
Abstract
Ribonucleic acid splicing is a crucial process to create a mature mRNA molecule by removing introns and ligating exons. This is a highly regulated process, but any alteration in splicing factors, splicing sites, or auxiliary components affects the final products of the gene. In diffuse large B-cell lymphoma, splicing mutations such as mutant splice sites, aberrant alternative splicing, exon skipping, and intron retention are detected. The alteration affects tumor suppression, DNA repair, cell cycle, cell differentiation, cell proliferation, and apoptosis. As a result, malignant transformation, cancer progression, and metastasis occurred in B cells at the germinal center. B-cell lymphoma 7 protein family member A (BCL7A), cluster of differentiation 79B (CD79B), myeloid differentiation primary response gene 88 (MYD88), tumor protein P53 (TP53), signal transducer and activator of transcription (STAT), serum- and glucose-regulated kinase 1 (SGK1), Pou class 2 associating factor 1 (POU2AF1), and neurogenic locus notch homolog protein 1 (NOTCH) are the most common genes affected by splicing mutations in diffuse large B cell lymphoma.
Introduction
Non-Hodgkin lymphoma is a heterogeneous group of cancers that starts during immune system differentiation.Citation1 Recent reports show that more than 100 different lymphoma types have been identified.Citation2 These lymphomas originate from cells of the immune system such as B cells, T cells, and dendritic cells. B-cell lymphomas are another type of cancer that develops from clonal expansion and subsequent B-cell invasion of immune organs.Citation3 The heterogeneity is derived from different stages of mature B-cell differentiation. Diffuse large B-cell lymphoma (DLBCL) is the most common lymphoid malignancy in adults that starts from the germinal center.Citation4,Citation5
Genetic alterations such as chromosomal translocations, sporadic somatic mutations, and copy number alterations, deletions, and amplifications of genes have been reported in the pathogenesis of DLBCL.Citation5–8 In addition, missense mutations such as immune-related epigenetic modifications that affect the function oncogenes have been involved in DLBCL genesis.Citation9–11 Furthermore, current evidence reveals that mutations during RNA splicing play a role in DLBCL pathogenesis. Thus, the identification of mutations in RNA splicing is important for the diagnosis, treatment, and prognosis of DLBCL.Citation12,Citation13
Human genes are highly complex; it contains coding sequence exons and non-coding sequence intron. Non-coding sequence should be removed from primary transcript through RNA splicing process.Citation14 RNA splicing is the process by which a precursor mRNA transcript is transformed into a mature mRNA. It is a crucial process to create an mRNA molecule by removing introns and subsequently ligating exons that direct the synthesis of the protein during translation.Citation13 The introns are removed from the transcript by cleavage at splice site (the 5′ and 3′ ends of intron). This process most commonly begins at dinucleotide GU and AG site at 5′ end and 3′ end of primary transcript, respectively ().
Figure 1 A schematic representative pre-mRNA, coding and non-coding sequence and splicing site.

The RNA splicing process contains more than five small nuclear ribonucleoproteins (snRNPs) and trans and cis-splicing elements such as more than 200 proteins (at the splice acceptor, donor, and splice site), branch points (BP), splice enhancers, splice silencers, and splicing reaction catalysts. A large RNA-protein complex called the spliceosome recognizes auxiliary elements to promote the excision of introns from the nuclear pre-mRNA.Citation12,Citation13
Based on their involvement, spliceosome is classified as major and minor. About 99.5% of introns are recognized and excised by major spliceosome, which is known as U2 dependent spliceosome (U1, U2, U4, U6, and U5 snRNPs).Citation15,Citation16 The major splicing reaction occurs in order to recognized relatively poorly conserved sequence of pre-mRNA at the 5’ and 3’ ends. On the other hand, remaining about 05% of introns are excised by minor spliceosome, it is called U12 dependent (U11, U12, U5, U4atac, and U6atac).Citation16,Citation17 They target a rare group of introns and highly conserved sequences at the 5’ and 3’ ends. Both of splicing reactions are important to formation of mature mRNA.Citation15,Citation18
The alteration during the RNA splicing could potentially alter mRNA maturation and the subsequent production of protein.Citation13,Citation19 Recurrent genetic abnormalities due to splicing mutations have been reported in all forms of myeloid neoplasms, several lymphoid neoplasms, and solid tumors. These abnormalities are implicated during pre-RNA splicing.Citation19 Furthermore, epigenetic changes can sometimes be linked to RNA splicing.Citation17
The abnormalities detected in DLBCL due to RNA splicing are mutant splice sites, aberrant alternative splicing site, exon skipping, and intron retention.Citation3,Citation4,Citation20 Alternative splicing is physiologic process, which occurs in most human gene and creates transcript variability. It is also allowing formation of various proteins from single transcript.Citation21 However, sometimes it leading to pathologic conditions and induces human diseases such DLBCL.Citation20
RNA splicing helps to regulate gene expression for cellular proliferation, survival, and differentiation.Citation22 Nevertheless, mutations in the RNA splicing could potentially affect the maturation of mRNA for most genes and the subsequent production of protein.Citation13 Most of these mutations affect the tumor suppressor activities and DNA repair activities of different genes.Citation23–26 Currently, many splicing site gene mutations in B-cell lymphomas are reported.Citation13 Mutated B cell has a high proliferation rate, a low differentiation rate, and a long lifetime, which leads it to malignant transformation.Citation27
Even though, different scholars published original articles related with splicing site mutations in DLBCL, there is scarcity of a comprehensive review which shows splicing mutation as a cause of DLBCL. Thus, this review tried to discuss the general mechanisms of splicing mutation and splicing regulation. In addition, the review elucidates the role of different splicing mutation in DLBCL pathogenesis. In order to achieve the objective, the articles were identified through searching of the literature published in English using National Library of Medicine, PubMed, Google Scholar, and Google databases. Articles already identified in the references were also manually searched and included. After article identified, imported to end note version 8.1 and exported to Microsoft word for citation.
RNA Splicing Regulation
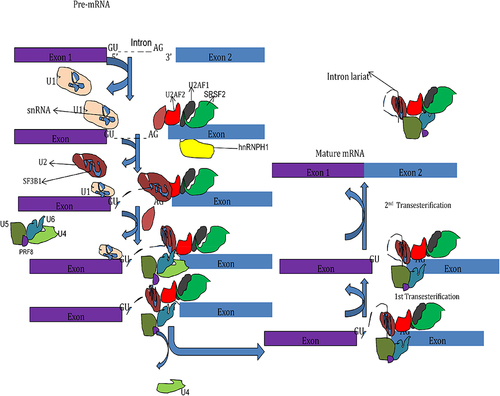
Ribonucleic acid splicing is a nuclear process catalyzed by the spliceosomes. It consists of snRNPs (U1, U2, U4, U5, and U6), over 200 related proteins, and other auxiliary components.Citation28 The first step in the process of RNA splicing is the binding of U1 snRNPs to the 5’ splice site. Splicing factor 1 then binds the BP. After that, the U2 auxiliary factor (U2AF) complex binds the polypyrimidine tract and 3’ splice site. Binding of U2AF either strengthens or repels the recruitment of the spliceosome complex to the splice site.Citation19,Citation28 These U2AF include members of the serine/arginine (SR) protein family that promote splicing by recognizing specific splicing sequences in pre-mRNA called exonic and intronic splicing enhancers. Conversely, heterogeneous nuclear ribonucleoproteins (hnRNPs) suppress splicing by interacting with exonic and intronic splicing silencers.Citation3 The splicing process consists of two sequential enzymatic steps named transesterification reactions. The BP nucleotide helps form the intron lariat through nucleophilic attack. After this, a ubiquitin-specific peptidase 59 (Usp59)-mediated attack on the ubiquitin-specific peptidase 39 (Usp39) occurred and led to the removal of the intron lariat from RNA for the formation of the spliced RNA product ().Citation3,Citation19,Citation28
Figure 2 Splicing regulation mechanism.
Mechanism of Splicing Site Mutation
The splicing process is a highly regulated one, but any alteration in splicing factors, splicing sites, or auxiliary components affects the final products of a gene.Citation3,Citation17 As a result of the alteration in mRNA, malignant transformation, cancer progression, and metastasis occur.Citation21 Splicing mutations may be formed by different mechanisms.Citation3,Citation13,Citation19,Citation29,Citation30 One of the mechanisms is related to cis-acting elements in RNA that act as an anchor for trans-acting factors to produce functional RNA transcripts. Mutations in cis-acting elements can lead to exon suppression, exon inclusion, blocking the binding of snRNPs, exon skipping, and intron retention. Consequently, the final product of mRNA is altered, which codes for tumor suppressors, oncogene inactivation, and DNA repair products.Citation3,Citation13
Another mechanism of splicing site mutation is related to alternative splicing. It is essential physiologically for the production of diverse proteins from a single pre-mRNA.Citation20 However, it can sometimes result in genetic changes such as exon skipping, intron retention, altered 5’ splice, altered 3’ splice, and mutually exclusive splicing.Citation22 These alterations may play an important role in tumor growth through tumor suppressor gene inactivation and oncogene activation, as well as inhibition of cell differentiation.Citation31
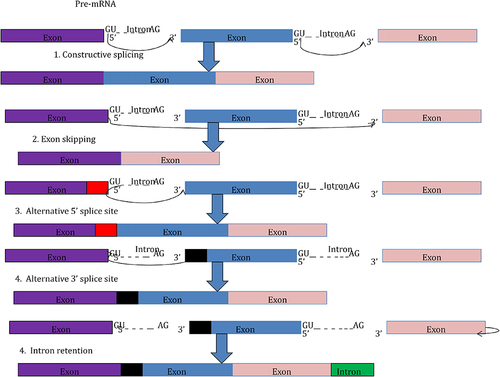
Dysregulation of splicing factors’ expression is another mechanism related to splicing mutations. Spliceosome recognizes intron-exon boundaries and removes the intron.Citation20 Exon skipping and intron retention are caused by mutations in the spliceosome complex.Citation13,Citation19 As a result of the spliceosome mutation, gene expression is dysregulated, and tumor cells proliferate ().Citation32,Citation33
Figure 3 Schematic representation of constitutive and alternative splicing dysregulation.
Splicing Mutation in DLBCL Pathogenesis
B-cell lymphoma is a heterogeneous group of hematological malignancies originating from B cells and accounts for up to 35% of non-Hodgkin’s lymphomas.Citation6,Citation27 DLBCL has a global annual incidence of more than 100,000 cases. As a result, identification of the cause, pathogenesis, prognosis, and presentation is critical for improving the diagnostic process, classification, outcome stratification, and personalized therapy. As a result, splicing site mutation also has a contribution to DLBCL diagnosis, pathogenesis, and prognosis.Citation5–8
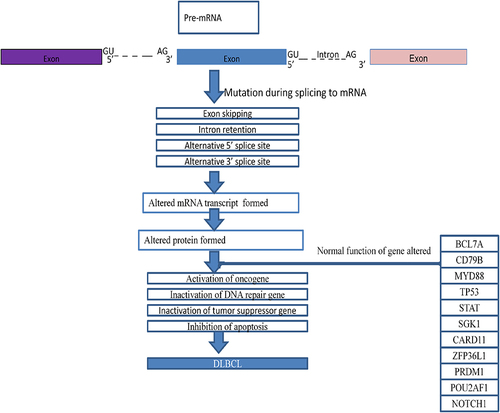
Mutations during splicing affect different genes, which contribute to the inactivation of oncogenes,Citation34 the activation of tumor suppressor genes,Citation35 the activation of DNA repair genes, and the inhibition of apoptosis.Citation27,Citation30 Splicing mutations most commonly affect B-cell lymphoma 7 protein family member A (BCL7A), cluster of differentiation 79B (CD79B), myeloid differentiation primary response gene 88 (MYD88), tumor protein P53 (TP53), signal transducer and activator of transcription (STAT), serum- and glucose-regulated kinase 1 (SGK1), caspase recruitment domain-containing protein 11 (CARD11), zinc finger proteins 36L1 (ZFP36L1), PR domain zinc finger protein 1 (PRDM1), pou class 2 associating factor 1 (POU2AF1), and neurogenic locus notch homolog protein 1 (NOTCH1) ().Citation6,Citation35,Citation36
Figure 4 Schematic representation of pathological mechanisms DLBCL in splicing mutation.
Role of Splice Site Mutation on BCL7A Gene
The BCL7A gene is located on chromosome 12 and is affected most commonly by a splicing mutation.Citation6 Its products play a role in B cell activation and tumor suppressor function. In the BCL7A gene, three different splicing site mutations, such as mutant splice site, cryptic splice site, and exon skipping, were found. So, splicing mutations in the BCL7A gene inactivate and alter its normal function.Citation5–7 The tumor suppressor function is one of the functions of genes affected by splice site mutations. The gene’s function is altered due to splice site mutations of the portion of the BCL7A protein’s amino terminal domain, which coded by the BCL7A gene.Citation7 As a result, binding of the BCL7A protein to the Switch/Sucrose non-fermentable complex was affected. The loss of Switch/Sucrose non-fermentable complex integrity suppresses the differentiation of B cells into plasma and memory cells. B cell activation is another function of a gene affected by splice site mutation. This is because BCL7A restoration induces transcriptomic changes in genes involved in B-cell activation. Furthermore, evidence suggests that the BCL7A gene is involved in oncogene inactivation.Citation3,Citation36
Role of Splice Site Mutation on MYD88, CARD11, CD79B and MYD88 Genes
MYD88, CARD11, and CD79B genes are among those whose functions are altered by RNA splicing mutations. Splicing mutations such as exon skipping in MYD88, intron retention in CARD11 and CD79B, and a mutant splice site in CARD1 affect the final product of the gene.Citation23,Citation30 Because there is participation in the inactivation of the nuclear factor kappa-light-chain-enhancer of activated B cells (NF-B) signaling pathway, alteration of the product of these genes plays a significant role in DLBCL pathogenesis.Citation25 The NF-kB signaling pathway has negative feedback for B cell differentiation and positive feedback for B cell proliferation.Citation6 According to the report of available literature, splicing mutations in MYD88, CARD11, and CD79B gene products are the driving force for activating the NF-κB pathway that is implicated in the pathogenesis of DLBCL.Citation37–39 Similarly, splicing factor mutations called spliceosome mutations in pre-mRNAs trigger exon skipping and intron retention. These changes are caused by mutations in the splicing factor 3A and 3B genes.Citation40,Citation41 These shorten the MyD88 gene transcript, which is essential to induce toll-like receptor signaling in cells and the differentiation of B cells. Lack of differentiation may cause DLBCL.Citation40,Citation42 In contrast, two studies found no link between splicing mutations and the MYD88 gene in the pathogenesis of DLBCL.Citation30,Citation34
Role of Splice Site Mutation on STAT Gene
The splicing mutations might be linked to changes in STAT gene products.Citation35 According to the literature, cryptic splice sites, intron retention, and exon skipping all play a role in STAT gene product alteration. Cryptic splicing leads to a lost serine codon at the acceptor sites, while exon skipping and intron retention lead to premature stop codons.Citation43,Citation44 These mutations affect STAT3’s normal function because it mediates the expression of various genes that play an important role in cell proliferation, survival, and differentiation. However, abnormal expression of STAT3 products promotes malignant transformation and tumor progression through oncogenic gene expression and blocking cell differentiation.Citation43
Role of Splice Site Mutation on SGK1, PRDM1, TP53, ZFP36L1, POU2AF1 and NOTCH1 Genes
Splicing mutations such as exon skipping, mutant splice sites, intron retention, cryptic splice sites, and aberrant alternative splicing are also involved in the SGK1 gene. Available data shows that SGK1 gene products are involved in the regulation of cell apoptosis and the cell cycle. So, alteration of this gene loses its tumor suppressor role and restricts B cell differentiation at the germinal center.Citation45,Citation46 Similarly, splicing mutations in the PR domain containing 1 with a zinc finger domain (PRDM1) affect B cell differentiation at the germinal center. Splicing mutations detected in PRDM1 are mutant splice site and exon skipping. It cause premature translation termination and inhibition of B cell differentiation at the germinal center, respectively.Citation6,Citation47,Citation48
Moreover, splicing mutations affect the products of a tumor suppressor gene called TP53. The gene mediates various stimuli for stressed cells through controlling the expression of target genes. In addition, splicing mutations altered the p53 protein, which participates in DNA repair and apoptosis.Citation49 The reports of different publications revealed that the splicing mutations found in the TP53 gene are aberrant alternative splicing, mutant splice sites, and exon skipping. As a result, the tumor suppressor, DNA repair, and apoptotic functions of genes were altered.Citation4,Citation27,Citation31,Citation35,Citation49–51 Similarly, splicing mutations such as exon skipping and mutant splice sites affect the product of ZFP36L1 and POU2AF1 genes that block B cell differentiation at the germinal center.Citation52,Citation53 Furthermore, exon skipping and mutant splice sites altered the apoptotic and tumor suppressor functions of POU2AF1 gene products.Citation6,Citation54
Furthermore, splicing mutations affect the NOTCH1 gene, which encodes a transmembrane protein for regulating cell differentiation, proliferation, and apoptosis. So, altered splicing of NOTCH gene transcripts may affect B-cell lineage development and cause malignant transformations.Citation55 So, altered splicing of NOTCH genes transcripts may affect B-cell lineage development and cause malignant transformations.Citation54 The findings of various studies show that the splicing mutations detected in the NOTCH gene transcript are aberrant alternative splicing and skipping of exon.Citation54,Citation55
Role of Splicing Mutation on SF1, CD58, TNFAIP3, NFKBIE, STXBP2, NFKBI, STXBP, CYLD and PTPN1 Genes
The splicing factor 1 (SF1) gene is one splicing factor gene out of 66 splicing factor coding genes. It recognizes the 3’ splice site by binding the BP of intron.Citation56 In addition, it has played a critical role in the retention of nuclear pre-mRNA during time-alternative splicing. Recently, there has been an increase in the evidence supporting mutations of the SF1 gene related to various cancers, but less in lymphoma.Citation57 Only a couple of studies report mutations in SF1 in DLBCL.Citation56,Citation58 In addition, mutant splice sites and exon skipping altered CD 58 gene products’ normal function. The loss of CD 58 receptors on B cells due to mutation allows tumor cells to avoid immune recognition.Citation51,Citation59 In addition, intron retention and aberrant splicing inhibit the expression of the CD 58 receptor on B cells. As a result, abnormal B cells escaped recognition by cytotoxic T cells and caused overproliferation of abnormal cells.Citation6,Citation59
As reported in different studies, splice site mutations in the TNFAIP3, NFKBIE, and STXBP2 genes play a significant role in DLBCL pathogenesis.Citation3,Citation6,Citation36,Citation60 The TNFAIP3 gene encodes a ubiquitin-modifying enzyme that is involved in the termination of NF-kB signaling pathways because its activation is important for malignant cell proliferation and blocks B cell differentiation.Citation39 Splicing mutation is one of the mechanisms involved in TNFAIP3 gene alteration. As evidence shows, mutant splicing sites and exclusion of exons are splicing mutations involved in the TNFAIP3 gene mutation.Citation8 As a result, the negative feedback for NF-Kb pathways is altered.Citation6,Citation30 Similarly, when the NFKBIA gene was mutated, the NF-B signaling pathway was activated because it codes for IkBs proteins that inhibit NF-kB dimers.Citation6,Citation38 Similarly, during mutation of NFKBIA gene, NF-κB signaling pathway activated because it codes IκBs proteins that inhibits NF-kB dimers.Citation3,Citation39 There are defects in the NFKBIA gene due to the exclusion of an exon that inhibits B cell differentiation at the germinal center, according to the literature.Citation6,Citation8,Citation27 Studies show that the STXBP2 gene is mutated due to exon skipping, but the details of the mechanisms need to be investigated further.Citation44,Citation61
Splicing mutations such as exon skipping, cryptic splices at acceptor sites of exons, and mutant splicing sites were detected in CYLD gene transcripts. These splicing mutations inhibit the tumor suppressor role of the gene through inactivation of NF-Kb signaling pathways. The pathway’s activation may contribute to malignant cells’ ability to proliferate.Citation39,Citation62 Splicing mutations like exon skipping are found in protein tyrosine phosphatase, nonreceptor type 1 (PTPN1) gene transcripts. This gene has crucial negative or positive regulators of JAK/STAT signaling. These functions of genes contribute to preventing the proliferation, differentiation, and survival of tumor cells.Citation63,Citation64 Splicing mutations altered the normal function of the PTPNI gene, leading to enhanced proliferation and survival of malignant B cells and blocking the differentiation of B cells.Citation63
Role of Splicing Mutation on ZEB2, CD70, FOXP1, EBF1 and CD37 Genes
Recently, mutant splice sites were detected in zinc finger E-box-binding homeobox 2 (ZEB2) transcripts. Consequently, the effects of ZEB2 on the growth, migration, invasion, cell cycle distribution, and apoptosis of B cells were explored.Citation6 As hypothesised by different scholars, alteration of ZEB2’s role may inhibit the differentiation of B cells and promote malignant transformation and metastasis.Citation6,Citation51,Citation60 Similarly, malignant transformation and metastasis are detected in the alteration of cluster of differentiation 70 (CD70). This is because, as evidence shows, its tumor suppressor and anti-apoptotic roles are affected due to exon skipping. This may lead to DLBCL formation.Citation6,Citation36 In contrast, one study found no significant link between splicing mutations and changes in CD70 expression.Citation65 It is convincing that CD70 only serves as a co-stimulator during the activation of a signal transduction pathway, and that their function is compensated for by the function of other co-stimulators.Citation3,Citation51
Forkhead box protein P1 (FOXP1) is a master regulator of stem and progenitor cells.Citation65 However, aberrant expression of the oncogenic transcription factor is due to a splicing mutation in FOXP1 detected commonly in DLBCL.Citation6,Citation39 This contributes to tumor cell survival because intron retention during RNA splicing leads to sphingosine-1-phosphate receptor 2 being repressed in the germinal center B-cell. It is important to activate downstream signaling pathways to induce apoptosis, restrict tumor growth, and inactivate oncogenes.Citation65 Similarly, early B-cell factor 1 (EBF1) functions as a regulator of committed B cell progenitors and B cell-specific gene expression programs. Despite the fact that splicing mutations known as exon skipping altered its function, recent studies revealed that splicing mutations affect efficient B cell commitment and differentiation.Citation6,Citation66,Citation67
Splicing mutations such as exon skipping affect cell cycle regulation and the apoptosis role in the proto-oncogene serine/threonine-protein kinase 1. This promotes malignant cell growth and survival through cell cycle dysregulation and inhibition of apoptosis in B cells.Citation68,Citation69 Similarly, due to exon skipping in pre-RNA, the apoptotic role of cluster of differentiation 37 (CD37) is affected. It causes malignant B cells to evade immune cells.Citation70
Role of Splicing Mutation on Others Minor Genes
According to recent findings, B cells in DLBCL fail to express cell-surface molecules required for tumor cell recognition by immune effector cells.Citation5,Citation6,Citation71 It may be due to splicing mutations (exon skipping) that inactivate the β2-microglobulin (B2M) gene. Inactivation of the gene prevents the expression of the cell-surface human leukocyte antigen 1 complex that is necessary for the recognition of tumor cells by CD8+ cytotoxic T cells. As a result, malignant B cells escape from immune surveillance and progress to DLBCL.Citation59,Citation71
In addition, ETS translocation variant 6 (ETV6), cluster of differentiation 83 (CD83), tumor necrosis factor receptor superfamily member 14 (TNFRSF14), myosin-Ie (MYO1E), dual specificity protein phosphatase 2 (DUSP2), lysosomal-associated transmembrane protein 5 (LAPTM5), histone-lysine N-methyltransferase 2D (KMT2D), and T-cell leukemia/lymphoma protein 1A (TCL1A) are less frequently detected genes in DLBCL. To identify the detailed mechanisms involved in the pathogenesis of DLBCL, they require further investigationCitation4,Citation6,Citation27,Citation36,Citation69,Citation70 ().
Table 1 Summarizes the Role of Splicing Mutation in Different Genes in DLBCL as Well as Their Mechanisms
Splicing Mutation and Targeted Drugs for the Treatment of DLBCL
Diffuse large B-cell lymphoma is a heterogeneous group of aggressive lymphoma that has specific clinical course and response to therapy.Citation75,Citation76 As a result, DLBCL is still remained challenging for physicians in developing specific targeted treatments for patients care. Recurrent mutations such as splicing mutation associated with poor clinical outcomes in DLBCL.Citation77 The drugs targeted to RNA binding regulatory proteins have predictive prognostic values to treat patients with DLBCL.Citation76,Citation78
As evidences show in the past decades, immunochemotheraphy such as rituximab plus cyclophosphamide, vincristine, etoposide, brentuximab, bleomycin, doxorubicin, and prednisone have a significant role in DLBCL patients to extend survival rate and cure rate as well as decrease relapse rate.Citation77,Citation79–81 Those drugs target RNA binding protein during splicing process. RNAs splicing is crucial process for the formation of mature mRNA which codes specific proteins. The proteins formed from translation regulate cell apoptosis, differentiation, DNA repair and inactivate oncogene.Citation35 Hence, the immunochemotheraphy drugs may target RNA binding factors and prevent the occurrence of mutation. However, prognosis of some patients still refractory to standard treatment. Thus, in order to improve prognostic outcomes, new treatment option is under the preclinical trials.Citation56,Citation77,Citation79
Furthermore, currently some drugs such as inhibitor of splice factor kinases (dasatinib), inhibitor of NF-kB pathway (ibrutinib), inhibitor of JAK-STAT pathway (niacinamide) and inhibitors bruton tyrosine kinase pathway (calabrutinib, tirabrutinib, and spebrutinib) on pre-clinical stage are reported as novel treatment option to treat patient with DLBCL.Citation75,Citation76,Citation82 The pathways targeted by drugs are important for malignant B cell proliferation and differentiation. Consequently, the drugs inactivate the malignant B cells proliferation, differentiation and induce malignant B cell apoptosis at germinal center during RNA splicing. Based on subtypes of DLBCL, providing and applying specific therapeutic strategies is important to treat patients with DLBCL.Citation76,Citation83
Conclusion
In conclusion, different types of splicing mutations were detected in DLBCL. These mutations are mutant splice site, aberrant alternative splicing, exon skipping and intron retention. Most of these RNA splicing mutations affect normal function of different genes. The genes most frequently affected by splicing mutation in DLBCL are BCL7A, CD79B, SF1, MYD88, TP53, STAT, SGK1, ZFP36L1 and POU2AF1. As a result of alteration in different gene by RNA splicing mutation, malignant transformation, cancer progression, and metastasis occur in DLBCL. Although, most of the literature published is focused on only the role of splicing mutation for DLBCL; risk factors, prevention mechanisms, and treatment options for these mutations were not well discussed. As a recommendation, it is important for researchers to find risk factors, prevention mechanisms, prognostic role and treatment options of RNA splicing mutation in DLBCL.
Abbreviations
BCL7A, B-cell Lymphoma 7 Protein Family Member; BP, branch points; CARD11, caspase recruitment domain-containing protein 11; CD, cluster of differentiation; CD79B, cluster of differentiation 79B; cHL, classical Hodgkin lymphoma; CYLD, cylindromatosis lysine differentiation gene; DLBCL: diffuse large B cell lymphoma; MYD88, myeloid differentiation primary response gene 88; NF-κB, nuclear factor kappa B cells; NOTCH1, neurogenic locus notch homolog protein 1; Pre-RNA, premature ribonucleic acid; SF1, splicing factor 1; snRNPS, small nuclear ribonucleoproteins; STAT, signal transducer and activator of transcription; TP53, tumor protein P53; U2AF, U2 auxiliary Factor.
Disclosure
The authors report no conflicts of interest in this work.
Acknowledgment
We would like to acknowledge the authors of the studies included in this review.
References
- de Leval L, Jaffe ES. Lymphoma classification. Cancer J. 2020;26(3):176–185. doi:10.1097/PPO.0000000000000451
- Swerdlow SH, Campo E, Pileri SA, et al. The 2016 revision of the World Health Organization classification of lymphoid neoplasms. Blood. 2016;127(20):2375–2390. doi:10.1182/blood-2016-01-643569
- Moghanibashi M, Mohamadynejad P. Splicing in Cancer. hematologica. 2022;14(23):214.
- Asmar F, Punj V, Christensen J, et al. Genome-wide profiling identifies a DNA methylation signature that associates with TET2 mutations in diffuse large B-cell lymphoma. Haematologica. 2013;98(12):1912. doi:10.3324/haematol.2013.088740
- Chapuy B, Stewart C, Dunford AJ, et al. Molecular subtypes of diffuse large B cell lymphoma are associated with distinct pathogenic mechanisms and outcomes. Nat med. 2018;24(5):679–690. doi:10.1038/s41591-018-0016-8
- Andrades A, Álvarez-Pérez JC, Patiño-Mercau JR, Cuadros M, Baliñas-Gavira C, Medina PP. Recurrent splice site mutations affect key diffuse large B-cell lymphoma genes. Blood. 2022;139(15):2406–2410. doi:10.1182/blood.2021011708
- Reddy A, Zhang J, Davis NS, et al. Genetic and functional drivers of diffuse large B cell lymphoma. Cell. 2017;171(2):481–94.e15. doi:10.1016/j.cell.2017.09.027
- Schmitz R, Wright GW, Huang DW, et al. Genetics and pathogenesis of diffuse large B-cell lymphoma. N Engl J Med. 2018;378(15):1396–1407. doi:10.1056/NEJMoa1801445
- Wang X, Hong Y, Meng S, et al. A novel immune-related epigenetic signature based on the transcriptome for predicting the prognosis and therapeutic response of patients with diffuse large B-cell lymphoma. Clin Immun. 2022;243:109105. doi:10.1016/j.clim.2022.109105
- Zhang S, Zhang T, Liu H, et al. Tracking the evolution of untreated high‐intermediate/high‐risk diffuse large B‐cell lymphoma by circulating tumour DNA. Br J Haematol. 2022;196(3):617–628. doi:10.1111/bjh.17894
- Zhang H, Lu Y, Zhang T, et al. PIM1 genetic alterations associated with distinct molecular profiles, phenotypes and drug responses in diffuse large B‐cell lymphoma. Clin Transl Med. 2022;12(4). doi:10.1002/ctm2.808
- Hahn CN, Venugopal P, Scott HS, Hiwase DK. Splice factor mutations and alternative splicing as drivers of hematopoietic malignancy. Immunol Rev. 2015;263(1):257–278. doi:10.1111/imr.12241
- Ebert B, Bernard OA. Mutations in RNA splicing machinery in human cancers. N Engl J Med. 2011;365(26):2534–2535. doi:10.1056/NEJMe1111584
- Saez B, Walter MJ, Graubert TA. Splicing factor gene mutations in hematologic malignancies. Blood. 2017;129(10):1260–1269. doi:10.1182/blood-2016-10-692400
- Akinyi MV, Frilander MJ. At the intersection of major and minor spliceosomes: crosstalk mechanisms and their impact on gene expression. Front Genet. 2021;12:700744. doi:10.3389/fgene.2021.700744
- Chen W, Moore MJ. Spliceosomes. Curr. 2015;25(5):R181–R183. doi:10.1016/j.cub.2014.11.059
- Kitamura K, Nimura K. Regulation of RNA splicing: aberrant splicing regulation and therapeutic targets in cancer. Cells. 2021;10(4):923. doi:10.3390/cells10040923
- Olthof AM, White AK, Mieruszynski S, et al. Disruption of exon-bridging interactions between the minor and major spliceosomes results in alternative splicing around minor introns. Nucleic Acids Res. 2021;49(6):3524–3545. doi:10.1093/nar/gkab118
- Chen S, Benbarche S, Abdel-Wahab O. Splicing factor mutations in hematologic malignancies. Blood. 2021;138(8):599–612. doi:10.1182/blood.2019004260
- Gurnari C, Pagliuca S, Visconte V. Alternative splicing in myeloid malignancies. Biomedicine. 2021;9(12):1844.
- Leivonen S, Taskinen M, Cervera A, et al. Alternative splicing discriminates molecular subtypes and has prognostic impact in diffuse large B-cell lymphoma. Blood Cancer j. 2017;7(8):596. doi:10.1038/bcj.2017.71
- Taylor J, Lee SC. Mutations in spliceosome genes and therapeutic opportunities in myeloid malignancies. Genes Chromosom Cancer. 2019;58(12):889–902. doi:10.1002/gcc.22784
- Ngo VN, Young RM, Schmitz R, et al. Oncogenically active MYD88 mutations in human lymphoma. Nature. 2011;470(7332):115–119. doi:10.1038/nature09671
- Schneider M, Schneider S, Zühlke‐Jenisch R, et al. Alterations of the CD58 gene in classical Hodgkin lymphoma. Gene Chromosom Cancer. 2015;54(10):638–645. doi:10.1002/gcc.22276
- Takeuchi T, Yamaguchi M, Kobayashi K, et al. MYD88, CD79B, and CARD11 gene mutations in CD5‐positive diffuse large B‐cell lymphoma. Cancer. 2017;123(7):1166–1173. doi:10.1002/cncr.30404
- Voropaeva EN, Pospelova TI, Voevoda MI, Maksimov VN, Orlov YL, Seregina OB. Clinical aspects of TP53 gene inactivation in diffuse large B-cell lymphoma. BMC Genom. 2019;12(2):35–44. doi:10.1186/s12920-019-0484-9
- Chapuy B, Stewart C, Dunford AJ, et al. Author Correction: molecular subtypes of diffuse large B cell lymphoma are associated with distinct pathogenic mechanisms and outcomes. Nat med. 2018;24(8):1290–1291. doi:10.1038/s41591-018-0097-4
- Wilkinson ME, Charenton C, Nagai K. RNA splicing by the spliceosome. Annu Rev Biochem. 2020;89(1):359–388. doi:10.1146/annurev-biochem-091719-064225
- Zhou J, Chng WJ. Aberrant RNA splicing and mutations in spliceosome complex in acute myeloid leukemia. Stem Cell Investig. 2017;4:6. doi:10.21037/sci.2017.01.06
- Dobashi A. Molecular pathogenesis of diffuse large B-cell lymphoma. JCEH. 2016;56(2):71–78. doi:10.3960/jslrt.56.71
- Jung H, Lee KS, Choi JK. Comprehensive characterisation of intronic mis-splicing mutations in human cancers. Oncogene. 2021;40(7):1347–1361. doi:10.1038/s41388-020-01614-3
- Yang H, Beutler B, Zhang D. Emerging roles of spliceosome in cancer and immunity. Protein Cell. 2021;13(8):559–579.
- Zhang Y, Qian J, Gu C, Yang Y. Alternative splicing and cancer: a systematic review. Signal Transduct Target Ther. 2021;6(1):78. doi:10.1038/s41392-021-00486-7
- Aggarwal V, Das A, Bal A, et al. MYD88, CARD11, and CD79B oncogenic mutations are rare events in the Indian cohort of de novo nodal diffuse large B-cell lymphoma. AIMM. 2019;27(4):311–318. doi:10.1097/PAI.0000000000000585
- Caeser R, Di Re M, Krupka JA, et al. Genetic modification of primary human B cells to model high-grade lymphoma. Natcommun. 2019;10(1):1–16.
- Baliñas-Gavira C, Rodríguez MI, Andrades A, et al. Frequent mutations in the amino-terminal domain of BCL7A impair its tumor suppressor role in DLBCL. Leukemia. 2020;34(10):2722–2735. doi:10.1038/s41375-020-0919-5
- Alizadeh AA, Eisen MB, Davis RE, et al. Distinct types of diffuse large B-cell lymphoma identified by gene expression profiling. Nature. 2000;403(6769):503–511. doi:10.1038/35000501
- Jallades L, Baseggio L, Sujobert P, et al. Exome sequencing identifies recurrent BCOR alterations and the absence of KLF2, TNFAIP3 and MYD88 mutations in splenic diffuse red pulp small B-cell lymphoma. Haematologica. 2017;102(10):1758–1766. doi:10.3324/haematol.2016.160192
- Koens L, Zoutman WH, Ngarmlertsirichai P, et al. Nuclear factor-κB pathway-activating gene aberrancies in primary cutaneous large B-cell lymphoma, leg type. J Invest Dermatol. 2014;134(1):290–292. doi:10.1038/jid.2013.265
- Cardona Gloria Y, Bernhart SH, Fillinger S, et al. Absence of non-canonical, inhibitory MYD88 splice variants in B cell lymphomas correlates with sustained NF-κB signaling. Front Immunol. 2021;12(8):616–651. doi:10.3389/fimmu.2021.616451
- Kishimoto T, Ying B-W, Tsuru S, et al. Molecular clock of neutral mutations in a fitness-increasing evolutionary process. PLoS Genet. 2015;11(7):e1005392. doi:10.1371/journal.pgen.1005392
- Pimentel H, Parra M, Gee S, et al. A dynamic alternative splicing program regulates gene expression during terminal erythropoiesis. Nucleic Acids Res. 2014;42(6):4031–4042. doi:10.1093/nar/gkt1388
- Turton KB, Annis DS, Rui L, Esnault S, Mosher DF. Ratios of four STAT3 splice variants in human eosinophils and diffuse large B cell lymphoma cells. PLoS One. 2015;10(5):e0127243. doi:10.1371/journal.pone.0127243
- Zhu F, Wang KB, Rui L. STAT3 activation and oncogenesis in lymphoma. Cancers. 2019;12(1):19. doi:10.3390/cancers12010019
- Gao J, Sidiropoulou E, Walker I, et al. SGK1 mutations in DLBCL generate hyperstable protein neoisoforms that promote AKT Independence. Blood. 2021;138(11):959–964. doi:10.1182/blood.2020010432
- Hartmann S, Schuhmacher B, Rausch T, et al. Highly recurrent mutations of SGK1, DUSP2 and JUNB in nodular lymphocyte predominant Hodgkin lymphoma. Leukemia. 2016;30(4):844–853. doi:10.1038/leu.2015.328
- Pasqualucci L, Compagno M, Houldsworth J, et al. Inactivation of the PRDM1/BLIMP1 gene in diffuse large B cell lymphoma. JEM. 2006;203(2):311–317. doi:10.1084/jem.20052204
- Mandelbaum J, Bhagat G, Tang H, et al. BLIMP1 is a tumor suppressor gene frequently disrupted in activated B cell-like diffuse large B cell lymphoma. Cancer Cell. 2010;18(6):568–579. doi:10.1016/j.ccr.2010.10.030
- Xu-Monette ZY, Medeiros LJ, Li Y, et al. Dysfunction of the TP53 tumor suppressor gene in lymphoid malignancies. Blood. 2012;119(16):3668–3683. doi:10.1182/blood-2011-11-366062
- Zlamalikova L, Moulis M, Ravcukova B, et al. Complex analysis of the TP53 tumor suppressor in mantle cell and diffuse large B-cell lymphomas. Oncol Rep. 2017;38(4):2535–2542. doi:10.3892/or.2017.5891
- De Miranda NF, Georgiou K, Chen L, et al. Exome sequencing reveals novel mutation targets in diffuse large B-cell lymphomas derived from Chinese patients. Blood. 2014;124(16):2544–2553. doi:10.1182/blood-2013-12-546309
- Hegde VV. Zinc Finger Protein 36L1 (ZFP36L1): Gene Expression, Regulation and Interactions with Immune Receptors in Human Tumour Cells. India: University of Essex; 2020.
- Oliveira C, Faoro H, Alves LR, Goldenberg S. RNA-binding proteins and their role in the regulation of gene expression in Trypanosoma cruzi and Saccharomyces cerevisiae. Genet Mol Biol. 2017;40(12):22–30. doi:10.1590/1678-4685-gmb-2016-0258
- Jespersen DS, Schönherz AA, Due H, Bøgsted M, Sondergaard TE, Dybkær K. Expression of NOTCH3 exon 16 differentiates Diffuse Large B-cell Lymphoma into molecular subtypes and is associated with prognosis. Sci Rep. 2019;9(1):335. doi:10.1038/s41598-018-36680-x
- Nowell CS, Radtke F. Notch as a tumour suppressor. Nat Rev Cancer. 2017;17(3):145–159. doi:10.1038/nrc.2016.145
- Zhang R, Lin P, Yang X, et al. Survival associated alternative splicing events in diffuse large B-cell lymphoma. Am J Transl Res. 2018;10(8):2636.
- Zhang Y, Dong W, Wang J, Cai J, Wang Z. Human omental adipose-derived mesenchymal stem cell-conditioned medium alters the proteomic profile of epithelial ovarian cancer cell lines in vitro. OncoTargets Ther. 2017;10(18):1655. doi:10.2147/OTT.S129502
- Fujimoto A, Okada Y, Boroevich KA, Tsunoda T, Taniguchi H, Nakagawa H. Systematic analysis of mutation distribution in three dimensional protein structures identifies cancer driver genes. Sci Rep. 2016;6(1):1–9. doi:10.1038/srep26483
- Challa-Malladi M, Lieu YK, Califano O, et al. Combined genetic inactivation of β2-Microglobulin and CD58 reveals frequent escape from immune recognition in diffuse large B cell lymphoma. Cancer Cell. 2011;20(6):728–740. doi:10.1016/j.ccr.2011.11.006
- Li Q, Ma L, Wu Z, et al. Zinc finger E‑box binding homeobox 2 functions as an oncogene in human laryngeal squamous cell carcinoma. Mol Med Rep. 2019;19(6):4545–4552. doi:10.3892/mmr.2019.10126
- Schmidt A, Schmitz R, Giefing M, et al. Rare occurrence of biallelic CYLD gene mutations in classical Hodgkin lymphoma. Genes Chromosom Cancer. 2010;49(9):803–809. doi:10.1002/gcc.20789
- Xu X, Wei T, Zhong W, et al. Down-regulation of cylindromatosis protein phosphorylation by BTK inhibitor promotes apoptosis of non-GCB-diffuse large B-cell lymphoma. Cancer Cell Int. 2021;21(1):195. doi:10.1186/s12935-021-01891-2
- Zahn M, Kaluszniak B, Möller P, Marienfeld R. The PTP1B mutant PTP1B∆ 2–4 is a positive regulator of the JAK/STAT signalling pathway in Hodgkin lymphoma. J Carcinog. 2021;42(4):517–527. doi:10.1093/carcin/bgaa144
- Zahn M, Marienfeld R, Melzner I, et al. A novel PTPN1 splice variant upregulates JAK/STAT activity in classical Hodgkin lymphoma cells. Blood. 2017;129(11):1480–1490. doi:10.1182/blood-2016-06-720516
- Walker MP, Stopford CM, Cederlund M, et al. FOXP1 potentiates Wnt/β-catenin signaling in diffuse large B cell lymphoma. Sci.Signal. 2015;8(362):ra12. doi:10.1126/scisignal.2005654
- Nutt SL, Kee BL. The transcriptional regulation of B cell lineage commitment. Immunity. 2007;26(6):715–725. doi:10.1016/j.immuni.2007.05.010
- Møller MB, Kania P, Ino Y, et al. Frequent disruption of the RB1 pathway in diffuse large B cell lymphoma: prognostic significance of E2F-1 and p16INK4A. Leukemia. 2000;14(5):898–904. doi:10.1038/sj.leu.2401761
- Alvarado Y, Giles FJ, Swords RT. The PIM kinases in hematological cancers. Expert Rev Hematol. 2012;5(1):81–96. doi:10.1586/ehm.11.69
- de Miranda NF, Peng R, Georgiou K, et al. DNA repair genes are selectively mutated in diffuse large B cell lymphomas. J Exp Med J EXP MED. 2013;210(9):1729–1742. doi:10.1084/jem.20122842
- Elfrink S, de Winde CM, van den Brand M, et al. High frequency of inactivating tetraspanin C D37 mutations in diffuse large B-cell lymphoma at immune-privileged sites. Blood. 2019;134(12):946–950. doi:10.1182/blood.2019001185
- Fan Z, Pei R, Sha K, Chen L, Wang T, Lu Y. Comprehensive characterization of driver genes in diffuse large B cell lymphoma. Oncol Lett. 2020;20(1):382–390. doi:10.3892/ol.2020.11552
- Krysiak K, Gomez F, White BS, et al. Recurrent somatic mutations affecting B-cell receptor signaling pathway genes in follicular lymphoma. Blood. 2017;129(4):473–483. doi:10.1182/blood-2016-07-729954
- Guo B, Huang Y, Duan Y, Liao C, Cen H. SGK1 mutation status can further stratify patients with germinal center B‐cell‐like diffuse large B‐cell lymphoma into different prognostic subgroups. Cancer Med. 2022;11(5):1281–1291. doi:10.1002/cam4.4550
- Gribben JG. The sequence of events in diffuse large B-cell lymphoma. Blood. 2013;122(7):1097–1098. doi:10.1182/blood-2013-06-506089
- Zhang J, Gu Y, Chen B. Drug-resistance mechanism and new targeted drugs and treatments of relapse and refractory DLBCL. Cancer Manag Res. 2023;245–255. doi:10.2147/CMAR.S400013
- Vaqué JP, Martínez N, Batlle-López A, et al. B-cell lymphoma mutations: improving diagnostics and enabling targeted therapies. Haematologica. 2014;99(2):222–231. doi:10.3324/haematol.2013.096248
- Shen R, Fu D, Dong L, et al. Simplified algorithm for genetic subtyping in diffuse large B-cell lymphoma. Signal Transduct Target Ther. 2023;8(1):145. doi:10.1038/s41392-023-01358-y
- Xu-Monette ZY, Wu L, Visco C, et al. Mutational profile and prognostic significance of TP53 in diffuse large B-cell lymphoma patients treated with R-CHOP: report from an International DLBCL Rituximab-CHOP Consortium Program Study. Blood. 2012;120(19):3986–3996. doi:10.1182/blood-2012-05-433334
- Kusowska A, Kubacz M, Krawczyk M, Slusarczyk A, Winiarska M, Bobrowicz M. Molecular aspects of resistance to immunotherapies advances in understanding and management of diffuse large B-cell lymphoma. Int J Mol Sci. 2022;23(3):1501. doi:10.3390/ijms23031501
- Wilson WH, Young RM, Schmitz R, et al. Targeting B cell receptor signaling with ibrutinib in diffuse large B cell lymphoma. Nat.Med. 2015;21(8):922–926. doi:10.1038/nm.3884
- Yu H, Sotillo E, Harrington C, et al. Repeated loss of target surface antigen after immunotherapy in primary mediastinal large B cell lymphoma. Am J Hematol. 2017;92(1):E11. doi:10.1002/ajh.24594
- Sciarrillo R, Wojtuszkiewicz A, Assaraf YG, et al. The role of alternative splicing in cancer: from oncogenesis to drug resistance. Drug Resist Updat. 2020;53:100728. doi:10.1016/j.drup.2020.100728
- Danilov AV, Magagnoli M, Matasar MJ. Translating the biology of diffuse large B-cell lymphoma into treatment. Oncologist. 2022;27(1):57–66. doi:10.1093/oncolo/oyab004