
Abstract
Diabetes mellitus type 1 (DM1) is an autoimmune disease in which β-cells of the pancreas islet are destroyed by T lymphocytes. Specific T cells are activated by antigen-presenting cells, mainly dendritic cells (DCs). It is already known that the regulation of tryptophan pathway in DC can be a mechanism of immunomodulation. The enzyme indoleamine 2,3-dioxygenase (IDO) is present in many cells, including DC, and participates in the metabolism of the amino acid tryptophan. Recent studies suggest the involvement of IDO in the modulation of immune response, which became more evident after the in vitro demonstration of IDO production by DC and of the ability of these cells to inhibit lymphocyte function through the control of tryptophan metabolism. Current studies on immunotherapies describe the use of DC and IDO to control the progression of the immune response that triggers DM1. The initial results obtained are promising and indicate the possibility of developing therapies for the treatment or prevention of the DM1. Clinical trials using these cells in DM1 patients represent an interesting alternative treatment. However, clinical trials are still in the initial phase and a robust group of assays is necessary.
Introduction
Diabetes mellitus type 1 (DM1) is an autoimmune disease with massive destruction of pancreatic β-cells, with lymphocyte and macrophage infiltrates in the islets of Langerhans.Citation1,Citation2 Although already approved therapies to DM1 are available, they are still poorly effective in controlling the progression of diabetes. Recent advances in the studies of DM intended to develop cellular therapies, such as the regeneration of pancreatic β-cells from transplantation of precursor stem cells.Citation3,Citation4 To reverse the immunopathogenesis of the DM1, some authors propose an early vaccination as prophylaxis. Also, restoration of immune tolerance to self-antigens in DM1 patients is considered a promising therapy.Citation5 In this sense, treatment with immunoregulatory cells specifically explores the modulatory capacity of indoleamine 2,3-dioxygenase (IDO).
It is known that the enzyme IDO, expressed in dendritic cells (DCs) and other cells, participates in the metabolism of tryptophan (TRP). Recent studies suggest the participation of IDO in the modulation of the immune response, which became more evident after the in vitro demonstration of IDO production by DC and of the ability of these cells to inhibit lymphocytes via the TRP pathway.Citation6–Citation10 In this subject, experiments with macrophages and human DC expressing IDO have demonstrated that the presence of IDO blocks in vitro proliferation of T cells.Citation7 IDO1 is an immunomodulatory molecule with potential effects on various diseases including cancer and autoimmunity.Citation11,Citation12 IDO2, as well as IDO1, is implicated in TRP catabolism and modulation of immune response. IDO2 seems to act in a distinct way from IDO1. However, the specific contributions of IDO2 to physiology are still unknown.Citation13
A recent study suggests that IDO1 activity may correlate with the pathogenesis of diabetes mellitus type 2 (DM2) since it was significantly increased in these patients.Citation14 Reduction in IDO1 levels also seems to be related to the pathogenesis of DM1,Citation15 since in experimental diabetes, toll-like receptor (TLR) 3-deficient mice presented a more robust disease, accompanied by the absence of IDO1 induction in pancreatic lymph nodes.
However, new mechanisms to control DM1 as well as to elucidate the participation of the IDO1 enzyme in the DC response are under development. In this context, the study of the mechanism of action of IDO1 and its relationship with the immunological mechanism can be relevant for the future development of vaccines or drugs. Hence, the aim of the present review was to describe the participation of DC in the control of the immune response on DM1, as well as the regulation of the IDO1 enzyme in these cells.
Immunological mechanisms of type 1 diabetes mellitus
Current studies demonstrate that DC can participate in the regulation of DM1,Citation16 since it is a disorder related to the action of autoreactive T cells against pancreatic β-cells.Citation17 CD11c+ CD11b+ CD8α− DC activates T lymphocytes that initiate the process of insulitis and eliminates pancreatic β-cells.Citation18,Citation19 Autoantibodies subsequently participate in inflammatory process and can be used as markers of immune response.Citation20 It is also known that the enzyme IDO, present in DC, participates in the immunity by regulating the proliferation of T cells.Citation7
Indoleamine 2,3-dioxygenase and the kynurenine pathway
IDO is one of the two enzymes that catalyze the limiting step of the degradation of TRP through the kynurenine (Kyn) pathway. Studies showed that the tryptophan 2,3-dioxygenase (TDO) and IDO enzymes catalyze the conversion of TRP to N-formyl kynurenine.Citation21,Citation22 Unlike TDO, expressed only in the liver, IDO is found in various organs, tissues, and cell types, such as stromal, hematopoietic, tumor, lymphoid, and dendritic cells,Citation21,Citation22 and expressed often in tissues with large mucosa, such as the gastrointestinal tract and lung.Citation23
IDO is an oxygenase enzyme with a heme groupCitation24 composed of two domains – major and minorCitation25 – encoded by two closely related genes (IDO1 and IDO2), located on human chromosome 8.Citation26,Citation27 In the TRP catabolism, IDO is responsible for the oxidative cleavage of double bond present in the indol fraction of TRP, resulting in the formation of N-formyl kynurenine.Citation25 The kynurenine pathway () is the main route of degradation of TRP, since 99% of this ingested amino acid is metabolized by this way. The kynurenines generated include kynurenic acid, 3-hydroxykynurenine, picolinic acid, quinolinic acid, and kynurenine itself.Citation25,Citation28,Citation29 IDO enzyme is able to inhibit the in vitro proliferation of T cells by the catabolism of TRP via kynurenines, participating in the modulation of immune response.Citation30
Figure 1 Kynurenine and serotonin pathway.
Several studies indicate that the transcription process is well controlled after the activation of IDO, with activation of genes of inflammatory mediators, such as interferon (IFN)-α, IFN-β, and IFN-γ. Among several IDO inducers, lipopolysaccharide (LPS)-induced IDO expression was found in blood, muscle, brain, liver, and lung. LPS administration induces increased concentrations of TNF-α, IL-10, and Kyn, and a depletion of TRP indicating an elevated activity of IDO.Citation28,Citation31
Although IDO expression is rapidly induced by IFN-γ alone or associated with interleukin (IL)-1β, IDO secretion also occurs in DC after signaling by transforming growth factor beta 1 (TGF-β1), which demonstrates a mediation of its regulatory function.Citation7,Citation30,Citation32–Citation34 The mechanism of control of IDO expression is not fully understood but post-translational modifications are relevant in this process. Also, the induction of IDO in DC can occur through immunomodulatory properties of molecules, that inhibit the pathways of T cells activation via T cell receptor (TCR), Cytotoxic T-lymphocyte-Associated Associated Antigen 4 (CTLA-4), Toll like receptors (TLR) and CD40.Citation23,Citation35
The immunoregulatory properties of IDO on DC
The role of IDO on immunosuppression was first discussed in 1998 when Munn et al demonstrated that when pregnant rats were treated with an inhibitor of the IDO enzyme called 1-methyl tryptophan, maternal lymphocytes initiated a process of rejection to the fetus. Subsequently, the studies of IDO in immunosuppression extended to regulation of tumors, inflammatory, allergic and autoimmune diseases, the enzyme being recognized as a regulator of immunity.Citation34,Citation36,Citation37 This regulatory action of IDO is possibly due to the combination of the effects of TRP degradation and of the accumulation of the kynurenine metabolites that act on the hydrocarbon ring present at the TCR.Citation17,Citation34
IDO can affect the ability of DC to present antigens and stimulate T cells. Also, the catabolism of the TRP is a central pathway to this immunosuppressive response. In this sense, IDO can induce the depletion of TRP and the accumulation of kynurenine metabolites, as exemplified in .Citation23,Citation34 The T cells activated by DC expressing IDO are able to initiate the cell cycle. However, the enzymatic activity of IDO leads to a significant decrease of TRP, inducing the cell cycle arrest and enhancing T cell apoptosis. IDO1 was described as the first rate-limiting enzyme acting on the degradation of TRP since the enzyme activity results in the depletion of TRP and inhibition of T lymphocytes.Citation38 The initial step of the kynurenine pathway involves the oxidation of TRP. TRP derivatives have pleiotropic effects on homeostasis processes. IDO and TDO are heme-containing enzymes that catalyze the O2-dependent oxidation of L-TRP in biological systems. Studies report that IDO and TDO react using the same mechanism. For IDO, a ferryl species accumulates in the steady state. However, there is no evidence for the accumulation of this compound during TDO catalysis. Serotonin and melatonin are two TRP-derived compounds produced in humans, among others. Also, indole ring breakdown through the “kynurenine shunt” produces a number of molecules involved in inflammation, immune response, excitatory neurotransmission, and many other functions, such as L-kynurenine, kynurenic and quinolinic acids, or the coenzyme nicotinamide adenine dinucleotide (NAD+).Citation39–Citation42
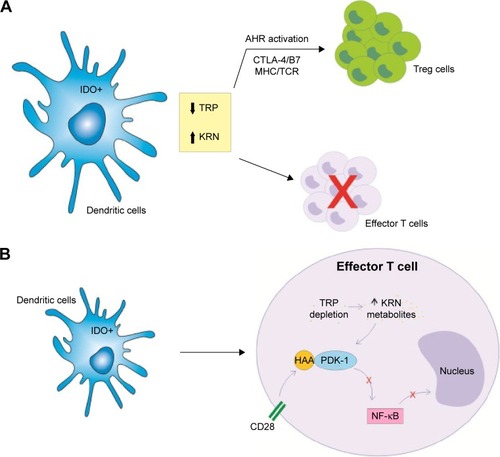
Figure 2 IDO and DC in the regulation of the T cell responses through the degradation of TRP and kynurenine metabolites.
Abbreviations: AHR, aryl hydrocarbon receptor; CTLA-4, cytotoxic T-lymphocyte-associated antigen 4; DCs, dendritic cells; HAA, 3-hydroxyanthranilic acid; IDO, indoleamine 2,3-dioxygenase; KRN, kynurenine; MHC, major histocompatibility complex; NF-kB, nuclear factor kappa B; PDK-1, phosphoinositide-dependent kinase 1; TCR, T cell receptor; Treg, T regulatory; TRP, tryptophan.
Recent research showed that catabolism of TRP mediated by IDO inhibits the immunoregulatory kinase mammalian target of rapamycin (mTOR), with the autophagy induction.Citation43 So, IDO was proposed to control inflammatory responses and immune tolerance coordinately by the pathways of TRP deficiency and sufficiency, via the integrated stress kinases namely general control non-depressible 2 (GCN2) and mTOR, respectively.Citation43,Citation44 However, the role of GCN2 in immune response and suppression is not clearly established. Conflicting data showed that, in lupus-prone mice, activation of GCN2 reduced anti-DNA antibodies, implicating GCN2 signal in limiting autoimmunity.Citation45 However, a recent study described that kinase GCN2 is not related to the suppression of antitumor response by TRP catabolism.Citation46
Tryptophan catabolites or starvation induce a regulatory environment affecting both CD8+ and CD4+ T cells. Recent studies showed that TRP catabolites result in GCN2 kinase-dependent downregulation of the TCR zeta-chain in CD8+ T cells in mouse model. Also, TRP catabolism includes the emergence of a regulatory phenotype in naive CD4+ CD25− T cells via TGF-β induction of the forkhead transcription factor Foxp3, capable of controlling diabetogenic T cells.Citation47,Citation48
The metabolites generated by the IDO activity are potent inhibitors of T cell activation. Some compounds such as 3-hydroxyanthranilic acid (3-HAA) are able to block T cells by interfering in the nuclear factor kappa B (NF-kB) pathway via inhibition of phosphoinositide-dependent kinase 1 (PDK-1), a mediator of NF-kB activation through CD28 signalingCitation49,Citation50 (). Besides that, kynurenine is also involved in the generation of T regulatory (Treg) in periphery, another important mechanism to control T cell responses. In lymph nodes, IDO controls conversion of Foxp3+ Tregs to T helper (Th) 17 cells.Citation51 In this way, TRP metabolites are able to ligate to aryl hydrocarbon receptor (AHR), a cytoplasmic transcription factor, favoring differentiation of naive T cells to Tregs but not to Th17 cells.Citation52 However, literature suggests that other signaling pathways are also relevant, like the CTLA-4/B7 signaling and the major histocompatibility complex (MHC)–TCR interaction.Citation23,Citation53
Related to IDO therapeutic potential, synergistic treatment with TGF-β can result in a burst of regulatory kynurenines that contribute to establishing a state of “infectious tolerance”. TGF-β activates the tolerogenic pathway of TRP catabolism – mediated by IDO.Citation54 IDO can be associated with the pathogenesis of persistent infections, and Treg cells might play an important role in the direct control of innate immune responses. According to an infectious tolerance model, Treg cells inhibit neutrophil function and promote their apoptosis by the induction of suppressor cell populations. LPS-activated Treg cells induce IL-10 and TGF-β1 secretion, inhibit IL-6 production, and also induce the expression of heme oxygenase-1 (HO-1) and the suppressor of cytokine signaling 3 molecule (SOCS3).Citation55,Citation56
Tregs induce IDO expression in DCs and tolerogenic DC induce conversion of naive T cells to Tregs. Both myeloid-derived suppressor cells (MDSCs) subset and tolerogenic DCs can control self-reactive immune responses by promoting the expansion of Treg population. MDSCs require direct contact with activated T cells, which leads to the production of IDO.Citation57
A recent report showed that high-dose IL-4 inhibits the CD103+ DC differentiation by inducing high levels of IDO. Interestingly, IL-4 diminished IDO expression in DCs in a dose-dependent manner. DCs generated under low concentrations of IL-4 favored Treg cells differentiation, which depend on IDO produced by CD103+ DCs.Citation58
Therapy with tolerogenic DC in diabetes mellitus
The conventional DM therapies basically include medications to control glucose and insulin levels. However, with the wide knowledge on cell biology, immunology, and biotechnology, cell therapy has been gaining prominence.Citation59 It can be used in the treatment of infections, autoimmune diseases, cancer, metabolic diseases, and tissue degeneration.Citation59,Citation60 Also, cell therapy is becoming the alternative treatment of DM1 and several studies approach this theme in different ways.Citation61
Studies demonstrate the possibility of development of in vitro insulin-producing cells with characteristics similar to pancreatic β-cells, using embryonic stem cells as source. Replacement of pancreatic β-cells has already been considered as an alternative to treat DM1.Citation62 Jalili et al prolonged islet allograft survival by preventing immune response. They induced a local immunosuppressive activity by stable IDO induction in bystander fibroblasts.Citation63 Also, administration of endovenous anti-CD3 monoclonal antibodies in patients newly diagnosed with DM1 results in the reduction of circulating lymphocytes as well as C-peptide, insulin, and glycated hemoglobin (HbAL1c) values.Citation64,Citation65 In addition, experiments combining transplantation of autologous hematopoietic stem cell and immunosuppression protocol based on endovenous cyclophosphamide and anticytoplasmic globulin restarted the immune system without autoreactive memory to the regenerated pancreatic β-cells. A high percentage of patients obtained insulin independence, without mortality or new toxic effects.Citation66–Citation68
Among therapies, DC therapies can be highlighted for autoimmune diseases such as DM1, since these cells participate in the pathogenesis of this disease.Citation69,Citation70 DC therapies are evaluated for the treatment of infectious diseases, tumor, autoimmune diseases, transplants, and allergies.Citation50,Citation69,Citation70 In this context, it is known that plasmacytoid type DC (pDC), that participate in the immune response and the generation of tolerance, can act in the DM1 in the control of the regulatory and pathogenic T cells.Citation17,Citation34
In order to develop therapies for DM1 using tolerogenic DC, a study was performed ablating the myeloid DC (mDC) and pDC mediated by diphtheria toxin receptor in nonobese diabetic mice (NOD). The results showed that, with the ablation of mDC, prevention of the development of DM was achieved. However, with the ablation of pDC, an increase in the severity of the insulitis was observed, accompanied by the loss of natural killer T cells (NKT) and a decrease of IDO. Since IDO enzyme is an inhibitor of T cell response, the decrease of pDC or the inhibition of IDO by the use of 1-methyl-D-tryptophan (1-MT) increased the aggressiveness of the insulitis and the destruction of pancreatic β-cells. Also, the ablation of pDC caused reallocation of NKT cells that aggravates the insulitis, while the restoration of pDC normalized the insulin and NKT levels. The IFN-γ secretion by the NKT induced IDO production by DC, interconnecting these responses. Finally, it was concluded that pDC, IDO, and NKT can act together to prevent insulin production and, consequently, the development of DM1.Citation71,Citation72
Another study demonstrated that NOD mice have a deficiency in the signaling and functioning of the IDO enzyme, even after stimulation of pDC with TGF-β1. IDO could only be stimulated by this cytokine in the presence of a basal amount of this enzyme in the pDC. So, in order to initiate the phosphorylation cascade and the consequent protein signaling, it was necessary to perform a forced induction of the IDO in the pDC and the stimulation with TGF-β1. IDO signaling generated a positive feedback with TGF-β1, alternatively activating the NF-kB pathway.Citation17
A recent study showed that a chimeric vaccine composed of the proinsulin and cholera toxin B subunit (CTB-INS) was able to suppress DM1 onset in NOD mice and upregulated enzyme IDO1 in human DC. This vaccine activated the tumor necrosis factor associated factor (TRAF) super-family receptor (TRAFR) pathway and led to the upregulation of IDO1 biosynthesis in CTB-INS inoculated DC. The control of NF-kB-inducing kinase (NIK) post-translational stability may be essential for the modulation of non-canonical NF-kB signaling pathway. Evidence suggests that TRAF2, TRAF3, and TRAF6 are critical to negative regulation of NIK activity. The CTB-INS vaccine acts in the TNF receptor-dependent signaling pathway of the noncanonical NF-kB signaling pathway resulting in the suppression of mediated DM1.Citation73
DCs present autoantigens of pancreatic β-cells for self-reactive T lymphocytes. They are also involved in the deletion of autoreactive T cells. A report showed that targeting of DCs with beta cell antigens was able to achieve deletion of autoreactive cytotoxic T cells in NOD model.Citation74
After treating DC with CTB-INS vaccine, an increase in the degradation of the TRP, a reduction in cell maturation functions, and an augment of the IDO1 synthesis were observed. This biosynthesis was reduced with NF-kB blockers reinforcing the role of this factor in the induction of IDO. All these findings, combined with the fact that this does not interfere with the monocytes differentiation into DC, open the possibility of developing safe and effective immunotherapeutic strategies for the prevention of autoimmunity that occurs in DM1.Citation16
A clinical trial using DC for the treatment of DM1 is currently underway.Citation75–Citation79 The vaccine consists of autologous monocyte-derived DC treated ex vivo with antisense phosphorothioate-modified oligonucleotides targeting the primary transcripts of the CD40, CD80, and CD86 co-stimulatory molecules, generating DC with immunoregulatory properties. The safety Phase I studyCitation80 allowed to conclude that the use of tolerogenic DC in patients with DM1 was well accepted and safe, without adverse effects or toxicity.Citation76 In Phase II, the modulatory capability of DC collected via leukapheresis and engineered ex vivo via incubation with antisense DNA oligonucleotides targeting the primary transcripts of CD40, CD80, and CD86 will be evaluated.Citation80,Citation81
Final considerations
Nowadays, DC therapies to DM1 have focus on the immuno-regulatory properties of these cells through their maturation and on the action of the IDO, an enzyme that participates in the metabolism of TRP and causes accumulation of the metabolites of kynurenines and consequent blockade of T lymphocyte proliferation. As shown here, the use of DCs in cell therapies in DM1, although recent, already presents promising results since it has been proven that the IDO enzyme can be modulated in DC in a tolerogenic way. In this sense, it was possible to conclude that recent studies have demonstrated that DC expressing tolerogenic IDO may be effective in regulating the immune response of DM1. Therefore, the data support the suggestion of the application of tolerogenic DCs to induce tolerance in DM1. It is important to highlight that studies with DC on DM1 are in clinical phase and the results obtained heretofore are promising since they indicate the possibility of developing therapies for the treatment or prevention of the DM1. However, clinical trials are still in the initial phase and a more robust group of assays to confirm this proposition is necessary.
Disclosure
The authors report no conflicts of interest in this work.
References
- ZhangCLZouXLPengJBXiangMImmune tolerance induced by adoptive transfer of dendritic cells in an insulin-dependent diabetes mellitus murine modelActa Pharmacol Sin2007281 98 10417184588
- LakhtakiaRThe history of diabetes mellitusSultan Qaboos Univ Med J2013133 368 37023984020
- GodfreyKJMathewBBulmanJCShahOClementSGallicanoGIStem cell-based treatments for type 1 diabetes mellitus: bone marrow, embryonic, hepatic, pancreatic and induced pluripotent stem cellsDiabet Med2012291 14 2321883442
- BouwensLHoubrackenIMfopouJKThe use of stem cells for pancreatic regeneration in diabetes mellitusNat Rev Endocrinol2013910 598 60623877422
- LeeCNLewAMWuLThe potential role of dendritic cells in the therapy of type 1 diabetesImmunotherapy201356 591 60623725283
- MellorAIndoleamine 2,3 dioxygenase and regulation of T cell immunityBiochem Biophys Res Commun20053381 20 2416157293
- MellorALMunnDHIDO expression by dendritic cells: tolerance and tryptophan catabolismNat Rev Immunol2004410 762 77415459668
- MunnDHSharmaMDLeeJRPotential regulatory function of human dendritic cells expressing indoleamine 2,3-dioxygenaseScience20022975588 1867 187012228717
- FavreDMoldJHuntPWTryptophan catabolism by indoleamine 2,3-dioxygenase 1 alters the balance of TH17 to regulatory T cells in HIV diseaseSci Transl Med2010232 32ra36
- XieFTCaoJSZhaoJYuYQiFDaiXCIDO expressing dendritic cells suppress allograft rejection of small bowel transplantation in mice by expansion of foxp3+ regulatory T cellsTranspl Immunol2015332 69 7726002283
- Mandik-NayakLMerloLIDO2: a pathogenic mediator of inflammatory autoimmunityClin Med Insights Pathol20169Suppl 1 21 2827891058
- LiFZhangRLiSLiuJIDO1: an important immunotherapy target in cancer treatmentInt Immunopharmacol201747 70 7728365507
- PrendergastGCMetzRMullerAJMerloLMMandik-NayakLIDO2 in immunomodulation and autoimmune diseaseFront Immunol20145 58525477879
- ZhangYRuanYZhangPWangLIncreased indoleamine 2,3-dioxygenase activity in type 2 diabetic nephropathyJ Diabetes Complications2017311 223 22727646613
- FallarinoFVolpiCZelanteTIDO mediates TLR9-driven protection from experimental autoimmune diabetesJ Immunol200918310 6303 631219841163
- MbongueJCNicholasDAZhangKInduction of indoleamine 2, 3-dioxygenase in human dendritic cells by a cholera toxin B subunit-proinsulin vaccinePLoS One2015102 e011856225714914
- PallottaMTOrabonaCBianchiRForced IDO1 expression in dendritic cells restores immunoregulatory signaling in autoimmune diabetesJ Cell Mol Med20141810 2082 209125215657
- TurleySPoirotLHattoriMBenoistCMathisDPhysiological beta cell death triggers priming of self-reactive T cells by dendritic cells in a type-1 diabetes modelJ Exp Med200319810 1527 153714623908
- FaustmanDLDavisMThe primacy of CD8 T lymphocytes in type 1 diabetes and implications for therapiesJ Mol Med(Berl)20098712 1173 117819693476
- SzablewskiLRole of immune system in type 1 diabetes mellitus pathogenesisInt Immunopharmacol2014221 182 19124993340
- TaylorMWFengGSRelationship between interferon-gamma, indoleamine 2,3-dioxygenase, and tryptophan catabolismFASEB J1991511 2516 25221907934
- MbongueJCNicholasDATorrezTWThe role of indoleamine 2,3-dioxygenase in immune suppression and autoimmunityVaccines201533 703 72926378585
- HuangLBabanBJohnsonBA3rdMellorALDendritic cells, indoleamina 2,3 dioxygenase and acquired immune privilegeInt Rev Immunol2010292 133 15520367139
- CurtiATrabanelliSSalvestriniVBaccaraniMLemoliRMThe role of indoleamine 2, 3-dioxygenase in the induction of immune tolerance: focus on hematologyBlood200911311 2394 240119023117
- SugimotoHOdaSOtsukiTHinoTYoshidaTShiroYCrystal structure of human indoleamine 2,3-dioxygenase: catalytic mechanism of O2 incorporation by a heme-containing dioxygenaseProc Natl Acad Sci U S A20061038 2611 261616477023
- MellorALChandlerPBabanBSpecific subsets of murine dendritic cells acquire potent T cell regulatory functions following CTLA4-mediated induction of indoleamine 2,3 dioxygenaseInt Immunol20041610 1391 140115351783
- MunnDHSharmaMDMellorALLigation of B7–1/B7-2 by human CD4+ T cells triggers indoleamine 2,3-dioxygenase activity in dendritic cellsJ Immunol20041727 4100 411015034022
- FatokunAHuntNHBallHJIndoleamine 2,3-dioxygenase 2 (IDO2) and the kynurenine pathway: characteristics and potential roles in health and diseaseAmino Acids2013456 1319 132924105077
- MoffettJRNamboodiriMATryptophan and the immune responseImmunol Cell Biol2003814 247 26512848846
- AllegriGCostaCVBertazzoABiasioloMRagazziEEnzyme activities of tryptophan metabolism along the kynurenine pathway in various species of animalsFarmaco2003589 829 83613679176
- WirthgenETuchschererMOttenWActivation of indoleamine 2,3-dioxygenase by LPS in a porcine modelInnate Immun2014201 30 3923606516
- FallarinoFGrohmannUPuccettiPIndoleamine 2,3-dioxygenase: from catalyst to signaling functionEur J Immunol2012428 1932 193722865044
- SarkarSAWongRHacklSIInduction of indoleamine 2,3- dioxygenase by interferon-gamma in human isletsDiabetes2007561 72 7917192467
- PallottaMTOrabonaCVolpiCIndoleamine 2,3-dioxygenase is a signaling protein in long-term tolerance by dendritic cellsNat Immunol2011129 870 87821804557
- HuangHDawickiWZhangXTownJGordonJRTolerogenic dendritic cells induce CD4+CD25hiFoxp3+ regulatory T cell differentiation from CD4+CD25-/loFoxp3- effector T cellsJ Immunol20101859 5003 501020870943
- MorrowLEGogineniVMaleskerMAProbiotic, prebiotic, and synbiotic use in critically ill patientsCurr Opin Crit Care2012182 186 19122343306
- Poormasjedi-MeibodMSJaliliRBHosseini-TabatabaeiAHartwellRGhaharyAImmuno-regulatory function of indoleamine 2,3 dioxygenase through modulation of innate immune responsesPLoS One201388 e7104423940687
- PlattenMvon Knebel DoeberitzNOezenIWickWOchsKCancer immunotherapy by targeting IDO1/TDO and their downstream effectorsFront Immunol20155 67325628622
- PalegoLBettiLRossiAGiannacciniGTryptophan biochemistry: structural, nutritional, metabolic, and medical aspects in humansJ Amino Acids20162016 895252026881063
- BasranJBoothESLeeMHandaSRavenELAnalysis of reaction intermediates in tryptophan 2,3-dioxygenase: a comparison with indoleamine 2,3-dioxygenaseBiochemistry20165549 6743 675027951658
- BatabyalDYehSRSubstrate-protein interaction in human tryptophan dioxygenase: the critical role of H76J Am Chem Soc20091319 3260 327019209904
- MengBWuDGuJOuyangSDingWLiuZJStructural and functional analyzes of human tryptophan 2,3-dioxygenaseProteins20148211 3210 321625066423
- MetzRRustSDuhadawayJBIDO inhibits a tryptophan sufficiency signal that stimulates mTOR: A novel IDO effector pathway targeted by D-1-methyl-tryptophanOncoimmunology201219 1460 146823264892
- MunnDHMellorALIndoleamine 2,3 dioxygenase and metabolic control of immune responsesTrends Immunol2013343 137 14323103127
- RavishankarBLiuHShindeRThe amino acid sensor GCN2 inhibits inflammatory responses to apoptotic cells promoting tolerance and suppressing systemic autoimmunityProc Natl Acad Sci U S A201511234 10774 1077926261340
- SonnerJKDeumelandtKOttMThe stress kinase GCN2 does not mediate suppression of antitumor T cell responses by tryptophan catabolism in experimental melanomasOncoimmunology2016512 e124085828123877
- FallarinoFGrohmannUYouSThe combined effects of tryptophan starvation and tryptophan catabolites down-regulate T cell receptor zeta-chain and induce a regulatory phenotype in naive T cellsJ Immunol200617611 6752 676116709834
- FallarinoFGrohmannUYouSTryptophan catabolism generates autoimmune-preventive regulatory T cellsTranspl Immunol2006171 58 6017157218
- HayashiTMoJHGongX3-Hydroxyanthranilic acid inhibits PDK1 activation and suppresses experimental asthma by inducing T cell apoptosisProc Natl Acad Sci U S A200710447 18619 1862418003900
- Silva PdeMBierJPaiattoLNTolerogenic dendritic cells on transplantation: immunotherapy based on second signal blockageJ Immunol Res20152015 85670726543876
- SharmaMDHouDYLiuYIndoleamine 2,3-dioxygenase controls conversion of Foxp3+ Tregs to TH17-like cells in tumor-draining lymph nodesBlood200911324 6102 611119366986
- PlattenMHoPPYoussefSTreatment of autoimmune neuroin-flammation with a synthetic tryptophan metaboliteScience20053105749 850 85516272121
- EleftheriadisTPissasGAntoniadiGLiakopoulosVStefanidisIIndoleamine 2,3-dioxygenase depletes tryptophan, activates general control non-derepressible 2 kinase and down-regulates key enzymes involved in fatty acid synthesis in primary human CD4+ T cellsImmunology20151462 292 30026147366
- BelladonnaMLOrabonaCGrohmannUPuccettiPTGF-β and kynurenines as the key to infectious toleranceTrends Mol Med2009152 41 4919162548
- BarthHRaghuramanSPersistent infectious diseases say – IDO. Role of indoleamine-2,3-dioxygenase in disease pathogenesis and implications for therapyCrit Rev Microbiol2014404 360 36823174025
- LewkowiczNKlinkMMyckoMPLewkowiczPNeutrophil – CD4+CD25+ T regulatory cell interactions: a possible new mechanism of infectious toleranceImmunobiology20132184 455 46422749980
- ZosoAMazzaEMBicciatoSHuman fibrocytic myeloid-derived suppressor cells express IDO and promote tolerance via Treg-cell expansionEur J Immunol20144411 3307 331925113564
- TuLChenJZhangHDuanLInterleukin-4 inhibits regulatory T cell differentiation through regulating CD103+ dendritic cellsFront Immunol20178 21428316599
- FischbachMABluestoneJALimWACell-based therapeutics: the next pillar of medicineSci Transl Med20135179 179ps7
- GarciaOCarraroGNavarroSCell-based therapies for lung diseaseBr Med Bull2012101 147 16122279079
- VoltarelliJCCouriCERodriguesMCTerapia celular no diabetes mellitusRev Bras Hematol Hemoter200931 149 156
- AssadySMaorGAmitMItskovitz-EldorJSkoreckiKLTzukermanMInsulin production by human embryonic stem cellsDiabetes2001508 1691 169711473026
- JaliliRBForouzandehFRezakhanlouAMLocal expression of indoleamine 2,3 dioxygenase in syngeneic fibroblasts significantly prolongs survival of an engineered three-dimensional Islet allograftDiabetes2010599 2219 222720522587
- HeroldKCHagopianWAugerJAAnti-CD3 monoclonal antibody in new-onset type 1 diabetes mellitusN Engl J Med200234622 1692 169812037148
- KeymeulenBVandemeulebrouckeEZieglerAGInsulin needs after CD3-antibody therapy in new-onset type 1 diabetesN Engl J Med200535225 2598 260815972866
- CouriCEFossMCVoltarelliJCSecondary prevention of type 1 diabetes mellitus: stopping immune destruction and promoting beta-cell regenerationBraz J Med Biol Res20063910 1271 128016941054
- StracieriABOliveiraMCSimoBPAutologous nonmyeloablative hematopoietic stem cell transplantation in newly diagnosed type 1 diabetes mellitusJAMA200729714 1568 157617426276
- VanikarAVTrivediHLThakkarUGStem cell therapy emerging as the key player in treating type 1 diabetes mellitusCytotherapy2016189 1077 108627424148
- SteinmanRMDendritic cells: understanding immunogenicityEur J Immuno200737Suppl 1 S53 S60
- SteinmanRMBanchereauJTaking dendritic cells into medicineNature20074497161 419 42617898760
- SaxenaVOndrJKMagnusenAFMunnDHKatzJDThe countervailing actions of myeloid and plasmacytoid dendritic cells control autoimmune diabetes in the nonobese diabetic mouseJ Immunol20071798 5041 505317911589
- NikolicTWelzen-CoppensJMLeenenPJDrexhageHAVersnelMAPlasmacytoid dendritic cells in autoimmune diabetes – potential tools for immunotherapyImmunobiology20092149–10 791 79919628297
- KimNSMbongueJCNicholasDAChimeric vaccine stimulation of human dendritic cell indoleamine 2, 3-dioxygenase occurs via the non-canonical NF-kB pathwayPLoS One2016112 e014750926881431
- MukhopadhayaAHanafusaTJarchumISelective delivery of cell antigen to dendritic cells in vivo leads to deletion and tolerance of autoreactive CD8+ T cells in NOD miceProc Natl Acad Sci U S A200810517 6374 637918430797
- EiblNSpatzMFischerGFImpaired primary immune response in type-1 diabetes: results from a controlled vaccination studyClin Immunol20021033 Pt 1 249 25912173299
- GiannoukakisNPhase I (safety) study of autologous tolerogenic dendritic cells in type 1 diabetic patientsDiabetes Care2011349 2026 203221680720
- HuoWLiGHQiRQClinical and immunologic results of local hyperthermia at 44°C for extensive genital warts in patients with diabetes mellitusInt J Hyperthermia2013291 17 2023311375
- GottliebPAAlkananiAKMichelsAWα1-Antitrypsin therapy downregulates toll-like receptor-induced IL-1β responses in monocytes and myeloid dendritic cells and may improve islet function in recently diagnosed patients with type 1 diabetesJ Clin Endocrinol Metab2014998 E1418 E142624527714
- OrbanTBeamCAXuPReduction in CD4 central memory T-cell subset in costimulation modulator abatacept-treated patients with recent-onset type 1 diabetes is associated with slower C-peptide declineDiabetes20146310 3449 345724834977
- GiannoukakisNPhillipsBFinegoldDHarnahaJTruccoMPhase I (safety) study of autologous tolerogenic dendritic cells in type 1 diabetic patientsDiabetes Care2011349 2026 203221680720
- HarnahaJMachenJWrightMInterleukin-7 is a survival factor for CD4+ CD25+ T-cells and is expressed by diabetes-suppressive dendritic cellsDiabetes2006551 158 17016380489