
Abstract
Aberrant canonical Wnt–β-catenin signaling has been reported in multiple sclerosis (MS), although the results are controversial. The present study aimed to examine the role of the Wnt–β-catenin pathway in experimental MS and also to test moringin (4-[α-L-rhamnopyranosyloxy]-benzyl isothiocyanate), resulting from exogenous myrosinase hydrolysis of the natural phytochemical glucomoringin 4(α-L-rhamnosyloxy)-benzyl glucosinolate as a modulator of neuroinflammation via the β-catenin–PPARγ axis. Experimental autoimmune encephalomyelitis (EAE), the most common model of MS, was induced in C57BL/6 mice by immunization with MOG35–55. Released moringin (10 mg/kg glucomoringin +5 μL myrosinase/mouse) was administered daily for 1 week before EAE induction and continued until mice were killed on day 28 after EAE induction. Our results clearly showed that the Wnt–β-catenin pathway was downregulated in the EAE model, whereas moringin pretreatment was able to avert this. Moringin pretreatment normalizes the aberrant Wnt–β-catenin pathway, resulting in GSK3β inhibition and β-catenin upregulation, which regulates T-cell activation (CD4 and FoxP3), suppresses the main inflammatory mediators (IL-1β, IL-6, and COX2), through activation of PPARγ. In addition, moringin attenuates apoptosis by reducing the expression of the Fas ligand and cleaved caspase 9, and in parallel increases antioxidant Nrf2 expression in EAE mice. Taken together, our results provide an interesting discovery in identifying moringin as a modulator of the Wnt–β-catenin signaling cascade and as a new potential therapeutic target for MS treatment.
Introduction
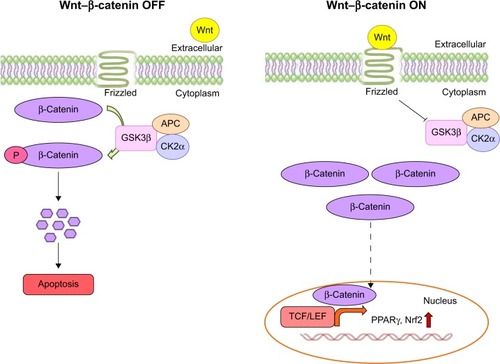
The central logic of the Wnt–β-catenin-dependent or Wnt canonical pathway has been revealed in the past two decades. The Wnt-signaling pathway regulates many biological events occurring in the developmental and adult phases of all animals.Citation1 Wnts are a family of secreted signaling proteins, which activate either β-catenin-dependent or -independent intracellular pathways by binding to the seven-pass transmembrane receptors of the Frizzled family.Citation2 Generally, the Wnt1 class acts via the canonical pathway, while the Wnt5 class acts via the noncanonical pathway.Citation3,Citation4 The key role of the canonical Wnt pathway is the stabilization of β-catenin present in the cytoplasm. When the Wnt ligand is absent, β-catenin binds with the adenomatous polyposis coli–Axin complex, which promotes β-catenin phosphorylation via the GSK3β enzyme. Then, phosphorylated β-catenin binds with the ubiquitin complex, resulting in degradation.Citation3,Citation5 Wnt signaling prevents GSK3β activity, and thus increases the amount of β-catenin, which translocates into the nucleus and associates with TCF/LEF transcription factors, leading to the regulation of Wnt target genes.Citation3
Wnt–β-catenin signaling is involved in the development of sensory and motor neurons present in the brain and spinal cord via regulation of all facets of neuronal activities, including differentiation, proliferation/senescence, and survival/apoptosis.Citation6–Citation8 Moreover, Wnt-signaling pathway aberrations are associated with the adulthood diseases of the central nervous system (CNS), which points to its critical role in the development of the mature CNS.Citation1,Citation8 Growing evidence shows the importance of the Wnt–β-catenin pathway to stabilize neuronal cell survival and death in neurodegenerative diseases, such as Alzheimer’s disease and Parkinson’s disease.Citation1,Citation9–Citation11 In addition, the Wnt–β-catenin pathway seems to be involved in the pathogenesis and modulation of chronic pain in experimental autoimmune encephalomyelitis (EAE) mice,Citation12 the most commonly used model for multiple sclerosis (MS). However, the role of the Wnt canonical pathway in MS is still unclear.
MS is a chronic inflammatory disease caused by an induction of autoreactive immune responses effected from T and B lymphocytes, which results in the demyelination of the myelin sheath around neurons in the CNS.Citation13,Citation14 It is noteworthy to emphasize that the present-day treatments for MS provide palliative relief, but do not cure the disease. Moreover, these treatments trigger many side effects that hinder their application for a prolonged period.Citation15 Therefore, new drugs that may act on the underlying etiology with no side effects are urgently required to treat MS.
In the last few decades, PPARγ, an important target for diabetes treatment,Citation16 has aroused great interest for its therapeutic role in brain disorders.Citation17 In addition, since PPARγ is linked with the Wnt–β-catenin pathway,Citation18,Citation19 identifying novel PPARγ activators through Wnt–β-catenin pathway regulation has become a promising therapeutic approach for brain diseases.
In this regard, we investigated moringin (4-[α-L-rhamnopyranosyloxy]-benzyl isothiocyanate) as a modulator of neuroinflammation via the β-catenin–PPARγ axis in an experimental model of MS. Moringin is released from the precursor 4-(α-L-rhamnosyloxy)-benzyl glucosinolate (glucomoringin [GMG]), found predominantly in the seeds of Moringa oleifera, the most widely distributed plant of the Moringaceae family, by myrosinase (β-thioglucoside glucohydrolase; EC 3.2.1.147)-catalyzed hydrolysis.
Moringin has recently been characterized and proven to possess a broad range of biological activities, including protective effects against neurodegenerative disorders and particularly MS.Citation20,Citation21 In the present study, we evaluated the regulatory role of moringin in the Wnt canonical pathway and PPARγ activation in a mouse EAE model.
Materials and methods
Isolation and purification of glucomoringin and myrosinase
GMG was isolated from M. oleifera seeds (PKM2 cake powder; Indena India Pvt Ltd, Bangalore, India) in two sequential steps – anion-exchange and size-exclusion chromatography – according to previously reported methods.Citation22,Citation23 The PKM2 cake powder was treated with boiling 70% ethanol, in order to quickly deactivate the endogenous enzyme myrosinase. GMG was extracted at medium speed using an Ultra-Turrax homogenizer for 15 minutes, and the resulting homogenate was centrifuged at 17,700× g for 30 minutes. The isolation of GMG from the extract was carried out by one-step anion-exchange chromatography.Citation24 The extract was loaded on a DEAE Sephadex A-25 (GE Healthcare UK Ltd, Little Chalfont, UK) anion-exchange column (150×26 mm) conditioned with 25 mM acetate buffer (pH 5.6). After being washed with 1 L of distilled water, GMG was eluted with 500 mL of aqueous K2SO4, 0.2 M. The eluate was concentrated to dryness using a rotary evaporator at 60°C–70°C under reduced pressure. Three subsequent extractions were carried out with 70–100 mL of boiling methanol. The alcoholic extract was then filtered and concentrated to 15%–20% of the initial volume. The solution was warmed and slowly added dropwise to 200 mL of absolute ethanol that had been previously cooled to −20°C. This led to the precipitation of a white powder. After centrifugation, the solid GMG (as potassium salt) was dried and sealed under vacuum to prevent moisture uptake by the highly hygroscopic solid. The purity of GMG was further improved by gel filtration performed using an XK 26/100 column packed with Sephadex G-10 (GE Healthcare UK Ltd) connected to a fast protein liquid chromatography (LC) system (AKTA; GE Healthcare UK Ltd).Citation25
The mobile phase was water at a flow rate of 2 mL/min, and the eluate absorbance was monitored at 254 nm. After the void volume was discarded, 5 mL fractions were collected and analyzed by high-performance LC (HPLC), and those containing GMG were pooled and freeze-dried.Citation26 The purity was assayed by HPLC analysis of the desulfo-derivative according to the ISO 9167-1 method,Citation27 yielding approximately 99% based on peak area value and more than 95% on a weight basis, due to its high hygroscopic properties,Citation28 determined by a calibration curve of standard desulfo-GMG available in our laboratory. The enzyme myrosinase was isolated from white mustard (Sinapis alba) seeds according to a reported method with some modifications.Citation29
Briefly, the enzyme was extracted from white-mustard seeds with water and purified by affinity chromatography on Con A Sepharose (GE Healthcare UK Ltd). Then, the active fractions coming from affinity chromatography were pooled and dialyzed against 50 mM phosphate buffer, pH 6.5, containing 0.15 M NaCl. The dialyzed Myr solution was concentrated and loaded onto a prepacked Superdex 200 HiLoad 26/60 gel-filtration column (GE Healthcare UK Ltd) equilibrated with 50 mM phosphate buffer, pH 6.5, containing 0.15 M NaCl connected with the fast protein LC system. The active fractions were pooled and concentrated with Amicon stirred cell model 8400 using an ultrafiltration membrane at 30 kDa molecular weight cutoff (EMD Millipore, Billerica, MA, USA). The stock solution used in the present study had a specific activity of 60 U/mg of soluble protein. Enzymatic activity was 32 U/mL, and the solution was stored at 4°C in sterile saline solution at neutral pH until use. One myrosinase unit was defined as the amount of enzyme able to hydrolyze 1 μmol/min of sinigrin at pH 6.5 and 37°C.Citation30
Enzyme bioactivation of glucomoringin
GMG (1 mg/mL) was suspended in phosphate-buffered saline (PBS; pH 7.2), and enzyme bioactivation of the phytochemical compound was required for mouse treatment. The in situ action of myrosinase (20 μL/mL) for 15 minutes at 37°C allowed the delivery of moringin quickly. Moreover, the total conversion of pure GMG into moringin before animal administration was confirmed by HPLC analysis, as reported in our previous work.Citation31
Induction of experimental autoimmune encephalomyelitis
This study was carried out in strict accordance with the recommendations in the guide for the care and use of laboratory animals of the National Institutes of Health. The protocol was approved by the Ministry of Health “General Direction of animal health and veterinary drugs” and this study was approved by “Animal Welfare Bodies” of IRCCS Centro Neurolesi Bonino-Pulejo, Messina, Italy. Particularly, animal care was in compliance with Italian regulations on the protection of animals used for experimental and other scientific purposes (D.lgs 26/2014). All efforts were made during experimental procedures, to minimize animal suffering and also to reduce the number of animal used. Male C57BL/6 mice (20–25 g) were anesthetized with an anesthetic cocktail consisting of tiletamine plus xylazine (10 mL/kg intraperitoneally). EAE was induced in mice using the peptide MOG35–55 (MEVGWYRSPFSRVVHLYRNGK, peak area by HPLC ≥95%; AnaSpec Inc., Fremont, CA, USA), based on Paschalidis et al.Citation32 Mice were subcutaneously injected with 300 μL/flank of the suspension consisting of an equal volume of 300 μg MOG35–55 in PBS and complete Freund’s adjuvant consisting of 300 μg heat-killed Mycobacterium tuberculosis H37Ra (BD, Franklin Lakes, NJ, USA). After MOG35–55 injection, the animals immediately received an intraperitoneal injection of 100 μL Bordetella pertussis toxin (500 ng/100 μL; Sigma-Aldrich Co., St Louis, MO, USA) and 48 hours later. EAE induction followed a sequence of progressive degeneration with visible signs, such as tail flaccidity and loss of hind-leg movement.
Experimental design
Mice were randomly separated into the following groups (n=35 total animals):
naïve group (n=5) – mice without any injection, serving as controls.
moringin control group (n=5) – mice not subjected to EAE induction, but injected with moringin (10 mg/kg GMG +5 μL Myr/mouse), killed as controls of drug safety and tolerance.
Myr control group (n=5) – mice not subjected to EAE damage, but only injected with Myr (5 μL Myr/mouse) to evaluate possible side effects, including allergenic reactions after administration.
EAE group (n=10) – mice receiving MOG injection.
EAE + moringin (n=10) – MOG-injected mice administered with moringin (10 mg/kg GMG +5 μL myrosinase/mouse); moringin administered intraperitoneally daily for 1 week before EAE induction and continued daily after EAE induction until death.
After 28 days of EAE induction, mice were killed and spinal cord tissues collected and processed for further analyses.
Clinical disease-score evaluation
Mice showed initial signs of MS, including loss of tail tonus, hind-limb paralysis, and loss of body weight, after 14 days of EAE induction. Clinical neurological score was assessed according to a standardized scoring system:Citation33 0, no visible signs; 1, partial flaccid tail; 2, complete flaccid tail; 3, hind limb hypotonia; 4, partial hind-limb paralysis; 5, complete hind-limb paralysis; and 6, moribund or dead animal. Mice with a clinical score ≥5 were killed to avoid animal suffering. The following clinical disease parameters were assessed: incidence, day of onset of clinical signs, peak disease score, cumulative disease score (CDS), and mortality (). The first assessment of neurological disease score was measured on the day of EAE induction (day 0), and all subsequent assessments were performed every 48 hours until mice were killed. Days’ difference in the disease parameter is reported compared to the day of EAE induction (day 0). Days are reported as mean ± standard error of mean of all mice for each experimental set.
Table 1 Clinical parameters of EAE
Immunohistochemistry
Spinal cord tissues removed from the cervical region were fixed in 10% (w/v) PBS-buffered formaldehyde and embedded in paraffin. The tissues were cut into 7 μm sections. After deparaffinization, tissues were incubated with 0.3% (v/v) hydrogen peroxide in 60% (v/v) methanol for 30 minutes to terminate endogenous peroxidase activity. Then, the tissue sections were blocked in 2% (v/v) normal goat serum in PBS for 20 minutes at room temperature.
Tissue sections were incubated overnight with the following primary antibodies:
anti-IL-1β polyclonal antibody (1:50 in PBS v/v; Santa Cruz Biotechnology Inc., Dallas, TX, USA)
anti-IL-6 polyclonal antibody (1:100 in PBS v/v; Abcam, Cambridge, UK)
anti-Fas-ligand polyclonal antibody (1:100 in PBS v/v; Abcam)
anti-cleaved caspase-9 monoclonal antibody (1:200 in PBS v/v; Abcam)
anti-CD4 polyclonal antibody (1:100 in PBS v/v; Santa Cruz Biotechnology Inc.)
anti-FoxP3 monoclonal antibody (1:100 in PBS v/v; Santa Cruz Biotechnology Inc.).
To avoid endogenous cross-reactivity of biotin- or avidin-binding sites, tissue sections were blocked separately with biotin and avidin for 15 minutes. Tissue sections were rinsed with PBS and incubated with secondary antibody. Specific labeling was performed using a biotin-conjugated antirabbit IgG and avidin–biotin peroxidase complex (VectaStain; Vector Laboratories, Burlingame, CA, USA). Then, the tissue sections were stained using a DAB peroxidase-substrate kit (Vector Laboratories), followed by hematoxylin counterstaining. In addition, tissue sections were incubated with either primary or secondary antibody to assess antibody specificity. In these cases, no positive staining was observed in the tissue sections, indicating that the immunoreactions were positive in all the experiments carried out. Immunohistochemical staining was evaluated using light microscopy (Leica DM 2000 combined with Leica ICC50 HD camera), and images were acquired by Leica Application Suite version 4.2.0 software.
Western blot analysis
Spinal cord tissues were homogenized in ice-cold extraction buffer consisting of 0.32 M sucrose, 10 mM Tris–HCl (pH 7.4), 5 mM NaN3, 2 mM ethylenediaminetetraacetic acid (EDTA), 1 mM ethyleneglycoltetraacetic acid (EGTA), 50 mM NaF, 10 mM 2-mercaptoethanol, and protease-inhibitor tablets (Hoffman-La Roche Ltd, Basel, Switzerland). The homogenates were kept on ice for 15 minutes, centrifuged (1,000 g for 10 minutes at 4°C), and the supernatant removed to estimate cytosolic proteins. The pellets were resuspended in ice-cold lysis buffer consisting of 10 mM Tris–HCl (pH 7.4), 150 mM NaCl, 1 mM EDTA, 1 mM EGTA, 1% Triton X-100, and protease-inhibitor tablets. The resuspended pellets were kept on ice for 10 minutes, centrifuged (15,000× g for 30 minutes at 4°C), and the supernatant removed to estimate nuclear proteins. Supernatants were preserved at −80°C until use. The quantity of protein was calculated with protein-assay reagent (Bio-Rad Laboratories Inc., Hercules, CA, USA). Bovine serum albumin was used as the standard. Proteins (30 μg) were resolved on sodium dodecyl sulfate polyacrylamide minigels (8% or 12%) and transferred onto polyvinylidene difluoride membranes (Immobilon-P transfer membrane; EMD Millipore). After transfer, membranes were blocked with PBS containing 5% nonfat dried milk (PBS-milk (PM)) for 1 hour at room temperature, and incubated for overnight at 4°C overnight with specific antibodies: Wnt1 (1:250; Santa Cruz Biotechnology), β-catenin (1:500; Cell Signaling Technology, Danvers, MA, USA), CK2α (1:250; Santa Cruz Biotechnology Inc.), GSK3β (1:250; Santa Cruz Biotechnology Inc.), p-β-catenin (1:500; Santa Cruz Biotechnology Inc.), PPARγ (1:250; Santa Cruz Biotechnology Inc.), COX2 (1:250; Santa Cruz Biotechnology Inc.), and Nrf2 (1:250; Santa Cruz Biotechnology Inc.). Then, membranes were washed in PBS and incubated with horseradish peroxidase-conjugated antimouse, -goat, or -rabbit IgG secondary antibody (1:2,000; Santa Cruz Biotechnology Inc.) for 1 hour at room temperature. Membranes were stripped and reprobed with β-actin (1:1,000; Santa Cruz Biotechnology Inc.) to confirm uniform protein loading. Protein bands were detected by an enhanced chemiluminescence system (Luminata Western; EMD Millipore), and images were taken by ChemiDoc™ MP (Bio-Rad Laboratories Inc.) and quantified for relative expression of proteins using ImageJ software.
Statistical analysis
Statistical analysis was performed using GraphPad Prism version 6.0 (GraphPad Software, Inc., La Jolla, CA, USA). The data were statistically analyzed by one-way analysis of variance and Bonferroni post hoc tests for multiple comparisons. A P-value ≤0.05 was considered statistically significant. Data are reported as mean ± standard error of mean of n experiments.
Results
Moringin ameliorates clinical disease score
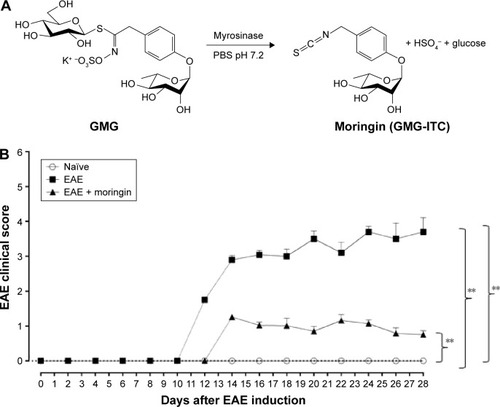
Clinical disease score was evaluated as a functional neurological parameter. EAE is a well-documented model of MS in the mouse and resembles the hallmarks of the disease, such as paralysis, loss of body weight, inflammation, demyelination in the CNS and blood–brain barrier leakage.Citation34 In order to study the role of moringin obtained by myrosinase hydrolysis () in regulating susceptibility to EAE, mice were pretreated with moringin and immunized with MOG35–55. The mice were monitored every 48 hours to assess clinical EAE signs. Compared to the EAE group, moringin pretreated mice showed a significant reduction in disease incidence and average CDS. The lower CDS in the EAE + moringin group was also reflected as a significant reduction in the severity of disease (). Indeed, EAE mice displayed a chronic-progressive clinical course, while significant reduction in clinical score was observed in EAE mice pretreated with moringin. No sign of motor deficits was noticed in naïve animals (). These results suggest that moringin reduces disease progression and increases recovery of neurological function in EAE mice.
Figure 1 Treatment with moringin ameliorates clinical score in EAE mice.
Abbreviations: EAE, experimental autoimmune encephalomyelitis; SEM, standard error of mean; GMG, glucomoringin; ITC, isothiocyanate.
Moringin regulates the Wnt–β-catenin signaling pathway in EAE development
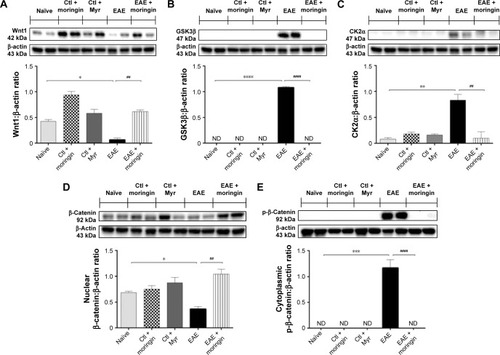
Western blot analysis was performed in order to observe the modulation of the Wnt–β-catenin signaling pathway after EAE induction in mouse spinal cord. Achieved results showed that the Wnt–β-catenin canonical pathway was downregulated in EAE mice. Lower Wnt1 expression () was found in spinal cord tissues taken from EAE mice compared to naïve and control ones, which led to enhanced expression of GSK3β (), which acts synergistically with CK2α () in the multiprotein complex that phosphorylates cytoplasmic β-catenin, encouraging its ubiquitination and degradation. Indeed, β-catenin nuclear translocation was prevented in spinal cord tissues taken from EAE mice (), whereas cytoplasmic p-β-catenin expression was increased (). On the contrary, moringin pretreatment positively regulated the Wnt–β-catenin signaling pathway in EAE mice. As demonstrated by Western blot analysis performed in spinal cord tissues, increased Wnt1 expression was found in pretreated EAE mice compared to untreated ones (). In addition, moringin pretreatment reduced the expression levels of GSK3β and CK2α (), by inhibiting phosphorylation of cytoplasmic β-catenin (). These results were further corroborated by enhanced expression of β-catenin in the nucleus ().
Figure 2 Moringin modulates the Wnt–β-catenin signaling pathway in EAE.
Abbreviations: EAE, experimental autoimmune encephalomyelitis; Ctl, control; ND, not detectable.
Moringin modulates apoptosis triggered by Wnt–β-catenin signaling pathway downregulation
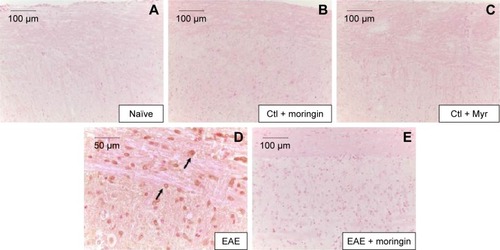
In order to investigate apoptosis, we studied Fas-ligand expression by immunohistochemistry in spinal cord sections. Results showed negative staining for Fas in naïve mice (), moringin controls (), and Myr control mice (). On the contrary, marked immunopositivity for Fas in untreated EAE mice (, arrows) was found. Moringin-pretreated EAE mice showed negative staining for Fas ().
Figure 3 Moringin modulates Fas-ligand expression in EAE.
Abbreviations: EAE, experimental autoimmune encephalomyelitis; Ctl, control.
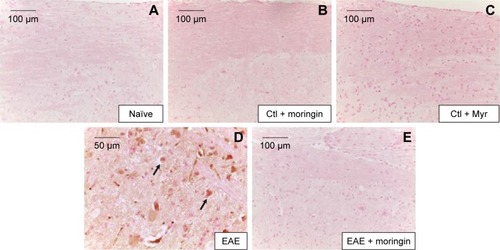
It is well known that activation of caspases is involved in apoptosis induction. Negative staining of cleaved caspase 9 was observed in naïve mice (), as well as in control groups (). Immunohistochemistry results also showed that cleaved caspase-9 level was considerably increased in EAE mice (). Moringin pretreatment totally suppressed the level of cleaved caspase 9 ().
Figure 4 Moringin modulates cleaved caspase-9 expression in EAE.
Abbreviations: EAE, experimental autoimmune encephalomyelitis; Ctl, control.
Moringin regulates production of CD4 and Treg cells
CD4 T-cell expression is involved in cell-mediated immunity and the pathogenesis of MS, with destruction of the axonal myelin sheath in several areas of the spinal cord mediated mainly by self-reactive CD4 T cells. Immunohistochemical analysis performed in spinal cord sections showed negative staining for CD4 in naïve (), moringin controls (), and Myr control mice (). Positive staining for CD4 was observed in EAE mice (); conversely, no positive staining for CD4 expression was obtained in mice pretreated with moringin (). In addition, in order to verify whether treatment with moringin could modulate the production of regulatory T (Treg) cells, we evaluated expression of the transcription factor FoxP3 by immunohistochemical analysis. Our results showed negative staining for FoxP3 in naïve mice () and in moringin, as well as the Myr control group (). Spinal cord sections from EAE mice showed positive staining for FoxP3 (), which was not observed in the tissues of EAE mice pretreated with moringin ().
Figure 5 Moringin modulates CD4 expression in EAE.
Abbreviations: EAE, experimental autoimmune encephalomyelitis; Ctl, control.
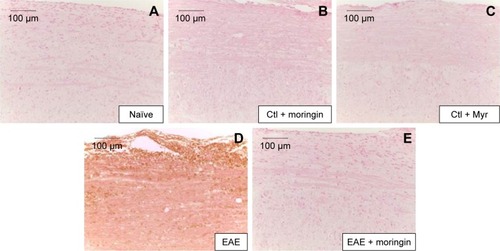
Figure 6 Moringin modulates FoxP3 expression in EAE.
Abbreviations: EAE, experimental autoimmune encephalomyelitis; Ctl, control.
Moringin modulates neuroinflammation triggered by Wnt–β-catenin signaling pathway downregulation
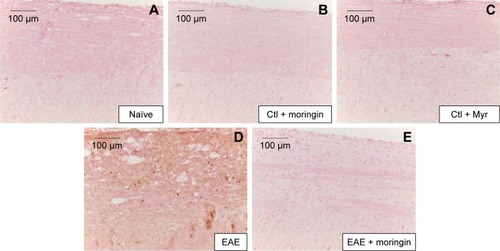
Enhanced expression of proinflammatory cytokines has been associated with many neurodegenerative diseases, including MS. Furthermore, as it is known that Wnt–β-catenin signaling can regulate cytokine production, we analyzed the expression of IL-1β and IL-6 in spinal cord tissues collected from EAE-untreated and moringin-pretreated EAE mice by immunohistochemical staining.
As shown in , no positive staining for IL-1β () or IL-6 () was obtained in naïve mice, in the moringin control group (), or in Myr ones (), while high levels of the these proinflammatory mediators were noticed in spinal cord tissues of EAE mice. Specifically, positive inflammatory cells are indicated by arrows in spinal cord sections stained with IL-1β and IL-6 (, respectively) and arrowheads show vascular endothelium positive for IL-6 (). Negative staining for IL-1β and IL-6 was observed in spinal cord tissues from EAE mice pretreated with moringin (, respectively). In addition, we investigated COX2 expression by Western blot analysis (). A basal level of COX2 was detected in naïve animals and control groups, while its level was significantly increased in EAE mice. Moringin-pretreated EAE mice showed reduced COX2 expression. In addition, we investigated whether moringin could be a potential activator of PPARγ via Wnt–β-catenin signaling. By Western blot analysis, a mild increase in PPARγ expression was found in EAE mice, while administration of moringin markedly increased PPARγ levels. Neither naïve mice nor the Myr control group showed any expression for PPARγ, whereas the moringin control group showed PPARγ expression ().
Figure 7 Moringin modulates IL-1β expression in EAE.
Abbreviations: EAE, experimental autoimmune encephalomyelitis; Ctl, control.
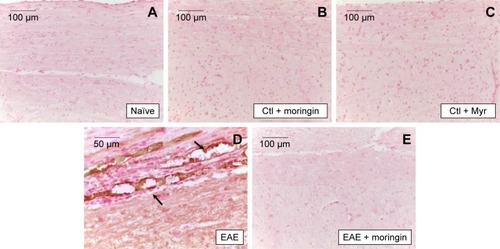
Figure 8 Moringin modulates IL-6 expression in EAE.
Abbreviations: EAE, experimental autoimmune encephalomyelitis; Ctl, control.
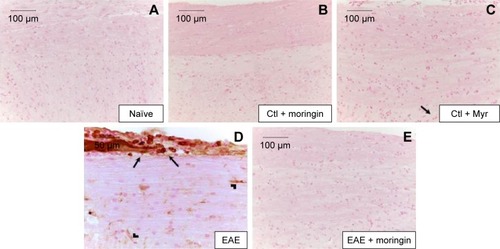
Figure 9 Moringin-modulated inflammatory mediators and Nrf2 activity in EAE.
Abbreviations: EAE, experimental autoimmune encephalomyelitis; Ctl, control; ND, not detectable.
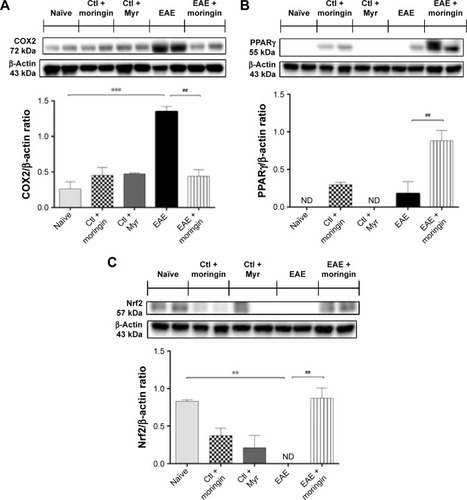
Moreover, as it is widely recognized that Glucosinolates (GLs) exert their protective effects by the capacity to induce expression of several enzymes via the Keap1–Nrf2–ARE pathway,Citation35,Citation36 we investigated expression of Nrf2. By Western blot analysis, we found that moringin pretreatment enhanced Nrf2 expression in EAE mouse spinal cord, while in untreated EAE mice, Nrf2 was absent ().
Discussion
MS is documented as the most common root of neurological disability.Citation37 Recent research has described the importance of the Wnt–β-catenin signaling pathway for normal functioning of the adult CNS, and its aberration has been reported in degenerative and inflammatory CNS diseases, like MS.Citation1,Citation9,Citation12,Citation38,Citation39 Moreover, Wnt signaling has been reported in immune cells present in the CNS, such as macrophages, microglia, and astrocytes, suggesting the critical role of the Wnt pathway in inflammation-mediated CNS injury and recovery.Citation40,Citation41 Although it is well known that the Wnt canonical pathway can modulate the immune system by repressing the inflammatory process during MS, the role of the Wnt canonical pathway in regulating remyelination remains controversial.Citation42 Some studies have supported the notion that the Wnt–β-catenin pathway may act as a negative modulator of the remyelination process, via inhibiting oligodendrocyte differentiation and myelin formation.Citation43–Citation45 Conversely, results from recent studies suggested that the Wnt canonical pathway may promote remyelination.Citation38,Citation46,Citation47
In the present study, we evaluated the involvement of the Wnt–β-catenin pathway in the etiopathology of an experimental EAE model and also tested a formulation of moringin, obtained from GMG by myrosinase-catalyzed hydrolysis, as a modulator of neuroinflammation via the β-catenin–PPARγ axis. In accordance with previous studies,Citation12,Citation48 our results showed that the canonical Wnt–β-catenin pathway is inactivated in EAE development. We found that in mice subjected to EAE, cytoplasmic β-catenin was constantly phosphorylated by increased expression of CK2α and GSK3β, which inhibited nuclear translocation of β-catenin and consequent activation of Wnt target genes involved in cell survival. The enhanced activity of CK2α and GSK3β was paralleled by Wnt1 suppression. Moreover, upregulation of GSK3β induced the degradation of β-catenin, which resulted in apoptosis of neurons. Apoptosis was confirmed by significant positive staining for proapoptotic Fas and cleaved caspase 9 in the spinal cord sections of EAE mice. Pretreatment with moringin markedly ameliorated the clinical score induced by EAE.
Interestingly, moringin pretreatment reverted the abnormal Wnt–β-catenin signaling of EAE mice. We found that moringin reduced levels of CK2α and GSK3β, which in turn increased Wnt1 and nuclear β-catenin levels. Reduction in GSK3β expression was further supported by the absence of cytoplasmic phosphorylated β-catenin, which resulted in the attenuation of apoptosis, evidenced by reduction in Fas and cleaved caspase 9. Moreover, we noticed that in the spinal cord of EAE mice, CD4 and FoxP3 levels were elevated, which indicated engagement and infiltration of Treg cells in the CNS. MOG-induced activation of CD4+/Foxp3+ Treg cells have been already demonstrated in EAE mice.Citation49 The proinflammatory mediators IL-1β, IL-6, and COX2 were also increased. Moringin repressed EAE-associated Treg-cell activation by diminishing CD4 and FoxP3 levels. Additionally, moringin decreased IL-1β, IL-6, and COX2 levels in EAE mice. Indeed, similar suppression of T-cell response in EAE has been demonstrated by sulforaphane, another widely examined isothiocyanate present in cruciferous vegetables.Citation50,Citation51 From our findings, we propose that moringin may control EAE-associated Treg-cell molecules via Wnt-signaling activation. It is well known that the Wnt-signaling pathway regulates T-cell activation.Citation52,Citation53 Increased Wnt1/β-catenin levels and decreased GSK3β level in moringin-administered EAE mice corroborated our notion, suggesting the active inhibition of MOG-induced T-cell activation via the moringin-mediated Wnt-signaling pathway.
Studies in recent years have demonstrated the beneficial efficacy of PPARγ agonists in the treatment of MS and other neurodegenerative diseases to suppress inflammatory and oxidative stress.Citation54–Citation56 Since PPARγ is associated with the Wnt–β-catenin pathway,Citation18,Citation19 we investigated whether moringin could be a potential activator of PPARγ via Wnt–β-catenin signaling. We observed a mild increase in PPARγ levels in EAE mice, which might have resulted from an innate anti-inflammatory response. Interestingly, EAE mice administered with moringin exhibited marked upregulation in PPARγ expression. We assume that the observed reduction of the proinflammatory mediators IL-1β, IL-6, and COX2 in moringin-treated EAE mice might be attributed to elevated levels of PPARγ.
Moreover, moringin pretreatment augmented antioxidant Nrf2 expression in EAE mice. It has been well documented that isothiocyanates may exert their antioxidative effects by Nrf2 activation.Citation36,Citation57 Of note, it is important to emphasize that GSK3β downregulation increases Nrf2 expression.Citation36 We assume that the enhanced expression of Nrf2 might have resulted from the moringin-regulated reduction of GSK3β. Our results are in parallel with previous studies, where it has been reported that synthetic sulforaphane, another widely examined isothiocyanate present in cruciferous vegetables, increased the level of Nrf2 by reducing GSK3β expression.Citation58–Citation60 shows a graphic representation of β-catenin signaling-mediated PPARγ and Nrf2 regulation in the presence or absence of Wnt.
Figure 10 Wnt–β-catenin canonical pathway.
Conclusion
Our results demonstrated that in EAE mice, moringin normalizes the aberrant Wnt–β-catenin pathway and inhibits GSK3β. Furthermore, moringin suppresses proinflammatory mediators via PPARγ activation and attenuates apoptosis. We propose that moringin might be a potential PPARγ agonist in the treatment of MS.
Authors contributions
SG performed the animal experiments and wrote the first draft of the manuscript. SG and TSR performed the biochemical analyses. GRDN and RI conducted the isolation and purification of GMG and myrosinase. EM performed immunohistochemical analysis. PB and EM designed the study. All authors contributed toward data analysis, drafting and critically revising the paper and agree to be accountable for all aspects of the work.
Acknowledgments
This study was supported by Current Research Funds 2014 of IRCCS Centro Neurolesi Bonino-Pulejo, Messina, Italy.
Disclosure
The authors report no conflicts of interest in this work.
References
- InestrosaNCArenasEEmerging roles of Wnts in the adult nervous systemNat Rev Neurosci2010112778620010950
- van AmerongenRMikelsANusseRAlternative Wnt signaling is initiated by distinct receptorsSci Signal2008135re918765832
- GordonMDNusseRWnt signaling: multiple pathways, multiple receptors, and multiple transcription factorsJ Biol Chem200628132224292243316793760
- KikuchiAYamamotoHSatoAMatsumotoSNew insights into the mechanism of Wnt signaling pathway activationInt Rev Cell Mol Biol2011291217122017973
- AberleHBauerAStappertJKispertAKemlerRβ-Catenin is a target for the ubiquitin-proteasome pathwayEMBO J19971613379738049233789
- PatapoutianAReichardtLFRoles of Wnt proteins in neural development and maintenanceCurr Opin Neurobiol200010339239910851180
- CianiLSalinasPCWnts in the vertebrate nervous system: from patterning to neuronal connectivityNat Rev Neurosci20056535136215832199
- OlivaCAVargasJYInestrosaNCWnts in adult brain: from synaptic plasticity to cognitive deficienciesFront Cell Neurosci2013722424348327
- ToledoEMColombresMInestrosaNCWnt signaling in neuroprotection and stem cell differentiationProg Neurobiol200886328129618786602
- Le GrandJNGonzalez-CanoLPavlouMASchwambornJCNeural stem cells in Parkinson’s disease: a role for neurogenesis defects in onset and progressionCell Mol Life Sci201572477379725403878
- BerwickDCHarveyKThe importance of Wnt signalling for neurodegeneration in Parkinson’s diseaseBiochem Soc Trans20124051123112822988876
- YuanSShiYTangSJWnt signaling in the pathogenesis of multiple sclerosis-associated chronic painJ Neuroimmune Pharmacol20127490491322547300
- TrappBDNaveKAMultiple sclerosis: an immune or neurodegenerative disorder?Annu Rev Neurosci20083124726918558855
- GovermanJAutoimmune T cell responses in the central nervous systemNat Rev Immunol20099639340719444307
- WeberMSMengeTLehmann-HornKCurrent treatment strategies for multiple sclerosis: efficacy versus neurological adverse effectsCurr Pharm Des201218220921922229582
- WillsonTMLambertMHKliewerSAPeroxisome proliferator-activated receptor gamma and metabolic diseaseAnnu Rev Biochem20017034136711395411
- HenekaMTLandrethGEHullMDrug insight: effects mediated by peroxisome proliferator-activated receptor-γ in CNS disordersNat Clin Pract Neurol20073949650417805244
- LecarpentierYClaesVDuthoitGHebertJLCircadian rhythms, Wnt/β-catenin pathway and PPAR α/γ profiles in diseases with primary or secondary cardiac dysfunctionFront Physiol2014542925414671
- SabatinoLPancioneMVotinoCEmerging role of the β-catenin-PPARγ axis in the pathogenesis of colorectal cancerWorld J Gastroenterol201420237137715124966585
- GiacoppoSGaluppoMMontautSAn overview on neuroprotective effects of isothiocyanates for the treatment of neurodegenerative diseasesFitoterapia2015106122126254971
- GaluppoMGiacoppoSDe NicolaGRAntiinflammatory activity of glucomoringin isothiocyanate in a mouse model of experimental autoimmune encephalomyelitisFitoterapia20149516017424685508
- BrunelliDTavecchioMFalcioniCThe isothiocyanate produced from glucomoringin inhibits NF-kB and reduces myeloma growth in nude mice in vivoBiochem Pharmacol20107981141114820006591
- MaldiniMMaksoudSANatellaFMoringa oleifera: study of phenolics and glucosinolates by mass spectrometryJ Mass Spectrom201449990091025230187
- Abdull RazisAFDe NicolaGRPagnottaEIoriRIoannidesC4-Methylsulfanyl-3-butenyl isothiocyanate derived from glucoraphasatin is a potent inducer of rat hepatic phase II enzymes and a potential chemopreventive agentArch Toxicol201286218319421960141
- Baasanjav-GerberCMonienBHMewisIIdentification of glucosinolate congeners able to form DNA adducts and to induce mutations upon activation by myrosinaseMol Nutr Food Res201155578379221213326
- BarillariJCervellatiRPaoliniMTatibouetARollinPIoriRIsolation of 4-methylthio-3-butenyl glucosinolate from Raphanus sativus sprouts (kaiware daikon) and its redox propertiesJ Agric Food Chem200553269890989616366671
- No authors listedEEC regulation 1864/90, enclosure VIIIOffic J Eur Commun1990L1702734
- GiacoppoSGaluppoMDe NicolaGRIoriRBramantiPMazzonE4(α-l-rhamnosyloxy)-benzyl isothiocyanate, a bioactive phytochemical that attenuates secondary damage in an experimental model of spinal cord injuryBioorg Med Chem2015231808825497964
- PessinaAThomasRMPalmieriSLuisiPLAn improved method for the purification of myrosinase and its physicochemical characterizationArch Biochem Biophys199028023833892369130
- GiacoppoSGaluppoMDe NicolaGRIoriRBramantiPMazzonETuscan black kale sprout extract bioactivated with myrosinase: a novel natural product for neuroprotection by inflammatory and oxidative response during cerebral ischemia/reperfusion injury in ratBMC Complement Altern Med20151539726545366
- GaluppoMGiacoppoSIoriRDe NicolaGRBramantiPMazzonEAdministration of 4-(α-L-rhamnosyloxy)-benzyl isothiocyanate delays disease phenotype in SOD1G93A rats: a transgenic model of amyotrophic lateral sclerosisBiomed Res Int2015201525941726075221
- PaschalidisNIqbalAJMaioneFModulation of experimental autoimmune encephalomyelitis by endogenous annexin A1J Neuroinflammation200963319912648
- RodriguesDHVilelaMCBarcelosLSPinhoVTeixeiraMMTeixeiraALAbsence of PI3Kγ leads to increased leukocyte apoptosis and diminished severity of experimental autoimmune encephalomyelitisJ Neuroimmunol20102221–2909420303183
- ConstantinescuCSFarooqiNO’BrienKGranBExperimental autoimmune encephalomyelitis (EAE) as a model for multiple sclerosis (MS)Br J Pharmacol201116441079110621371012
- HuRHebbarVKimBRIn vivo pharmacokinetics and regulation of gene expression profiles by isothiocyanate sulforaphane in the ratJ Pharmacol Exp Ther2004310126327114988420
- Dinkova-KostovaATHoltzclawWDColeRNDirect evidence that sulfhydryl groups of Keap1 are the sensors regulating induction of phase 2 enzymes that protect against carcinogens and oxidantsProc Natl Acad Sci U S A20029918119081191312193649
- NoseworthyJHLucchinettiCRodriguezMWeinshenkerBGMultiple sclerosisN Engl J Med20003431393895211006371
- MarchettiBPluchinoSWnt your brain be inflamed? Yes, it Wnt!Trends Mol Med201319314415623312954
- KimHWonSHwangDYDownregulation of Wnt/β-catenin signaling causes degeneration of hippocampal neurons in vivoNeurobiol Aging201132122316.e1e15
- HalleskogCMulderJDahlströmJWnt signaling in activated microglia is proinflammatoryGlia201159111913120967887
- L’EpiscopoFTiroloCTestaNReactive astrocytes and Wnt/β-catenin signaling link nigrostriatal injury to repair in 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine model of Parkinson’s diseaseNeurobiol Dis201141250852721056667
- XieCLiZZhangGXGuanYWnt signaling in remyelination in multiple sclerosis: friend or foe?Mol Neurobiol20144931117112524243343
- FancySPBaranziniSEZhaoCDysregulation of the Wnt pathway inhibits timely myelination and remyelination in the mammalian CNSGenes Dev200923131571158519515974
- ShimizuTKagawaTWadaTMuroyamaYTakadaSIkenakaKWnt signaling controls the timing of oligodendrocyte development in the spinal cordDev Biol2005282239741015950605
- GaesserJMFyffe-MaricichSLIntracellular signaling pathway regulation of myelination and remyelination in the CNSExp Neurol Epub201635
- HanafyKASloaneJARegulation of remyelination in multiple sclerosisFEBS Lett2011585233821382821443876
- YeFChenYHoangTHDAC1 and HDAC2 regulate oligodendrocyte differentiation by disrupting the β-catenin-TCF interactionNat Neurosci200912782983819503085
- SwaffordDManicassamySWnt signaling in dendritic cells: its role in regulation of immunity and toleranceDiscov Med20151910530331025977193
- Zorzella-PezaventoSFChiuso-MinicucciFFrancaTGPersistent inflammation in the CNS during chronic EAE despite local absence of IL-17 productionMediators Inflamm2013201351962723970813
- GeiselJBrückJGlocovaISulforaphane protects from T cell-mediated autoimmune disease by inhibition of IL-23 and IL-12 in dendritic cellsJ Immunol201419283530353924639357
- LiBCuiWLiuJSulforaphane ameliorates the development of experimental autoimmune encephalomyelitis by antagonizing oxidative stress and Th17-related inflammation in miceExp Neurol201325023924924120440
- van LoosdregtJFleskensVTiemessenMMCanonical Wnt signaling negatively modulates regulatory T cell functionImmunity201339229831023954131
- StaalFJLuisTCTiemessenMMWnt signalling in the immune system: Wnt is spreading its wingsNat Rev Immunol20088858159318617885
- DrewPDXuJRackeMKPPAR-γ: therapeutic potential for multiple sclerosisPPAR Res2008200862746318604287
- KaundalRKSharmaSSPeroxisome proliferator-activated receptor γ agonists as neuroprotective agentsDrug News Perspect201023424125620520853
- MrakRELandrethGEPPARγ, neuroinflammation, and diseaseJ Neuroinflammation200411515285797
- ErnstIMPalaniKEsatbeyogluTSchwarzKRimbachGSynthesis and Nrf2-inducing activity of the isothiocyanates iberverin, iberin and cheirolinPharmacol Res201370115516223403058
- RadaPRojoAIEvrard-TodeschiNStructural and functional characterization of Nrf2 degradation by the glycogen synthase kinase 3/β-TrCP axisMol Cell Biol201232173486349922751928
- ShangGTangXGaoPSulforaphane attenuation of experimental diabetic nephropathy involves GSK-3 β/Fyn/Nrf2 signaling pathwayJ Nutr Biochem201526659660625724107
- RojoAIRadaPEgeaJRosaAOLopezMGCuadradoAFunctional interference between glycogen synthase kinase-3 β and the transcription factor Nrf2 in protection against kainate-induced hippocampal cell deathMol Cell Neurosci200839112513218619545