
Abstract
Dioctadecyldimethylammonium bromide (DODAB) is an antimicrobial lipid that can be dispersed as large closed bilayers (LV) or bilayer disks (BF). Gramicidin (Gr) is an antimicrobial peptide assembling as channels in membranes and increasing their permeability towards cations. In mammalian cells, DODAB and Gr have the drawbacks of Gram-positive resistance and high toxicity, respectively. In this study, DODAB bilayers incorporating Gr showed good antimicrobial activity and low toxicity. Techniques employed were spectroscopy, photon correlation spectroscopy for sizing and evaluation of the surface potential at the shear plane, turbidimetric detection of dissipation of osmotic gradients in LV/Gr, determination of bacterial cell lysis, and counting of colony-forming units. There was quantitative incorporation of Gr and development of functional channels in LV. Gr increased the bilayer charge density in LV but did not affect the BF charge density, with localization of Gr at the BF borders. DODAB/Gr formulations substantially reduce Gr toxicity against eukaryotic cells and advantageously broaden the antimicrobial activity spectrum, effectively killing Escherichia coli and Staphylococcus aureus bacteria with occurrence of cell lysis.
Introduction
Peptides used in therapy should be resistant to the action of proteases.Citation1 Many peptides have been used in a number of biotechnological applications, ranging from neutralizing toxins to antitumor or antimicrobial agents.Citation2 Antimicrobial peptides (AMPs) are considered an important class of molecules requiring development of suitable antibacterial formulations. AMPs have been known for many years, yet very few have been used extensively in the clinic due to their considerable toxicity and high manufacturing costs.Citation2 Further, the development of resistance by pathogenic bacteria requires multiple lines of urgent action, such as the design and synthesis of model peptides with potent antimicrobial activity and low toxicityCitation3 and the invention of novel supramolecular assemblies for the available peptides, allowing combination of different mechanisms of action.Citation4–Citation6 For example, small cationic peptides were recently shown to delocalize peripheral membrane proteins, thereby impacting important cellular processes such as respiration and cell wall biosynthesis.Citation7 Lipid bilayer disksCitation8 have been used as carriers for AMPs, leading to novel, nontoxic, and efficacious formulations which fully protect the peptide against degradation but preserve its activity against bacteria.Citation9 Some polysaccharides have also been evaluated as suitable vehicles for AMPs, with confirmation of the advantages of prolonged retention of peptides at the site of application due to bioadhesion and slowing of proteolytic degradation.Citation10,Citation11
Gramicidin A (Gr) is an antimicrobial peptide with a unique mechanism of hampering the function of bacterial plasma membranes, thereby preventing development of bacterial resistance. Gr is a 15-residue linear peptide extracted from Bacillus brevis, that acquires a β-helix secondary structure with an internal pore and forms transmembrane channels consisting of two Gr molecules associated head-to head.Citation12,Citation13 This channel allows permeation of cations across the membrane, changing the ionic balanceCitation14 and ultimately generating the observed antibiotic activity of Gr.Citation15,Citation16 However, Gr is insoluble in water and highly toxic to mammalian cellsCitation17 over a range of bactericidal concentrations.Citation18 Further, Gr was reported to be ineffective against Gram-negative bacteria.Citation19,Citation20 On the other hand, some cationic lipids assembling as artificial membranes in aqueous solution have been described as potent microbicidal agents against Gram-negative bacteria, but are much less efficient against Gram-positive bacteriaCitation21–Citation23 and fungi.Citation24–Citation26
In the present work, a broad-spectrum combination of Gr and dioctadecyldimethylammonium bromide (DODAB) as closed bilayers (LV) or bilayer disks (BF)Citation27 was characterized with regard to its physical properties, antibacterial activity, and toxicity in a eukaryotic cell model, ie, Saccharomyces cerevisae.Citation28 Such novel combinations have the advantages of low toxicity and broad-spectrum activity via a mechanism that does involve bacterial cell lysis.
Materials and methods
Reagents
DODAB, gramicidin D (a peptide mixture consisting mostly of Gr), ethanol, chloroform, and 2,2,2-trifluoroethanol (TFE) were purchased from Sigma-Aldrich (St Louis, MO, USA).
Preparation of lipid dispersions
LV were obtained from hydration and vortexing of DODAB films with 1 mM sodium chloride aqueous solution (60°C) until the dispersions became homogeneous at a final DODAB concentration equal to 0.002 M.Citation29,Citation30
The ultrasonic dispersion of LV with a macrotip (85 W nominal output for 20 minutes at 70°C) followed by centrifugation (10,000 g for 60 minutes at 15°C) to eliminate titanium particles has been previously described as a procedure for yielding a BF dispersion.Citation29 Disruption of LV in this manner generates the BF. DODAB quantitative analysis was performed by bromide microtitration, as described elsewhere.Citation31
Preparation and characterization of DODAB/Gr assemblies
Aliquots of a stock solution of Gr (6.4 mM) in TFE were added to the DODAB bilayers before incubation (one hour at 70°C) at a DODAB to Gr molar ratio of 10:1. The size distribution, hydrodynamic diameter, polydispersity index, and zeta potential of the dispersions were obtained by photon correlation spectroscopy or dynamic light scattering at 90° using the Brookhaven Zeta Plus-Zeta Potential apparatus (Brookhaven Instruments Corporation, Holtsville, NY, USA). The log-normal fitting function of the apparatus software was used to calculate the mean diameters of the size distributions.Citation32 The zeta potential was given as ζ = μη/ε, where μ, η, and ε are the electrophoretic mobility in 1 mM NaCl solution (25°C), the medium viscosity, and the dielectric constant, respectively.
Evaluation of Gr incorporation in DODAB LV or BF from fluorescence spectra
LV/Gr and BF/Gr (0.2 mM DODAB) were filtered using polycarbonate membranes (with a cutoff of 0.2 μm). The total area under the Gr fluorescence spectra before (Abefore) and after (Aafter) filtering gave %Gr =100× Aafter/Abefore. The fluorescence emission spectra for Gr were determined at 25°C using a F4500 fluorescence spectrofluorometer (Hitachi, Tokyo, Japan) at λexc =280 nm. Excitation and emission slits were fixed at 2.5 nm. The molar extinction coefficient for Gr at 280 nm is 20,700 m−1 cm−1.Citation33
Circular dichroism spectra for Gr in different types of medium
Spectra were acquired at 25°C using a 720 spectropolarimeter (Jasco Inc, Tokyo, Japan) in a 0.1 cm quartz cell with 0.5 nm wavelength increments and a 4-second response in the 200–280 nm range (100 nm per minute). Each spectrum is the average of five scans, with a full-scale sensitivity of 10 m deg. All spectra were corrected for background by subtraction of appropriate blanks in the absence of Gr (DODAB dispersion or Gr solvent). Spectra smoothing kept the overall spectral shape. The ellipticities θ (in deg dmol−1 cm2) were plotted as a function of wavelength.
Evaluation of Gr-induced changes in permeability of DODAB closed bilayers at 2 mM DODAB
Absorbance derived from light scattered by the LV at 400 nm was recorded as a function of time upon addition of hyperosmotic KCl or glucose solutions to LV or LV/Gr dispersions by means of a U2000 spectrophotometer (Hitachi) at room temperature. No hyperosmotic solution was added to the reference cuvette; instead, water was added. Absorbances were normalized to the initial value. The turbidity (400 nm) kinetics was followed after adding 20 mM KCl or 40 mM D-glucose to LV in water.
Microbial cultures
Escherichia coli (ATCC 25922; American Type Culture Collection, Manassas, VA, USA) and Staphylococcus aureus (ATCC 25923) were incubated for 3 hours at 37°C under shaking in tryptic soy broth (Merck, Darmstadt, Germany), spread onto Mueller-Hinton agar plates (Hi-Media Laboratories Pvt, Mumbai, India), and incubated for 24 hours at 37°C. Some colonies from the plates were shaken in tryptic soy broth (160 rpm at 37°C for 2 hours) to reach the exponential phase of growth before pelleting (8,000 rpm for 15 minutes) and washing with a 0.264 M D-glucose solution. Washing was performed twice, and the bacteria were then mixed with the DODAB BF or LV with or without Gr in the same D-glucose solution. Tube 0.5 of the McFarland scale at 625 nm (about 1.5×108 colony-forming units [CFU]/mL) was used as the reference for preparation of the bacteria suspension. Thereafter, bacteria and the DODAB, Gr, or DODAB/Gr dispersions were allowed to interact for counting of CFU, as described in the next section.
S. cerevisae was cultured in yeast extract peptone dextrose medium for both broth and solid agar cultures as previously described.Citation28 Some isolated colonies from a fresh S. cerevisae culture on solid medium were transferred to yeast extract peptone dextrose broth, and the yeast suspension was then incubated at 32°C under shaking (120 rpm for 5 hours) before centrifuging (10,000 rpm for 10 minutes), washing the pellet three times with 0.264 M D-glucose solution, and adjusting the cell concentration to 3–4×105 cells/mL as determined by CFU counting.Citation28
Determination of effects of DODAB or DODAB/Gr on cell viability
Microbes and dispersions were mixed and interacted for one hour before being diluted up to 20,000-fold for plating of 0.1 mL of each in triplicate and incubation (24 hours for 37°C) for CFU counting. Cell survival (%), taken as the mean ± standard deviation, was plotted against DODAB and/or Gr concentration. As a control for cell viability in the absence of DODAB or DODAB/Gr dispersions, a standard bacterial suspension was added to 0.264 M D-glucose solution, diluted, and spread on the agar plate.
S. cerevisae and the dispersions (Gr, BF, LV, BF/Gr or LV/Gr) interacted for one hour. The control sample in the absence of the antimicrobial agent was a mixture of the standard S. cerevisae suspension and 0.264 M D-glucose solution. Aliquots of each diluted mixture (dilution up to 1,000 fold) were plated onto yeast extract peptone dextrose agar. CFU counting was performed after incubation for 48 hours at 32°C. Cell viability (%) was taken as the mean ± standard deviation and plotted against DODAB and/or Gr concentration. The minimum bactericidal concentration (MBC) is the concentration resulting in 99% cell death.
Determination of leakage of phosphorylated compounds from bacteria
Bacteria were grown on plates of Mueller–Hinton agar and incubated for 18–24 hours at 37°C to prepare a cell suspension in 1 mM NaCl solution with turbidity at 625 nm adjusted to 0.400. Aliquots were then transferred to Eppendorf tubes and pelleted (6,000 rpm for 15 minutes) before resuspension in the DODAB, Gr, or DODAB/Gr dispersions in 1 mM NaCl. Bacteria and the assemblies were allowed to interact for one hour, centrifuged (6,000 rpm for 15 minutes), and used to determine the inorganic phosphorus concentration in the supernatant (ie, Pi supernatant), as previously described.Citation34 As a positive (Pi) control, inorganic phosphorus was determined for bacteria in NaCl 1 mM. As a negative control, inorganic phosphorus was determined in the supernatant of bacterial suspensions after centrifugation. Final concentrations in the interaction mixtures ranged from 3.7×108 to 4.8×109 CFU/mL (E. coli) and from 5.2×109 to 5.1×1010 CFU/mL (S. aureus). Bacterial lysis is related to the concentration of phosphorylated compounds in the supernatant as percent leakage =100× Pisupernatant/(Picontrol).Citation21
Evaluation of cell morphology by scanning electron microscopy
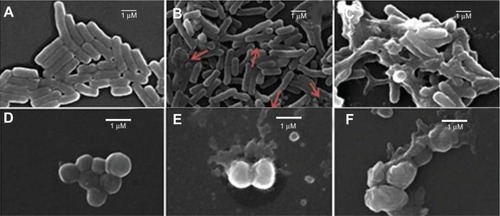
Bacteria interacted with DODAB/Gr dispersions above the MBC (37°C for 2 hours) before being pelleted (12,000 rpm for 10 minutes), progressively dehydrated by ethanol as previously described,Citation35 and then placed on a silicon wafer for evaporation of ethanol at room temperature, gold sputtering, and imaging by a JSM-6460LV scanning electron microscope (JEOL Ltd, Tokyo, Japan) at 20 kV.
Results
Sizing, zeta potential, polydispersity index, and gramicidin conformation and functionality in DODAB/Gr assemblies
shows the steps involved in preparing the DODAB/Gr dispersions. Preparation of BF/Gr and LV/Gr with insertion of Gr in LV and adsorption of Gr at the borders of BF are schematically illustrated. The open nature of the BF in the absence of Gr has been often reported in the literature, both by electron microscopyCitation36 and by cryotransmission electron microscopy.Citation37 The sonication procedure not only disperses the cationic lipid to promote its self-assembly as bilayer vesicles but also disrupts the closed vesicles, producing the bilayer fragments or disks.Citation8
Figure 1 Preparation of BF/Gr and LV/Gr dispersions and representation of possible conformations for Gr in the DODAB bilayer: dimeric channel conformation in LV and intertwined Gr molecules at the borders of BF.
Abbreviations: DODAB, dioctadecyldimethylammonium bromide; LV, closed bilayers; BF, bilayer disks; Gr, Gramicidin; TFE, 2,2,2-trifluoroethanol.
Incorporation of the peptide in BF caused an increase in hydrodynamic diameter, whilst for LV, incorporation of Gr decreased the hydrodynamic diameter (). Gr possibly induced a slight aggregation of BF due to a small degree of intertwining between adsorbed Gr molecules at different BF. On the other hand, insertion of Gr in the LV improved the colloidal stability of the LV, possibly due to the presence of Gr tryptophan residues at the bilayer/water interface. Incorporation of Gr in DODAB BF slightly affected the zeta potential, which changed from 55±3 to 43±4 mV (). For DODAB LV, incorporation of Gr substantially increased the zeta potential from 46±2 mV to 61±3 mV. When Gr becomes incorporated in the LV bilayer, the charge density at the level of the polar heads increases for the LV/Gr dispersion but this does not occur for the BF upon incorporation of Gr: in the latter case, the charge density and zeta potential remain practically unchanged. For BF/Gr, Gr induced some aggregation of BF, as suggested by the increase in hydrodynamic diameter from 61±0 nm to 104±1 nm. This would be consistent with interactions between Gr molecules at the border of different BF causing a certain extent of aggregation. The polydispersity of the assemblies was reduced for both BF/Gr and LV/Gr as compared with the nonloaded bilayers, suggesting some degree of stabilization of the DODAB dispersions induced by the peptide. For LV/Gr, the colloidal stabilization induced by the peptide is confirmed further by a reduction in hydrodynamic diameter from 379±10 nm to 247±2 nm (), indicating reduced aggregation for LV assemblies with Gr.
Table 1 Physical characterization of DODAB BF or LV, DODAB BF/Gr, or DODAB LV/Gr dispersions prepared in a 1 mM solution of NaCl
The intrinsic fluorescence of Gr due to tryptophans was used to determine incorporation of the peptide in the DODAB BF or LV dispersions. The filtration procedure for the BF/Gr dispersion yielded in the filtrate a mean molar percentage of 38%±11% for DODAB and 28%±1% for Gr. Within the limits of the experimental error, these two values are practically equal. The mean percentage of DODAB and Gr in the filtrate of the LV/Gr dispersion was 0%, indicating retention of LV or LV/Gr by the filter (). The 200 nm cutoff filter retained the majority of the LV/Gr but permitted flow of smaller BF/Gr assemblies. The results shown in also demonstrate quantitative incorporation of Gr in the DODAB bilayers. When DODAB from BF is present in the filtrate, so is Gr. When DODAB from LV is retained by the filter, so is Gr, and in the same proportion ().
Table 2 DODAB or Gr concentrations (%) in BF/Gr or LV/Gr dispersions before and after filtering
The fluorescent tryptophans in the Gr molecular structure may act as intrinsic labels for the peptide. shows the fluorescence spectra for Gr in different types of medium, such as TFE, ethanol, and DODAB dispersions. Although the four spectra share similar shapes and peaks, the intensity of fluorescence for Gr in BF is more similar to that for Gr in ethanol than for Gr in TFE (). For DODAB LV/Gr, tryptophan residues sense a microenvironment similar to the one in TFE. Thus, Gr perceives a more polar medium in BF than in LV.
Figure 2 Gr intrinsic fluorescence at λexc =280 nm and slit widths of 2.5 nm in different types of medium (A). Gr circular dichroism spectra at 25°C in different types of medium (B). In both subfigures, the Gr concentration is 0.02 mM and the DODAB concentration is 0.2 mM.
Abbreviations: DODAB, dioctadecyldimethylammonium bromide; LV, closed bilayers; BF, bilayer disks; Gr, Gramicidin; TFE, 2,2,2-trifluoroethanol.
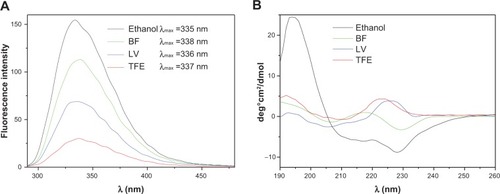
shows the circular dichroism (CD) spectra of Gr in ethanol, TFE, BF, and LV. The Gr spectrum in LV has a general shape that resembles the spectrum of Gr in TFE. In BF, Gr displays a CD spectrum with the negative peak at about 230 nm, which is similar to that for Gr in ethanol (). These data support the conclusion that the Gr β-helix in BF senses a microenvironment more like that in ethanol than that in TFE, whereas in LV, the Gr β-helix perceives a medium more like that in TFE than that in ethanol. In the literature, the negative peak around 230 nm and the positive peak around 197 nm in ethanol typically show intertwining of Gr molecules.Citation38 The CD spectrum for Gr in BF is more similar in shape to that of Gr in ethanol than that of Gr in TFE. However, in the case of Gr in BF, one should not expect to see the same extensive intertwining of Gr molecules in ethanol since the intertwining event may take place at a lower frequency at the BF borders than it does in ethanol. For example, the intertwining between Gr molecules adsorbed on different BF may be the reason for aggregation of BF with the observed increase in size (). However, this increase in size is not so large as to be consistent with extensive aggregation of BF.
The effect of adding hypertonic solutions to LV is shown in . Turbidity at 400 nm increased with time when the water from the inner vesicle compartment flowed in accordance with the solute gradient, resulting in shrinkage of LV (). It has been previously reported for vesicles of similar size that turbidimetry allows monitoring of vesicle shrinkage or swelling, which corresponds respectively to an increase or decrease in turbidity kinetics at 400 nm.Citation29,Citation39 In fact, the osmotic behavior of LV depended on the osmolarity of the outer medium, but in the presence of Gr, this behavior changed significantly, ie, instead of shrinking, the LV swelled on addition of the hypertonic external medium ().
Figure 3 Effect of KCl or glucose concentration gradients across the LV on the normalized turbidity of the dispersions without (A–C) or with (D–F) Gr inserted in the DODAB LV. The Gr concentration is 0.1 mM and the DODAB concentration is 1.0 mM. The kinetics were obtained at 25°C.
Abbreviations: DODAB, dioctadecyldimethylammonium bromide; LV, closed bilayers; Gr, Gramicidin.
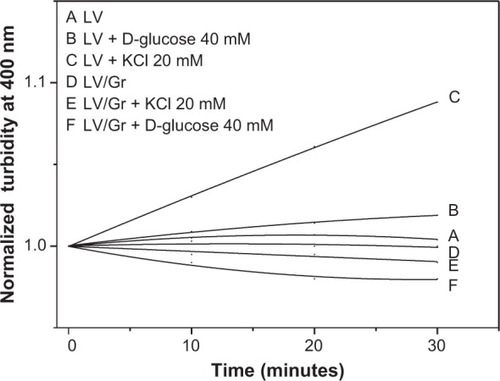
The Gr channel conformation in LV modifies the usually low permeation of cations through the DODAB bilayer. Permeation of D-glucose through the DODAB LV/Gr bilayer also increases substantially in comparison with permeation through the DODAB LV bilayer. The entry of D-glucose or KCl solutes via the Gr channel from the outer to the inner vesicle compartment is accompanied by entry of water, which causes the vesicle to swell ().
Antimicrobial activity, mechanism of action, and differential cytotoxicity of DODAB/Gr assemblies
The antimicrobial activity of DODAB, Gr, and DODAB/Gr assemblies was evaluated against E. coli, S. aureus, and S. cerevisae in order to gain further insights into the differential cytotoxicity of the combined formulations (). There was poor Gr activity against E. coli and S. cerevisae as shown from the survival of microbes (%) over a range of Gr concentrations, in contrast with the action of Gr on S. aureus, consistent with previous reports in the literature.Citation18–Citation20
Figure 4 Cell viability (%) of Escherichia coli, Staphylococcus aureus, and Saccharomyces cerevisae as a function of Gr or DODAB. DODAB was dispersed as BF or LV, that were further added of Gr for a final DODAB to Gr molar ratio of about 10:1. The interaction time between the microbial cells and the dispersions was one hour, at final cell concentrations of 1.0×108 CFU/mL (E. coli or S. aureus) or 1.0×105 CFU/mL (S. cerevisae).
Abbreviations: CFU, colony-forming units; DODAB, dioctadecyldimethylammonium bromide; LV, closed bilayers; BF, bilayer disks; Gr, Gramicidin.
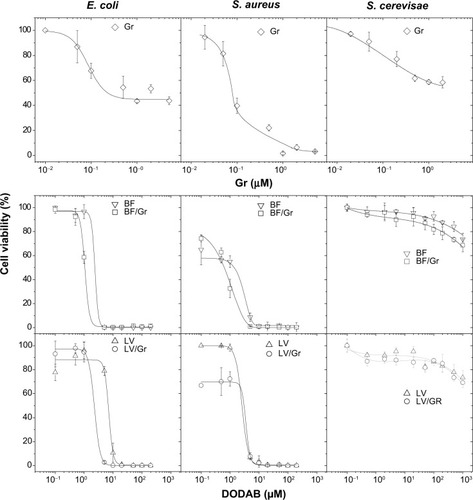
The activity of DODAB as shown in was also consistent with previous data in the literature reporting its high activity against E. coliCitation21,Citation22 and poor activity against the yeast.Citation24–Citation26 The novelty of these combinations was the broadening of antimicrobial activity to encompass E. coli and S. aureus as representatives of Gram-negative and Gram-positive bacteria. Of note, the microbicidal activity occurred over a range of low Gr and DODAB concentrations which are not toxic to S. cerevisae ().
The MBCs taken from the viability curves for all dispersions are shown in . The MBCs are lower for DODAB and Gr in combination in comparison with those obtained for the separate DODAB and Gr dispersions (). The lowest MBCs were obtained for DODAB BF/Gr, which revealed superior performance for delivering Gr to bacteria in comparison with DODAB LV/Gr. The high DODAB concentrations needed to kill S. aureus reconfirm the emergence of resistance of Gram-positive bacteria to cationic antimicrobial agents.Citation40 In this respect, delivering Gr becomes very important since Gr does not bear cationic moieties and acts by a different mechanism. The excellent activity of DODAB against Gram-negative bacteria complements the poor activity of the peptide against these bacteria. The excellent activity of Gr against Gram-positive bacteria complements the poor activity of DODAB against these bacteria. The combined formulations barely affected the S. cerevisae cells, suggesting limited toxicity if used for further therapeutic application.
Table 3 Minimum bactericidal concentrations (in μM) and cell lysis at the minimum bactericidal concentration (% L) for DODAB, Gr, and DODAB/Gr dispersions
In order to shed some light on the mechanism of antimicrobial action for the assemblies, leakage of phosphorylated compounds from bacteria was determined over a range of DODAB concentrations (), and no cell rupture was seen over a range of low DODAB concentrations. This remained so up to the MBC, as also seen in . Above the MBC, the leakage increased, suggesting some important lytic events taking place in the cells. The bacterial morphology examined by scanning electron microscopy before and after interaction with DODAB/Gr at DODAB and Gr concentrations above the MBCs also suggested some cellular distortions departing from the morphology of untreated cells ().
Figure 5 Leakage of phosphorylated compounds (%) from bacteria for different dispersions at 3.7×108 to 4.8×109 CFU/mL (Escherichia coli) or 5.2×109 to 5.1×1010 CFU/mL (Staphylococcus aureus). The dispersions were tested over a range of DODAB or DODAB/Gr concentrations after one hour of interaction with both types of bacteria. As a control, over a range of Gr concentrations (10−2 to 10−4 mM), no leakage was detected from bacteria after the same interaction time.
Abbreviations: CFU, colony-forming units; DODAB, dioctadecyldimethylammonium bromide; LV, closed bilayers; BF, bilayer disks; Gr, Gramicidin.
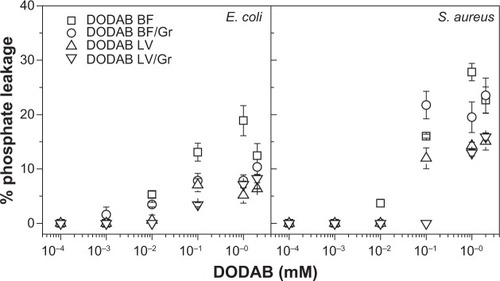
Figure 6 Micrographs of Escherichia coli (A–C) or Staphylococcus aureus (D–F) cells untreated (A, D) or treated with DODAB BF/Gr (B, E) or DODAB LV/Gr dispersions (C, F), obtained by scanning electron microscopy. Cells appear enlarged by 10,000× (E. coli) or 20,000× (S. aureus). Against E. coli cells, in (B), DODAB =0.005 mM (DODAB BF/Gr) and, in (C), DODAB =0.01 mM (DODAB LV/Gr). Against S. aureus, in (E), DODAB =0.01 mM (DODAB BF/Gr) and, in (F), DODAB =0.05 mM (DODAB LV/Gr).
Abbreviations: DODAB, dioctadecyldimethylammonium bromide; LV, closed bilayers; BF, bilayer disks; Gr, Gramicidin.
Discussion
The physical behavior of Gr in the DODAB dispersions was very similar to that previously reported for Gr in composite bilayers of phospholipid and DODAB at a molar proportion of 1:1.Citation39 Fluorescence and CD data reported the Gr microenvironment. For DODAB BF, the Gr helical moiety perceives the hydrophobic borders of the bilayer disks and the aqueous medium surrounding the disks. The intertwined Gr conformation in BF was depicted in the CD spectra. The Gr beta-helix in LV or TFE was demonstrated by the CD positive peak around 225 nm and associated with the functional channel present in LV (). Tryptophans at the membrane/water surface help to ensure appropriate conformation and activity in the Gr channel.Citation12,Citation38,Citation39 Gr fluorescence is similar in LV and TFE, showing localization of the tryptophans in a hydrophobic medium (). In contrast, Gr fluorescence is also similar in BF and ethanol, showing localization of the tryptophans in a more polar medium (). Gr possibly attaches to the edges of the disks at low concentrations, but with increasing Gr concentration, other Gr molecules from the outer solution can intertwine with those already attached at the disk border.
For LV, shrinking or swelling causes changes in the turbidity of the dispersions.Citation29 In accordance with the Joebst equation, the turbidity of the spheres in the dispersion varies with 1/R2, where R is the mean particle radius and is useful for inferring changes in vesicle size taking place upon establishment of osmotic gradients through the LV bilayers. DODAB LV shrinkage and swelling would correspond to the kinetics of increasing and decreasing turbidity, respectively (). The Gr inserted in LV form channels, that change the LV permeation to solutes, such as D-glucose and KCl consistent with previous reports of similar vesicular dispersions in the presence of Gr.Citation39
Hydrophobic drugs solubilize at the hydrophobic borders of BF, in contrast with the absence of drug solubilization in LV.Citation4 Given the hydrophobic nature of the Gr beta-helix, the behavior of Gr in the DODAB bilayers is similar to that of hydrophobic drugs in good organic solvents and in DODAB bilayers.Citation4 However, delivery of Gr to the bacterial cells seems to be favored by its combination with BF, possibly due to peripheral localization of Gr in the BF ().
The antimicrobial activity of the DODAB cationic lipid observed at a very low DODAB concentration is related to the quaternary nitrogen in its molecular structure. The differential cytotoxicity of DODAB has been systematically studied, eg, at 0.5 mM, DODAB kills about 50% of fibroblasts cultured as a subconfluent monolayer.Citation1 The antimicrobial activity of Gr over the micromolar range of concentrations against Gram-positive bacteria has also been well established,Citation18 as well as its differential cytotoxicity.Citation17,Citation18 The minimum inhibitory concentration of Gr against S. aureus is 2.5 μM.Citation18 The survival of mammalian HeLa cells at this concentration is 15%, indicating that the peptide is highly toxic.Citation18 In addition, there is about 50% survival of cultured renal cells at 2.5 μg/mL Gr,Citation17 reconfirming its high toxicity at concentrations that kill the bacteria. Here the MBC for Gr against S. aureus is about 4 μM (, ) and is consistent with the previously reported minimum inhibitory concentration of 2.5 μM.Citation10 Gr is indeed very toxic in mammalian cells,Citation17,Citation18 and also against eukaryotic cells, as exemplified by its activity against S. cerevisae (). However, in formulations containing DODAB, this toxicity is substantially reduced, as indicated by comparison of the viability curves for S. cerevisae using Gr alone and using DODAB/Gr (). Cell viability remained very high for both DODAB BF/Gr and DODAB LV/Gr, clearly showing improvement in delivery of Gr by the DODAB/Gr combinations. Further, DODAB is not only a vehicle but also displays good activity against E. coli, thus broadening the spectrum of antimicrobial activity for Gr.
The mechanism of action of these novel formulations involves bacterial cell lysis since leakage of phosphorylated compounds took place from the MBC values (, ). Some lysis was detected at doses above the MBC, and visualization of cells by scanning electron microscopy indeed showed distortions in cell morphology (). The detergent-like mechanism often ascribed to the interaction between antimicrobial peptides and surfactants seems to be important for determining antimicrobial activity. For the DODAB/Gr combinations, the mechanism involved affects membrane function and selectivity in the transport of ions and nutrients and ion distribution in the cell. Death of bacterial cells possibly takes place with some substantial membrane rupture and leak of intracellular bacterial compounds. Eventually, damage to certain transmembrane proteins may also be involved in the death mechanism.
Acknowledgments
This work was supported by grants 502231/2010-6 and 470105/2010-0 from The Conselho Nacional de Desenvolvimento Científico e Tecnológico and grants 2011/00046-5 and 2012/24534-1 from The Fundacçaão de Amparo à Pesquisa do Estado de Saão Paulo. The technical assistance of Rodrigo Tadeu Ribeiro is also gratefully acknowledged.
Disclosure
The authors report no conflicts of interest in this work.
References
- BracciLFalcianiCLelliBSynthetic peptides in the form of dendrimers become resistant to protease activityJ Biol Chem200327847465904659512972419
- RosciaGFalcianiCBracciLPiniAThe development of antimicrobial peptides as new antibacterial drugsCurr Protein Pept Sci201314864164924384032
- KimS-JKimJ-SLeeY-SStructural characterization of de novo designed L5K5W model peptide isomers with potent antimicrobial and varied hemolytic activitiesMolecules201318185987623344198
- Carmona-RibeiroAMBiomimetic nanoparticles: preparation, characterization and biomedical applicationsInt J Nanomedicine2010524925920463941
- Carmona-RibeiroAMBarbassaLMeloLDAntimicrobial biomimeticsCavrakMBiomimetic Based ApplicationsRijeka, CroatiaInTech2011
- Carmona-RibeiroAMde Melo CarrascoLDCationic antimicrobial polymers and their assembliesInt J Mol Sci20131459906994623665898
- WenzelMChiriacAIOttoASmall cationic antimicrobial peptides delocalize peripheral membrane proteinsProc Natl Acad Sci U S A201411114E1409E141824706874
- Carmona-RibeiroAMLipid bilayer fragments and disks in drug deliveryCurr Med Chem200613121359137016719782
- ZetterbergMMReijmarKPraentingMEngstroemAAnderssonDIEdwardsKPEG-stabilized lipid disks as carriers for amphiphilic antimicrobial peptidesJ Control Release2011156332332821903146
- RuissenALvan der ReijdenWAvan’t HofWVeermanECNieuw AmerongenAVEvaluation of the use of xanthan as vehicle for cationic antifungal peptidesJ Control Release1999601495610370170
- GawandePVLeungKPMadhyasthaSAntibiofilm and antimicrobial efficacy of DispersinB®-KSL-W peptide-based wound gel against chronic wound infection associated bacteriaCurr Microbiol201468563564124445333
- KelkarDAChattopadhyayAThe gramicidin ion channel: a model membrane proteinBiochim Biophys Acta2007176892011202517572379
- UrbanBWHladkySBHaydonDAIon movements in gramicidin pores. An example of single-file transportBiochim Biophys Acta198060223313546159005
- ClementNRGouldJMKinetics for development of gramicidin-induced ion permeability in unilamellar phospholipid vesiclesBiochemistry1981206154415486164386
- HaroldFMBaardaJRGramicidin, valinomycin, and cation permeability of Streptococcus faecalisJ Bacteriol196794153604961416
- HamadaTMatsunagaSFujiwaraMSolution structure of polytheonamide B, a highly cytotoxic nonribosomal polypeptide from marine spongeJ Am Chem Soc201013237129411294520795624
- SorochkinaAIPlotnikovEYRokitskayaTIN-terminally glutamate-substituted analogue of gramicidin A as a protonophore and selective mitochondrial uncouplerPLoS One201277e4191922911866
- WangFQinLPaceCJWongPMalonisRGaoJSolubilized gramicidin A as potential systemic antibioticsChem Bio Chem20121315155
- MollGNvan den EertweghVTournoisHRoelofsenBOp den KampJAvan DeenenLLGrowth inhibition of Plasmodium falciparum in in vitro cultures by selective action of tryptophan-N-formylated gramicidin incorporated in lipid vesiclesBiochim Biophys Acta1991106222062101706202
- RammelkampCHWeinsteinLToxic effects of tyrothricin, gramicidin and tyrocidineJ Infect Dis1942712166173
- MartinsLMSMamizukaEMCarmona-RibeiroAMCationic vesicles as bactericidesLangmuir1997132155835587
- CampanhaãMTMamizukaEMCarmona-RibeiroAMInteractions between cationic liposomes and bacteria: the physical-chemistry of the bactericidal actionJ Lipid Res19994081495150010428986
- MeloLDMamizukaEMCarmona-RibeiroAMAntimicrobial particles from cationic lipid and polyelectrolytesLangmuir20102614123001230620578678
- CampanhaãMTMamizukaEMCarmona-RibeiroAMInteractions between cationic vesicles and Candida albicansJ Phys Chem B20011053482308236
- MeloLDCarmona-RibeiroAMFungicidal nanoparticles of low toxicity from cationic lipid and polyelectrolytesNSTI Nanotech20123350353
- Carmona-RibeiroAMCarrascoLDMFungicidal assemblies and their mode of actionOA Biotechnology20132325
- Carmona-RibeiroAMSynthetic amphiphile vesiclesChem Soc Rev199221209214
- BuschiniAPoliPRossiCSaccharomyces cerevisiae as an eukaryotic cell model to asses cytotoxicity and genotoxicity of three anticancer anthraquinonesMutagenesis2003181252612473732
- Carmona-RibeiroAMChaimovichHPreparation and characterization of large dioctadecyldimethylammonium chloride liposomes and comparison with small sonicated vesiclesBiochim Biophys Acta198373311721796882751
- SobralCNSotoMACarmona-RibeiroAMCharacterization of DODAB/DPPC vesiclesChem Phys Lipids20081521384418258186
- SchalesOSchalesSA simple and accurate method for the determination of chloride in biological fluidsJ Biol Chem1941140879884
- GrabowskiEMorrisonIMeasurements of suspended particles by quasi-elastic light scatteringDahnekeBMeasurements of Suspended Particles by Quasi-Elastic Light ScatteringNew York, NY, USAWiley Interscience1983
- KillianJAPrasadKUHainsDUrryDWThe membrane as an environment of minimal interconversion. A circular dichroism study on the solvent dependence of the conformational behavior of gramicidin in diacylphosphatidylcholine model membranesBiochemistry19882713484848552458757
- Carmona-RibeiroAMPreparation and characterization of biomimetic nanoparticles for drug deliveryMethods Mol Biol201290628329422791441
- HoqueJAkkapeddiPYarlagaddaVUppuDSKumarPHaldarJCleavable cationic antibacterial amphiphiles: synthesis, mechanism of action, and cytotoxicitiesLangmuir20122833122251223422838496
- Carmona-RibeiroAMCastumaCESessoASchreierSBilayer structure and stability in dihexadecyl phosphate dispersionsJ Phys Chem19919553615366
- AnderssonMHammarstromLEdwardsKEffect of bilayer phase transitions of vesicle structure, and its influence on the kinetics of viologen reductionJ Phys Chem199599391453114538
- Abdul-ManantNHintonJFConformation states of gramicidin A along the pathway to the formation of channels in model membranes determined by 2D NMR and circular dichroism spectroscopyBiochemistry19943322677367837515684
- CarvalhoCAOlivares-OrtegaCSoto-ArriazaMACarmona-RibeiroAMInteraction of gramicidin with DPPC/DODAB bilayer fragmentsBiochim Biophys Acta20121818123064307122960286
- OttoMStaphylococcus epidermidis – the “accidental” pathogenNat Rev Microbiol20097855556719609257