaviesMAAKT-independent signaling downstream of oncogenic PIK3CA mutations in human cancerCancer Cell2009161213219573809
- VanhaesebroeckBGuillermet-GuibertJGrauperaMBilangesBThe emerging mechanisms of isoform-specific PI3K signallingNat Rev Mol Cell Biol201011532934120379207
- FrumanDARegulatory subunits of class IA PI3KCurr Top Microbiol Immunol201034622524420563711
- VogtPKHartJRGymnopoulosMPhosphatidylinositol 3-kinase: the oncoproteinCurr Top Microbiol Immunol20103477910420582532
- JiaSRobertsTMZhaoJJShould individual PI3 kinase isoforms be targeted in cancer?Curr Opin Cell Biol200921219920819200708
- WongKKEngelmanJACantleyLCTargeting the PI3K signaling pathway in cancerCurr Opin Genet Dev2010201879020006486
- SajiMRingelMDThe PI3K-Akt-mTOR pathway in initiation and progression of thyroid tumorsMol Cell Endocrinol20103211202819897009
- ShawRJLKB1 and AMP-activated protein kinase control of mTOR signalling and growthActa Physiol (Oxf)20091961658019245654
- CantleyLCThe phosphoinositide 3-kinase pathwayScience200229655731655165712040186
- MarkmanBDienstmannRTaberneroJTargeting the PI3K/Akt/mTOR pathway – beyond rapalogsOncotarget20101753054321317449
- HuPMargolisBSkolnikEYLammersRUllrichASchlessingerJInteraction of phosphatidylinositol 3-kinase-associated p85 with epidermal growth factor and platelet-derived growth factor receptorsMol Cell Biol19921239819901372091
- McGladeCJEllisCReedijkMSH2 domains of the p85 alpha subunit of phosphatidylinositol 3-kinase regulate binding to growth factor receptorsMol Cell Biol19921239919971372092
- ZhuGDeckerSJSaltielARDirect analysis of the binding of Srchomology 2 domains of phospholipase C to the activated epidermal growth factor receptorProc Natl Acad Sci U S A19928920955995631384057
- SamuelsYVelculescuVEOncogenic mutations of PIK3CA in human cancersCell Cycle20043101221122415467468
- SamuelsYWangZBardelliAHigh frequency of mutations of the PIK3CA gene in human cancersScience2004304567055415016963
- MarkmanBAtzoriFPerez-GarciaJTaberneroJBaselgaJStatus of PI3K inhibition and biomarker development in cancer therapeuticsAnn Oncol201021468369119713247
- ZhaoJJChengHJiaSThe p110alpha isoform of PI3K is essential for proper growth factor signaling and oncogenic transformationProc Natl Acad Sci U S A200610344162961630017060635
- MaoCYangZYHuXFChenQTangJLPIK3CA exon 20 mutations as a potential biomarker for resistance to anti-EGFR monoclonal antibodies in KRAS wild-type metastatic colorectal cancer: a systematic review and meta-analysisAnn Oncol20122361518152522039088
- SamuelsYWaldmanTOncogenic mutations of PIK3CA in human cancersCurr Top Microbiol Immunol2010347214120535651
- PhilpAJCampbellIGLeetCThe phosphatidylinositol 3′-kinase p85alpha gene is an oncogene in human ovarian and colon tumorsCancer Res200161207426742911606375
- IsakoffSJEngelmanJAIrieHYBreast cancer-associated PIK3CA mutations are oncogenic in mammary epithelial cellsCancer Res20056523109921100016322248
- SamuelsYDiazLAJrSchmidt-KittlerOMutant PIK3CA promotes cell growth and invasion of human cancer cellsCancer Cell20057656157315950905
- SamuelsYEricsonKOncogenic PI3K and its role in cancerCurr Opin Oncol2006181778216357568
- BaderAGKangSZhaoLVogtPKOncogenic PI3K deregulates transcription and translationNat Rev Cancer200551292192916341083
- ZhaoJJLiuZWangLShinELodaMFRobertsTMThe oncogenic properties of mutant p110alpha and p110beta phosphatidylinositol 3-kinases in human mammary epithelial cellsProc Natl Acad Sci U S A200510251184431844816339315
- GymnopoulosMElsligerMAVogtPKRare cancer-specific mutations in PIK3CA show gain of functionProc Natl Acad Sci U S A2007104135569557417376864
- VogtPKGymnopoulosMHartJRPI 3-kinase and cancer: changing accentsCurr Opin Genet Dev2009191121719185485
- ShekarSCWuHFuZMechanism of constitutive phosphoinositide 3-kinase activation by oncogenic mutants of the p85 regulatory subunitJ Biol Chem200528030278502785515932879
- CastellanoEDownwardJRAS interaction with PI3K: more than just another effector pathwayGenes Cancer20112326127421779497
- BackerJMThe regulation and function of Class III PI3Ks: novel roles for Vps34Biochem J2008410111718215151
- YanYBackerJMRegulation of class III (Vps34) PI3KsBiochem Soc Trans200735Pt 223924117371248
- YanYFlinnRJWuHSchnurRSBackerJMhVps15, but not Ca2+/CaM, is required for the activity and regulation of hVps34 in mammalian cellsBiochem J2009417374775518957027
- ThoresenSBPedersenNMLiestolKStenmarkHA phosphatidylinositol 3-kinase class III sub-complex containing VPS15, VPS34, Beclin 1, UVRAG and BIF-1 regulates cytokinesis and degradative endocytic trafficExp Cell Res2010316203368337820643123
- NobukuniTJoaquinMRoccioMAmino acids mediate mTOR/raptor signaling through activation of class 3 phosphatidylinositol 3OH-kinaseProc Natl Acad Sci U S A200510240142381424316176982
- ByfieldMPMurrayJTBackerJMhVps34 is a nutrient-regulated lipid kinase required for activation of p70 S6 kinaseJ Biol Chem200528038330763308216049009
- GulatiPGaspersLDDannSGAmino acids activate mTOR complex 1 via Ca2+/CaM signaling to hVps34Cell Metab20087545646518460336
- LaplanteMSabatiniDMmTOR signaling in growth control and diseaseCell2012149227429322500797
- KimEGoraksha-HicksPLiLNeufeldTPGuanKLRegulation of TORC1 by Rag GTPases in nutrient responseNat Cell Biol200810893594518604198
- SancakYPetersonTRShaulYDThe Rag GTPases bind raptor and mediate amino acid signaling to mTORC1Science200832058821496150118497260
- FindlayGMYanLProcterJMieuletVLambRFA MAP4 kinase related to Ste20 is a nutrient-sensitive regulator of mTOR signallingBiochem J20074031132017253963
- KimSKimSFMaagDAmino acid signaling to mTOR mediated by inositol polyphosphate multikinaseCell Metab201113221522121284988
- HannanKMBrandenburgerYJenkinsAmTOR-dependent regulation of ribosomal gene transcription requires S6K1 and is mediated by phosphorylation of the carboxy-terminal activation domain of the nucleolar transcription factor UBFMol Cell Biol200323238862887714612424
- YangXYangCFarbermanAThe mammalian target of rapamycin-signaling pathway in regulating metabolism and growthJ Anim Sci200886Suppl 14E36E5017998426
- WangXProudCGThe mTOR pathway in the control of protein synthesisPhysiology (Bethesda)20062136236916990457
- JamesMJZomerdijkJCPhosphatidylinositol 3-kinase and mTOR signaling pathways regulate RNA polymerase I transcription in response to IGF-1 and nutrientsJ Biol Chem2004279108911891814688273
- CorradettiMNGuanKLUpstream of the mammalian target of rapamycin: do all roads pass through mTOR?Oncogene200625486347636017041621
- HemmingsBARestucciaDFPI3K-PKB/Akt pathwayCold Spring Harb Perspect Biol201249a01118922952397
- SancakYThoreenCCPetersonTRPRAS40 is an insulinregulated inhibitor of the mTORC1 protein kinaseMol Cell200725690391517386266
- Vander HaarELeeSIBandhakaviSGriffinTJKimDHInsulin signalling to mTOR mediated by the Akt/PKB substrate PRAS40Nat Cell Biol20079331632317277771
- LiangJSlingerlandJMMultiple roles of the PI3K/PKB (Akt) pathway in cell cycle progressionCell Cycle20032433934512851486
- BrunetABonniAZigmondMJAkt promotes cell survival by phosphorylating and inhibiting a forkhead transcription factorCell199996685786810102273
- DattaSRDudekHTaoXAkt phosphorylation of BAD couples survival signals to the cell-intrinsic death machineryCell19979122312419346240
- MayoLDDonnerDBA phosphatidylinositol 3-kinase/Akt pathway promotes translocation of Mdm2 from the cytoplasm to the nucleusProc Natl Acad Sci U S A20019820115981160311504915
- RomashkovaJAMakarovSSNF-kappaB is a target of AKT in anti-apoptotic PDGF signallingNature19994016748869010485711
- ChanJCHannanKMRiddellKAKT promotes rRNA synthesis and cooperates with c-MYC to stimulate ribosome biogenesis in cancerSci Signal20114188ra5621878679
- StaalSPMolecular cloning of the akt oncogene and its human homologues AKT1 and AKT2: amplification of AKT1 in a primary human gastric adenocarcinomaProc Natl Acad Sci U S A19878414503450373037531
- BellacosaAde FeoDGodwinAKMolecular alterations of the AKT2 oncogene in ovarian and breast carcinomasInt J Cancer19956442802857657393
- HennessyBTSmithDLRamPTLuYMillsGBExploiting the PI3K/AKT pathway for cancer drug discoveryNat Rev Drug Discov2005412988100416341064
- CarptenJDFaberALHornCA transforming mutation in the pleckstrin homology domain of AKT1 in cancerNature2007448715243944417611497
- YuanZQSunMFeldmanRIFrequent activation of AKT2 and induction of apoptosis by inhibition of phosphoinositide-3-OH kinase/Akt pathway in human ovarian cancerOncogene200019192324233010822383
- StahlJMSharmaACheungMDeregulated Akt3 activity promotes development of malignant melanomaCancer Res200464197002701015466193
- CristianoBEChanJCHannanKMA specific role for AKT3 in the genesis of ovarian cancer through modulation of G(2)-M phase transitionCancer Res20066624117181172517178867
- Cancer Genome Atlas Research NetworkIntegrated genomic analyses of ovarian carcinomaNature2011474735360961521720365
- WuRHendrix-LucasNKuickRMouse model of human ovarian endometrioid adenocarcinoma based on somatic defects in the Wnt/beta-catenin and PI3K/Pten signaling pathwaysCancer Cell200711432133317418409
- ZhangWHainesBBEffersonCEvidence of mTOR Activation by an AKT-independent mechanism provides support for the combined treatment of PTEN-deficient prostate tumors with mTOR and AKT inhibitorsTransl Oncol20125642242923323157
- XuJWanMHeQSGK3 is associated with estrogen receptor expression in breast cancerBreast Cancer Res Treat2012134253154122576469
- CreightonCJA gene transcription signature of the Akt/mTOR pathway in clinical breast tumorsOncogene200726324648465517213801
- SommerEMDryHCrossDGuichardSDaviesBRAlessiDRElevated SGK1 predicts resistance of breast cancer cells to Akt inhibitorsBiochem J2013452349950823581296
- De MarcoCRinaldoNBruniPMultiple genetic alterations within the PI3K pathway are responsible for AKT activation in patients with ovarian carcinomaPLoS One201382e5536223408974
- LiuDYangXSongyangZIdentification of CISK, a new member of the SGK kinase family that promotes IL-3-dependent survivalCurr Biol200010191233123611050396
- DaiFYuLHeHCloning and mapping of a novel human serum/glucocorticoid regulated kinase-like gene, SGKL, to chromosome 8q12.3-q13.1Genomics1999621959710585774
- WangYZhouDPhungSMasriSSmithDChenSSGK3 is an estrogen-inducible kinase promoting estrogen-mediated survival of breast cancer cellsMol Endocrinol2011251728221084382
- EllsonCDAndrewsSStephensLRHawkinsPTThe PX domain: a new phosphoinositide-binding moduleJ Cell Sci2002115Pt 61099110511884510
- VirbasiusJVSongXPomerleauDPZhanYZhouGWCzechMPActivation of the Akt-related cytokine-independent survival kinase requires interaction of its phox domain with endosomal phosphatidylinositol 3-phosphateProc Natl Acad Sci U S A20019823129081291311606732
- XuJLiuDGillGSongyangZRegulation of cytokine-independent survival kinase (CISK) by the Phox homology domain and phosphoinositidesJ Cell Biol2001154469970511514587
- KobayashiTDeakMMorriceNCohenPCharacterization of the structure and regulation of two novel isoforms of serum- and glucocorticoid-induced protein kinaseBiochem J1999344Pt 118919710548550
- FirestoneGLGiampaoloJRO’KeeffeBAStimulus-dependent regulation of serum and glucocorticoid inducible protein kinase (SGK) transcription, subcellular localization and enzymatic activityCell Physiol Biochem200313111212649597
- Garcia-MartinezJMAlessiDRmTOR complex 2 (mTORC2) controls hydrophobic motif phosphorylation and activation of serum-and glucocorticoid-induced protein kinase 1 (SGK1)Biochem J2008416337538518925875
- LuMWangJJonesKTmTOR complex-2 activates ENaC by phosphorylating SGK1J Am Soc Nephrol201021581181820338997
- LyoDXuLFosterDAPhospholipase D stabilizes HDM2 through an mTORC2/SGK1 pathwayBiochem Biophys Res Commun2010396256256520438709
- ParkJLeongMLBusePMaiyarACFirestoneGLHemmingsBASerum and glucocorticoid-inducible kinase (SGK) is a target of the PI 3-kinase-stimulated signaling pathwayEMBO J199918113024303310357815
- TessierMWoodgettJRRole of the Phox homology domain and phosphorylation in activation of serum and glucocorticoid-regulated kinase-3J Biol Chem200628133239782398916790420
- KobayashiTCohenPActivation of serum- and glucocorticoid-regulated protein kinase by agonists that activate phosphatidylinositide 3-kinase is mediated by 3-phosphoinositide-dependent protein kinase-1 (PDK1) and PDK2Biochem J1999339Pt 231932810191262
- Sanchez CanedoCDemeulderBGinionAActivation of the cardiac mTOR/p70(S6K) pathway by leucine requires PDK1 and correlates with PRAS40 phosphorylationAm J Physiol Endocrinol Metab20102984E761E76920051528
- SlagsvoldTMarcheseABrechAStenmarkHCISK attenuates degradation of the chemokine receptor CXCR4 via the ubiquitin ligase AIP4EMBO J200625163738374916888620
- FrodinMAntalTLDummlerBAA phosphoserine/threonine-binding pocket in AGC kinases and PDK1 mediates activation by hydrophobic motif phosphorylationEMBO J200221205396540712374740
- CollinsBJDeakMArthurJSArmitLJAlessiDRIn vivo role of the PIF-binding docking site of PDK1 defined by knock-in mutationEMBO J200322164202421112912918
- BiondiRMKielochACurrieRADeakMAlessiDRThe PIF-binding pocket in PDK1 is essential for activation of S6K and SGK, but not PKBEMBO J200120164380439011500365
- XuJLiaoLQinJLiuDSongyangZIdentification of flightless-I as a substrate of the cytokine-independent survival kinase CISKJ Biol Chem200928421143771438519293151
- McCormickJAFengYDawsonKTargeted disruption of the protein kinase SGK3/CISK impairs postnatal hair follicle developmentMol Biol Cell20041594278428815240817
- AlonsoLOkadaHPasolliHASgk3 links growth factor signaling to maintenance of progenitor cells in the hair follicleJ Cell Biol2005170455957016103225
- BiLOkabeIBernardDJWynshaw-BorisANussbaumRLProliferative defect and embryonic lethality in mice homozygous for a deletion in the p110alpha subunit of phosphoinositide 3-kinaseJ Biol Chem199927416109631096810196176
- GrahammerFArtuncFSandulacheDRenal function of gene-targeted mice lacking both SGK1 and SGK3Am J Physiol Regul Integr Comp Physiol20062904R945R95016537821
- MauroTMMcCormickJAWangJAkt2 and SGK3 are both determinants of postnatal hair follicle developmentFASEB J20092393193320219433625
- YaoLJMcCormickJAWangJNovel role for SGK3 in glucose homeostasis revealed in SGK3/Akt2 double-null miceMol Endocrinol201125122106211821980074
- ChoHMuJKimJKInsulin resistance and a diabetes mellitus-like syndrome in mice lacking the protein kinase Akt2 (PKB beta)Science200129255221728173111387480
- BrunetAParkJTranHHuLSHemmingsBAGreenbergMEProtein kinase SGK mediates survival signals by phosphorylating the forkhead transcription factor FKHRL1 (FOXO3a)Mol Cell Biol200121395296511154281
- HoKKMyattSSLamEWMany forks in the path: cycling with FoxOOncogene200827162300231118391972
- DaiFYuLHeHHuman serum and glucocorticoid-inducible kinase-like kinase (SGKL) phosphorylates glycogen syntheses kinase 3 beta (GSK-3beta) at serine-9 through direct interactionBiochem Biophys Res Commun200229341191119612054501
- HannanKMSanijEHeinNHannanRDPearsonRBSignaling to the ribosome in cancer – it is more than just mTORC1IUBMB Life2011632798521360636
- EngelmanJALuoJCantleyLCThe evolution of phosphatidylinositol 3-kinases as regulators of growth and metabolismNat Rev Genet20067860661916847462
- FailorKLDesyatnikovYFingerLAFirestoneGLGlucocorticoid-induced degradation of glycogen synthase kinase-3 protein is triggered by serum- and glucocorticoid-induced protein kinase and Akt signaling and controls beta-catenin dynamics and tight junction formation in mammary epithelial tumor cellsMol Endocrinol200721102403241517595317
- GassmannPHaierJSchluterKCXCR4 regulates the early extravasation of metastatic tumor cells in vivoNeoplasia200911765166119568410
- MullerAHomeyBSotoHInvolvement of chemokine receptors in breast cancer metastasisNature20014106824505611242036
- LangFVoelklJTherapeutic potential of serum and glucocorticoid inducible kinase inhibitionExpert Opin Investig Drugs2013226701714