
Abstract
Liver fibrosis is a dynamic reversible pathological process in the development of chronic liver disease to cirrhosis. However, the current treatments are not administered for a long term due to their various side effects. Autophagy is initiated to decompose damaged or excess organelles, which had been found to alter the progression of liver fibrosis. In this article, we hypothesized that fucoidan from Fucus vesiculosus may attenuate liver fibrosis in mice by inhibition of the extracellular matrix and autophagy in carbon tetrachloride- and bile duct ligation-induced animal models of liver fibrosis. The results were determined using enzyme-linked immunosorbent assay, quantitative real-time polymerase chain reaction, Western blotting, and immunohistochemical staining. Fucoidan from F. vesiculosus could inhibit the activation of hepatic stellate cells and the formation of extracellular matrix and autophagosomes, and its effect may be associated with the downregulation of transforming growth factor beta 1/Smads pathways. Fucoidan, as an autophagy and transforming growth factor beta 1 inhibitor, could be a promising potential therapeutic agent for liver fibrosis.
Introduction
Liver fibrosis, a dynamic reversible pathological process that can cause chronic liver disease to develop into cirrhosis (mainly seen in intrahepatic fibrous dysplasia), results in the production and deposition of extracellular matrix (ECM) and an imbalance between synthesis and degradation.Citation1 Approximately 70% of patients with chronic liver disease have associated liver fibrosis. If the process is not stopped, 25% will deteriorate further to decompensated cirrhosis accompanied by digestive tract hemorrhage, hepatic encephalopathy, and hepatorenal syndrome.Citation2–Citation4 The current treatments to tackle liver fibrosis include antiviral, antioxidant, and immune regulation therapies that are not administered in the long term due to their various side effects.Citation5,Citation6 Therefore, safe and effective screening of drugs used to control liver fibrosis plays an important role in cirrhosis and hepatoma prevention.
Hepatic ECM contains bioactive substances. Under normal physiological conditions, the ECM provides nutrition and is involved in the immune response. When stimulated by various cytokines, the ECM evolves to liver fibrosis due to synthesis of its components, including collagen I, fibronectin, laminin (LN), and hyaluronic acid, or reduced degradation induced by an imbalance between matrix metalloproteinases (MMPs) and tissue inhibitor of matrix metalloproteinases (TIMPs).Citation7,Citation8 A number of studies have demonstrated that the activation and proliferation of hepatic stellate cells (HSCs) are the key points in the production of ECM and includes the participation of other cells such as hepatic sinus endothelial cells, Kupffer cells, neutrophils, T-cells, and natural killer cells.Citation1,Citation7,Citation9,Citation10 HSCs differentiate into myofibroblasts (MFBs) with abundant production of α-smooth muscle actin (α-SMA) and ECM following liver damage, eventually leading to liver fibrosis.Citation11 The related signaling pathways that influence each other in this process mainly include the transforming growth factor beta (TGF-β), platelet-derived growth factor, Wnt/β-catenin, Toll-like receptor, and other important pathways.Citation7,Citation12 The establishment of animal models that simulate the pathogenic mechanism has always been difficult, and commonly used drugs that induce liver fibrosis include carbon tetrachloride (CCl4) and dimethylnitrosamine. Another liver fibrosis model is induced by cholestasis via surgery with bile duct ligation (BDL).Citation13–Citation15 CCl4 can lead to the degeneration and necrosis of hepatic cells by directly dissolving liver cell membranes to initiate lipid peroxidation, which is similar to human liver fibrosis in morphology and pathophysiology.Citation16 The CCl4-induced liver fibrosis model is widely used due to its stable pathological characteristics and good repeatability, while BDL-induced disease is akin to the biochemical and histological changes in human small nodular cirrhosis.Citation17 These two animal models were jointly adopted in the present study to conduct drug screening for liver fibrosis.
Autophagy, defined by Ashford and PorterCitation18 using an electron microscope when observing human liver cells, is a naturally occurring physiological process in which damaged or excess organelles are destroyed, to allow the synthesis of new organelles in the absence of nutrients and energy.Citation18–Citation21 Programmed cell death plays an essential role in acute liver damage, ischemia reperfusion, and other systemic diseases; however, less research has been carried out on liver fibrosis.Citation22–Citation27 In recent years, numerous studies have found that autophagy can alter the progression of liver fibrosis. Hernandez-Gea et al found that cells gained energy to supply materials for the activation of HSCs following lipid autophagic degradation.Citation28 Liu et al showed that the autophagy inhibitor 3-methyladenine affected the proliferation and activation of HSCs to alleviate fibrosis.Citation29 Therefore, autophagy could be regarded as a target in the prevention of liver fibrosis. Fucoidan, a polysaccharide extracted from brown seaweed, was reported to have many benefits, such as anti-inflammation and anticancer activities, and it affects oxidative stress and vascular physiology.Citation30–Citation32 In 2008, Hayashi et alCitation33 demonstrated the protective effect of fucoidan on CCl4-induced liver fibrosis, while Hong et alCitation15 reported that fucoidan could protect against liver fibrogenesis induced by dimethylnitrosamine through the inhibition of oxidative stress and inflammatory cytokine release.Citation13,Citation15 However, it is unclear whether fucoidan can ameliorate CCl4- and BDL-induced liver fibrosis as autophagy-targeting drugs, and its relationship with the TGF-β/Smad pathway is unknown.
In the present study, we hypothesized that fucoidan from Fucus vesiculosus may attenuate liver fibrosis in mice by inhibition of the ECM and autophagy in CCl4- and BDL-induced animal models of liver fibrosis. The potential mechanism of action of fucoidan was also investigated.
Materials and methods
Mice and fibrosis induction
Male C57 mice weighing 20–22 g were purchased from Shanghai Laboratory Animal Co., Ltd. (Shanghai, People’s Republic of China). All animals were housed in an air-conditioned room at a temperature of 25°C with a 12-hour alternating light and dark cycle and permitted free access to food and water. In the liver fibrosis models, mice were injected intraperitoneally with 1 mL/kg body weight of CCl4 (1:10 v/v; Sigma-Aldrich Co., St Louis, MO, USA) in olive oil twice a week for 8 weeks.Citation33 Fucoidan (Sigma-Aldrich Co.) was diluted in saline and was administered orally at daily doses of 10 mg/kg, 25 mg/kg, and 50 mg/kg once a day. In BDL-induced liver fibrosis, all mice were anesthetized with 1.25% Nembutal (Sigma-Aldrich Co.) after a 24-hour fast. When the reflection and pain stimulation disappeared, the abdominal cavity was opened along the linea alba. A wet swab was used to expose the diaphragm and umbilical region and separate them from the bile duct, flank portal vein, and hepatic artery. The bile duct was ligated with two surgical knots using 5-0 suture and cut in between the two knots.
The abdominal cavity was washed with saline, and the two abdominal layers and skin were closed.Citation17 The mice were fed in a warm and humid environment until fully awake and active and treated with and without drugs for 2 weeks. All animal experiments were performed in accordance with legal regulations and approved by the Animal Care and Use Committee of Shanghai Tongji University. Serum and liver tissues were obtained for the following determinations:
Mice with CCl4-induced liver fibrosis were randomly divided into six groups (eight mice per group) as follows:
control (CCl4) group: saline by gavage;
fucoidan (CCl4) group: fucoidan (50 mg/kg) daily by gavage;
CCl4 group: CCl4 injected intraperitoneally;
CCl4 + fucoidan (10) group: CCl4 injected intraperitoneally with fucoidan (10 mg/kg) daily by gavage;
CCl4 + fucoidan (25) group: CCl4 injected intraperitoneally with fucoidan (25 mg/kg) daily by gavage; and
CCl4 + fucoidan (50) group: CCl4 injected intraperitoneally with fucoidan (50 mg/kg) daily by gavage.
The BDL-induced liver fibrosis mice were randomly divided into groups as described earlier, including the control (BDL) group, fucoidan (BDL) group, BDL group, BDL + fucoidan (10) group, BDL + fucoidan (25) group, and BDL + fucoidan (50) group (eight mice per group).
Serum analysis
Serum alanine aminotransferase (ALT) and aspartate aminotransferase (AST) were determined using an automated chemistry analyzer (Olympus AU1000; Olympus Corporation, Tokyo, Japan). Hydroxyproline and LN were determined using kits according to the manufacturers’ instructions.
Hematoxylin and eosin staining and Masson’s trichrome staining
Fresh liver samples were embedded in paraffin for histological examination using hematoxylin and eosin (H&E) and Masson’s trichrome staining. Sections (5 μm thick) were cut and prepared for further experiments. Collagen fiber stained blue, cytoplasm stained red, and nucleus stained blue-violet by Masson dye were observed under a light microscope with a digital camera (Leica Microsystems, Wetzlar, Germany).
Real-time polymerase chain reaction
Total RNA was extracted from frozen liver tissue by the pyrolysis of Trizol, chloroform, isopropyl alcohol, and ethanol (Thermo Fisher Scientific, Waltham, MA, USA) and transcribed into cDNA using the reverse transcription kit (TaKaRa Biotechnology, Dalian, People’s Republic of China). Real-time polymerase reaction (PCR) was carried out according to the protocols described by SYBR Premix EX Taq (TaKaRa Biotechnology) using a 7900HT Fast Real-Time PCR system (Thermo Fisher Scientific). The ratio of target genes and β-actin was calculated on the basis of the solubility curve. All primers used in the experiments are shown in .
Table 1 Nucleotide sequences of the primers used for qRT-PCR
Western blotting analysis
The total tissue protein was extracted by radio immunoprecipitation assay lysis buffer with phenylmethanesulfonyl fluoride, and the concentration was calculated using the bicinchoninic acid protein assay (Kaiji, People’s Republic of China). Equal amounts of samples were added to 8%–12.5% sodium dodecyl sulfate-polyacrylamide gels and transferred onto polyvinylidene fluoride membranes that were sequentially blocked with defatted milk and incubated with specific primary and secondary antibodies. The primary antibodies used were as follows: α-SMA, TIMP1, MMP-9, Co-I, Beclin-1, P62 (1:500, all from Proteintech Group, Inc., Chicago, IL, USA); TGF-β1, smad2, p-smad2, LC3, β-actin (1:1,000, all from Cell Signaling Technology, Danvers, MA, USA); and smad3, p-smad3 (1:500, both from ABclonal, Cambridge, MA, USA). The Odyssey two-color infrared laser imaging system (LI-COR Biosciences, Lincoln, NE, USA) was used to scan the membranes for image acquisition.
Immunohistochemical staining
The prepared paraffin sections were dewaxed and rehydrated with different concentrations of alcohol. After washing with phosphate-buffered saline solution, the sections were pretreated using a microwave antigen retrieval technique with heating to 95°C and cooling to room temperature (four cycles). Antibodies, including α-SMA (1:100), TIMP1 (1:100), MMP-9 (1:100), Co-I (1:100), Beclin-1 (1:100), TGF-β1 (1:200), p-smad2 (1:100), LC3 (1:50), and p-smad3 (1:100), were added to the slices and then incubated at 4°C overnight. Following incubation with secondary antibody, a DAB kit was used and positive areas were observed by a microscope with a digital camera.
Electron microscopy
The liver tissues were prefixed with 3% glutaraldehyde buffered with 0.2 mmol/L cacodylate for 4 hours and postfixed in 1% osmium tetroxide for 1 hour. Autophagosomes were viewed by electron microscopy (JEM-1230; JEOL, Tokyo, Japan), and images were acquired.
Statistical analysis
The statistical analysis was performed using the SPSS 20.0 software package (IBM Corporation, Armonk, NY, USA). All data were compared by calculating the mean ± standard deviation using the Student–Newman–Keuls test or one-way analysis of variance. A P-value <0.05 was considered statistically significant. The positively stained areas were evaluated by integrated optical density and analyzed using Image-Pro Plus 6.0 (Media Cybernetics, Silver Spring, MD, USA).
Results
Fucoidan improved the liver function in liver fibrosis
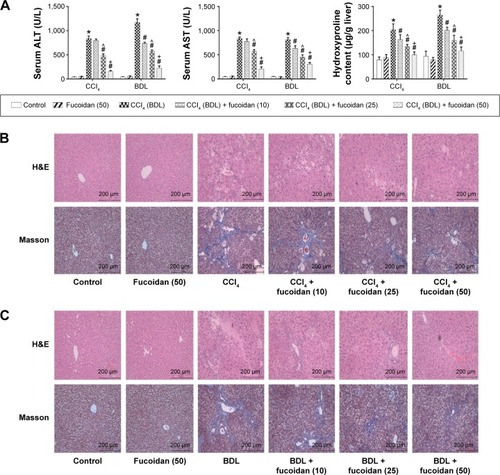
ALT and AST are important indices for reflecting the status of liver function. We conducted a series of experiments to clarify the effect of fucoidan on liver enzyme levels. The serum levels of liver enzymes in the CCl4 and BDL groups were significantly elevated, which indicated serious liver damage. With increasing fucoidan dose (10 mg/kg, 25 mg/kg, and 50 mg/kg), the levels of ALT and AST declined gradually. However, 10 mg/kg of fucoidan had no obvious influence on the fibrosis caused by CCl4. Hydroxyproline, which can reflect the severity of liver fibrosis, is a characteristic amino acid in the synthesis of collagens. The hydroxyproline levels in liver tissue were higher in the model groups (both BDL and CCl4 groups) and decreased with increasing fucoidan concentration (). H&E and Masson staining were used to determine general morphology and collagen fibers in liver tissues. According to the results of H&E staining, the model groups showed inflammatory infiltrates, degeneration and necrosis in liver cells around the central vein, and connective tissue hyperplasia. The degree of damage in the fucoidan treatment groups was reduced. Consistent with the changes in liver enzymes and H&E staining, Masson staining of liver sections showed blue areas of collagen fiber and red areas of cytoplasm. Collagen fiber in the fucoidan groups showed a decreasing trend relative to the model groups (). These results indicated that different concentrations of fucoidan significantly alleviated liver fibrosis in a dose-dependent manner.
Figure 1 Effect of fucoidan on liver cytolysis.
Abbreviations: AST, aspartate aminotransferase; SD, standard deviation; CCl4, carbon tetrachloride; BDL, bile duct ligation; H&E, hematoxylin and eosin; ALT, alanine aminotransferase.
Fucoidan inhibited the formation of ECM
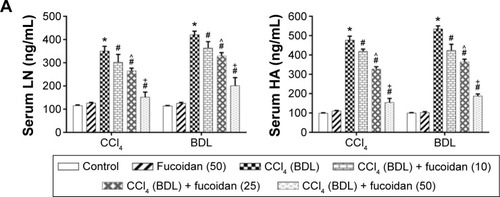
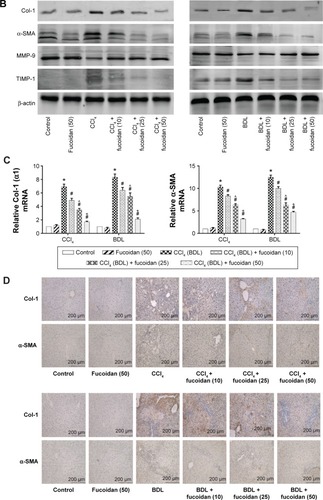
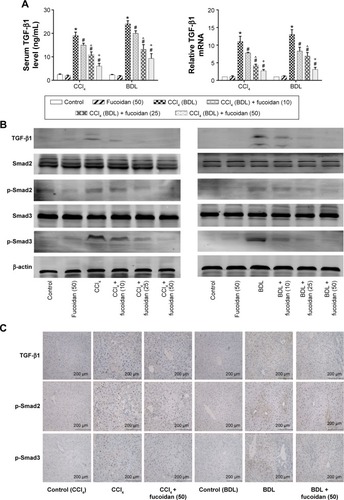
Hyaluronic acid, fibronectin, LN, collagen I, α-SMA, MMPs, and TIMPs are involved in the synthesis of ECM. Enzyme-linked immunosorbent assay kits were used to detect serum hyaluronic acid and LN, and the results were consistent with those for ALT and AST. Increased serum levels in both model groups indicated severe fibrosis, and decreased levels in the fucoidan groups indicated attenuation of liver lesions (). PCR and Western blot were used to determine changes in the gene and protein levels of ECM components in liver tissues in the different treatment groups, respectively (). The results showed increased expression of collagen I, α-SMA, and TIMP1 in the model groups, and fucoidan inhibited this increase. Similarly, immunohistochemical staining of liver biopsies showed that the changes in these components in liver tissues were consistent with the results for gene and protein expression (). These results demonstrate that the production of ECM was inhibited by fucoidan to block the progression of liver fibrosis.
Figure 2 Effect of fucoidan on ECM in liver fibrosis.
Abbreviations: ECM, extracellular matrix; HA, hyaluronic acid; LN, laminin; α-SMA, α-smooth muscle actin; MMP, matrix metalloproteinase; TIMP, tissue inhibitor of matrix metalloproteinase; mRNA, messenger ribonucleic acid; SD, standard deviation; CCl4, carbon tetrachloride; BDL, bile duct ligation.
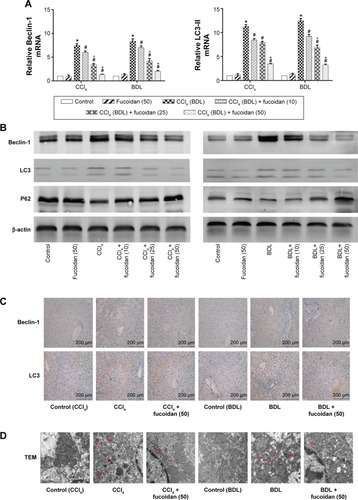
Fucoidan decreased the formation of autophagosomes by downregulation of Beclin-1 and LC3
Autophagy can provide energy for the acceleration of liver fibrosis via the degradation of cell protein. Beclin-1, LC3, and P62 are important markers of autophagosome formation. To assess autophagy levels in liver tissues after fucoidan treatment, PCR, Western blot, and immunohistochemical staining were used to show the gene and protein tissue levels of these indices, respectively. The results showed that proteins, such as Beclin-1 and LC3, promoted autophagy and were significantly increased in the model groups with fibrosis (BDL and CCl4 groups). Fucoidan decreased cell damage induced by autophagy; thus, Beclin-1 and LC3 levels in the fucoidan-treated groups (10 mg/kg, 25 mg/kg, and 50 mg/kg) gradually decreased in a dose-dependent manner. P62 – an autophagy-related transporter – decreased in the model groups and increased following fucoidan treatment (). All markers showed the formation of autophagosomes detected by electron microscopy, including the observation of microstructure. Compared with the control group, the number of autophagosomes in the BDL and CCl4 groups increased, and following fucoidan administration, the agglutinated chromatin in mitochondria and autophagy corpuscles was not easily seen. In summary, fucoidan mitigated the injury induced by autophagy via the inhibition of autophagosome formation by downregulating Beclin-1 and LC3.
Figure 3 Effect of fucoidan on the process of autophagy in liver fibrosis.
Abbreviations: mRNA, messenger ribonucleic acid; SD, standard deviation; BDL, bile duct ligation; CCl4, carbon tetrachloride; TEM, transmission electron microscopy.
Fucoidan prevented the activation of HSCs via the TGF-β1/Smads signaling pathway
TGF-β1 is mainly found in HSCs, hepatic sinus endothelial cells, and inflammatory cells during liver fibrosis and plays an important role in tissues and organs. As an important factor in improving liver fibrosis, TGF-β1 can mediate necrosis and autophagy by the activation of smads, which are phosphorylated in the nuclear region. In order to confirm the conduction pathway, we determined the levels of TGF-β1 in serum and tissues and its gene and protein expression (). The results showed that the levels of these factors were upregulated in the model groups with liver fibrosis induced by BDL and CCl4 and declined after fucoidan treatment with statistically significant differences observed. We also focused on the phosphorylation of smads, such as smad2 and smad3, and found a significant increase in the BDL and CCl4 groups and a dose-dependent decrease following fucoidan treatment. Western blot and immunohistochemical staining showed consistent findings (). These results suggest that fucoidan reduced the expression of TGF-β1, which mediated negative activation of downstream smads signaling pathways, which may be an essential pathway in the induction of necrosis and autophagy during liver fibrosis.
Figure 4 Effect of fucoidan on the TGF-β1/Smads pathways in liver fibrosis.
Abbreviations: TGF-β1, transforming growth factor beta 1; mRNA, messenger ribonucleic acid; SD, standard deviation; CCl4, carbon tetrachloride; BDL, bile duct ligation.
Discussion
Liver fibrosis is a scar formation induced by a compensatory response to a variety of chronic liver injuries. As the common pathological process in many liver diseases, liver fibrosis also leads to cirrhosis.Citation34 It is important to identify new, safe, and effective drug therapies due to the lack of surgical procedures as a result of diffuse pathological changes. Fucoidan, a polysaccharide based on fucose, has attracted the attention of scientists.Citation30,Citation35,Citation36 In the present study, two animal models of liver fibrosis (CCl4 and BDL) were established to determine the protective effect and mechanism of action of fucoidan by analyzing biochemical indicators (ALT, AST, and hydroxyproline), ECM components (collagenase I, MMPs, and TIMPs), and other key proteins located in signaling pathways (TGF-β1 and Smads).
The study by Senties-Gomez et al confirmed that increased synthesis and insufficient degradation of the ECM, which leads to excess deposition and interactions with other cytokines, occur in the initiation and development of liver fibrosis.Citation37 Collagen type I, found in late pathological changes, is an important component of the ECM. As the characteristic amino acid of collagen type I is hydroxyproline, the serum level of this amino acid may be an important indicator of fibrosis. MMPs, a group of enzymes that contain Ca2+ and Zn2+, could play a role in ECM degradation, and TIMPs, secreted by activated HSCs, are specific inhibitors of MMPs.Citation11 ECM metabolism is dependent on the balance between MMPs and TIMPs, mainly MMP-9/TIMP1.Citation38,Citation39 As the key cells in ECM production, HSCs can transform into MFBs, which express the unique molecular marker protein α-SMA.Citation10 Hong et al and Moon et al verified that fucoidan increased MMP-9 secretion in the monocytic cell line U937 and downregulated type I procollagen and α-SMA synthesis in human skin fibroblasts.Citation15,Citation40 In our experiments, the expression of collagen type I and α-SMA detected by PCR, Western blot, and immunohistochemical staining in the CCl4 and BDL model groups showed an upward trend with TIMP1 regarded as an ECM metabolic inhibitory enzyme but decreased in a dose-dependent manner in the fucoidan-treated groups (10 mg/kg, 25 mg/kg, and 50 mg/kg). In contrast, MMP-9, the enzyme that promotes the metabolism of ECM, decreased in the model groups and increased after fucoidan treatment. These results showed that fucoidan can protect against liver injury induced by CCl4 and BDL through the effective inhibition of ECM production and preventing the change from HSCs to MFBs, resulting in reduced liver fibrosis. Based on detection of the above indicators, measurement of ALT, AST, and hydroxyproline suggested that liver function changed with ECM formation. Compared with the model groups, liver biopsy pathology in the fucoidan-treated groups showed less collagen fiber formation and focal necrosis that was dose dependent. In summary, fucoidan inhibited HSCs and reduced ECM formation and α-SMA release to protect against liver fibrosis.
Autophagy, referred to as type II programmed cell death widely exists in eukaryotic cell biology, occurs less frequently under normal circumstances but can be induced to maintain the balance between proteins and organelles under stress. However, excess activation of autophagy can also increase cell damage and death.Citation41,Citation42 In recent years, an increasing number of studies have verified the role of autophagy in chronic liver damage, such as nonalcoholic and alcoholic fatty liver disease, viral hepatitis, and cirrhosis.Citation43–Citation46 During liver fibrosis, the occurrence of autophagy is associated with the activation of HSCs acting on the TGF-β, platelet-derived growth factor, Wnt/β-catenin, and Toll-like receptor pathways.Citation47–Citation51 Therefore, we hypothesize that inhibition of autophagy may be a new drug target in the prevention of liver fibrosis. Beclin-1, LC3, and P62 were assessed to demonstrate the expression of autophagy in this study. The results showed that autophagy was upregulated in the CCl4 and BDL model groups and decreased with increased fucoidan concentration. Abundant autophagosomes and autophagolysosomes were seen in liver tissues of the model groups but were rarely seen following treatment with fucoidan. These findings show that fucoidan can effectively inhibit autophagy, which provides the energy for the activation of HSCs and thus slows down the process of liver fibrosis.
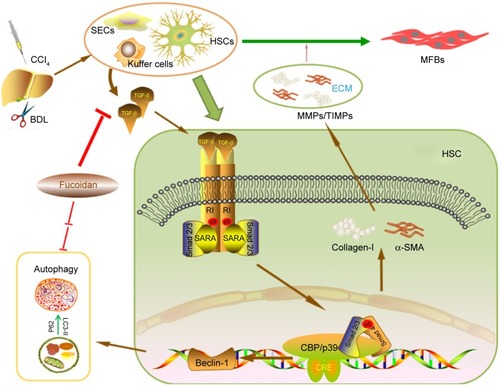
How does fucoidan regulate the formation of ECM and autophagy in liver fibrosis? Shen et al, He et al, and Ghavami et al found that autophagy is a regulator of the TGF-β1 pathway in atrial MFBs and HSCs, and these findings were verified by other researchers.Citation12,Citation23,Citation28,Citation29,Citation45,Citation47,Citation48,Citation50 The TGF-β family, which includes various functional cytokines, can regulate cell growth, proliferation, differentiation, migration, and apoptosis. Research evidence shows that TGF-β, mainly TGF-β1 with the highest biological activity located in intrahepatic cells, is highly expressed in HSCs, hepatic sinus, endothelial cells, and the inflammatory cells that affect liver fibrosis.Citation52,Citation53 HSCs were activated by CCl4 and cholestasis and transformed into MFBs, which secreted TGF-β1. TGF-β1 can expedite the processes of collagen, and TIMP1 in the ECM and TGFR1 on the surface of cell membranes can combine with TGF-β1 to stimulate Smad2/3 located in the cytoplasm.Citation54–Citation56 The examination of signal molecules in the TGF-β1/Smads pathway showed an increased trend in the model groups compared with the normal group, which declined following fucoidan treatment. The gene and protein expression results indicated that fucoidan effectively reduced secreted TGF-β1, thus inhibiting the downstream pathway. In summary, fucoidan inhibited the activation of HSCs, the formation of the ECM, and the release of TGF-β1. Insufficient TGF-β1 formed complexes with its receptor, which led to reduced phosphorylation of Smad2/3. Smad2/3 cannot be transferred from the pulp to the nucleus to combine with specific DNA sequences to promote Beclin-1 transcription due to its unphosphorylated state.Citation51 Beclin-1 usually promotes the occurrence of autophagy through an interaction with PI3K, which induces the conversion of LC3-I to LC3-II.Citation19,Citation57 Following fucoidan treatment, the inhibition of TGF-β1 hindered the formation of autophagosomes, which degraded organelles to protect liver cells via the downregulated TGF-β1/Smad signaling pathways ().
Figure 5 Mechanism of fucoidan action.
Abbreviations: HSCs, hepatic stellate cells; TGF-β1, transforming growth factor beta 1; DNA, deoxyribose nucleic acid; CCl4, carbon tetrachloride; BDL, bile duct ligation; MFBs, myofibroblasts; ECM, extracellular matrix; MMPs, matrix metalloproteinases; TIMPs, tissue inhibitor of matrix metalloproteinases; α-SMA, α-smooth muscle actin; SEC, sinusoidal endothelial cells. CRE, c-AMP response element.
Conclusion
In conclusion, we preliminarily found that the activation of HSCs to secrete TGF-β1, which induced formation of the ECM and autophagy, was inhibited by fucoidan from F. vesiculosus during liver fibrosis in mice. As new drug targets, autophagy and TGF-β1 inhibitors could be promising potential therapeutic agents for liver fibrosis. Of course, the safety and other related mechanisms of fucoidan require further investigation prior to clinical application.
Authors contributions
All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in either drafting the article or revising it critically for important intellectual content; gave final approval of the version to be published; and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in this work.
References
- FriedmanSLHepatic fibrosis – overviewToxicology2008254312012918662740
- LimYSKimWRThe global impact of hepatic fibrosis and end-stage liver diseaseClin Liver Dis200812473374618984463
- ZhangYLiSHeLCombination therapy of fenofibrate and ursodeoxycholic acid in patients with primary biliary cirrhosis who respond incompletely to UDCA monotherapy: a meta-analysisDrug Des Devel Ther2015927572766
- ZhangHYangJZhuRCombination therapy of ursodeoxycholic acid and budesonide for PBC-AIH overlap syndrome: a meta-analysisDrug Des Devel Ther20159567574
- SchwabeRBatallerRLiver fibrosisSemin Liver Dis2015352959625974895
- ChenRJWuHHWangYJStrategies to prevent and reverse liver fibrosis in humans and laboratory animalsArch Toxicol201589101727175025963329
- LeeUEFriedmanSLMechanisms of hepatic fibrogenesisBest Pract Res Clin Gastroenterol201125219520621497738
- SchuppanDAfdhalNHLiver cirrhosisLancet2008371961583885118328931
- DuvalFMoreno-CuevasJEGonzalez-GarzaMTRodriguez-MontalvoCCruz-VegaDEProtective mechanisms of medicinal plants targeting hepatic stellate cell activation and extracellular matrix deposition in liver fibrosisChin Med2014912725606051
- ReevesHLFriedmanSLActivation of hepatic stellate cells – a key issue in liver fibrosisFront Biosci20027d808d82611897564
- PucheJESaimanYFriedmanSLHepatic stellate cells and liver fibrosisCompr Physiol2013341473149224265236
- ShenMChenKLuJProtective effect of astaxanthin on liver fibrosis through modulation of TGF-beta1 expression and autophagyMediators Inflamm2014201495450224860243
- HayashiSItohAIsodaKKondohMKawaseMYagiKFucoidan partly prevents CCl4-induced liver fibrosisEur J Pharmacol2008580338038418068155
- TungHLeeFWangSThe beneficial effects of P2X7 antagonism in rats with bile duct ligation-induced cirrhosisPLoS One2015105e124654
- HongSWJungKHLeeHSSuppression by fucoidan of liver fibrogenesis via the TGF-beta/Smad pathway in protecting against oxidative stressBiosci Biotechnol Biochem20117583384021597183
- MontfortIPerez-TamayoRCollagenase in experimental carbon tetrachloride cirrhosis of the liverAm J Pathol1978922411420209692
- TagCGSauer-LehnenSWeiskirchenSBile duct ligation in mice: induction of inflammatory liver injury and fibrosis by obstructive cholestasisJ Vis Exp20151096e52438
- AshfordTPPorterKRCytoplasmic components in hepatic cell lysosomesJ Cell Biol19621219820213862833
- GordyCHeYWThe crosstalk between autophagy and apoptosis: where does this lead?Protein Cell201231172722314807
- ChengPChenKXiaYHydrogen sulfide, a potential novel drug, attenuates concanavalin A-induced hepatitisDrug Des Devel Ther2014812771286
- MaoYWangJYuFGhrelin reduces liver impairment in a model of concanavalin A-induced acute hepatitis in miceDrug Des Devel Ther2015953855396
- JiaGSowersJRAutophagy: a housekeeper in cardiorenal metabolic health and diseaseBiochim Biophys Acta20151852221922424984281
- DoriaAGattoMPunziLAutophagy in human health and diseaseN Engl J Med201336819184523656659
- LiJXiaYLiuTProtective effects of astaxanthin on ConA-induced autoimmune hepatitis by the JNK/p-JNK pathway-mediated inhibition of autophagy and apoptosisPLoS One2015103e120440
- ShenMLuJDaiWEthyl pyruvate ameliorates hepatic ischemia-reperfusion injury by inhibiting intrinsic pathway of apoptosis and autophagyMediators Inflamm2013201346153624453420
- ChengPWangFChenKHydrogen sulfide ameliorates ischemia/reperfusion-induced hepatitis by inhibiting apoptosis and autophagy pathwaysMediators Inflamm2014201493525124966472
- WangCChenKXiaYN-acetylcysteine attenuates ischemia-reperfusion-induced apoptosis and autophagy in mouse liver via regulation of the ROS/JNK/Bcl-2 pathwayPLoS One201499e10885525264893
- Hernandez-GeaVGhiassi-NejadZRozenfeldRAutophagy releases lipid that promotes fibrogenesis by activated hepatic stellate cells in mice and in human tissuesGastroenterology2012142493894622240484
- LiuMHeYZhangJEffect of autophagy inhibitor 3-methyladenine on proliferation and activation of hepatic stellate cellsXi Bao Yu Fen Zi Mian Yi Xue Za Zhi2013298809812 Chinese23948405
- FittonJHTherapies from fucoidan; multifunctional marine polymersMar Drugs20119121731176022072995
- KwakJFucoidan as a marine anticancer agent in preclinical developmentMar Drugs201412285187024477286
- SenthilkumarKManivasaganPVenkatesanJKimSBrown seaweed fucoidan: biological activity and apoptosis, growth signaling mechanism in cancerInt J Biol Macromol20136036637423817097
- PanRLXiangLXWangPLow-molecular-weight fibroblast growth factor 2 attenuates hepatic fibrosis by epigenetic down-regulation of delta-like1Hepatology20156151708172025501710
- TsukadaSParsonsCJRippeRAMechanisms of liver fibrosisClin Chim Acta20063641–2336016139830
- LimJLeeSKimTFucoidan from Fucus vesiculosus protects against alcohol-induced liver damage by modulating inflammatory mediators in mice and HepG2 cellsMar Drugs20151321051106725690093
- LiCGaoYXingYZhuHShenJTianJFucoidan, a sulfated polysaccharide from brown algae, against myocardial ischemia-reperfusion injury in rats via regulating the inflammation responseFood Chem Toxicol20114992090209521645579
- Senties-GomezMDGalvez-GastelumFJMeza-GarciaEArmendariz-BorundaJHepatic fibrosis: role of matrix metalloproteases and TGFbetaGac Med Mex20051414315322 Spanish16164129
- JacksonPLXuXWilsonLHuman neutrophil elastase-mediated cleavage sites of MMP-9 and TIMP-1: implications to cystic fibrosis proteolytic dysfunctionMol Med2010165–615916620111696
- FuWWangYJinZLosartan alleviates renal fibrosis by down-regulating HIF-1alpha and up-regulating MMP-9/TIMP-1 in rats with 5/6 nephrectomyRen Fail201234101297130423006043
- MoonHJLeeSHKuMJFucoidan inhibits UVB-induced MMP-1 promoter expression and down regulation of type I procollagen synthesis in human skin fibroblastsEur J Dermatol200919212913419129083
- MurrowLDebnathJAutophagy as a stress-response and quality-control mechanism: implications for cell injury and human diseaseAnnu Rev Pathol2013810513723072311
- LiJWangFXiaYAstaxanthin pretreatment attenuates hepatic ischemia reperfusion-induced apoptosis and autophagy via the ROS/MAPK pathway in miceMar Drugs20151363368338726023842
- KwantenWJMartinetWMichielsenPPFrancqueSMRole of autophagy in the pathophysiology of nonalcoholic fatty liver disease: a controversial issueWorld J Gastroenterol201420237325733824966603
- OsnaNAThomesPGDonohue JrTMInvolvement of autophagy in alcoholic liver injury and hepatitis C pathogenesisWorld J Gastroenterol201117202507251421633655
- WangJKangRHuangHHepatitis C virus core protein activates autophagy through EIF2AK3 and ATF6 UPR pathway-mediated MAP1LC3B and ATG12 expressionAutophagy201410576678424589849
- SasakiMYoshimura-MiyakoshiMSatoYNakanumaYA possible involvement of endoplasmic reticulum stress in biliary epithelial autophagy and senescence in primary biliary cirrhosisJ Gastroenterol201550998499525552342
- HeWWangBYangJChloroquine improved carbon tetrachloride-induced liver fibrosis through its inhibition of the activation of hepatic stellate cells: role of autophagyBiol Pharm Bull20143791505150925177034
- HeYZhuJHuangYGaoHZhaoYAdvanced glycation end product (AGE)-induced hepatic stellate cell activation via autophagy contributes to hepatitis C-related fibrosisActa Diabetol201552595996926002589
- ThoenLFGuimaraesELGrunsvenLAAutophagy: a new player in hepatic stellate cell activationAutophagy20128112612822082960
- GhavamiSCunningtonRHGuptaSAutophagy is a regulator of TGF-β1-induced fibrogenesis in primary human atrial myofibroblastsCell Death Dis201563e169625789971
- PanCCKumarSShahNEndoglin regulation of Smad2 function mediates beclin1 expression and endothelial autophagyJ Biol Chem201529024148841489225931117
- GressnerAMWeiskirchenRModern pathogenetic concepts of liver fibrosis suggest stellate cells and TGF-beta as major players and therapeutic targetsJ Cell Mol Med2006101769916563223
- MassagueJBlainSWLoRSTGFbeta signaling in growth control, cancer, and heritable disordersCell2000103229530911057902
- SchnablBKweonYOFrederickJPWangXFRippeRABrennerDAThe role of Smad3 in mediating mouse hepatic stellate cell activationHepatology20013418910011431738
- RaoCLinSLRuanWJWenHWuDJDengHHigh expression of IGFBP7 in fibroblasts induced by colorectal cancer cells is co-regulated by TGF-beta and Wnt signaling in a Smad2/3-Dvl2/3-dependent mannerPLoS One201491e8534024427302
- SamarakoonRChitnisSSHigginsSPHigginsCEKrepinskyJCHigginsPJRedox-induced Src kinase and caveolin-1 signaling in TGF-beta1-initiated SMAD2/3 activation and PAI-1 expressionPLoS One201167e2289621829547
- NikoletopoulouVMarkakiMPalikarasKTavernarakisNCrosstalk between apoptosis, necrosis and autophagyBiochim Biophys Acta20131833123448345923770045