
Abstract
Background
Necrotizing enterocolitis (NEC) is a devastating gastrointestinal disease that primarily affects premature infants. It is characterized by inflammation and leukocyte infiltration that can progress to intestinal necrosis, perforation, systemic inflammatory response, and death. In this study, we examined the effect of FLLL32, a curcumin analog, on an NEC-like neonatal intestinal injury model.
Methods
NEC was induced in CD-1 mice pups using the Paneth cell ablation and Klebsiella infection model. Pups were divided into sham, NEC, and NEC + FLLL32 groups. At the end of the experiment, pups were euthanized; whole blood and small intestines were harvested. Intestinal inflammatory cytokine profile, in vivo intestinal permeability using serum fluorescein isothiocyanate-dextran, and histological injury scores were compared between the groups.
Results and conclusion
FLLL32-treated pups had lower intestinal injury, improved intestinal permeability, and lower proinflammatory cytokine profiles compared to those in untreated pups with NEC. These results suggest that FLLL32 plays a protective role in NEC.
Introduction
Necrotizing enterocolitis (NEC) is a devastating gastrointestinal disease characterized by inflammation and injury to the small intestine.Citation1,Citation2 Despite advances in care, NEC remains the leading cause of morbidity and mortality in infants born weighing less than 1500 g.Citation1,Citation3,Citation4 Although the etiology of NEC is unclear, evidence suggests that inflammation plays a central part in the pathogenesis.Citation5,Citation6 Specifically, activation of Toll-like receptor 4 (TLR4) is essential for the development of NEC.Citation7–Citation9 Thus, strategies to inhibit TLR activation or inflammatory cytokines have been proposed to help ameliorate injury in models of NEC.Citation10,Citation11
Curcumin is a natural compound found in Curcuma longa.Citation12 Curcumin interacts with numerous targets that are involved in inflammation.Citation12–Citation15 It downregulates inflammatory cytokines, such as tumor necrosis factor-alpha (TNF-α) and interleukin (IL)-1, IL-6, and IL-8, by inhibiting TLR4/nuclear factor kappa B (NF-κB) activation.Citation14,Citation15 In intestinal inflammation models, curcumin attenuates inflammation and injury through inhibition of COX-2 expressionCitation16 and the Janus kinase (JAK)/signal transducers and activators of transcription (STAT) signaling pathways.Citation17–Citation20
The safety and tolerability of curcumin at high doses are well established. However, under physiologic conditions, curcumin is unstable and has poor bioavailability. FLLL32 is a diketone analog of curcumin that exhibits improved potency and biochemical properties, specifically as an inhibitor of the inflammatory signaling molecule Jak2/STAT3.Citation21,Citation22 The use of FLLL32 in intestinal inflammation has not been explored. Thus, the purpose of this study was to determine the effect of FLLL32 in a neonatal NEC-like intestinal inflammation model.
Methods
Effect of FLLL32 and curcumin on IL-6-induced reduction of transepithelial electrical resistance (TEER) in T84 monolayer
T84 intestinal epithelial cells (CCL-248) purchased from the American Type Cell Culture (ATCC, Manassas, VA, USA) were grown under standard conditions as described previously.Citation23,Citation24 Cells were seeded in 24-well Transwells (Cat#: 353495, 6.5 mm diameter, 0.4 mm pore size, PET track-etched membrane) at a density of 6 × 104 cells/800 mL/well (BD Biosciences, San Jose, CA, USA). Cells were grown for 7 days on the Transwells prior to the experiments, until TEER indicated established tight junction formation (TEER>500 Ω cm2). One day prior to the experiment, cells were serum starved overnight. Cells were pretreated with FLLL32 (50 µM), curcumin (50 µM), or carrier (dimethyl sulfoxide) for 1 hour in serum-free medium. Recombinant human IL-6 (10 ng/mL) was applied to the cell monolayers (Miltenyioe Biotech, San Diego, CA, USA). After 60 minutes, the cells were supplemented with serum, and TEER, as an indicator of tight junction permeability, was measured at 24, 48, and 72 hours.Citation24 TEER was measured using Millcell-ERS (Millipore, Bedford, MA, USA) and calculated as ohms × cm2 (Ω cm2) by multiplying it by the surface area of the monolayer. Changes in TEER during experimental conditions were calculated as the percentage of corresponding basal values. Triplet cell monolayers were used for each group in each experiment.
Determination of no-observable-adverse-effect-level (NOAEL)
To determine the safety and tolerability of FLLL32 (Calbiochem®) in neonatal mice, P8-P14 CD-1 mouse pups were orally gavaged 25 and 50 mg/kg of FLLL32 at a volume of 10 µL/g once daily at the same time every day for 7 days. The pups were weighed daily and observed for any adverse effects, such as lethargy, abnormal behavior, weight loss, and death.
Experimental model of NEC
All animal procedures were approved by the Institutional Animal Care and Use Committee at the University of Oklahoma Health Sciences Center and were performed in strict accordance with the recommendations in the Guide for the Care and Use of Laboratory Animals. Briefly, CD-1 (Charles River Laboratory) pups were divided into three groups: 1) Sham (n=6), 2) NEC (n=18), and 3) NEC + FLLL32 (n=12). The NEC + FLLL32 group received 25 mg/kg of FLLL32 by oral gavage from P9 to P14. This was based on prior studies and the NOAEL effects listed previously.Citation22 NEC was induced at P15 using the Paneth-cell ablation and Klebsiella infection model, also known as the dithizone/Klebsiella model (DK model).Citation25 Pups received an intraperitoneal (ip) injection of 33 mg/kg of dithizone (Sigma-Aldrich, St. Louis, MO, USA) or an equivalent volume of vehicle alone at P15. Six hours after the injection, mice pups received an enteral gavage of 108 CFU/g body weight Klebsiella pneumoniae (ATCC#10031, Manassas, VA, USA). Mice pups were then continuously monitored for 10 hours after which they were euthanized, and blood and tissues were collected.
Histological evaluation of NEC
Tissues were fixed with 10% formalin buffer and stained with hematoxylin and eosin for microscopic examination. Mucosal injury was evaluated by two blinded pathologists and graded on a 4-point scale: grade 0, no injury (normal); grade 1, separation of the villous core; grade 2, villous core separation and epithelial sloughing; and grade 3, denudation of epithelium with loss of villi, full-thickness necrosis. The analysis was performed on three to five sections. Animals with histological scores ≥2 were defined as having NEC.Citation26
In vivo intestinal barrier function assay
To investigate the effect of FLLL32 on intestinal permeability barrier function in the NEC model, an in vivo permeability assay was performed using fluorescein isothiocyanate (FITC)-dextran.Citation27,Citation28 Surviving mice pups were gavaged 4 kDa FITC-dextran (44 mg/100 g body weight). After 4 hours, blood was collected and centrifuged at 4°C, 3000 g for 10 min. Plasma was diluted in an equal volume of phosphate-buffered saline (PBS) and analyzed for DX-4000–FITC concentration with a fluorescence spectrophotometer (Tecan, Maennedorf, Switzerland) at an excitation wavelength of 480 nm and emission wavelength of 520 nm.
Mouse inflammatory cytokine quantification
Small intestinal samples were homogenized in lysis buffer. Intestinal cytokines were analyzed using custom designed Procarta Plex Mouse Cytokine & Chemokine Panel 1A (eBioscience, San Diego, CA, USA) as per manufacturer instructions. Final cytokine levels were normalized to total protein concentration (milligram/milliliter) and reported as pictogram/microgram protein.
Statistical analysis
Data are presented as mean ± standard error of mean (SEM) and analyzed using one-way analysis of variance (ANOVA) followed by Dunnett’s multiple comparisons test. Data were considered significant when p<0.05. All the analyses were performed using GraphPad Prism version 6.00 for Windows (GraphPad Software, La Jolla, CA, USA).
Results
FLLL32 protects T84 monolayers against inflammation-induced paracellular permeability
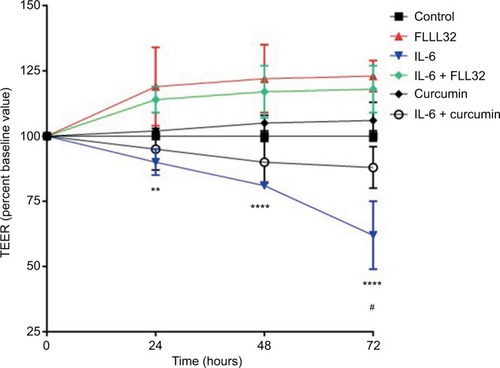
To evaluate the effect of FLLL32 on intestinal permeability in vitro, we measured the TEER in T84 cell monolayers. Similar to previous studies,Citation24 IL-6 decreased the TEER in a time-dependent manner, indicating increased tight junction permeability to ionic solutes. As seen in , the addition of 10 mg/mL IL-6 to the culture medium for 72 hours induced more than 25% decrease in the TEER reading compared to baseline in a time-dependent manner. Pretreatment with FLLL32 ameliorated IL-6 effect on TEER at 24, 48, and 72 hours (). In contrast to curcumin pretreatment, at the same molar concentration, FLLL32 had no effect on IL-6-induced reduction in TEER at 24 and 48 hours. At 72 hours, a modest alleviation on TEER is seen when compared with FLLL32 pretreatment. These results could suggest that FLLL32 is more superior to curcumin in preserving barrier function in vitro compared to curcumin.
Figure 1 Effect of FLLL32 and curcumin on IL-6-induced reduction of TEER in T84 monolayer.
Notes: TEER value of T84 monolayers incubated with cell culture medium for 0–72 hours in the presence of IL-6 (10 ng/mL) with FLLL32 (50 µM), curcumin (50 µM), or carrier (dimethyl sulfoxide) for 1 hour in serum-free medium. **p=0.001 (IL-6 vs. IL-6 + FLLL32 at 24 hours), ****p<0.0001 (IL-6 vs. IL-6 + FLLL32 at 48 and 72 hours), and #p=0.003 (IL-6 vs. IL-6 + curcumin).
Abbreviations: IL, interleukin; TEER, transepithelial electrical resistance.
Determination of NOAEL
Although the use of FLLL32 has been used extensively in different murine models of malignancy, its use in neonatal mice pups has not been reported. Thus, we sought to determine its safety and tolerability in CD-1 mice pups for 7 days starting P8 till P 14. We administered FLLL32 orally at two different doses, 25 mg/kg and orally 50 mg/kg at a volume of 10 µL/g once daily. The pups were weighed daily and observed for any clinical signs of distress such as lethargy, abnormal behavior, weight loss, or death. Pups receiving 25 mg/kg of FLLL32 had no clinical signs of adverse events and had no difference in weight gain compared to controls. In contrast, pups receiving 50 mg/kg daily had lower weight gain on days 6 and 7 compared to both control and the 25 mg/kg group ().
Figure 2 Determination of no-observable-adverse-effect-level (NOAEL).
Notes: P8-P14 CD-1 mouse pups were orally gavaged 25 and 50 mg/kg of FLLL32 once daily for 7 days. Pups were weighed daily and observed for side effects. *p<0.05 (FLLL32 [50 mg/kg] vs. control at 6 days), **p<0.01 (FLLL32 [50 mg/kg] vs. control at 7 days). #p<0.001 (FLLL32 [50 mg/kg] vs. FLLL32 [25 mg/kg] at 6 days). ##p<0.0001 (FLLL32 [50 mg/kg] vs. FLLL32 [25 mg/kg]).
![Figure 2 Determination of no-observable-adverse-effect-level (NOAEL).Notes: P8-P14 CD-1 mouse pups were orally gavaged 25 and 50 mg/kg of FLLL32 once daily for 7 days. Pups were weighed daily and observed for side effects. *p<0.05 (FLLL32 [50 mg/kg] vs. control at 6 days), **p<0.01 (FLLL32 [50 mg/kg] vs. control at 7 days). #p<0.001 (FLLL32 [50 mg/kg] vs. FLLL32 [25 mg/kg] at 6 days). ##p<0.0001 (FLLL32 [50 mg/kg] vs. FLLL32 [25 mg/kg]).](/cms/asset/31096784-0111-4feb-a5d6-b67ae564d366/djir_a_131051_f0002_b.jpg)
FLLL32 reduces incidence and severity of intestinal injury in DK NEC model
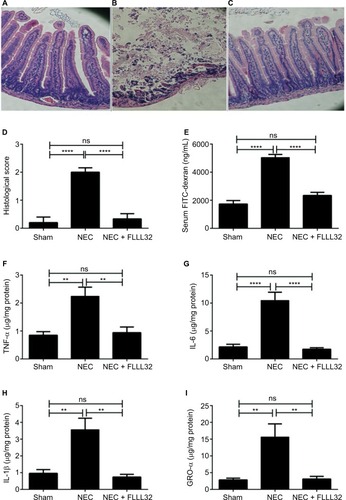
NEC was induced using the DK NEC model as described in the “Methods” section. shows representative histological pictures of sections obtained from the three groups. As expected, sham pups had normal villi and sub-mucosal structure (). The untreated NEC group had moderate to severe intestinal villi damage (). More importantly, pups in the NEC + FLLL32 had mild evidence of intestinal injury (). To further determine the incidence and severity of NEC in the groups, we used the modified NEC scoring system.Citation26 Pretreatment with FLLL32 reduced intestinal injury score from a mean of 2 in the untreated group to a mean of 0.33 in the NEC + FLLL32 group (). In addition, the FLLL32 reduced the incidence of NEC from 72% (13/18) in the untreated NEC group to 8% (1/12) in the treated group (p=0.001).
Figure 3 FLLL32 attenuates intestinal inflammation and injury in DK NEC model.
Notes: Representative H&E pictures from pups in the sham group (A), untreated NEC group (B), and NEC + FLLL32 group (C); 20× magnification. (D) Histological NEC scoring was obtained by two pathologists blinded to the groups. (E) FLLL32 preserved intestinal permeability in the NEC + FLLL32 group compared to the untreated group and control group. FLLL32 pretreatment reduced the levels of proinflammatory cytokines, TNF-α (F, p=0.001), IL-6 (G, p<0.001), IL-1β (H, p=0.009), and GRO-α levels (I, p=0.034) compared to pups in the untreated NEC group. Data are mean ± SEM. Results are representative of at least three separate experiments. **p<0.05; ****p<0.001.
Abbreviations: DK, dithizone/Klebsiella; FITC, fluorescein isothiocyanate; GRO-α, growth-regulated oncogene-alpha; H&E, hematoxylin and eosin; NEC, necrotizing enterocolitis; TNF-α, tumor necrosis factor-alpha; IL, interleukin; SEM, standard error of mean; ns, not significant.
FLLL32 protects intestinal barrier in a DK NEC model
Although the etiology of NEC remains incompletely understood, evidence shows that breakdown of the intestinal barrier is a predisposing factor in the development of NEC.Citation9,Citation29–Citation31 To determine whether FLLL32 protection against NEC was associated with preservation of intestinal barrier, intestinal permeability was assessed by oral administration of the FITC-dextran (4 kDa) to all surviving pups. Concentrations of FITC-dextran in serum were measured 4 hours after administration. As shown in , the sham group had low FITC-dextran levels in the blood. Pups in the untreated NEC group had the highest levels of FITC-dextran in the blood, reflecting increased intestinal permeability. Pups in NEC + FLLL32 group had significantly decreased FITC-dextran in blood to near sham levels, indicating that pretreatment with FLLL32 protected intestinal barrier (p<0.0001).
FLLL32 decreases intestinal inflammatory cytokine protein levels in DK NEC model
Consistent evidence demonstrates the important role of inflammatory cytokines in the pathogenesis of NEC.Citation5–Citation9,Citation32 To determine the effect of FLLL32 on intestinal cytokine levels, we compared the levels of IL-1β, IL-6, growth-regulated oncogene-alpha (GRO-α), and TNF-α in intestinal tissue lysates from pups in the three groups. Similar to other studies,Citation25 levels of all inflammatory cytokines were significantly increased compared to sham (). Interestingly, pups in the NEC + FLLL32 group resulted in 2.5-fold decrease in TNF-α (, p=0.001), 5.8-fold decrease in IL-6 (, p<0.001), 4.8-fold decrease in IL-1β (, p=0.009), and 5-fold decrease in GRO-α levels (, p=0.034) compared to pups in the untreated NEC group. Intestinal IL-10 levels did not differ between NEC + FLLL32 and untreated NEC groups (data not shown).
Discussion
NEC is a devastating disease characterized by inflammation and injury to the small intestine. Despite early and aggressive treatment, NEC can quickly progress to frank bowel necrosis and death.Citation4,Citation33 Although the pathogenesis of NEC is still unclear, observations suggest a complex interplay between prematurity, formula feeding, hypoxia, and altered bacterial colonization.Citation9,Citation34,Citation35 In premature infants, developmental immaturity of mucosal barrier and increased expression of TLR4 on the surface of the intestinal epithelium render the gut highly reactive and susceptible to injury from various stimuli.Citation7 This leads to an exaggerated proinflammatory cytokine release, leukocyte infiltration, altered epithelial barrier, intestinal necrosis, and bacterial translocation across the lumen.Citation36,Citation37
FLLL32 is a curcumin analog with improved solubility and bioavailability compared to the parent product.Citation22 Although FLLL32 has been extensively used in cancer models,Citation22,Citation38,Citation39 its use in inflammation, specifically in NEC, has not been explored. Our results demonstrate that FLLL32 given orally at 25 mg/kg is well tolerated in neonatal mice pups. We also show that similar to curcumin,Citation16 prophylactic administration of FLLL32 reduces the incidence and severity of NEC in DK mode. FLLL32 also preserved intestinal permeability and reduced intestinal proinflammatory cytokines, both of which play important roles in the pathogenesis of NEC.Citation5,Citation6,Citation9,Citation29–Citation31 Although we did not perform a comparison of FLLL32 and curcumin in NEC, our results show that FLLL32 was superior to curcumin in preserving intestinal barrier function in vitro. The protective mechanism of FLLL32 could be through STAT3 inhibition, which has been shown to modulate PAF-induced TLR4 expression in intestinal epithelial cells in vitro.Citation40
Conclusion
Oral FLLL32 decreases intestinal inflammation and injury in a DK NEC model. Further studies are needed to better understand the mechanisms underlying its protective effects.
Acknowledgments
We would like to acknowledge Randle Gallucci, PhD, and Kathryn Burge, PhD, in helping with manuscript editing. This paper was presented in part at the Pediatric Academic Society Meeting, April 28, 2015.Citation41
Disclosure
The authors report no conflicts of interest in this work.
References
- NeuJWalkerWANecrotizing enterocolitisN Engl J Med2011364325526421247316
- RoweMIReblockKKKurkchubascheAGHealeyPJNecrotizing enterocolitis in the extremely low birth weight infantJ Pediatr Surg1994298987990 discussion 990–9817965535
- PatelRMKandeferSWalshMCEunice Kennedy Shriver National Institute of Child Health and Human Development Neonatal Research NetworkCauses and timing of death in extremely premature infants from 2000 through 2011N Engl J Med2015372433134025607427
- SrinivasanPSBrandlerMDD’SouzaANecrotizing enterocolitisClin Perinatol2008351251272x18280885
- FrostBLJillingTCaplanMSThe importance of pro-inflammatory signaling in neonatal NECSemin Perinatol200832210010618346533
- CaplanMSSimonDJillingTThe role of PAF, TLR, and the inflammatory response in neonatal necrotizing enterocolitisSemin Pediatr Surg200514314515116084401
- SodhiCPShiXHRichardsonWMToll-like receptor-4 inhibits enterocyte proliferation via impaired beta-catenin signaling in necrotizing enterocolitisGastroenterology2010138118519619786028
- EganCESodhiCPGoodMToll-like receptor 4–mediated lymphocyte influx induces neonatal necrotizing enterocolitisJ Clin Invest2016126249550826690704
- LeaphartCLCavalloJGribarSCA critical role for TLR4 in the pathogenesis of necrotizing enterocolitis by modulating intestinal injury and repairJ Immunol200717974808482017878380
- NealMDJiaHEyerBDiscovery and validation of a new class of small molecule Toll-like receptor 4 (TLR4) inhibitorsPLoS One201386e6577923776545
- GoodMSiggersRHSodhiCPAmniotic fluid inhibits Toll-like receptor 4 signaling in the fetal and neonatal intestinal epitheliumProc Natl Acad Sci U S A201210928113301133522733781
- JurenkaJSAnti-inflammatory properties of curcumin, a major constituent of Curcuma longa: a review of preclinical and clinical researchAltern Med Rev200914214115319594223
- GoelAKunnumakkaraABAggarwalBBCurcumin as “Curecumin”: from kitchen to clinicBiochem Pharmacol200875478780917900536
- BasnetPSkalko-BasnetNCurcumin: an anti-inflammatory molecule from a curry spice on the path to cancer treatmentMolecules2011166456721642934
- ZhongKCurcumin mediates a protective effect via TLR-4/NF-κB signaling pathway in rat model of severe acute pancreatitisCell Biochem Biophys201573117518025711190
- JiaSHWeiHYuJLWeiXDZhangXPLiJCProtective effects of curcumin on neonatal rats with necrotizing enterocolitisZhongguo Dang Dai Er Ke Za Zhi [Chin J Contemp Pediatr]2010122132136
- SalhBAssiKTemplemanVCurcumin attenuates DNB-induced murine colitisAm J Physiol Gastrointest Liver Physiol20032851G235G24312637253
- ZhangXWuJYeBWangQXieXShenHProtective effect of curcumin on TNBS-induced intestinal inflammation is mediated through the JAK/STAT pathwayBMC Complement Altern Med201616129927544348
- AliTShakirFMortonJCurcumin and inflammatory bowel disease: biological mechanisms and clinical implicationDigestion201285424925522950087
- LubbadAOriowoMAKhanICurcumin attenuates inflammation through inhibition of TLR-4 receptor in experimental colitisMol Cell Biochem20093221–212713519002562
- LiuYFuchsJLiCLinJIL-6, a risk factor for hepatocellular carcinoma: FLLL32 inhibits IL-6-induced STAT3 phosphorylation in human hepatocellular cancer cellsCell Cycle20109173423342720818158
- BillMANicholasCMaceTAStructurally modified curcumin analogs inhibit STAT3 phosphorylation and promote apoptosis of human renal cell carcinoma and melanoma cell linesPLoS One201278e4072422899991
- IkenouchiJFuruseMFuruseKSasakiHTsukitaSTsukitaSTricellulin constitutes a novel barrier at tricellular contacts of epithelial cellsJ Cell Biol2005171693994516365161
- SuzukiTYoshinagaNTanabeSInterleukin-6 (IL-6) regulates claudin-2 expression and tight junction permeability in intestinal epitheliumJ Biol Chem201128636312633127121771795
- ZhangCShermanMPPrinceLSPaneth cell ablation in the presence of Klebsiella pneumoniae induces necrotizing enterocolitis (NEC)-like injury in the small intestine of immature miceDis Model Mech20125452253222328592
- MusemecheCCaplanMHsuehWSunXKellyAExperimental necrotizing enterocolitis: the role of polymorphonuclear neutrophilsJ Pediatr Surg199126910471049 discussion 1049–10501941482
- CaniPDBibiloniRKnaufCChanges in gut microbiota control metabolic endotoxemia-induced inflammation in high-fat diet-induced obesity and diabetes in miceDiabetes20085761470148118305141
- GuptaJNebredaARAnalysis of intestinal permeability in miceBio-protocol2014422e1289
- HackamDJGoodMSodhiCPMechanisms of gut barrier failure in the pathogenesis of necrotizing enterocolitis: Toll-like receptors throw the switchSemin Pediatr Surg2013222768223611610
- LiuYFathereeNYMangalatNRhoadsJMLactobacillus reuteri strains reduce incidence and severity of experimental necrotizing enterocolitis via modulation of TLR4 and NF-κB signaling in the intestineAm J Physiol Gastrointest Liver Physiol20123026G608G61722207578
- HalpernMDDenningPWThe role of intestinal epithelial barrier function in the development of NECTissue Barriers201531–2e100070725927016
- ClaudECWalkerWABacterial colonization, probiotics, and necrotizing enterocolitisJ Clin Gastroenterol200842suppl 2S46S5218520617
- HullMAFisherJGGutierrezIMMortality and management of surgical necrotizing enterocolitis in very low birth weight neonates: a prospective cohort studyJ Am Coll Surg201421861148115524468227
- JillingTSimonDLuJThe roles of bacteria and TLR4 in rat and murine models of necrotizing enterocolitisJ Immunol200617753273328216920968
- LuPSodhiCPHackamDJToll-like receptor regulation of intestinal development and inflammation in the pathogenesis of necrotizing enterocolitisPathophysiology2014211819324365655
- ClaudECWalkerWAHypothesis: inappropriate colonization of the premature intestine can cause neonatal necrotizing enterocolitisFASEB J20011581398140311387237
- LinPWNasrTRStollBJNecrotizing enterocolitis: recent scientific advances in pathophysiology and preventionSemin Perinatol2008322708218346530
- BillMAFuchsJRLiCThe small molecule curcumin analog FLLL32 induces apoptosis in melanoma cells via STAT3 inhibition and retains the cellular response to cytokines with anti-tumor activityMol Cancer2010916520576164
- FosseySLBearMDLinJThe novel curcumin analog FLLL32 decreases STAT3 DNA binding activity and expression, and induces apoptosis in osteosarcoma cell linesBMC Cancer20111111221443800
- SolimanAMichelsenKSKarahashiHPlatelet-activating factor induces TLR4 expression in intestinal epithelial cells: implication for the pathogenesis of necrotizing enterocolitisPLoS One2010510e1504420976181
- EckertJScottBLawrenceSMFLLL32, a curcumin analog, ameliorates intestinal injury in necrotizing enterocolitisPoster presented at: Pediatric Academic Society MeetingApril 28, 2015San Diego, CA, USA