
Abstract
Degenerative fundus disease encompasses a spectrum of ocular diseases, including diabetic retinopathy (DR) and age-related macular degeneration (AMD), which are major contributors to visual impairment and blindness worldwide. The development and implementation of effective strategies for managing and preventing the onset and progression of these diseases are crucial for preserving patients’ visual acuity. Melatonin, a neurohormone primarily produced by the pineal gland, exhibits properties such as circadian rhythm modulation, antioxidant activity, anti-inflammatory effects, and neuroprotection within the ocular environment. Furthermore, melatonin has been shown to suppress neovascularization and reduce vascular leakage, both of which are critical in the pathogenesis of degenerative fundus lesions. Consequently, melatonin emerges as a promising therapeutic candidate for degenerative ocular diseases. This review provides a comprehensive overview of melatonin synthesis, its localization within ocular tissues, and its mechanisms of action, particularly in regulating melatonin production, thereby underscoring its potential as a therapeutic agent for degenerative fundus diseases.
Introduction
Fundus degeneration encompasses a spectrum of ocular conditions, notably age-related macular degeneration (AMD) and diabetic retinopathy (DR).Citation1 In developed nations, AMD stands as the primary cause of legal blindness in individuals aged 60 and above, particularly among those of Caucasian descent.Citation2 Projections suggest that the number of diagnosed AMD cases will rise to 196 million by 2020 and 288 million by 2040.Citation3 The etiology of AMD is complex and not yet fully elucidated, although certain risk factors have been identified as potential contributors to the development of the condition.Citation4 Smoking and sunlight exposure are known to worsen environmental risk factors, with age being identified as the primary demographic risk factor. Additionally, research indicates that nutritional factors, gut microbiota, hypertension, and genetic variability play a role in the development of wet AMD.Citation5 Advanced wet AMD can lead to severe visual impairment due to choroidal neovascularization.Citation6
DR is associated with various microvascular complications, including blindness, particularly in individuals under the age of 40.Citation7 The estimated prevalence of diabetes is 400 million individuals, with one third experiencing DR and one tenth affected by vision-threatening conditions such as diabetic macular edema and proliferative DR (PDR).Citation8 Worldwide, DR and AMD are the major causes of vision impairment and blindness.Citation9 Given the increasing prevalence of diabetes and aging populations, the prevention and treatment of fundus degenerative diseases are of heightened importance.
Melatonin, a neurohormone predominantly produced by the pineal gland, has been the subject of extensive research and application across various disciplines.Citation10 Eye research has particularly focused on the interaction between melatonin and the eyes, especially retinal ganglion cells. Additionally, studies have explored the impact of eye lesions on the circadian rhythm regulation of melatonin.Citation11 Besides its role in regulating circadian rhythms, melatonin serves crucial physiological functions in ocular structures. Evidence suggests that a deficiency in melatonin may contribute to the development of AMD, as indicated by reduced melatonin production in AMD patients compared to controls.Citation12 Furthermore, individuals with AMD exhibit elevated melatonin levels during daytime hours, suggesting a disruption in melatonin’s diurnal cycle in individuals with this condition.Citation13 Consequently, heightened levels of melatonin during daylight hours may have deleterious effects on ocular structures, potentially leading to the onset of light-induced retinal degeneration.
Research has shown that melatonin suppresses the phosphoinositide 3-kinase (PI3K)/ protein kinase B (Akt)/signal transducer and activator of transcription 3 (STAT3)/nuclear factor-kappaB (NF-κB) signaling pathway in experimental DR, thereby preserving the integrity of the blood-retinal barrier.Citation14 Genome-wide association studies have revealed that variations in genes encoding melatonin synthase and receptors are implicated in the pathogenesis of type 2 diabetes.Citation15,Citation16 It has been hypothesized that melatonin may play a role in various forms of retinopathy; however, population-based studies have not investigated these potential associations. Melatonin exerts its effects through multiple mechanisms, such as antioxidant, anti-inflammatory, and neuroprotective actions. It scavenges free radicals, diminishes the release of inflammatory molecules, and shields ocular tissues from oxidative stress and inflammatory harm in AMD and DR.Citation17,Citation18 Furthermore, melatonin inhibits neovascularization and vascular leakage, regulating the development of fundus degenerative lesions.Citation19
Studies have found that exogenous melatonin is safe in the human body and has a high ability to cross the blood-retinal barrier.Citation20,Citation21 Therefore, melatonin may become a potential novel therapy in degenerative eye diseases. Interestingly, research has found that the synthesis of melatonin in the retina decreases with age, further suggesting that melatonin may be involved in age-related pathologies.Citation22
Melatonin Biosynthesis and Physiological Functions
Mechanisms of Melatonin Synthesis and Release
Melatonin, a neurohormone primarily synthesized by the pineal gland, undergoes complex regulatory mechanisms during its synthesis and release.Citation23 Various enzymes, hormones, and regulatory factors are involved in this process, and once produced, melatonin diffuses freely out of the cell.Citation24,Citation25 Understanding the processes involved in the production and secretion of melatonin is crucial to uncovering its significance in fundus degeneration.
Melatonin is synthesized from tryptophan, an essential amino acid, through a series of enzymatic reactions, among which tryptophan hydroxylase is involved.Citation26 In the following step, 5-hydroxytryptophan is converted into 5-hydroxytryptophan via the hydroxytryptophan decarboxylase. The cytochrome P450 enzyme is also involved in the regulation of the conversion to 5-hydroxytryptamine.Citation27 Melatonin is eventually converted from 5-hydroxytryptophan inside the cell through a variety of enzyme-catalyzed modifications.Citation28 Key enzymes in this pathway include the melatonin N-acetyltransferase (AANAT) and the hydroxytryptamine-O-methyltransferase (HIOMT).Citation29 The enzyme AANAT catalyzes the transfer of an acetyl group to serotonin, forming N-acetylserotonin, which is then methylated by HIOMT to produce melatonin.Citation30
The Regulation of Melatonin Synthesis and Release
Light primarily regulates melatonin synthesis and release through the retino-pineal pathway.Citation31 Light signals are transmitted from the retina, the light-sensitive tissue of the eye, to the pineal gland.Citation32 Retinoid molecules in photoreceptor cells trigger a series of biochemical reactions that ultimately lead to neural signals. As these signals travel through the optic nerve to the pineal gland, the pineal gland modulates melatonin synthesis and release by releasing neurotransmitters and activating melatonin receptors. During daytime light exposure, neurotransmitters released from the retina inhibit the activity of melatonin synthase and AANAT in the pineal gland, decreasing melatonin production.Citation33 In contrast, in darkness or at night, the absence of light signaling reduces the release of retinal neurotransmitters, increasing the activity of melatonin synthase and AANAT, thus promoting melatonin synthesis and release.Citation34
In addition to light, melatonin synthesis and release are influenced by endogenous factors, involving various systems such as the nervous, endocrine, and immune systems.Citation35 Neurotransmitters in the nervous system, including dopamine, norepinephrine, and epinephrine, can inhibit melatonin synthesis and release, whereas neuropeptides such as oxytocin can promote it.Citation36 Hormones within the endocrine system, such as cortisol, thyroid hormones, and sex hormones, also affect melatonin synthesis by binding to receptors in pineal cells and regulating intracellular signaling pathways.Citation37 Additionally, immune system cytokines and inflammatory responses can influence melatonin production and release. Cytokines involved in inflammatory processes, such as tumor necrosis factor-α (TNF-α) and interleukin-1β (IL-1β), have been shown to inhibit melatonin synthesis and release.Citation38
The pineal gland plays a crucial role as an endogenous regulator in the synthesis and release of melatonin. Pineals are small brain glands that are primarily responsible for melatonin synthesis and release.Citation39 Various hormones, such as β-adrenoceptors and cholinergic neurons, control pineal gland activity.Citation40 Adrenaline suppresses melatonin synthesis, while cholinergic neurons stimulate it.Citation41 The synthesis of melatonin in the pineal gland is regulated by the daily rhythm of β-adrenergic receptor stimulation, which initiates the transcription of AANAT genes and activates the corresponding enzymes.Citation42 Melatonin demonstrates a protective and enhancing influence on cholinergic neurons responsible for the regulation of circadian rhythm and sleep. However, a definitive correlation between these two systems has not been established.Citation43 These hormones and neuromodulators regulate intracellular signaling pathways by binding to receptors on the surface of pineal gland cells, thereby affecting melatonin synthesis and release.
Melatonin receptors have been identified in many retinal neurons, including intrinsic photosensitive retinal ganglion cells (ipRGCs) containing melanopsin, suggesting that melatonin modulates the physiological function of these photoreceptors in the inner layers of the retina.Citation44 ipRGCs are located in the inner layers of the retina are responsible for circadian regulation, and are directly stimulated by light through the activation of the photoreceptor pigment melanopsin, which projects to the supraoptic nucleus of the optic cross, and eventually projects to the pineal gland and controls the release of the sleep hormone melatonin.Citation45
In summary, the production and secretion of melatonin is an intricate and carefully controlled process. It is regulated by several factors, including enzymes in the synthesis pathway, pineal hormone regulation, light exposure, and endogenous regulation of the nervous, endocrine, and immune systems. An in-depth understanding of the mechanisms of melatonin synthesis and release is important for revealing its role and therapeutic potential in fundus degenerative diseases.
The Presence and Role of Melatonin in the Structure of the Eye
Melatonin is prevalent in ocular structures and plays a critical role in several physiological processes. Below are the key aspects of melatonin’s presence and function in ocular tissues.
Melatonin in the Retina
Melatonin is synthesized in retinal photoreceptors and acts as a local neuromodulator.Citation46 In addition to endogenous production, melatonin can be transported into ocular tissues via the bloodstream. Melatonin receptors have been identified in multiple retinal structures, including retinal pigment epithelium (RPE), photoreceptors, horizontal cells, bipolar cells, amacrine cells, ganglion cells, and the inner and outer plexiform layers.Citation47 The melatonin hormone has paracrine and autocrine impacts that are controlled by its G-protein-coupled receptors, melatonin receptor type 1 (MT1) and MT2, as well as potentially MT3, or can occur without receptor involvement.Citation48 It is currently believed that it is the MT1 and MT2 receptors that play a major role in the eye.Citation49 The retinal cells with receptors play several crucial roles, such as absorbing light that is scattered, preserving the well-being of photoreceptor cells in the retina, and offering nutritional assistance.Citation50 Melatonin is present in the eye’s retina, with a high concentration in the RPE.Citation51 Melatonin is released in the dark and dopamine in the light within the eye, with melatonin suppressing the release of dopamine from anaplastic cells.Citation52 MT1 receptors, present on dopaminergic and GABAergic amacrine cells in guinea pigs, humans, and rhesus monkeys, have the potential to induce this effect.Citation53 Furthermore, exposure to light stimulates the release of dopamine from anaphase cells, which bind to D2 and D4 receptors on photoreceptors, thereby inhibiting the activity of AANAT and modulating cAMP and calcium concentrations, ultimately leading to a reduction in melatonin production.Citation54 Therefore, the widespread distribution of melatonin receptors in the retina suggests an intricate interplay between neuronal processes involving melatonin, dopamine, and melanopsin.
Research indicates that melatonin supports retinal health by controlling RPE cell activity, reducing oxidative stress, and protecting against damage.Citation55 Knockout studies in mice have shown that defects in MT receptors lead to lipofuscin accumulation and decreased cell survival.Citation56 Melatonin also influences phototransduction and modulates photoreceptor and bipolar cell sensitivity.Citation57 Additionally, it mitigates cell death and inflammation in RPE cells, thereby maintaining retinal function.Citation58
Melatonin in the Sclera and Uvea
Melatonin is present in the uvea, where it plays several physiological roles. It regulates ocular blood flow by modulating vasoconstriction and vasodilation and exhibits anti-neovascularization properties, inhibiting endothelial cell proliferation and migration.Citation59–61 Melatonin also reduces ocular inflammation by modulating immune responses and decreasing cytokine levels and inflammatory mediator production.Citation62 Melatonin receptors are expressed in the sclera, which may contribute to the maintenance of scleral elasticity.Citation63 Melatonin receptors in the sclera may be involved in the progression of ocular axial length due to myopia in adolescents. As a result of the blue light (480 nm) interacting with the retina and sclera, melanopsin synthesis is specifically stimulated and melatonin MT1 receptor expression and melatonin concentration were decreased.Citation64 The findings are consistent with those of a study in guinea pigs in which melatonin protease was inhibited by an antagonist administered intravitreally.Citation65 This melatonin protease inhibition leads to an abnormal increase in eye growth, which is directly related to increased melatonin levels in the retina.
Melatonin in the Lens and Vitreous
Melatonin is also found in the lens, where it contributes to lens transparency and health.Citation66 It was once thought that melatonin reached the lens via atrial fluid absorption, but recent studies suggest that it is also synthesized by human lens epithelial cells. Light activates melanopsin, reducing melatonin synthesis. Likewise, darkness increases AANAT expression and melatonin production in lens epithelial cells.Citation67 Lenses are transparent tissues inside the eyes used for focusing light.Citation68 Oxidative stress and inflammatory responses affect lens clarity and function. Melatonin, which has antioxidant and anti-inflammatory effects in the lens, can maintain lens health by inhibiting the production of free radicals and reducing oxidative stress, protecting lens cells from oxidative stress damage.Citation69 The lens is unable to avoid UV exposure, and after UV exposure, exogenously administered melatonin interventions result in decreased lipid peroxidation, increased antioxidant enzyme activity, and decreased CA2+, thereby reducing cataract formation.Citation70 Furthermore, melatonin reduces inflammatory damage to lens cells by modulating the inflammation response and inhibiting the production of inflammatory mediators.Citation71 Located between the lens and retina, vitreous humor is a clear gel-like substance that maintains the shape of the eye and supports the retina. There is no melatonin production in the vitreous, it is generally secreted and delivered to the vitreous and has been shown to be effective in rescuing photoreceptor degeneration in experimental studies.Citation72
Melatonin in the Ciliary Body
The unpigmented ciliary epithelium is also thought to provide melatonin, and the fact that AANAT and HIOMT are localized to the human ocular ciliary body supports a ciliary source for melatonin.Citation73 Researchers have identified melatonin receptors in rabbit iris and ciliary body, as well as melatonin metabolites in the ciliary bulge Melatonin receptors have been identified in the iris and ciliary body of rabbits, as well as localized to the ciliary epithelium.Citation74,Citation75 The ciliary body secretes melatonin, which is responsible for aqueous humor production and maintaining intraocular pressure variations throughout the day.Citation76 Melatonin is highest at night, when intraocular pressure (IOP) is generally lowest; therefore, melatonin may be negatively correlated with IOP. Moreover, topical administration of melatonin analogs has been shown to lower IOP, thereby altering the circadian pattern of IOP.Citation77 Currently, it is believed that the mechanism of action is to reduce chloride ion efflux, which in turn reduces the formation of aqueous humor caused by fluid movement in the unpigmented ciliary body.Citation78 It is possible that melatonin plays a role in the pathophysiology and treatment of glaucoma because of its association with an increase in aqueous humor production and intraocular pressure.Citation79 Glaucoma-induced damage to the inner retina layer can impact retinal ganglion cells, potentially causing disruptions in melatonin levels and contributing to the prevalence of sleep disorders among individuals with glaucoma.Citation80,Citation81 Melatonin, on the other hand, has the potential to be a new treatment option for glaucoma if it has both IOP-lowering and neuroprotective effects.
Melatonin in the Cornea
The distribution of melatonin receptors is also detected in the cornea, which is mainly localized in the epithelium, stroma, and endothelium of the rabbit cornea.Citation82 Both rabbits and humans have melatonin in their tears. Various potential impacts of melatonin on the eye surface have been observed. Melatonin facilitates the shedding of epithelial cells by controlling the development of tight junctions and the detachment of surface cells.Citation83 Melatonin has been shown in in vitro study to affect corneal hydration and corneal wound healing.Citation84 Furthermore, melatonin may play a part in the growth of corneal epithelial cells.Citation85 Melatonin has also been shown to have antioxidant properties that protect corneal epithelial cells from ultraviolet (UV) irradiation damage, and melatonin-treated rabbit corneal epithelial cells showed a decrease in lipid peroxidation and an increase in mitochondrial viability after UV exposure compared to untreated control tissue.Citation86
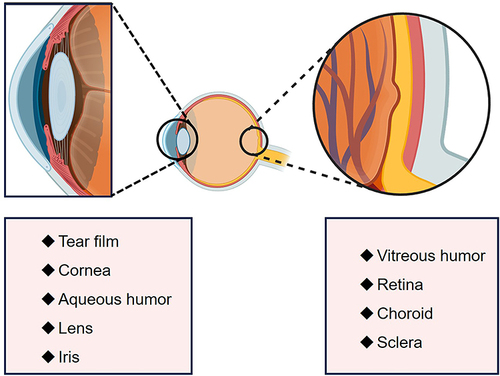
In Conclusion, melatonin plays a significant role in various physiological functions within the eye, including phototransduction and photoreceptor renewal, aqueous humor production regulation, intraocular pressure, wound healing, and providing antioxidant protection (). Melatonin receptors are widely distributed across ocular tissues, enabling diverse responses to melatonin.Citation87 Understanding melatonin’s synthesis and effects in ocular tissues highlights its importance in maintaining ocular health and its potential therapeutic applications in treating ocular diseases.
Figure 1 The distribution of melatonin in various tissues of the eye.
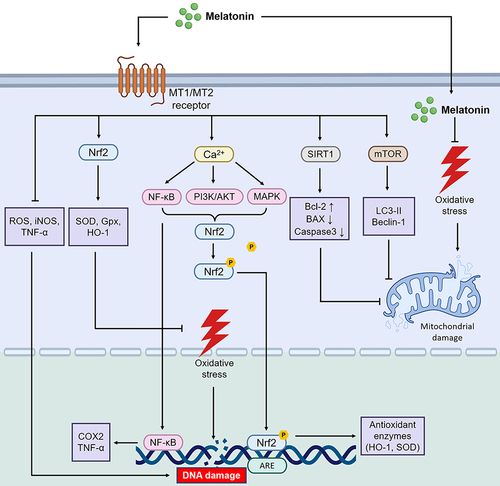
Melatonin Exerts Antioxidant, Anti-Inflammatory, Cytoprotective, Cell-Cycle-Regulating and Proliferative Effects in Degenerative Fundus Lesions
Antioxidant Effects
Melatonin exerts significant antioxidant and anti-inflammatory effects in degenerative diseases of the fundus, playing a crucial role in maintaining ocular tissue health and mitigating lesion progression Oxidative stress is a primary factor in the development of degenerative fundus lesions, causing irreversible damage to cellular and tissue structures. Melatonin plays a neuroprotective role by activating its receptor MT1 to inhibit the expression of amyloid precursor protein processing enzyme in the hyperglycemia induced diabetes cell model.Citation88 Additionally, melatonin inhibits mTOR and autophagy markers (LC3-II and Beclin-1) through its MT2 receptors, promoting autophagy to reduce retinal oxidative stress.Citation89 It also modulates the expression of antioxidant enzymes and neuronal-type nitric oxide synthase (nNOS), while triggering the activation of inducible nitric oxide synthase (inNOS).Citation90,Citation91
Melatonin serves as a mitochondria-targeted antioxidant, protecting retinal cells from oxidative stress.Citation92 It enhances the survival of retinal cells, including photoreceptors, retinal ganglion cells, and Müller cells, which are affected by DR.Citation93 In diabetic retinas, melatonin prevents nitro-oxidative stress and the inflammatory response induced by high glucose levels.Citation94 Studies have shown that melatonin synthesis is reduced in the retinas of diabetic rats, correlating with the progression of DR.Citation95–97
In age-related macular degeneration (AMD), reduced nocturnal melatonin production contributes to oxidative damage, highlighting the importance of melatonin’s antioxidant effects.Citation98,Citation99 Melatonin regulates antioxidant gene transcription via nuclear factor erythroid 2–related factor 2 (Nrf2), which is compromised in aging RPE cells.Citation100,Citation101 Exogenous melatonin may thus play a role in preventing and treating AMD by modulating oxidative stress and Nrf2-mediated defenses.Citation102 Additionally, in fundus oxidative stress injury, reactive oxygen species (ROS) can facilitate telomere erosion, thereby contributing to the senescence of RPE cells.Citation103 The retina contains active telomerase, and defects in telomerase activity in AMD can also lead to RPE cell damage; melatonin, which regulates telomerase activity, may play a role in slowing the damage.Citation104 Melatonin appears to modulate telomerase, suggesting it may be useful to prevent or treat AMD.Citation105 Since melatonin is an antioxidant, a mitochondrial protector, and an inflammatory modulator, its deficiency may promote the progression of DR and AMD lesions. Further clinical trials are necessary to evaluate its efficacy and safety.
Melatonin’s direct antioxidant effects should not be overlooked.Citation106 It neutralizes free radicals, reducing oxidative stress and protecting eye tissues from damage.Citation107–109 It also enhances mitochondrial function and intracellular antioxidant capacity by boosting the activity of enzymes like superoxide dismutase and glutathione peroxidase.Citation110 Melatonin has demonstrated efficacy in protecting mitochondrial DNA from oxidative damage, although it did not confer protection against ischemia/reperfusion-induced retinal damage.
Melatonin demonstrated efficacy in safeguarding the mitochondrial DNA of ARPE-19 cells against damage induced by hydrogen peroxide.Citation111 Ischemia/reperfusion is an important source of free radical production, and subcutaneous injection of melatonin can provide protection against retinal damage caused by ischemia/reperfusion in many rats.Citation112 In autopsy studies of AMD-affected eyes, mitochondrial DNA was found to be excessively damaged by oxidative stress.Citation113 Furthermore, melatonin’s antioxidant properties are expressed in its ability to reduce oxidative damage to DNA.Citation114 Inflammation can also be triggered by mitochondrial signaling pathways. Melatonin attenuates mitochondrial damage and oxidative stress generated during inflammation by modulating mitochondrial function and the activity of the mitochondrial respiratory chain. Regulation of the mitochondrial pathway reduces the production and release of inflammatory mediators, thereby attenuating the inflammatory response and cellular damage. Melatonin reduces ocular inflammatory responses and damage by regulating the production and release of cytokines and maintaining the balance of cytokines and inflammatory mediators.Citation115 Thus, the involvement of melatonin in mitochondrial regulation may be one of the ways to address AMD fundus lesions. It is predicted that mitochondria may play a significant role in melatonin synthesis, which supports the relationship between melatonin and mitochondria.Citation116
Anti-Inflammatory Effects
Inflammatory responses play an important role in the development of degenerative fundus lesions, leading to tissue damage and apoptosis. Moreover, the inflammatory process plays a significant role in the pathogenesis of DR, rendering it a low-grade chronic inflammatory condition. Upregulation of inflammatory mediators including nNOS, Cyclooxygenase-2, vascular endothelial growth factor (VEGF), mitogen-activated protein kinases (MAPK) and NF-κB is observed in DR, with NF-κB activation leading to cytokine release and increased production of ROS.Citation117 A significant part of melatonin’s anti-inflammatory effect is its ability to reduce the inflammatory response and inflammatory mediator production.Citation118 There are several mechanisms by which melatonin exerts its anti-inflammatory effects. First, it inhibits the production and release of inflammatory mediators, such as cytokines and inflammatory mediators. Specifically, melatonin suppresses the synthesis of inflammatory cytokines, including (TNF-α) and interleukin-6, leading to a decrease in inflammatory response.Citation119 Second, melatonin also regulates inflammatory signaling pathways such as NF-κB and mitochondrial pathways.Citation120 NF-κB is a key transcription factor involved in the regulation of inflammatory responses and apoptosis, among other processes. Melatonin inhibits NF-κB activation through multiple mechanisms. It inhibits the nuclear translocation and nuclear transcriptional activity of NF-κB, thereby suppressing the expression of inflammation-related genes.Citation121 As well as blocking NF-κB activation signaling, melatonin inhibits the activity of IKK receptors.Citation122 Furthermore, melatonin regulates cytokine signaling pathways, including the Janus kinase/signal transducer and activator of transcription (JAK/STAT) family pathway, impacting immune cell function and the development of inflammatory responses.Citation123
These signaling pathways play an important role in the inflammatory response, and melatonin inhibits the onset and progression of the inflammatory response by regulating their activity.Citation124 In addition, melatonin reduces inflammation-induced apoptosis and cellular damage. It protects eye tissues from inflammation-induced damage by inhibiting oxidative stress and regulating the expression of proteins associated with cell survival and apoptosis.
Regulates Cell Activity and Exerts Cytoprotective and Anti-Apoptotic Effects
Melatonin also affects the activity of immune cells, regulates the immune response and inflammatory processes, and modulates immune cells such as macrophages and T-lymphocytes that play an important role in fundus degeneration.Citation125 In dendritic cells, melatonin influences immune function via the Nrf2/hemoglobin oxygenase-1 (HO-1) axis.Citation126 Melatonin regulates dendritic cells to modulate the body’s immune system, but further basic and clinical studies are needed to confirm this in the field of ophthalmology.Citation127 Melatonin was effective in ameliorating retinal degeneration in a mouse model, and the 5’-nucleotidase ecto/Tet methylcytosine dioxygenase 2 (NT5E/TET2) pathway in Tregs may be the mechanism of the anti-retinal degeneration response to melatonin treatment, which was first demonstrated to regulate immune homeostasis through Tregs.Citation128 Melatonin also affects the proliferation and differentiation of immune cells and regulates their function. By affecting the function of immune cells, it reduces the aggregation and activation of inflammatory cells and decreases the secretion and release of inflammatory mediators. In addition, melatonin can inhibit the migration of immune cells and reduce the inflammatory response and damage to ocular tissues.Citation129 In addition, melatonin acts as a cytoprotectant, reducing apoptosis and damage to eye tissue.Citation130 The apoptotic pathway is inhibited by melatonin, thereby increasing cell survival and functional recovery. According to a study, melatonin inhibits the expression of apoptosis-related proteins, including members of the Bcl-2 family and cysteine protease family.Citation131 The expression and activity of these proteins play an important role in apoptosis, and melatonin inhibits it in ocular tissues. The action of melatonin interferes with cell survival signaling pathways, thereby regulating the balance between cell survival and apoptosis. A study showed that melatonin can promote cell survival and proliferation by activating the PI3K/Akt pathway.Citation132 Furthermore, in terms of neuroprotection, melatonin treatment increased levels of brain-derived neurotrophic factor (BNDF) and its downstream phosphorylated tropomyosin receptor kinase B (Trkb)/Akt/extracellular regulated protein kinase (ERK)/p-cAMP response element binding protein (CREB) levels, and hypoxia-ischemia-induced increases in cleaved caspase-3 and Bax protein levels, as well as reductions in Bcl-2 protein levels, were reduced.Citation133 Besides, activation of the NF-κB signaling pathway triggered apoptosis of retinal capillary cells in the DR, increasing the production of ROS and inflammatory factors.Citation134 Melatonin regulates dendritic cells to modulate the body’s immune system, but further basic and clinical studies are needed to confirm this in the field of ophthalmology.Citation135 Melatonin induces the expression of SIRT1 gene to reduce the expression of BAX and caspase 3, increase the expression of Bcl2, and play a role in reducing mitochondrial and nuclear DNA damage.Citation136 Melatonin exerts its cytoprotective effects against DR and AMD through these cytoprotective and anti-apoptotic mechanisms, attenuating cellular damage and inflammatory responses in ocular tissues.
In summary, melatonin’s mechanism of action in degenerative fundus diseases involves its antioxidant, anti-inflammatory, cytoprotective, cell proliferation regulator and anti-apoptosis (). A comprehensive understanding melatonin’s role in fundus degenerative disease is crucial for revealing its therapeutic potential and developing appropriate therapeutic strategies.
Figure 2 Melatonin plays a role in antioxidant, anti-inflammatory, and reducing cell apoptosis in degenerative fundus lesions.
Vasomodulation of Melatonin in Degenerative Fundus Lesions
Abnormal neovascularization is an important feature in fundus degenerative diseases such as AMD and DR. Melatonin exerts a role in reducing neoangiogenesis, decreasing vascular damage due to inflammatory responses in vascular tissue and regulating vasoconstriction (). Melatonin regulates retinal VEGF secretion, promoting physiologic secretion to protect the retina from oxidative stress and reducing pathologic secretion to inhibit neovascularization.Citation137 Inhibition of proliferation and migration of vascular endothelial cells is one way in which melatonin inhibits neovascularization.Citation138 In addition, melatonin inhibits the production and release of VEGF and matrix metalloproteinase (MMP) in vascular endothelial cells, which are involved in regulating the process of neovascularization.Citation139,Citation140 VEGF is a major cause of neovascularization, and melatonin protects the retina from oxidative stress and hemorrhage by regulating VEGF secretion in the retina and reducing pathological secretion to inhibit neovascularization.Citation141 The antioxidant and antiangiogenic characteristics of melatonin position it as a promising therapeutic option for addressing AMD and DR. The diminished synthesis of melatonin in the retinas of DR rats, in comparison to control subjects, may be linked to the advancement of DR fundus lesions.Citation142 As well, serum melatonin concentrations are reduced at night in diabetics with PDR.Citation97 Furthermore, studies have shown that melatonin exhibits a protective effect against pathological retinal neovascularization, preservation of glial cells, and mitigation of inflammatory response through the inhibition of the HIF-1α/VEGF signaling pathway.Citation143 Additionally, melatonin has been found to suppress the activity of angiopoietin receptors, crucial components of angiogenesis-related signaling pathways that contribute significantly to angiogenesis and tissue remodeling.Citation137 Melatonin has been shown to suppress RhoA/ROCK signaling in mouse models of choroidal neovascularization, leading to a transition of macrophages and microglia from M2 to M1 phenotypes following prolonged laser exposure.Citation144 These findings indicate the potential for melatonin intervention as a novel therapeutic approach for AMD and DR. Through the regulation of key molecules, melatonin can impact the proliferation, migration, and lumen formation of vascular endothelial cells, ultimately influencing vascular structure and function.
Figure 3 The vascular regulatory role of melatonin in retinal degenerative diseases.
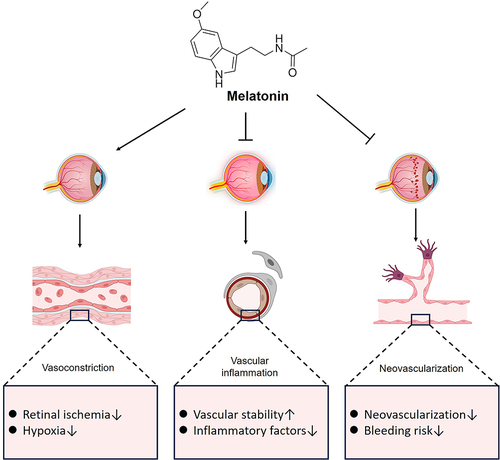
In regulating vasoconstriction, melatonin has been shown to have different effects on blood vessels and blood flow in different organs and tissues of the body. Mechanisms of melatonin action on the vasculature have been shown to include direct activation of melatonin receptors and through intercellular pathways.Citation145,Citation146 The prevalence of MT1 receptors in retinal vascular endothelial cells suggests that melatonin binding to these receptors on vascular smooth muscle cells can induce vasoconstriction through the enhancement of norepinephrine signaling.Citation147 The presence of MT1 has been identified in the outer membranes of retinal arteries and veins, particularly in the papillary region rather than in the ciliary and choroidal vessels.Citation148 The localization of MT1 in retinal vessels to the outer membrane of the vessel wall aligns with previous observations in human cerebral vessels.Citation149 The primary role of MT1 receptors is thought to be mediating vasoconstriction.Citation150 Therefore, we hypothesized that melatonin may act in an indirect manner on retinal vascular smooth muscle to regulate contraction. And there is also a study demonstrating that the onset of AMD may lead to smaller diameters of small blood vessels.Citation151 In addition, patients with DR also show narrowing changes in the diameter of the lumen of the retinal blood vessels in the fundus, resulting in decreased blood flow to the fundus.Citation152 Thus, the possible vasoconstrictive effect of melatonin on the fundus has the potential to exacerbate fundus changes in patients at an early stage. In spite of melatonin’s ability to modulate vasoconstriction, its effect on blood pressure appears to be dose-dependent. A reduction in arterial blood pressure was observed in subject men and women with 1 mg of melatonin.Citation153 However, in another study it was found that ingesting 3mg of melatonin did not change the subjects’ blood pressure.Citation154 In order to better understand the relationship between melatonin levels and vascular changes, adrenal nerve activity, and hormonal responses, additional studies are needed for various doses of the drug.Citation146,Citation155 Melatonin’s protective effects on retinal inflammation, oxidative stress, angiogenesis, and apoptosis have been demonstrated in animal models, suggesting that melatonin may help prevent diabetic-related retinal damage.Citation156 However, due to a lack of clinical studies, it is still unknown whether melatonin has clinical benefits, despite having been taken as a supplement by a relatively large number of people worldwide.
Melatonin for Stem Cell Therapy in Fundus Degenerative Disease
Melatonin has demonstrated effective application in various diseases when used in pretreatment or in combination with scaffolds including chronic kidney disease, neurodegenerative diseases, and orthopedic disorders, whereas its use in ocular diseases is rare.Citation157,Citation158 It has been widely demonstrated that stem cells are effective in treating eye diseases.Citation159 Melatonin promotes the proliferation, differentiation and survival of stem cells and improves their effectiveness in therapy.Citation160 For treatment of retinal diseases and vision restoration, reprogramming endogenous neural stem cells is considered to be the most promising approach.Citation161 In vitro, melatonin inhibits cell apoptosis pathways and regulates DNA methylation to promote the survival of endogenous neural stem cells derived from bovine retina.Citation162 The proliferation of pluripotent stem cells induced by endogenous neural stem cells can be further enhanced through activation of the ERK1/2 signaling pathway downstream of melatonin. Additionally, melatonin has been shown to significantly increase retinal neural stem cell proliferation and expression of the marker nestin via the ERK and TGF-β/Smad pathway-mediated MT1.Citation163
Moreover, melatonin has been shown to impact signaling pathways and gene expression in stem cells, modulate the equilibrium between cell proliferation and differentiation, and serve as an antioxidant to mitigate oxidative stress-induced apoptosis and augment stem cell functionality.Citation164,Citation165 However, the specific role of melatonin in the regulation of corneal epithelial stem cells and its efficacy in the treatment of ocular diseases in animal models have yet to be elucidated. Given the demonstrated superiority and significant potential of melatonin in stem cell therapy, it is imperative to prioritize further investigation into its therapeutic modalities, effects, and mechanisms. Additionally, preliminary small-scale clinical trials in patients may be warranted to assess the potential for melatonin’s clinical application to enhance therapeutic outcomes, promote improvements in visual acuity, and facilitate the repair of fundus tissues.
Melatonin Dosing Patterns
Clinical trials have focused on oral melatonin administration, with early results showing protection of the retina and slow progression of AMD when administered daily (3 mg).Citation166 In recent studies, however, it has been demonstrated that due to the liver’s first pass metabolism, its bioavailability is only about 15%.Citation167 Whereas the efficacy of melatonin varies by dosage form, with continuous-release and absorbed forms being more efficacious than immediate-release forms, the short half-life and ultra-high maximal plasma concentration may further contribute to the low bioavailability of melatonin due to under-absorption and tolerability issues.Citation168 Ocular surface drug delivery is convenient and safe, and is currently the most common route of administration in the treatment of ophthalmic diseases. The ophthalmic administration of melatonin has been widely used in animal experiments, and nano micelle formulation eye drops can penetrate into the eyes of rats.Citation169 However, if clinical application is required, further research is needed on the concentration, safety, and bioavailability of drugs entering the human eye. The use of vitreous cavity injections, which circumvent the ocular barrier and facilitate the delivery of drugs to the targeted treatment site, is currently under observational investigation for the treatment of retinal diseases. Nevertheless, a prior study documented the occurrence of adverse effects, specifically retinal cell degeneration, following high-dose melatonin injections.Citation170 Hence, it is imperative to conduct further research on the mechanism of action and potential toxicity of melatonin in the treatment of ocular diseases, as well as to establish the optimal therapeutic dosage for varying ocular conditions and individual patients. Additionally, the optimal timing of melatonin administration and the therapeutic efficacy of alternative routes of delivery, such as subconjunctival injection, warrant exploration through extensive clinical trials.
Efforts to enhance melatonin’s efficacy in treating degenerative fundus lesions focus on developing drug delivery systems that sustain release, increase local tissue drug concentrations, improve therapeutic efficacy, and reduce side effects. The delivery of ocular drugs using nanotechnology can penetrate the surface of the eye and provide sustained release. Nanocarriers can be classified into four distinct categories according to their geometries: 0D-like nanoparticles, 1D-like nanofibers, 2D-like nanomembranes, and 3D-like nanogels.Citation171 Research has demonstrated that the encapsulation of melatonin within 0D and 1D nanocarriers enhances both bioavailability and therapeutic effectiveness.Citation172 Melatonin-loaded PLGA-PEG nanoparticles synthesized by solvent substitution method can also reduce IOP twice as long as melatonin aqueous solution, and the results show good tolerance.Citation173 Moreover, melatonin is delivered through cationic and mucus carriers, which enhance its ability to cross the ocular surface barrier and prolong its retention.Citation174 A recent study discovered that melatonin solution encapsulated within ethylcellulose nanoparticles exhibited enhanced corneal penetration attributed to its adhesion to mucin and slower release kinetics in comparison to free melatonin solution. This formulation significantly augmented retinal thickness and decreased apoptosis of retinal ganglion cells by approximately 16% in a retinal detachment model, indicating a superior retinoprotective effect.Citation175 Nevertheless, there remains a scarcity of advanced high-dimensional nanocarriers with high drug loading capacity and stimulus responsiveness for the loading of melatonin in ocular drug delivery, necessitating further development and exploration for potential clinical application. We learned that synergistic therapeutic effects based on melatonin, including co-administration strategies with glial cell-derived neurotrophic factors and neuroprotective drugs, have also been proposed, and the prognosis of co-administration may be better compared with single drugs. The incorporation of melatonin into multidrug systems results in improved sustained release of formulations, while advancements in drug delivery systems facilitate enhanced detection and monitoring. However, it is important to acknowledge potential challenges such as competition for drug delivery sites and the formation of drug co-crystals.Citation176 Therefore, despite the development of nano-loading technologies, mass production of melatonin nanomedicines for ophthalmic applications remains challenging and clinical trials are still lacking. In addition to short-term clinical studies, it is important to evaluate the long-term efficacy and safety of the application of various dosage forms of melatonin in the treatment of fundus degenerative diseases.
Conclusion
Melatonin exhibits diverse mechanisms of action and therapeutic potential in managing fundus degenerative diseases (). Clinical and laboratory research underscores its role in modulating vasomotor function, maintaining fundus vasculature, and mitigating complications like fundus hemorrhage through anti-angiogenic and anti-inflammatory effects. Melatonin’s antioxidant properties neutralize free radicals, reduce oxidative stress, and protect retinal cells from apoptosis, promoting cell survival, repair, and normal turnover in retinal tissue. Despite these promising findings, further research is essential to optimize melatonin's clinical application. Long-term studies are crucial for evaluating its efficacy, safety, and individualized treatment approaches tailored to genetic predispositions, age, and health status. Comprehensive understanding of melatonin's molecular mechanisms in fundus degenerative diseases will inform clinical practice and guide development of optimized drug delivery systems. Continued exploration of melatonin’s therapeutic potential and refinement of delivery systems represent promising avenues for advancing treatment outcomes in ocular diseases.
Figure 4 Melatonin is primarily responsible for fundus degeneration.
Disclosure
The authors report no conflicts of interest in this work.
Additional information
Funding
References
- Chen X, Rong SS, Xu Q, et al. Diabetes mellitus and risk of age-related macular degeneration: a systematic review and meta-analysis. PLoS One. 2014;9(9):e108196. doi:10.1371/journal.pone.0108196
- Pascolini D, Mariotti SP, Pokharel GP, et al. 2002 global update of available data on visual impairment: a compilation of population-based prevalence studies. Ophthalmic Epidemiol. 2004;11(2):67–115. doi:10.1076/opep.11.2.67.28158
- Wong WL, Su X, Li X, et al. Global prevalence of age-related macular degeneration and disease burden projection for 2020 and 2040: a systematic review and meta-analysis. Lancet Glob Health. 2014;2(2):e106–116. doi:10.1016/s2214-109x(13)70145-1
- Heesterbeek TJ, Lorés-Motta L, Hoyng CB, Lechanteur YTE, den Hollander AI. Risk factors for progression of age-related macular degeneration. Ophthalmic Physiol Opt. 2020;40(2):140–170. doi:10.1111/opo.12675
- Zisimopoulos A, Klavdianou O, Theodossiadis P, Chatziralli I. The role of the microbiome in age-related macular degeneration: a review of the literature. Ophthalmologica. 2021;244(3):173–178. doi:10.1159/000515026
- Hernández-Zimbrón LF, Zamora-Alvarado R, Ochoa-de la paz L, et al. Age-related macular degeneration: new paradigms for treatment and management of AMD. Oxid Med Cell Longev. 2018;2018:8374647. doi:10.1155/2018/8374647
- Das A. Diabetic retinopathy: battling the global epidemic. Indian J Ophthalmol. 2016;64(1):2–3. doi:10.4103/0301-4738.178155
- Ting DS, Cheung GC, Wong TY. Diabetic retinopathy: global prevalence, major risk factors, screening practices and public health challenges: a review. Clin Exp Ophthalmol. 2016;44(4):260–277. doi:10.1111/ceo.12696
- Bourouki E, Dimitriou E, Chatzipantelis A, et al. Co-existence of age-related macular degeneration and diabetic retinopathy in a tertiary referral center in Greece. Cureus. 2022;14(11):e31051. doi:10.7759/cureus.31051
- Vasey C, McBride J, Penta K. Circadian rhythm dysregulation and restoration: the role of melatonin. Nutrients. 2021;13(10). doi:10.3390/nu13103480
- Andrews CD, Foster RG, Alexander I, et al. Sleep-wake disturbance related to ocular disease: a systematic review of phase-shifting pharmaceutical therapies. Transl Vis Sci Technol. 2019;8(3):49. doi:10.1167/tvst.8.3.49
- Rosen R, Hu DN, Perez V, et al. Urinary 6-sulfatoxymelatonin level in age-related macular degeneration patients. Mol Vis. 2009;15:1673–1679.
- Schmid-Kubista KE, Glittenberg CG, Cezanne M, Holzmann K, Neumaier-Ammerer B, Binder S. Daytime levels of melatonin in patients with age-related macular degeneration. Acta Ophthalmol. 2009;87(1):89–93. doi:10.1111/j.1755-3768.2008.01173.x
- Tang L, Zhang C, Lu L, et al. Melatonin maintains inner blood-retinal barrier by regulating microglia via inhibition of PI3K/Akt/Stat3/NF-κB signaling pathways in experimental diabetic retinopathy. Front Immunol. 2022;13:831660. doi:10.3389/fimmu.2022.831660
- Bonnefond A, Clément N, Fawcett K, et al. Rare MTNR1B variants impairing melatonin receptor 1B function contribute to type 2 diabetes. Nat Genet. 2012;44(3):297–301. doi:10.1038/ng.1053
- Li C, Shi Y, You L, Wang L, Chen ZJ. Melatonin receptor 1A gene polymorphism associated with polycystic ovary syndrome. Gynecol Obstet Invest. 2011;72(2):130–134. doi:10.1159/000323542
- Mehrzadi S, Hemati K, Reiter RJ, Hosseinzadeh A. Mitochondrial dysfunction in age-related macular degeneration: melatonin as a potential treatment. Expert Opin Ther Targets. 2020;24(4):359–378. doi:10.1080/14728222.2020.1737015
- Dehdashtian E, Mehrzadi S, Yousefi B, et al. Diabetic retinopathy pathogenesis and the ameliorating effects of melatonin; involvement of autophagy, inflammation and oxidative stress. Life Sci. 2018;193:20–33. doi:10.1016/j.lfs.2017.12.001
- Kaur C, Foulds WS, Ling EA. Blood-retinal barrier in hypoxic ischaemic conditions: basic concepts, clinical features and management. Prog Retin Eye Res. 2008;27(6):622–647. doi:10.1016/j.preteyeres.2008.09.003
- Buscemi N, Vandermeer B, Hooton N, et al. The efficacy and safety of exogenous melatonin for primary sleep disorders. A meta-analysis. J Gen Intern Med. 2005;20(12):1151–1158. doi:10.1111/j.1525-1497.2005.0243.x
- Costa EJ, Lopes RH, Lamy-Freund MT. Permeability of pure lipid bilayers to melatonin. J Pineal Res. 1995;19(3):123–126. doi:10.1111/j.1600-079x.1995.tb00180.x
- Tosini G, Chaurasia SS, Michael Iuvone P. Regulation of arylalkylamine N-acetyltransferase (AANAT) in the retina. Chronobiol Int. 2006;23(1–2):381–391. doi:10.1080/07420520500482066
- Bhattacharya S, Patel KK, Dehari D, Agrawal AK, Singh S. Melatonin and its ubiquitous anticancer effects. Mol Cell Biochem. 2019;462(1–2):133–155. doi:10.1007/s11010-019-03617-5
- Claustrat B, Leston J. Melatonin: physiological effects in humans. Neurochirurgie. 2015;61(2–3):77–84. doi:10.1016/j.neuchi.2015.03.002
- Tosini G, Baba K, Hwang CK, Iuvone PM. Melatonin: an underappreciated player in retinal physiology and pathophysiology. Exp Eye Res. 2012;103:82–89. doi:10.1016/j.exer.2012.08.009
- Martínez A, Knappskog PM, Haavik J. A structural approach into human tryptophan hydroxylase and its implications for the regulation of serotonin biosynthesis. Curr Med Chem. 2001;8(9):1077–1091. doi:10.2174/0929867013372616
- Hardeland R. Taxon- and site-specific melatonin catabolism. Molecules. 2017;22(11). doi:10.3390/molecules22112015
- Bruni O, Angriman M, Melegari MG, Ferri R. Pharmacotherapeutic management of sleep disorders in children with neurodevelopmental disorders. Expert Opin Pharmacother. 2019;20(18):2257–2271. doi:10.1080/14656566.2019.1674283
- Kurhaluk N, Tkachenko H. Melatonin and alcohol-related disorders. Chronobiol Int. 2020;37(6):781–803. doi:10.1080/07420528.2020.1761372
- Jauhari A, Baranov SV, Suofu Y, et al. Melatonin inhibits cytosolic mitochondrial DNA-induced neuroinflammatory signaling in accelerated aging and neurodegeneration. J Clin Invest. 2020;130(6):3124–3136. doi:10.1172/jci135026
- Brennan R, Jan JE, Lyons CJ. Light, dark, and melatonin: emerging evidence for the importance of melatonin in ocular physiology. Eye. 2007;21(7):901–908. doi:10.1038/sj.eye.6702597
- Cipolla-Neto J, Amaral FGD. Melatonin as a hormone: new physiological and clinical insights. Endocr Rev. 2018;39(6):990–1028. doi:10.1210/er.2018-00084
- Drijfhout WJ, Van der linde AG, Kooi SE, Grol CJ, Westerink BH. Norepinephrine release in the rat pineal gland: the input from the biological clock measured by in vivo microdialysis. J Neurochem. 1996;66(2):748–755. doi:10.1046/j.1471-4159.1996.66020748.x
- Maronde E, Wicht H, Taskén K, et al. CREB phosphorylation and melatonin biosynthesis in the rat pineal gland: involvement of cyclic AMP dependent protein kinase type II. J Pineal Res. 1999;27(3):170–182. doi:10.1111/j.1600-079x.1999.tb00613.x
- Shin JW. Neuroprotective effects of melatonin in neurodegenerative and autoimmune central nervous system diseases. Encephalitis. 2023;3(2):44–53. doi:10.47936/encephalitis.2022.00094
- Polyakova VO, Kvetnoy IM, Anderson G, Rosati J, Mazzoccoli G, Linkova NS. Reciprocal interactions of mitochondria and the neuroimmunoendocrine system in neurodegenerative disorders: an important role for melatonin regulation. Front Physiol. 2018;9:199. doi:10.3389/fphys.2018.00199
- Masters A, Pandi-Perumal SR, Seixas A, Girardin JL, McFarlane SI. Melatonin, the hormone of darkness: from sleep promotion to Ebola treatment. Brain Disord Ther. 2014;4(1). doi:10.4172/2168-975x.1000151
- Huang CC, Chiou CH, Liu SC, et al. Melatonin attenuates TNF-α and IL-1β expression in synovial fibroblasts and diminishes cartilage degradation: implications for the treatment of rheumatoid arthritis. J Pineal Res. 2019;66(3):e12560. doi:10.1111/jpi.12560
- Watanabe K, Araki M, Iwasaki H. The embryonic pineal body as a multipotent organ. Microsc Res Tech. 1992;21(3):218–226. doi:10.1002/jemt.1070210305
- Pires-Lapa MA, Carvalho-Sousa CE, Cecon E, Fernandes PA, Markus RP. β-adrenoceptors trigger melatonin synthesis in phagocytes. Int J Mol Sci. 2018;19(8). doi:10.3390/ijms19082182
- Won E, Na KS, Kim YK. Associations between melatonin, neuroinflammation, and brain alterations in depression. Int J Mol Sci. 2021;23(1). doi:10.3390/ijms23010305
- Simonneaux V, Ribelayga C. Generation of the melatonin endocrine message in mammals: a review of the complex regulation of melatonin synthesis by norepinephrine, peptides, and other pineal transmitters. Pharmacol Rev. 2003;55(2):325–395. doi:10.1124/pr.55.2.2
- Aykan U, Güvel M, Paykal G, Uluoglu C. Neuropharmacologic modulation of the melatonergic system. Explorat Neurosci. 2023;2:287–306. doi:10.37349/en.2023.00029
- Provencio I, Rodriguez IR, Jiang G, Hayes WP, Moreira EF, Rollag MD. A novel human opsin in the inner retina. J Neurosci. 2000;20(2):600–605. doi:10.1523/jneurosci.20-02-00600.2000
- Abbott KS, Queener HM, Ostrin LA. The ipRGC-driven pupil response with light exposure, refractive error, and sleep. Optom Vis Sci. 2018;95(4):323–331. doi:10.1097/opx.0000000000001198
- Zawilska JB, Nowak JZ. Regulatory mechanisms in melatonin biosynthesis in retina. Neurochem Int. 1992;20(1):23–36. doi:10.1016/0197-0186(92)90122-8
- Fujieda H, Scher J, Hamadanizadeh SA, Wankiewicz E, Pang SF, Brown GM. Dopaminergic and GABAergic amacrine cells are direct targets of melatonin: immunocytochemical study of mt1 melatonin receptor in Guinea pig retina. Vis Neurosci. 2000;17(1):63–70. doi:10.1017/s0952523800171068
- Emet M, Ozcan H, Ozel L, Yayla M, Halici Z, Hacimuftuoglu A. A review of melatonin, its receptors and drugs. Eurasian J Med. 2016;48(2):135–141. doi:10.5152/eurasianjmed.2015.0267
- Gianesini C, Hiragaki S, Laurent V, Hicks D, Tosini G. Cone viability is affected by disruption of melatonin receptors signaling. Invest Ophthalmol Vis Sci. 2016;57(1):94–104. doi:10.1167/iovs.15-18235
- Rosen RB, Hu DN, Chen M, McCormick SA, Walsh J, Roberts JE. Effects of melatonin and its receptor antagonist on retinal pigment epithelial cells against hydrogen peroxide damage. Mol Vis. 2012;18:1640–1648.
- Liang FQ, Green L, Wang C, Alssadi R, Godley BF. Melatonin protects human retinal pigment epithelial (RPE) cells against oxidative stress. Exp Eye Res. 2004;78(6):1069–1075. doi:10.1016/j.exer.2004.02.003
- Boatright JH, Rubim NM, Iuvone PM. Regulation of endogenous dopamine release in amphibian retina by melatonin: the role of GABA. Vis Neurosci. 1994;11(5):1013–1018. doi:10.1017/s0952523800003941
- Scher J, Wankiewicz E, Brown GM, Fujieda H. AII amacrine cells express the MT1 melatonin receptor in human and macaque retina. Exp Eye Res. 2003;77(3):375–382. doi:10.1016/s0014-4835(03)00123-4
- Ivanova TN, Alonso-Gomez AL, Iuvone PM. Dopamine D4 receptors regulate intracellular calcium concentration in cultured chicken cone photoreceptor cells: relationship to dopamine receptor-mediated inhibition of cAMP formation. Brain Res. 2008;1207:111–119. doi:10.1016/j.brainres.2008.02.025
- Fukuhara PS, Lee DH, Chwa M, Kuppermann BD, Kenney MC. Effects of melatonin on human Retinal Pigment Epithelial cells (ARPE-19) and Müller cells (MIO-M1) exposed to cobalt chloride (CoCl2) in vitro. Invest Ophthalmol Visual Sci. 2021;62(8):2722.
- Laurent V, Sengupta A, Sánchez-Bretaño A, Hicks D, Tosini G. Melatonin signaling affects the timing in the daily rhythm of phagocytic activity by the retinal pigment epithelium. Exp Eye Res. 2017;165:90–95. doi:10.1016/j.exer.2017.09.007
- Wiechmann AF, Yang XL, Wu SM, Hollyfield JG. Melatonin enhances horizontal cell sensitivity in salamander retina. Brain Res. 1988;453(1–2):377–380. doi:10.1016/0006-8993(88)90182-5
- Yu HS, Hernandez V, Haywood M, Wong CG. Melatonin inhibits the proliferation of retinal pigment epithelial (RPE) cells in vitro. In Vitro Cell Dev Biol Anim. 1993;29a(5):415–418. doi:10.1007/bf02633991
- Hagström A, Kal Omar R, Williams PA, Stålhammar G. The rationale for treating uveal melanoma with adjuvant melatonin: a review of the literature. BMC Cancer. 2022;22(1):398. doi:10.1186/s12885-022-09464-w
- Kal Omar R, Hagström A, Stålhammar G. Adjuvant melatonin for uveal melanoma (AMUM): protocol for a randomized open-label phase III study. Trials. 2023;24(1):230. doi:10.1186/s13063-023-07245-9
- Roberts JE, Wiechmann AF, Hu DN. Melatonin receptors in human uveal melanocytes and melanoma cells. J Pineal Res. 2000;28(3):165–171. doi:10.1034/j.1600-079x.2001.280306.x
- Ku LC, Sheu ML, Cheng HH, et al. Melatonin protects retinal integrity through mediated immune homeostasis in the sodium iodate-induced mouse model of age-related macular degeneration. Biomed Pharmacother. 2023;161:114476. doi:10.1016/j.biopha.2023.114476
- Wiechmann AF, Udin SB, Summers Rada JA. Localization of Mel1b melatonin receptor-like immunoreactivity in ocular tissues of Xenopus laevis. Exp Eye Res. 2004;79(4):585–594. doi:10.1016/j.exer.2004.07.004
- Wang F, Zhou J, Lu Y, Chu R. Effects of 530 nm green light on refractive status, melatonin, MT1 receptor, and melanopsin in the Guinea pig. Curr Eye Res. 2011;36(2):103–111. doi:10.3109/02713683.2010.526750
- Zheng W, Chen Y, Zhou X, et al. Regulation of retinal melanopsin on lens-induced myopia in Guinea Pigs. Optom Vis Sci. 2020;97(7):489–495. doi:10.1097/opx.0000000000001529
- Alkozi HA, Wang X, Perez de Lara MJ, Pintor J. Presence of melanopsin in human crystalline lens epithelial cells and its role in melatonin synthesis. Exp Eye Res. 2017;154:168–176. doi:10.1016/j.exer.2016.11.019
- Alkozi HA, Perez de Lara MJ, Sánchez-Naves J, Pintor J. TRPV4 stimulation induced melatonin secretion by increasing Arylalkymine N-acetyltransferase (AANAT) protein level. Int J Mol Sci. 2017;18(4). doi:10.3390/ijms18040746
- Spadea L, Maraone G, Verboschi F, Vingolo EM, Tognetto D. Effect of corneal light scatter on vision: a review of the literature. Int J Ophthalmol. 2016;9(3):459–464. doi:10.18240/ijo.2016.03.24
- Kiliç A, Selek S, Erel O, Aksoy N. Protective effects of melatonin on oxidative-antioxidative balance and cataract formation in rats. Ann Ophthalmol. 2008;40(1):22–27.
- Anwar MM, Moustafa MA. The effect of melatonin on eye lens of rats exposed to ultraviolet radiation. Comp Biochem Physiol C Toxicol Pharmacol. 2001;129(1):57–63. doi:10.1016/s1532-0456(01)00180-6
- Lledó VE, Alkozi HA, Sánchez-Naves J, Fernandez-Torres MA, Guzman-Aranguez A. Melatonin counteracts oxidative damage in lens by regulation of Nrf2 and NLRP3 inflammasome activity. Exp Eye Res. 2022;215:108912. doi:10.1016/j.exer.2021.108912
- Li C, Tian Y, Yao A, Zha X, Zhang J, Tao Y. Intravitreal delivery of melatonin is protective against the photoreceptor loss in mice: a potential therapeutic strategy for degenerative retinopathy. Front Pharmacol. 2019;10:1633. doi:10.3389/fphar.2019.01633
- Martin XD, Malina HZ, Brennan MC, Hendrickson PH, Lichter PR. The ciliary body--The third organ found to synthesize indoleamines in humans. Eur J Ophthalmol. 1992;2(2):67–72. doi:10.1177/112067219200200203
- Chiquet C, Claustrat B, Thuret G, Brun J, Cooper HM, Denis P. Melatonin concentrations in aqueous humor of glaucoma patients. Am J Ophthalmol. 2006;142(2):325–327.e321. doi:10.1016/j.ajo.2006.03.040
- Alkozi H, Sánchez-Naves J, de Lara MJ, et al. Elevated intraocular pressure increases melatonin levels in the aqueous humour. Acta Ophthalmol. 2017;95(3):e185–e189. doi:10.1111/aos.13253
- Pintor J, Martin L, Pelaez T, Hoyle CH, Peral A. Involvement of melatonin MT(3) receptors in the regulation of intraocular pressure in rabbits. Eur J Pharmacol. 2001;416(3):251–254. doi:10.1016/s0014-2999(01)00864-0
- Andrés-Guerrero V, Molina-Martínez IT, Peral A, de Las Heras B, Pintor J, Herrero-Vanrell R. The use of mucoadhesive polymers to enhance the hypotensive effect of a melatonin analogue, 5-MCA-NAT, in rabbit eyes. Invest Ophthalmol Vis Sci. 2011;52(3):1507–1515. doi:10.1167/iovs.10-6099
- Huete-Toral F, Crooke A, Martínez-águila A, Pintor J. Melatonin receptors trigger cAMP production and inhibit chloride movements in nonpigmented ciliary epithelial cells. J Pharmacol Exp Ther. 2015;352(1):119–128. doi:10.1124/jpet.114.218263
- Rosenstein RE, Pandi-Perumal SR, Srinivasan V, Spence DW, Brown GM, Cardinali DP. Melatonin as a therapeutic tool in ophthalmology: implications for glaucoma and uveitis. J Pineal Res. 2010;49(1):1–13. doi:10.1111/j.1600-079X.2010.00764.x
- Gao J, Provencio I, Liu X. Intrinsically photosensitive retinal ganglion cells in glaucoma. Front Cell Neurosci. 2022;16:992747. doi:10.3389/fncel.2022.992747
- Wang H, Zhang Y, Ding J, Wang N. Changes in the circadian rhythm in patients with primary glaucoma. PLoS One. 2013;8(4):e62841. doi:10.1371/journal.pone.0062841
- Crooke A, Guzman-Aranguez A, Mediero A, et al. Effect of melatonin and analogues on corneal wound healing: involvement of Mt2 melatonin receptor. Curr Eye Res. 2015;40(1):56–65. doi:10.3109/02713683.2014.914540
- Wiechmann AF, Hollaway LR, Rada JA. Melatonin receptor expression in Xenopus laevis surface corneal epithelium: diurnal rhythm of lateral membrane localization. Mol Vis. 2009;15:2384–2403.
- Wahl C, Li T, Takagi Y, Howland H. The effects of light regimes and hormones on corneal growth in vivo and in organ culture. J Anat. 2011;219(6):766–775. doi:10.1111/j.1469-7580.2011.01429.x
- Sasaki M, Masuda A, Oishi T. Circadian rhythms of corneal mitotic rate, retinal melatonin and immunoreactive visual pigments, and the effects of melatonin on the rhythms in the Japanese quail. J Comp Physiol A. 1995;176(4):465–471. doi:10.1007/bf00196412
- Ciuffi M, Pisanello M, Pagliai G, et al. Antioxidant protection in cultured corneal cells and whole corneas submitted to UV-B exposure. J Photochem Photobiol B. 2003;71(1–3):59–68. doi:10.1016/j.jphotobiol.2003.07.004
- Rusciano D, Russo C. The therapeutic trip of melatonin eye drops: from the ocular surface to the retina. Pharmaceuticals. 2024;17(4). doi:10.3390/ph17040441
- Nopparat C, Chaopae W, Boontem P, Sopha P, Wongchitrat P, Govitrapong P. Melatonin attenuates high glucose-induced changes in beta amyloid precursor protein processing in human neuroblastoma cells. Neurochem Res. 2022;47(9):2568–2579. doi:10.1007/s11064-021-03290-5
- Chang CC, Huang TY, Chen HY, et al. Protective effect of melatonin against oxidative stress-induced apoptosis and enhanced autophagy in human retinal pigment epithelium cells. Oxid Med Cell Longev. 2018;2018:9015765. doi:10.1155/2018/9015765
- Jung KH, Hong SW, Zheng HM, et al. Melatonin ameliorates cerulein-induced pancreatitis by the modulation of nuclear erythroid 2-related factor 2 and nuclear factor-kappaB in rats. J Pineal Res. 2010;48(3):239–250. doi:10.1111/j.1600-079X.2010.00748.x
- Jung KH, Hong SW, Zheng HM, Lee DH, Hong SS. Melatonin downregulates nuclear erythroid 2-related factor 2 and nuclear factor-kappaB during prevention of oxidative liver injury in a dimethylnitrosamine model. J Pineal Res. 2009;47(2):173–183. doi:10.1111/j.1600-079X.2009.00698.x
- Jiang T, Chang Q, Cai J, Fan J, Zhang X, Xu G. Protective effects of melatonin on retinal inflammation and oxidative stress in experimental diabetic retinopathy. Oxid Med Cell Longev. 2016;2016:3528274. doi:10.1155/2016/3528274
- Jiang T, Chang Q, Zhao Z, et al. Melatonin-mediated cytoprotection against hyperglycemic injury in Müller cells. PLoS One. 2012;7(12):e50661. doi:10.1371/journal.pone.0050661
- Xie M, Hu A, Luo Y, Sun W, Hu X, Tang S. Interleukin-4 and melatonin ameliorate high glucose and interleukin-1β stimulated inflammatory reaction in human retinal endothelial cells and retinal pigment epithelial cells. Mol Vis. 2014;20:921–928.
- Do Carmo Buonfiglio D, Peliciari-Garcia RA, Do Amaral FG, et al. Early-stage retinal melatonin synthesis impairment in streptozotocin-induced diabetic Wistar rats. Invest Ophthalmol Vis Sci. 2011;52(10):7416–7422. doi:10.1167/iovs.10-6756
- Chen H, Liu X, Zhu W, et al. SIRT1 ameliorates age-related senescence of mesenchymal stem cells via modulating telomere shelterin. Front Aging Neurosci. 2014;6:103. doi:10.3389/fnagi.2014.00103
- Hikichi T, Tateda N, Miura T. Alteration of melatonin secretion in patients with type 2 diabetes and proliferative diabetic retinopathy. Clin Ophthalmol. 2011;5:655–660. doi:10.2147/opth.S19559
- Fu Y, Tang M, Fan Y, Zou H, Sun X, Xu X. Anti-apoptotic effects of melatonin in retinal pigment epithelial cells. Front Biosci. 2012;17(4):1461–1468. doi:10.2741/3997
- Liang FQ, Aleman TS, ZaixinYang CAV, Jacobson SG, Bennett J. Melatonin delays photoreceptor degeneration in the rds/rds mouse. Neuroreport. 2001;12(5):1011–1014. doi:10.1097/00001756-200104170-00029
- Flynn-Evans EE, Tabandeh H, Skene DJ, Lockley SW. Circadian rhythm disorders and melatonin production in 127 blind women with and without light perception. J Biol Rhythms. 2014;29(3):215–224. doi:10.1177/0748730414536852
- Tokarz P, Piastowska-Ciesielska AW, Kaarniranta K, Blasiak J. All-trans retinoic acid modulates DNA damage response and the expression of the VEGF-A and MKI67 genes in ARPE-19 cells subjected to oxidative stress. Int J Mol Sci. 2016;17(6). doi:10.3390/ijms17060898
- Tokarz P, Kaarniranta K, Blasiak J. Role of antioxidant enzymes and small molecular weight antioxidants in the pathogenesis of age-related macular degeneration (AMD). Biogerontology. 2013;14(5):461–482. doi:10.1007/s10522-013-9463-2
- Honda S, Hjelmeland LM, Handa JT. Oxidative stress--induced single-strand breaks in chromosomal telomeres of human retinal pigment epithelial cells in vitro. Invest Ophthalmol Vis Sci. 2001;42(9):2139–2144.
- Lau BW, Tsao GS, So KF, Yip HK. Expression of telomerase reverse transcriptase in adult goldfish retina. J Mol Neurosci. 2007;32(2):160–167. doi:10.1007/s12031-007-0031-6
- Blasiak J, Reiter RJ, Kaarniranta K. Melatonin in retinal physiology and pathology: the case of age-related macular degeneration. Oxid Med Cell Longev. 2016;2016:6819736. doi:10.1155/2016/6819736
- Reiter RJ, Mayo JC, Tan DX, Sainz RM, Alatorre-Jimenez M, Qin L. Melatonin as an antioxidant: under promises but over delivers. J Pineal Res. 2016;61(3):253–278. doi:10.1111/jpi.12360
- Aranda ML, Fleitas MFG, Dieguez H, et al. Melatonin as a therapeutic resource for inflammatory visual diseases. Curr Neuropharmacol. 2017;15(7):951–962. doi:10.2174/1570159x15666170113122120
- Korkmaz A, Reiter RJ, Topal T, Manchester LC, Oter S, Tan DX. Melatonin: an established antioxidant worthy of use in clinical trials. Mol Med. 2009;15(1–2):43–50. doi:10.2119/molmed.2008.00117
- Siu AW, Reiter RJ, To CH. Pineal indoleamines and vitamin E reduce nitric oxide-induced lipid peroxidation in rat retinal homogenates. J Pineal Res. 1999;27(2):122–128. doi:10.1111/j.1600-079x.1999.tb00606.x
- Tan DX, Manchester LC, Esteban-Zubero E, Zhou Z, Reiter RJ. Melatonin as a potent and inducible endogenous antioxidant: synthesis and metabolism. Molecules. 2015;20(10):18886–18906. doi:10.3390/molecules201018886
- Bardak H, Uğuz AC, Bardak Y. Protective effects of melatonin and memantine in human retinal pigment epithelium (ARPE-19) cells against 2-ethylpyridine-induced oxidative stress: implications for age-related macular degeneration. Cutan Ocul Toxicol. 2018;37(2):112–120. doi:10.1080/15569527.2017.1354218
- Yilmaz T, Celebi S, Kükner AS. The protective effects of melatonin, vitamin E and octreotide on retinal edema during ischemia-reperfusion in the Guinea pig retina. Eur J Ophthalmol. 2002;12(6):443–449. doi:10.1177/112067210201200601
- Karunadharma PP, Nordgaard CL, Olsen TW, Ferrington DA. Mitochondrial DNA damage as a potential mechanism for age-related macular degeneration. Invest Ophthalmol Vis Sci. 2010;51(11):5470–5479. doi:10.1167/iovs.10-5429
- Ramis MR, Esteban S, Miralles A, Tan DX, Reiter RJ. Protective effects of melatonin and mitochondria-targeted antioxidants against oxidative stress: a review. Curr Med Chem. 2015;22(22):2690–2711. doi:10.2174/0929867322666150619104143
- Bonilla E, Valero N, Chacín-Bonilla L, et al. Melatonin increases interleukin-1beta and decreases tumor necrosis factor alpha in the brain of mice infected with the Venezuelan equine encephalomyelitis virus. Neurochem Res. 2003;28(5):681–686. doi:10.1023/a:1022897314108
- Manchester LC, Coto-Montes A, Boga JA, et al. Melatonin: an ancient molecule that makes oxygen metabolically tolerable. J Pineal Res. 2015;59(4):403–419. doi:10.1111/jpi.12267
- Kern TS. Contributions of inflammatory processes to the development of the early stages of diabetic retinopathy. Exp Diabetes Res. 2007;2007:95103. doi:10.1155/2007/95103
- Wang L, He C. Nrf2-mediated anti-inflammatory polarization of macrophages as therapeutic targets for osteoarthritis. Front Immunol. 2022;13:967193. doi:10.3389/fimmu.2022.967193
- Bang J, Chang HW, Jung HR, et al. Melatonin attenuates clock gene cryptochrome1, which may aggravate mouse anti-type II collagen antibody-induced arthritis. Rheumatol Int. 2012;32(2):379–385. doi:10.1007/s00296-010-1641-9
- Colombo J, Jardim-Perassi BV, Ferreira JPS, et al. Melatonin differentially modulates NF-кB expression in breast and liver cancer cells. Anticancer Agents Med Chem. 2018;18(12):1688–1694. doi:10.2174/1871520618666180131112304
- Muxel SM, Pires-Lapa MA, Monteiro AW, et al. NF-κB drives the synthesis of melatonin in RAW 264.7 macrophages by inducing the transcription of the arylalkylamine-N-acetyltransferase (AA-NAT) gene. PLoS One. 2012;7(12):e52010. doi:10.1371/journal.pone.0052010
- Li SJ, Cheng WL, Kao YH, Chung CC, Trang NN, Chen YJ. Melatonin inhibits NF-κB/CREB/Runx2 signaling and alleviates aortic valve calcification. Front Cardiovasc Med. 2022;9:885293. doi:10.3389/fcvm.2022.885293
- Mihanfar A, Yousefi B, Azizzadeh B, Majidinia M. Interactions of melatonin with various signaling pathways: implications for cancer therapy. Cancer Cell Int. 2022;22(1):420. doi:10.1186/s12935-022-02825-2
- Chen F, Liu H, Wang X, et al. Melatonin activates autophagy via the NF-κB signaling pathway to prevent extracellular matrix degeneration in intervertebral disc. Osteoarthritis Cartilage. 2020;28(8):1121–1132. doi:10.1016/j.joca.2020.05.011
- Xia Y, Chen S, Zeng S, et al. Melatonin in macrophage biology: current understanding and future perspectives. J Pineal Res. 2019;66(2):e12547. doi:10.1111/jpi.12547
- Qin T, Feng D, Zhou B, et al. Melatonin attenuates lipopolysaccharide-induced immune dysfunction in dendritic cells. Int Immunopharmacol. 2023;120:110282. doi:10.1016/j.intimp.2023.110282
- Abd-Elhafeez HH, Hassan AHS, Hussein MT. Melatonin administration provokes the activity of dendritic reticular cells in the seminal vesicle of Soay ram during the non-breeding season. Sci Rep. 2021;11(1):872. doi:10.1038/s41598-020-79529-y
- Lin LW, Wang SW, Huang WC, et al. Melatonin inhibits VEGF-induced endothelial progenitor cell angiogenesis in neovascular age-related macular degeneration. Cells. 2023;12(5). doi:10.3390/cells12050799
- Zhu Z, Peng R, Shen H, et al. Treatment with melatonin after corneal graft attenuates rejection. Front Pharmacol. 2021;12:778892. doi:10.3389/fphar.2021.778892
- Macchi MM, Bruce JN. Human pineal physiology and functional significance of melatonin. Front Neuroendocrinol. 2004;25(3–4):177–195. doi:10.1016/j.yfrne.2004.08.001
- Osborne NN, Nash MS, Wood JP. Melatonin counteracts ischemia-induced apoptosis in human retinal pigment epithelial cells. Invest Ophthalmol Vis Sci. 1998;39(12):2374–2383.
- Tang L, Zhang C, Yang Q, et al. Melatonin maintains inner blood-retinal barrier via inhibition of p38/TXNIP/NF-κB pathway in diabetic retinopathy. J Cell Physiol. 2021;236(8):5848–5864. doi:10.1002/jcp.30269
- Huang R, Xu Y, Lu X, et al. Melatonin protects inner retinal neurons of newborn mice after hypoxia-ischemia. J Pineal Res. 2021;71(1):e12716. doi:10.1111/jpi.12716
- Romeo G, Liu WH, Asnaghi V, Kern TS, Lorenzi M. Activation of nuclear factor-kappaB induced by diabetes and high glucose regulates a proapoptotic program in retinal pericytes. Diabetes. 2002;51(7):2241–2248. doi:10.2337/diabetes.51.7.2241
- Qin T, Feng D, Zhou B, Bai L, Yin Y. Melatonin suppresses LPS-induced oxidative stress in dendritic cells for inflammatory regulation via the Nrf2/HO-1 axis. Antioxidants. 2022;11(10). doi:10.3390/antiox11102012
- Zheng B, Meng J, Zhu Y, Ding M, Zhang Y, Zhou J. Melatonin enhances SIRT1 to ameliorate mitochondrial membrane damage by activating PDK1/Akt in granulosa cells of PCOS. J Ovarian Res. 2021;14(1):152. doi:10.1186/s13048-021-00912-y
- Klettner A, Kampers M, Töbelmann D, Roider J, Dittmar M. The Influence of melatonin and light on VEGF secretion in primary RPE cells. Biomolecules. 2021;11(1). doi:10.3390/biom11010114
- Yang L, Zhang Y, Ma Y, et al. Effect of melatonin on EGF- and VEGF-induced monolayer permeability of HUVECs. Am J Physiol Heart Circ Physiol. 2019;316(5):H1178–H1191. doi:10.1152/ajpheart.00542.2018
- Choudhary P, Roy T, Chatterjee A, Mishra VK, Pant S, Swarnakar S. Melatonin rescues swim stress induced gastric ulceration by inhibiting matrix metalloproteinase-3 via down-regulation of inflammatory signaling cascade. Life Sci. 2022;297:120426. doi:10.1016/j.lfs.2022.120426
- Wiechmann AF, Smith AR. Melatonin receptor RNA is expressed in photoreceptors and displays a diurnal rhythm in Xenopus retina. Brain Res Mol Brain Res. 2001;91(1–2):104–111. doi:10.1016/s0169-328x(01)00134-6
- Ostrin LA. Ocular and systemic melatonin and the influence of light exposure. Clin Exp Optom. 2019;102(2):99–108. doi:10.1111/cxo.12824
- Luo N, Wang Y, Ma Y, Liu Y, Liu Z. Melatonin alleviates renal injury in diabetic rats by regulating autophagy. Mol Med Rep. 2023;28(5). doi:10.3892/mmr.2023.13101
- Xu Y, Lu X, Hu Y, et al. Melatonin attenuated retinal neovascularization and neuroglial dysfunction by inhibition of HIF-1α-VEGF pathway in oxygen-induced retinopathy mice. J Pineal Res. 2018;64(4):e12473. doi:10.1111/jpi.12473
- Xu Y, Cui K, Li J, et al. Melatonin attenuates choroidal neovascularization by regulating macrophage/microglia polarization via inhibition of RhoA/ROCK signaling pathway. J Pineal Res. 2020;69(1):e12660. doi:10.1111/jpi.12660
- Masana MI, Doolen S, Ersahin C, et al. MT(2) melatonin receptors are present and functional in rat caudal artery. J Pharmacol Exp Ther. 2002;302(3):1295–1302. doi:10.1124/jpet.302.3.1295
- Ting N, Thambyraja A, Sugden D, Scalbert E, Delagrange P, Wilson VG. Pharmacological studies on the inhibitory action of melatonin and putative melatonin analogues on porcine vascular smooth muscle. Naunyn Schmiedebergs Arch Pharmacol. 2000;361(3):327–333. doi:10.1007/s002109900198
- Vandeputte C, Giummelly P, Atkinson J, Delagrange P, Scalbert E, Capdeville-Atkinson C. Melatonin potentiates NE-induced vasoconstriction without augmenting cytosolic calcium concentration. Am J Physiol Heart Circ Physiol. 2001;280(1):H420–H425. doi:10.1152/ajpheart.2001.280.1.H420
- Meyer P, Pache M, Loeffler KU, et al. Melatonin MT-1-receptor immunoreactivity in the human eye. Br J Ophthalmol. 2002;86(9):1053–1057. doi:10.1136/bjo.86.9.1053
- Savaskan E, Olivieri G, Brydon L, et al. Cerebrovascular melatonin MT1-receptor alterations in patients with Alzheimer’s disease. Neurosci Lett. 2001;308(1):9–12. doi:10.1016/s0304-3940(01)01967-x
- Doolen S, Krause DN, Dubocovich ML, Duckles SP. Melatonin mediates two distinct responses in vascular smooth muscle. Eur J Pharmacol. 1998;345(1):67–69. doi:10.1016/s0014-2999(98)00064-8
- Trinh M, Kalloniatis M, Nivison-Smith L. Vascular changes in intermediate age-related macular degeneration quantified using optical coherence tomography angiography. Transl Vis Sci Technol. 2019;8(4):20. doi:10.1167/tvst.8.4.20
- Ueno Y, Iwase T, Goto K, et al. Association of changes of retinal vessels diameter with ocular blood flow in eyes with diabetic retinopathy. Sci Rep. 2021;11(1):4653. doi:10.1038/s41598-021-84067-2
- Arangino S, Cagnacci A, Angiolucci M, et al. Effects of melatonin on vascular reactivity, catecholamine levels, and blood pressure in healthy men. Am J Cardiol. 1999;83(9):1417–1419. doi:10.1016/s0002-9149(99)00112-5
- Ray CA. Melatonin attenuates the sympathetic nerve responses to orthostatic stress in humans. J Physiol. 2003;551(Pt 3):1043–1048. doi:10.1113/jphysiol.2003.043182
- Forsling ML, Wheeler MJ, Williams AJ. The effect of melatonin administration on pituitary hormone secretion in man. Clin Endocrinol. 1999;51(5):637–642. doi:10.1046/j.1365-2265.1999.00820.x
- Yan M, Wang H, Gu Y, Li X, Tao L, Lu P. Melatonin exerts protective effects on diabetic retinopathy via inhibition of Wnt/β-catenin pathway as revealed by quantitative proteomics. Exp Eye Res. 2021;205:108521. doi:10.1016/j.exer.2021.108521
- Ramezani M, Komaki A, Hashemi-Firouzi N, Mortezaee K, Faraji N, Golipoor Z. Therapeutic effects of melatonin-treated bone marrow mesenchymal stem cells (BMSC) in a rat model of Alzheimer’s disease. J Chem Neuroanat. 2020;108:101804. doi:10.1016/j.jchemneu.2020.101804
- Yoon YM, Lee JH, Song KH, Noh H, Lee SH. Melatonin-stimulated exosomes enhance the regenerative potential of chronic kidney disease-derived mesenchymal stem/stromal cells via cellular prion proteins. J Pineal Res. 2020;68(3):e12632. doi:10.1111/jpi.12632
- Lin Y, Ren X, Chen Y, Chen D. Interaction between mesenchymal stem cells and retinal degenerative microenvironment. Front Neurosci. 2020;14:617377. doi:10.3389/fnins.2020.617377
- Salih M, Shaharuddin B, Abdelrazeg S. A concise review on mesenchymal stem cells for tissue engineering with a perspective on ocular surface regeneration. Curr Stem Cell Res Ther. 2020;15(3):211–218. doi:10.2174/1574888x15666200129145251
- Madelaine R, Mourrain P. Endogenous retinal neural stem cell reprogramming for neuronal regeneration. Neural Regen Res. 2017;12(11):1765–1767. doi:10.4103/1673-5374.219028
- Bai C, Li X, Gao Y, et al. Melatonin improves reprogramming efficiency and proliferation of bovine-induced pluripotent stem cells. J Pineal Res. 2016;61(2):154–167. doi:10.1111/jpi.12334
- Gao Y, Ma L, Bai C, Zhang X, Yang W. Melatonin promotes self-renewal and nestin expression in neural stem cells from the retina. Histol Histopathol. 2019;34(6):645–654. doi:10.14670/hh-18-065
- Ping Z, Hu X, Wang L, et al. Melatonin attenuates titanium particle-induced osteolysis via activation of Wnt/β-catenin signaling pathway. Acta Biomater. 2017;51:513–525. doi:10.1016/j.actbio.2017.01.034
- Giannaccare G, Carnevali A, Senni C, Logozzo L, Scorcia V. Umbilical cord blood and serum for the treatment of ocular diseases: a comprehensive review. Ophthalmol Ther. 2020;9(2):235–248. doi:10.1007/s40123-020-00239-9
- Yi C, Pan X, Yan H, Guo M, Pierpaoli W. Effects of melatonin in age-related macular degeneration. Ann N Y Acad Sci. 2005;1057:384–392. doi:10.1196/annals.1356.029
- Harpsøe NG, Andersen LP, Gögenur I, Rosenberg J. Clinical pharmacokinetics of melatonin: a systematic review. Eur J Clin Pharmacol. 2015;71(8):901–909. doi:10.1007/s00228-015-1873-4
- Seiden DJ, Shah SM. A randomized, crossover, pharmacokinetics evaluation of a novel continuous release and absorption melatonin formulation. Prim Care Companion CNS Disord. 2019;21(4). doi:10.4088/PCC.19m02450
- Dal Monte M, Cammalleri M, Amato R, et al. A topical formulation of melatoninergic compounds exerts strong hypotensive and neuroprotective effects in a rat model of hypertensive glaucoma. Int J Mol Sci. 2020;21(23). doi:10.3390/ijms21239267
- Tao Y, Hu B, Ma Z, et al. Intravitreous delivery of melatonin affects the retinal neuron survival and visual signal transmission: in vivo and ex vivo study. Drug Deliv. 2020;27(1):1386–1396. doi:10.1080/10717544.2020.1818882
- Yu H, Wu W, Lin X, Feng Y. Polysaccharide-based nanomaterials for ocular drug delivery: a perspective. Front Bioeng Biotechnol. 2020;8:601246. doi:10.3389/fbioe.2020.601246
- Ahn JH, Kim HD, Abuzar SM, et al. Intracorneal melatonin delivery using 2-hydroxypropyl-β-cyclodextrin ophthalmic solution for granular corneal dystrophy type 2. Int J Pharm. 2017;529(1–2):608–616. doi:10.1016/j.ijpharm.2017.07.016
- Musumeci T, Bucolo C, Carbone C, Pignatello R, Drago F, Puglisi G. Polymeric nanoparticles augment the ocular hypotensive effect of melatonin in rabbits. Int J Pharm. 2013;440(2):135–140. doi:10.1016/j.ijpharm.2012.10.014
- Carbone C, Manno D, Serra A, et al. Innovative hybrid vs polymeric nanocapsules: the influence of the cationic lipid coating on the ”4S”. Colloids Surf B Biointerfaces. 2016;141:450–457. doi:10.1016/j.colsurfb.2016.02.002
- Bessone CDV, Martinez SM, Luna JD, et al. Neuroprotective effect of melatonin loaded in ethylcellulose nanoparticles applied topically in a retinal degeneration model in rabbits. Exp Eye Res. 2020;200:108222. doi:10.1016/j.exer.2020.108222
- Arranz-Romera A, Davis BM, Bravo-Osuna I, et al. Simultaneous co-delivery of neuroprotective drugs from multi-loaded PLGA microspheres for the treatment of glaucoma. J Control Release. 2019;297:26–38. doi:10.1016/j.jconrel.2019.01.012