
Abstract
Recently, more and more works have focused and used extensive resources on atherosclerosis research, which is one of the major causes of death globally. Alongside traditional risk factors, such as hyperlipidemia, smoking, hypertension, obesity, and diabetes, mechanical forces, including shear stress, pressure and stretches exerted on endothelial cells by flow, is proved to be crucial in atherosclerosis development. Studies have recognized the mechanosensitive Piezo1 channel as a special sensor and transducer of various mechanical forces into biochemical signals, and recent studies report its role in atherosclerosis through different mechanical forces in pressure, stretching and turbulent shear stress. Based on our expertise in this field and considering the recent advancement of atherosclerosis research, we will be focusing on the function of Piezo1 and its involvement in various cellular mechanisms and consequent involvement in the development of atherosclerosis in this review. Also, we will discuss various functions of Piezo1 involvement in atherosclerosis and come up with new mechanistic insight for future research. Based on the recent findings, we suggest Piezo1 as a valid candidate for novel therapeutic innovations, in which deep exploration and translating its findings into the clinic will be a new therapeutic strategy for cardiovascular diseases, particularly atherosclerosis.
Graphical Abstract
Introduction
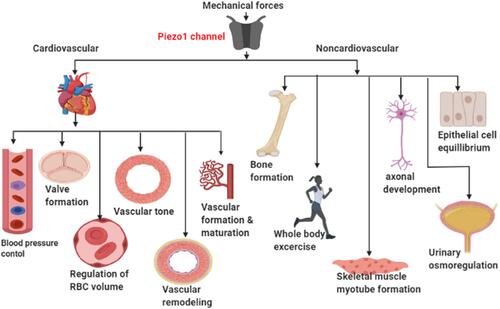
Atherosclerosis (AS) is a major global killer, and is the basis of cardiovascular diseases (CVDs) comprising myocardial infarction (MI), ischemic heart disease (IHD) and stroke. AS is a multifactorial inflammatory disorder,Citation1–Citation4 and is the root of cardiovascular disease.Citation5 AS remains a leading cause of death globally, claiming millions of lives every year, as estimated in the 2015 global burden of disease study (17.9 million) about 30% of whole death globally.Citation6 There is an increased in its prevalence due to the global rise in obesity and diabetes, with more than half of the world population becoming obese, leading to a steep rise in cardiovascular diseases (CVDs) burden; it is estimated that the global death from CVDs may go beyond 23.6 million by the year 2030. At the onset of AS, endothelial cell (EC) defense is triggered by a number of factors, including mechanical forces such as shear stress and stretching, which leads to EC adaptive responses thereby developing a distinct phenotype to interact with other traditional risk factors,Citation7 and facilitate the initiation of AS. The significance of mechanical forces in cardiovascular physiology and pathology have been reported for decades. Shear stress mediated by Piezo1 is among the significant factors and the initiator of inflammation and vascular endothelial cell (VEC) dysfunction.Citation8 Cells detect and liberate mechanical forces using various biochemical and molecular mechanisms. In the cardiovascular system, frictional force and pressure are generated in the vessels, endothelial Piezo1 senses and transduces such forces which are the determinant of the physiological competence of the system throughout life. Piezo1 serves a crucial function in angiogenesis, embryonic development and is crucial for endothelial shear stress-senses, blood pressure regulation, and physical activities during exercise etc.Citation9 As our knowledge of this field increases, various functions of Piezo1 are emerging. Piezo1 is a significant player in cardiovascular physiology (), renal and hematopoietic systems.Citation10 Its function is known in embryonic vessel development,Citation11 RBC stability in both human and mouse,Citation9 control of blood pressure, physical activities, hypertension-dependent arterial remodeling, urinary osmoregulation, epithelial equilibrium and axonal development.Citation11 It also serves the function of adjusting erythrocyte volume,Citation12–Citation14 and in prostate cancer.Citation15 Piezo1 global knockout in mice leads to embryonic fatality,Citation10,Citation12,Citation16–Citation19 indicating its significance during embryogenesis.Citation20 Additionally, severe defects in vessel maturation and remodeling, and disability in NO production and vessel dilation responding to flow were both observed following Piezo1 global deletion or EC-specific disruption.Citation21 Also, Piezo1 in partnership with Piezo2 serves the function of baroreceptor reflex as their dual disruption obliterates baroreceptor responses to sodium nitroprusside or phenylephrine.Citation22 Piezo1 appears to function in a wide area of cell biology, as illustrated in , and its pathologic importance in humans was also indicated.Citation11,Citation23–Citation25 The broad concept of Piezo1 functions was also reviewed in recent articles.Citation9,Citation26–Citation28 Collectively, Piezo1 remains a significant player in a broad area of cell physiology, considering its expression and functions in diverse tissues, not limited to the cardiovascular system (). This indicates that defects or mutations in this channel may impair various functions in ECs and other cell types. Therefore, pharmacological manipulation of either activation or inhibition of this channel may come up with a new trend in clinical practice. Different abnormalities or up/downregulation of cellular activities were observed in several studies following the silencing/blocking, deletion, cell specific or global knockout and deficiency of Piezo1.Citation9,Citation28–Citation33 Since the recognition of Piezo1 &2 in 2010, they have been regarded as one of the most significant classes of mechanosensory proteins. It is now feasible to investigate the mechanism of Piezo1 engagement in physiological and pathophysiological processes as a result of recent developments in understanding their topology, the identification of the agonists Yoda1, Jedi1&2 as well as the antagonist GsMTx4, Ruthenium Red. Recently, cardiovascular aspects of Piezo1 channel have been reviewed elsewhere, but here we review the current understanding of Piezo1 channel and specifically focus on its immune/inflammatory mechanisms of atherosclerosis involving different mechanotransduction processes as summarized in the graphic abstract, which have not been reviewed to date. We also raise awareness of the realization of its pharmacology and suggested new directions for future research toward the development of advanced therapeutic strategies for prevention and management of atherosclerosis.
Figure 1 Cardiovascular and non-cardiovascular functions of Piezo1: The schematic diagram demonstrating a wide range of Piezo1 functions in different areas of physiology including cardiovascular and non-cardiovascular roles from the activation of Piezo1 by mechanical force branching from left to right arrows, pointing different organ and cell /tissue.
Structure of Piezo1 Channel
Human mechanically activated (MA) channel molecular identity was mysterious for a long time until the revolutionary discovery of Piezo1 & 2, which may increase our understanding of cellular signaling and mechanotransduction. Piezo1 proteins are collectively arranged to form a three blade-like propeller inserted within the lipid bilayer which makes the central ion pore that senses mechanical forces.Citation9,Citation34–Citation36 Its extracellular propeller domain serves as a detector of flow related-shear and other mechanical stress.Citation16,Citation37 In respect to structural components, Piezo1 can be categorized as follows.
Exceptional 38‑TM Topography
With advancement of Piezo1 research, high-resolution structures of mice Piezo1 (mPiezo1) were shortly disclosed, showing that each subdomain has a special 38-TM topography. The inner helixes (IH) and outer helix (OH) of the center of the pore module are the 2 TM subdomains (TM37 and TM38) nearest to the protein’s core. The remaining 36 TM regions (TM1-36) are organized into a blade resembling a curved structure with 9 repeating folds comprising four TM regions each, known as transmembrane helical units (THUs).Citation38–Citation40
Exceptionally Curving Blades
Each subdomain’s 9 peripheral THUs produce bladelike architectures, each twisted clockwise. As seen from a line parallel to the plasma membranous planes, next to the TM25–TM36 and periphery of the TM13-24 these are based at a 100° and a 140° angle, respectively. The L-shaped helical structures produced by TM13, TM17, TM21, TM25, and TM29 are another essential feature of the blades. Both structural characteristics tend to be suitable for inducing regional membrane curvature as well as mechano-sensing. Surprisingly, the peripheral TM13-24 tends to be located inside a strongly bent membrane plane, implying that the Piezo1 channel has the ability to bend the membrane in which it sits. Previous research suggested that cell membrane deflection and tension may control Piezo1.Citation26,Citation27,Citation38,Citation39,Citation41
Central Cap
Using topographical predicting simulation, a study by Kamajaya’s team,Citation42 discovered that Piezo1 residues 2210 to 2457 produced an extracellular loop next to the last TM region from the C-terminus, known as C-terminal extracellular domain (CED). The central cap was not expressed when residues 2218 to 2453 were removed from the Piezo1 protein, implying that this area trimerized to produce the central cap. The central cap comprises the CED in the shape of trimeric complexes that encapsulated the extracellular-vestibule (EV) with apertures, disclosed by another study.Citation27,Citation38,Citation39,Citation43
The Ion Conducting Pore
Piezo proteins compose a trimeric ion conducting channel comprising residues 2189 to 2547, that composes the last two TMs. An EV in the cap region, a transmembrane-vestibule (MV) within the membrane, and the intracellular-vestibule (IV) beneath the membrane make up the continuous central channel. The MVs are located on top of and beneath the membrane, and both the EV and IV have an opening that binds them. DEEED (2393–2397), a patch of negative charge residues located in the opening of the extracellular “cap” structure made up of the CED, is necessary for effective ion conducting and determining the preference for cations than anions. Furthermore, divalent calcium ion selectivity, unitary conductance, and aperture blocking can be caused by two essential acidic residues, E2495 and E2496, situated at the CTD-constituting IV.Citation26,Citation39,Citation40,Citation44
The Intracellular Beam
Piezo1 possess 3 beam-like structures from intracellular aspect, each measuring 90 nm in length and arranged at a 30° angle to the membranous plane. The beam composition is made up of residues H1300-S1362. The long intracellular THU7-8 loop has about 390 residues, which could supply the beam with a structural foundation for force transmitting. The 3 longer intracellular beams serve as a functional barrier between the distal THUs and the central ion conducting orifice, as well as supporting the entire TM framework. The mutated protein was missing after residues 1280 to 1360 were removed, indicating the beam’s structural significance.Citation26,Citation27,Citation39
The Anchor
The OH-IH pair is connected into the C-terminal domains (CTD) planes by a hairpin structure called the anchor, which pushes the OH-CED-IH-composing area of 1 subdomain to the adjacent subdomain in a dextral aspect. Three helices (α1, α2, and α3) make up the anchor. The inverted V-shaped structure formed by helices α1 and α2 was discovered to keep the ion-conducting pore’s stability. The longer α3 helical unit interacts with the polar residue-rich α2–3 turning in the anchor and the glutamate-rich portion of the CTD through a lysine-rich anchor-OH linker which runs parallel onto the membranous plane. A few mutations in Piezo1 have been identified to account for serious disorders at regions such as KKKK (2182-K2185), T2143, T2142 (T2127 in human Piezo1), R2514, E2523, and E2522, which are situated at α3 in the anchor. SERCA2, a Piezo-interactor protein, also inhibits Piezo1 by interacting with the anchor-OH linker. These results affirm the anchor portion’s structural and functional significance.Citation26,Citation38,Citation40
Piezo1 Functions
The Role of Piezo1 in Helical Flow
The circulatory system generates a variety of flow patterns within the vascular beds, including laminar, disturbed or turbulent, and oscillatory or helical flow, depending on the nature of vessel.Citation8 Piezo1 channel was recently recognized as a specialized mechanosensor that senses blood flow and integrates the signal into a genealogical program for vascular formation and maturation.Citation45–Citation47 Due to the effects of flow pattern on vascular health, helical flow patterns have received considerable attention in modern days. The helical pattern of flow is symbolized by increased velocityCitation48 with a high shear stress that is known to be sensed by Piezo1,Citation9,Citation27 and is now considered as a physiologic type of flow.Citation48,Citation49 It is now thought to be an atheroprotective flow,Citation50 considering that it could minimize blood cell adherence to the vessel wall, prohibit LDL buildup, and improve perfusion and oxygen delivery. Furthermore, hemodynamic efficacy of a vascular device is enhanced by helical flow,Citation48,Citation51 as summarized in . Piezo1 may sense helical flow and act as a key regulator of vascular health through various mechanisms, as it senses and transduces different flow patterns into biochemical signals. Therefore, through Piezo1 mechanotransductions, the roles of helical flow might minimize the burdens on vasculature as well as protecting it from atherosclerotic pathologies, intimal hyperplasia and thromboembolism, through the suppression of proinflammatory signaling resulting from low and turbulent shear stress.Citation52 Altogether, these data demonstrate the physiological importance of Piezo1 mediated helical flow, its mechanisms in cardiovascular biology signaling and its therapeutic potentials for targeting atherosclerosis and other CVDs.
Table 1 Summarizing the Suggested Clinical Significance of Helical Flow
Piezo1 in VSMCs Shear Stress-Induced Plaque Formation
Piezo1 is the specialized sensor and transducer of mechanical forces including shear stress. It enables EC,Citation9,Citation53 and vascular smooth muscle cells (VSMCs),Citation20 to detect and respond to changes in their environment. Turbulent shear senses via Piezo1,Citation8 increases EC protein and gene expressions of VSMC mitogens including platelet-derived growth factors (PDGF), ET-1, and vascular endothelial growth factor (VEGF). EC expression of VSMCs migration inhibitor (plasminogen activator inhibitor (PAI)-1) and effective suppressors of cellular growth and migration, such as NO and TGFβ, are all downregulated by turbulent shear. The EC downregulation of these growth inhibitors by disturbed shear encourages VSMCs to migrate into the intima via an interrupted internal elastic lamina (IEL). VSMCs develop synthetic phenotype in the intima, replicate and generate collagen, including other (ECM) proteins. With time, VSMCsCitation54 together with fibroblast that is known to be regulated by Piezo1,Citation55 form the fibrous cap along with lipid core, building up what is known as atherosclerosis plaque.
Piezo1 Channel as Shear Sensors
Shear sensing is vital in cardiovascular physiology, and shear stress is among the factors guiding the developing vessel during embryogenesis.Citation17 Piezo1 is a specialized and authentic shear stress sensor, as proved by much evidence. ECs detect shear stress and convey the signaling inside the cells, which induces cell responses to changes in its surroundings; abnormality in these responses could result in various disorders, including CVDs.Citation56 In response to shear stress, Piezo1 activates cascades of downstream mechanisms, which alter the cell behavior based on the type of stimuli.Citation17,Citation57 It senses physiological shear to enhance angiogenesis and vessel maturation.Citation34 Earlier discoveries about Piezo1 unveiled its importance for shear stress detection.Citation18 With Piezo1, EC oriented and aligned in the flow direction, while in its absence, EC failed to do so.Citation12,Citation34 Collectively, these qualify Piezo1 as an authentic shear stress sensor; and tuning Piezo1 through pharmacological approaches may change the story of clinical practice. Shear stress triggers EC Piezo1, which causes Ca2+ inflow and phosphorylating of AKT and eNOS, resulting in enhanced NO generation and VSMC-dependent vessel dilatation. Comparatively, VSMC Piezo1 is triggered by stretch and is active in vascular remodeling mechanisms under pathological conditions, resulting in a reduction in vessel diameter. Both Piezo1-dependent processes successfully sustain basal BP control.Citation58 In human embryonic kidney cells, transfecting Piezo1 caused prompt shear stress–evoked Ca2+ influx or ion current, shear stress quickly stimulates endogenous Piezo1 in membrane patches extracted from native endothelium, and Piezo1 knockout disrupts embryonic vascular maturation, which is thought to be caused by shear stress.Citation9
Piezo1 Correlation with Inflammation and Atherosclerosis
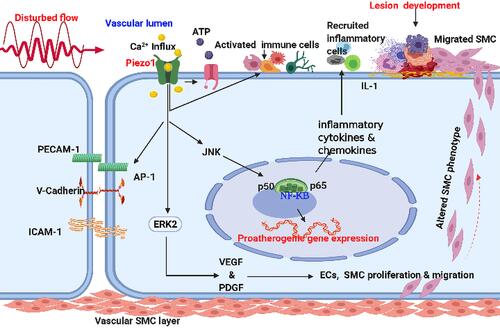
Increasing evidence has reported Piezo1 involvement in various mechanisms of cell biology, including health and disease. Nearly all cells, tissues and organs are exposed to various degrees of mechanical forces, and Piezo1 is the principal mechano-sensor in various cells and tissues, thus linking Piezo1 to multiple mechanisms of inflammation and various disease pathogenesis particularly atherosclerosis. Mechanical forces, including shear stress, are the primary stimuli for cell physiology, since they supply signaling for cellular homeostasis. Piezo1 is known to mediate wide varieties of such forces, therefore dysregulation of Piezo1 signal may alter homeostasis and result in disease processes such as atherosclerosis. ECs respond to these stimuli such as laminar or turbulent shear mediated by Piezo1, and this triggers pro- or anti-inflammatory signals leading to inflammation and atherosclerosis depending on the flow pattern. Piezo1’s significance in cardiovascular physiology and pathology is becoming more widely recognized, hence the next section will discuss in detail the Piezo1 mechanisms in inflammation and atherosclerosis. Furthermore, evidence shows that unidirectional and turbulent flow senses by Piezo1, causes distinct signaling in ECs, leading to an anti- or pro-atherogenic phenotype.Citation8 Each of the unidirectional and turbulent flow triggers initiate signal thoroughfare involving Piezo1.Citation8,Citation59 The trigger of NF-κB upregulates proinflammatory as well as pro-atherogenic genes,Citation60,Citation61 leading to growth of AS, while KLF triggering sustains vessel architecture ( and ). Furthermore, various in vivo and ex vivo studies on pre-flowed ECs culture, demonstrate that unidirectional and turbulent flow-induced athero-protective and atherogenic signals both require Piezo1.
Figure 2 Piezo1 flow induced anti-atherogenic signaling mechanism: schematic diagram, demonstrating laminar flow (anti-atherogenic flow) activates Piezo1 channel, leading to Ca2+ influx and then triggers the generation of eNOS that leads to the expression of KLF2 & KLF4 through MAPK and results in the deactivation of adhesion molecules and AP-1 (red inhibition arrow) as well as downregulations of growth factors VEGF/PDGF, proinflammatory chemokines and cytokines, cell proliferation and migration followed by anti-atherogenic gene expression and athero-protection.
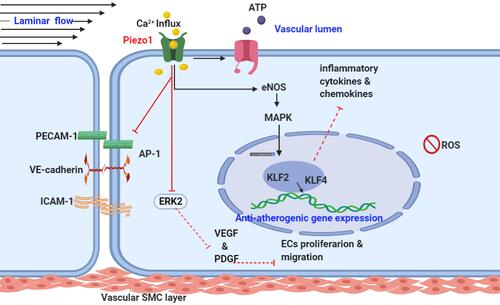
Figure 3 Piezo1 flow induced proatherogenic signaling mechanism: schematic diagram demonstrating disturbed/oscillatory (pro-atherogenic flow) activation of the Piezo1 channel, leading to Ca2+ influx followed by the activation of JNK, NF-KB, p50 and p65, ERK2 pathway. And the atherogenic gene expression, growth factors VEGF/PDGF, proinflammatory chemokines, cytokines. And cellular adhesion molecules, which result in the activation of immune cells, inflammatory cell recruitment, and ECs and SMCs proliferation and migration, leading to endothelial inflammation, increased permeability, cell damage, plaque formation, and atherosclerosis development.
Piezo1 Mediated Mechanisms in the Initiation and Progression of Inflammation and Atherosclerosis
The majority of prior studies on atherosclerosis pointed to elevated LDL levels as the primary culprit.Citation62 Only recent works recognized that hypercholesterolemia and atherosclerosis development are linked via inflammatory processes. Therefore, immunological response and inflammation have been regarded as key contributors in the initiation and amplification of atherosclerosis.Citation63,Citation64 Mechanosensitive Piezo1 channel was recently reported to play a crucial role in immune cells and inflammatory processes.Citation65–Citation68 Over time, research has shed more light on inflammatory responses in atherosclerosis, providing strong evidence that inflammation is a major factor in all stages of atherogenesis.Citation69 Furthermore, inflammatory signaling increases thrombosis, which is responsible for ischemic heart disease and the majority of myocardial infarctions and cerebrovascular diseases.
Stages of Atheroma Formation
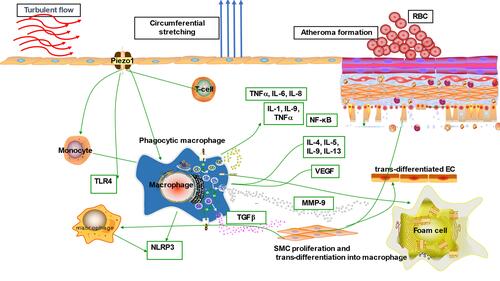
(A) Initially EC undergo inflammatory triggers in response to noxious stimuli,Citation70 including atherogenic shear from flow mediated by Piezo1,Citation71 as discussed in Piezo1 Mediated Shear-Induced Inflammation and Atherosclerosis; (B) monocytes recruitment to atheroma region discussed in 5.3; (C) cytokines and chemoattractive molecules engaged in promoting the recruiting of additional immune/inflammatory cells to the intima discussed in 5.5; (D) later monocyte differentiates into macrophages and engulfs lipoprotein particles to transform into lipid engorged foam cells; (E) then foam cells release proinflammatory molecules, and growth factors including reactive leading to the proliferation, migration, discussed in 5.5, and phenotypic switching or transdifferentiation of VSMCs into SMC derived macrophages in the intima.Citation72,Citation73 (F) Apoptosis of macrophages leading to the development of “necrotic” core of the mature plaque; (G) macrophages and SMC augment the process,Citation74 through secretion of matrix metalloproteinases (MMPs) particularly MMP-9,Citation75 which leads to the degradation of extracellular matrix, resulting in the plaque’s fibrous cap thinning; (H) plaque rupture due to weakness of the fibrous cap leading to thrombogenic coagulation and finally thrombosis and obstruction of vessel lumen,Citation76 which limits the tissue perfusion and results in ischemic heart disease.Citation69,Citation70 Piezo1 signaling is known to play crucial roles in other immune/inflammatory mechanisms of atherosclerosis as discussed in 5.4.
Piezo1 Mediated Shear-Induced Inflammation and Atherosclerosis
Beside the established risk factors of atherosclerosis, it is convincing that turbulent shear mediated by Piezo1 is crucial for EC activation, leading to initiating of inflammation and atherosclerosis.Citation8,Citation77,Citation78 Disturbed flow sensed via Piezo1 produces turbulent shear, which is a considerable risk for atherosclerosis, as studies reported its effect on EC.Citation78,Citation79 Turbulent shearCitation80 triggers pro-inflammatory EC phenotype; disorders such as AS appeared to originate from such EC phenotypes.Citation8 Numerous studies suggested Piezo1 associates with different inflammatory pathways and signaling mechanisms in the development of AS (). Piezo1 was recently found to be expressed and function in inflammatory cells including monocytes, macrophagesCitation65 and T-cells.Citation81 Studies reported that activation of monocytes and macrophage by Piezo1 triggers proinflammatory signals leading to expression of various cytokines and chemokines,Citation65–Citation67 a crucial event during atherogenesis. Cytokines such as TNF-α, IL-1, 2, 3, 6, 10, 12, 15, 18, CXCL8, IFN-γ, M-CSF, TGF-β1, 2, and 3 are critical components of inflammation and play a significant role in AS pathogenesis. The most interesting fact is that in the lack of Piezo1, inflammatory cytokines and chemokines, as well as the transcriptional factor, hypoxia-inducible factor 1 (HIF-1), were not expressed by macrophages and monocytes subjected to pressure cycles. Additionally, PECAM-1, VCAM-1 and ICAM-1Citation81 are also found in disturbed flow regions of ECs, and reduced shear upregulates the P-selectinCitation82 and monocyte chemotactic protein-1 (MCP-1) mRNA expression in correlation with an increased number of monocytes bounded to EC. Interestingly, following the loss of Piezo1, the rise in Vcam-1 expression and the number of CD68-positive cells was markedly decreased, and turbulent flow-evoked inflammation signals were also blocked. Comparatively, subjecting human umbilical artery endothelial cells (HUA ECs) to turbulent flow leads to NF-κB triggering, as evidenced by p65 phosphorylating at serine 536. Furthermore, in athero-susceptible sites, there were less integrin activities, inflammatory signals and atherosclerosis, following the selective depletion of endothelial Piezo1 or Gq/G11 in mouse. Both in vivo and ex vivo EC Piezo1 and Gq/G11 mediate inflammatory EC signal in response to turbulent flow, leading to EC dysfunction and AS, also equal signaling pathway detects laminar flow and triggers eNOS.Citation21,Citation81,Citation83 Hence, turbulent and laminar flow tend to stimulate Piezo1- and Gq/G11-mediating signals, which mediates both athero-protective and proatherogenic signals, described in the figure legendsCitation8,Citation84–Citation86 ( and ). In AS, inflammation involves several mechanisms and cross-talk between various pathways, therefore targeting inflammation through Piezo1 approach could be wise. Altogether, these findings support the idea that Piezo1 might not just mediate unidirectional flow-evoked anti-atherogenic signals but is also necessary for turbulent flow-mediated endothelial inflammation and atherogenesis.
Piezo1 Mediated Immune/ Inflammatory Response in Atherosclerosis
Mechanical stress exerted on EC in the form of disturbed flow is among the primary activators of the endothelial immune defense system. Activation of EC is always followed by the recruitment of immune cells such as monocytes, which later differentiate into macrophages and transform to lipid-engorged foam cells, the hallmark of AS. Numerous studies reported the dominance of immune cells on the site of an earlier plaque. Triggered T cells, including macrophages, have been revealed to reside on early and part of matured AS plaques, and their transcription components speed up the plaque advancement, indicating the significant function served by innate and adaptive immunity in AS cytopathology.Citation64,Citation87,Citation88 Turbulent flow is thought to activate the endothelial immune system that initiates the process of AS. Traditionally ECs oppose adherence of inflammatory cells, but EC exposure to turbulent shear mediated by Piezo1 activates the endothelial expressions of adhesive molecules that promote the attaching of immune cells (). Studies reported the significant role of mechanical force on immune cell functioning, and Piezo1 appears to be crucial in T-cell activation and recruitment.Citation89,Citation90 Additionally, findings by Solis et al. revealed the physiologic function of Piezo1, including mechano-sensing in immunity.Citation65 Adaptive immunity has a significant effect on atherogenesis, with pro- and anti-atherogenic impacts exerted by T cells sub-classes.Citation91 As reported by Solis, Piezo1 detects oscillatory pressure in myeloid cells, and triggers pro-inflammatory responses. Mechanical activation of macrophages, including monocytes, stimulates expressions of pro-inflammatory and chemo-attractive mediators, which all rely on Piezo1.Citation65 Intriguingly, in lack of Piezo1, macrophages, including monocytes subjected to cyclical pressure, could not express inflammatory cytokines and chemokines.Citation67 Altogether these findings demonstrated that Piezo1 is crucial for immune response during atherogenesis ().
Piezo1 Activation of NLRP3 Inflammasome and TLR4 Signaling in Regulation of Inflammation and Atherosclerosist
Atherosclerosis is a chronic progressive inflammatory disorder that is believed to be linked to the triggering of NLRP3 inflammasome and TLRs signaling. According to a growing body of data, NLRP3 inflammasome and TLR4 appears to have a causal role in the onset and advance of atherosclerosis.Citation92,Citation93 Given the importance of mechanical forces in regulating cellular and tissue growth, as well as in disease pathogenesis,Citation94 including atherosclerosis, mechanosensitive Piezo1 channel could be considered as a crucial player in various mechanisms of CVDs particularly atherosclerosis.Citation28 Piezo1 channel was reported to be expressed and function in vascular cells including ECs,Citation95 VSMCs,Citation20 as well as immune cells (myeloid cells) such as monocyte, macrophages,Citation65 B and T cells.Citation68,Citation90 On the other hand, NLRP3 and TLRs,Citation96,Citation97 are also expressed by both immune cells and endothelium. NLRP3 inflammasomes and TLRs are well-known multi-protein complex immunological sensors that augment inflammation in response to a variety of danger-signaling ligands inclusive of pathogen-associated molecular patterns (PAMPs) from invading microbes (e.g., LPS) or mislocated commensal pathogens, as well as danger-associated molecular patterns (DAMPs) or alarmins like endogenous factors,Citation98 inclusive of RNA and DNA, HMGB1, amyloid-β, cholesterol crystals, mitochondrial damage, or ROS, as well as noxious stimuli like disturbed shear mediated by Piezo1, which results in accumulation and triggering of caspase-1. Innate immune responses mainly rely on the detection of PAMPs and DAMPs via the pattern-recognition receptors (PPR), including Toll-like receptors and NLRs, particularly TRL4 and NLRP3. A recent study shows that application of Yoda1 in BV2 cell upregulates the expressions of TLR4, moreover, significant elevation of TLR4 and Piezo1 expressions were seen following the application of Yoda1 and LPS; this finding suggest that Piezo1 is crucial for TRL4 signaling.Citation99 NLRP3 activators are thought to cause one or more downstream cellular processes or diseases rather than directly interacting with NLRP3. The NLRP3 inflammasome is triggered by two distinct signals: toll-like receptor 4 (TLR4) ligands lipopolysaccharide (LPS) binding to its receptor, which causes NLRP3 and pro-IL-1 transcriptional upregulating via NF-B (1st signaling). TLR4 can also deliver 1st signaling independently of new protein synthesis through its adaptors myeloid differentiation factor 88 (MyD88), interleukin 1 receptor-associated kinase 1 (IRAK1), and IRAK4. NLRP3 activation requires a posttranscriptional alteration; NLRP3 deubiquitination mediates by BRCA1/BRCA2-containing complex subunit 3 (BRCC3) (2nd signaling). The NLRP3 inflammasome is assembled and activated by the second signal through NLRP3-activating substances (e.g., ATP, ROS, oxidized mitochondrial DNA (mtDNA), and other noxious stimuli including mechanical stretch and disturbed or oscillatory shear mediated by Piezo1, followed by proinflammatory caspase-mediated pyroptosis.Citation97,Citation100 Recent findingsCitation92,Citation100–Citation102 suggested that in ECs, various insults like disturbed flow, initiate the NLRP3 inflammasome triggering, and Piezo1 is a newly discovered mechanosensitive channel that senses and transduces different flow patterns into biochemical signals.Citation103,Citation104 In ECs simulation of turbulent flow and oscillating shear extensively intensifies the generation of active caspase-1 and IL-1β.Citation105 Furthermore, Sun et al. reported that Piezo1 triggering enhances the assembly of NLRP3 inflammasome, as indicated by caspase 1 upregulating and generation of IL-1β which was reversed after Piezo1 siRNA transfecting. Interestingly, the abrogation of Piezo1 dependent NLRP3 inflammasome trigger was observed following the Ca2+/NF-κB pathway suppression. The author further revealed that mechanical stretching enhanced expression of Piezo1 and intracellular Ca2+ accumulation that upregulates NLRP3 through the NF-B pathway triggering. It also acts as a direct secondary stimulus, promoting NLRP3 assembly, caspase-1 triggering, and the generation of IL-1β.Citation106 In tissue-resident mice alveolar macrophages, Wu et al. found that cyclical stretching known to be mediated by Piezo1, triggers the NLRP3 inflammasome through mitochondrial ROS generation, suggesting that this mechanism might be connected to lung inflammation caused by mechanical ventilation. Mechanical stress appears to be a risk factor for NLRP3 inflammasome activation, according to this study.Citation107 Whereas another study found that application of prolonged compressive stress to epidermal tissue increased NLRP3 and caspase-1 protein expressions while reduced IL-1β expression.Citation108 Contrarily, another study revealed that in macrophages, cyclical stretching inhibits the NLRP3 inflammasome.Citation109 This implies that further studies are necessary to explain these contradictory findings. The mechanosensitive Piezo1 channel appeared as principal transducer of mechanical cues into Ca2+ dependent signaling, and Ca2+ signaling was critical in various cellular physiology and pathologies. Piezo1 triggering is necessary for various Ca2+ dependent pathways. Many downstream pathways including that of TLR4, NF-B, mTOR, and JNK1,Citation110 which are controlled by intracellular Ca 2+ signals, have been demonstrated to be the major molecular processes involved in immune processes including inflammation and atherosclerosis.
Piezo1 Mechanical Trigger of Proliferation, Migration and Apoptosis in Atherogenesis
Owing to the pulsative aspect of blood supply, vascular cells (VECs & VSMCs) are exposed to hemodynamic forces in the form of shear stress, pressure, circumferential/ distention or stretches. Mechanosensitive Piezo1 senses such forces in various intensities, allowing their translation into biological signals within the cell, leading to triggering of downstream pathways.Citation9,Citation111,Citation112 Depending on whether the cells are subjected to physiologic or sub-physiologic/supraphysiologic mechanical forces, this trigger can differ. Significant physiological mechanical force is crucial in maintaining vascular homeostasis.Citation72 While sub-physiologic or supraphysiologic forces may induce alterations in gene expression that encourage inflammation, cell proliferation, migration, apoptosis and vascular remodeling, that are pivotal process in atherosclerosis development.Citation113,Citation114 Multiple studies demonstrated the effects of Piezo1-mediated mechanical signals on vascular and other cells (immune/inflammatory cells) involving proliferation, migration, apoptosis and remodeling during disease pathogenesis,Citation65,Citation66,Citation68,Citation90,Citation105 particularly atherosclerosis (). Pathological inflammation, cell proliferation, migration and apoptosis of EC, VSMCs and macrophages is the hallmark of atherosclerosis pathogenesis, including several CVDs.Citation115 Piezo1 is necessary for sensing and transduction of various mechanical forces including shear stress, pressure, and cyclic stretch. VSMCs proliferation was reported to be increased by cyclic stretch in vitro.Citation72,Citation73 Piezo1 is a well-recognized mechanically triggered channel that enables Ca2+ influx,Citation47,Citation116 and Ca2+ is a highly recognized modulator of cell proliferation, migration and apoptosis, thus, it serves a crucial function in atherogenesis.Citation8,Citation9 Additionally, ERK and Akt/mTOR pathways are crucial for cell survival, differentiation, proliferation, migrating and apoptosis,Citation117–Citation119 and intracellular Ca2+ signaling can control these pathways.Citation120 Stretching was shown to enhance mice and rabbit VSMCs proliferation by activating the extracellular signal-regulated-kinase (ERK) pathway in a synergistic manner with oxidized LDL and norepinephrine. Furthermore, 15% stretching of mouse aortic SMCs exhibited higher ERK and Akt triggering, as well as an increase in insulin-induced cellular proliferation.Citation72 In pathological stretching, however, unregulated proliferation of ECs was seen due to upregulating of oncogene c-Myc expression in human umbilical vein endothelial cells (HUVEC). HUVECs depleted in Piezo1 display decreased expression of VEGF, cell proliferation, and migration in stationary states, demonstrating the significance of Piezo1 in this mechanism.Citation12 When ECs were stretched, they transdifferentiated into SMCs as elevated expression of specific SMC marker genes (SM22, -SMA, caldesmon-1, SM MHC, and calponin) was observed, while endothelial markers were reduced. The appearance of SMC markers on EC means that during mechanical stretching, EC plasticity towards the SMC phenotype occurs, which may lead to the plaque progression.Citation73 Comparatively, Piezo1 overexpression or dysregulation can lead to increase in Ca2+ overload. The elevated level of intracellular Ca2+ can activate the Akt/mTOR pathways through calmodulin (CaM) or CaM-dependent protein kinase II (CaMKII) and, as a result, accelerate cellular proliferation and migration, important processes in plaque formation and cancer. Piezo1 plays a crucial role in cell migration, as MCF-7 cells can migrate better when Piezo1 is overexpressed. While application of Piezo1 agonist GsMTx-4 impaired the migrating of MCF-7 breast cancer cells.Citation28 Collectively, these results demonstrate that Piezo1 is needed for inflammation cell proliferation, migration, apoptosis and, that these are critical stages in the development of AS.
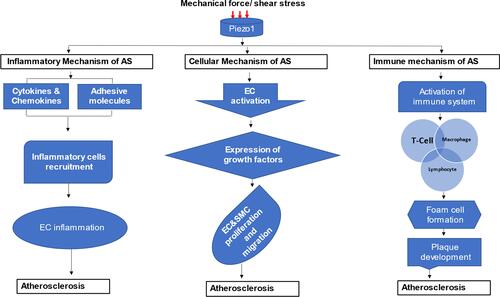
Figure 4 Different mechanosignaling mechanisms of atherosclerosis: schematic diagram demonstrating different signaling thoroughfares and stages of atherosclerotic development from Piezo1 activation by mechanical force, the arrow branching to inflammatory, cellular proliferation and migration, and immune atherosclerosis development.
Piezo1 Pharmacology
The accumulated reports and preceding experiments collectively demonstrate the mechanosensitive value of Piezo1 in both physiology and pathology, particularly atherosclerosis. They may be used as diagnostic biomarkers, as well as pharmacologic and genetic targets for innovative and advanced therapeutic approaches. Despite the infancy of Piezo1 research and its pharmacology, the rapid discovery of its pharmacologic agents presents an outstanding therapeutic opportunity. Recent studies reported the recognition of a few molecules including Yoda1,Citation121 and Jedi1/2,Citation122 as potent activators of Piezo1, and GsMTx-4,Citation123–Citation126 Ruthenium Red (RR)Citation47,Citation127 as its non-specific inhibitors, inclusive of Yoda1 analog called Dooku1,Citation11 and the most recent molecule Tubeimoside1 (TBMS1),Citation128 that have reversible antagonizing capacity of Yoda1-evoked Piezo1 trigger, as well as the binding site including activating mechanisms.Citation112 Altogether, the agonistic and antagonistic capability of the molecules on Piezo1 demonstrated the medicinal and pathophysiological significance of Piezo1 in atherosclerosis and including cancer and other disorders. Nevertheless, Piezo1 drug targeting for therapeutic approaches remains challenging and a difficult task that requires further investigations.
Future Research
Piezo1 channel is a newly identified ion channel, which touches many areas in cell biology. It is expressed in different cell types, and its function may differ in different species, tissues, and local cellular environments. Despite the rapid advancement in Piezo1 research, more roles are yet to be explored. Piezo1 will be a novel candidate for global researchers to further explore its physiological and pathological functions as well as thorough understanding of its pharmacology. Additionally, future investigation on Piezo1 regulation of accompanying mechanisms related to AS will shine a light toward the future prospective and promising therapeutic strategies that may help to reduce the burden of disease and cost of treatment in various conditions, particularly AS in CVDs.
Conclusion
Atherosclerosis is the basis of CVDs, and the critical subject in cardiovascular research and the long-term major issue of global health care. Great efforts and advancement in biomedical as well as clinical research have brought significant achievements in the management of CVDs burden, particularly AS. Despite these remarkable achievements, residual risks yet remain, as many individuals suffer the disability of heart functions leading to the increased CVDs epidemic. AS is a multifactorial disorder; that makes it a complex disease which requires multiple approaches beyond lipid-lowering and lifestyle improvement. Besides these primary and other risks, considering mechanical forces including shear stress in form of disturbed flow should also be crucial. Consistent with the accumulating evidence that specific plaque location is in the curved and branching regions of the arteries where the flow is low and disturbed,Citation129 and immune/inflammatory mechanisms involving Piezo1 signaling play critical role during the process, this shows that even newborn babies are counted in this risk, due to the branching and curved nature of blood vessels. This indicates the importance of turbulent shear, mediated by Piezo1 in the development and progression of atherosclerosis. There have been several experiments that have elucidated the mechanisms of Piezo1 signaling and its regulatory roles in the trigger of various inflammatory signals that initiate downstream events including activation of NLRP3 inflammasome and TLR4. However, we need to be clear that the initiation of signaling necessary inflammatory mechanisms and their downstream effects in NLRP3 inflammasome triggering play critical roles at different stages of triggering and the impacts they generate are miscellaneous. Piezo1 signaling has various impacts on inflammatory cells and their function including the initiation of inflammatory mechanisms and downstream triggering of the NLRP3 inflammasome, TLR4 and related Ca2+ dependent signaling which play crucial roles in the initiation and progression of atherosclerosis. This paper provides a new perspective where not just Piezo1 inflammatory signals and the downstream activation of TLR4 and NLRP3 inflammasome but also the signaling hubs between them are promising strategies to target against atherosclerosis that are worthwhile for future research. In this regard the scientific community should come up with new strategies using interdisciplinary approaches to resolve the challenges of disparity between in vivo physiological systems that have heterogeneity in cell behavior across vascular beds compared with our current homogenic in vitro experimental system. To achieve this, the following arguments should be considered.
Endothelial Model
HUVEC is the most commonly used model in research; the disparity of endothelial heterogeneity across vascular beds is rarely taken into account, in which endothelium from other origins could give different outcomes to the same approach. Indeed for clarity, a single study approach should be tested on endothelial models of multiple origin, as the cell behavior is spatially different across vascular beds.
Individual Mechano-Sensing Protein, Its Correlation with Other Proteins and Signaling Pathways
Several studies reported Piezo1 as an independent mechano-sensor and transducer of itself, independent of other ancillary proteins, while other works reported Piezo1 functions in correlation with other mechanosensitive proteins, such as TRPV4. This could be due to the cross-talk between different proteins in different cellular mechanisms and correlation with signaling pathways in a particular cell activity. Therefore, new approaches and strategies are required to resolve this argument. Further study targeting several mechanisms involving various proteins and their correlation in regulating multiple cellular signaling pathways are necessary, to elucidate more on particular function of each protein individually, and in correlation with others in different mechanism of cellular physiology.
Physiological Model of Flow in Different Vascular Beds and Rhythmic Changes
With physiological flow in vivo, shear pattern is transiently dynamic due to physiological variations in vascular bed, cardiac output or other metabolic needs, compared with ex vivo experimental systems that are commonly employed, the laminar or oscillatory shear pattern applied is always steady with no variation in intensity, amplitude or pattern (seen with in vivo) throughout the experiment. This creates a considerable gap among ex vivo and in vivo systems. Therefore, the shear sensing of Piezo1, the downstream impact, endothelial response and consequent gene expression may vary between ex vivo and in vivo. The transient variations of shear intensity and amplitude which commonly happen with in vivo should be considered when using in vitro experimental systems, since the common experimental systems used only apply steady, laminar, oscillatory or low turbulent shear which is not enough to precisely symbolize in vivo physiological shear experienced by EC. Here arises the requirement for advancing the current ex vivo experimental system to precisely meet the conditions similar to that of the in vivo physiological flow system.
In summary, Piezo1 is a promising candidate in cardiovascular research, due to the evidence already available on its physiological and pathological relevance in various angle of cardiovascular, regardless of its novelty. Its broad expression makes it likely to serve vital functions in regulating undiscovered aspects of cardiovascular physiology and pathology, and produce multiple functional outcomes depending on cell type and mechanisms involved. Therefore, innovative strategies through Piezo1 pharmacological approach will be advantageous for multiple therapies in cardiovascular disease, especially atherosclerosis.
Lastly, an intriguing fact in the Piezo1 pharmacology is the recent discovery of its activator (Yoda1), and inhibitor Grammostola spatulata mechano-toxin 4 (GsMTx4). It shows that with deep understanding of both Piezo1 and its pharmacology, their translation into the clinic may unlock the door and could be the promising therapeutic target for AS burden and other CVDs.
Abbreviations
AS, atherosclerosis; CVDs, cardiovascular disease; MI, myocardial infarction; IHD, ischemic heart disease; ECs, endothelial cells; VEC, vascular endothelial cell; RBCs, red blood cells; VEGF, vascular endothelial growth factor; PDGF, platelet derived growth factor; IFN-γ, interferon gamma; IL, interleukin; MCP-1, monocyte chemotactic protein-1; PECAM-1, platelet endothelial cell adhesion molecule; ICAM-1, intercellular adhesion molecule; VCAM, vascular cell adhesion molecule; KLF, Kruppel like factor; NF-κB, nuclear factor kappa B; eNOS, endothelial nitric oxide synthase; GM-CSF, granulocyte-macrophage colony-stimulating factor; ROX, reactive oxygen species; HUVECs, human umbilical vein endothelial cells.
Author Contributions
All authors made a significant contribution to the work reported, right from conception, study design, execution, acquisition of data, analysis and interpretation, and took part in drafting, revising and critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Acknowledgments
Great appreciation to National Natural Science Foundation of China, Collaborative Innovation Center for Prevention and Treatment of Cardiovascular Disease of Sichuan Province, and Luzhou Municipal People’s Government-Southwestern Medical University Science and Technology Strategic Coopration Project for supporting this work, and special regards to all the teams and staff of the Department of cardiovascular surgery and Clinical Research Center of Southwest Medical University for raising the discussions and exciting research environment for this paper.
Disclosure
The authors report no conflicts of interest in this work.
Additional information
Funding
References
- Libby P. Inflammation in atherosclerosis. Arterioscler Thromb Vasc Biol. 2012;32(9):2045–2051. doi:10.1161/ATVBAHA.108.179705
- Pourcet B, Staels B. Alternative macrophages in atherosclerosis: not always protective! J Clin Invest. 2018;128(3):910–912. doi:10.1172/JCI120123
- Ross R. Atherosclerosis--an inflammatory disease. N Engl J Med. 1999;340(2):115–126. doi:10.1056/NEJM199901143400207
- Ruscica M, Corsini A, Ferri N, Banach M, Sirtori CR. Clinical approach to the inflammatory etiology of cardiovascular diseases. Pharmacol Res. 2020;159:104916.
- O’Morain VL, Ramji DP. The potential of probiotics in the prevention and treatment of atherosclerosis. Mol Nutr Food Res. 2020;64(4):e1900797. doi:10.1002/mnfr.201900797
- Song P, Fang Z, Wang H, et al. Global and regional prevalence, burden, and risk factors for carotid atherosclerosis: a systematic review, meta-analysis, and modelling study. Lancet Global Health. 2020;8(5):e721–e729. doi:10.1016/S2214-109X(20)30117-0
- Zhou J, Li YS, Chien S. Shear stress-initiated signaling and its regulation of endothelial function. Arterioscler Thromb Vasc Biol. 2014;34(10):2191–2198. doi:10.1161/ATVBAHA.114.303422
- Albarran-Juarez J, Iring A, Wang S, et al. Piezo1 and Gq/G11 promote endothelial inflammation depending on flow pattern and integrin activation. J Exp Med. 2018;215(10):2655–2672. doi:10.1084/jem.20180483
- Beech DJ, Kalli AC. Force sensing by piezo channels in cardiovascular health and disease. Arterioscler Thromb Vasc Biol. 2019;39(11):2228–2239. doi:10.1161/ATVBAHA.119.313348
- Lukacs V, Mathur J, Mao R, et al. Impaired PIEZO1 function in patients with a novel autosomal recessive congenital lymphatic dysplasia. Nat Commun. 2015;6:8329. doi:10.1038/ncomms9329
- Evans EL, Cuthbertson K, Endesh N, et al. Yoda1 analogue (Dooku1) which antagonizes Yoda1-evoked activation of Piezo1 and aortic relaxation. Br J Pharmacol. 2018;175(10):1744–1759. doi:10.1111/bph.14188
- Bagriantsev SN, Gracheva EO, Gallagher PG. Piezo proteins: regulators of mechanosensation and other cellular processes. J Biol Chem. 2014;289(46):31673–31681. doi:10.1074/jbc.R114.612697
- Coste B, Murthy SE, Mathur J, et al. Piezo1 ion channel pore properties are dictated by C-terminal region. Nat Commun. 2015;6:7223. doi:10.1038/ncomms8223
- Lewis AH, Grandl J. Mechanical sensitivity of Piezo1 ion channels can be tuned by cellular membrane tension. Elife. 2015;4:e12088. doi:10.7554/eLife.12088
- Han Y, Liu C, Zhang D, et al. Mechanosensitive ion channel Piezo1 promotes prostate cancer development through the activation of the Akt/mTOR pathway and acceleration of cell cycle. Int J Oncol. 2019;55(3):629–644. doi:10.3892/ijo.2019.4839
- Geng J, Zhao Q, Zhang T, Xiao B. In touch with the mechanosensitive piezo channels: structure, ion permeation, and mechanotransduction. Curr Top Membr. 2017;79:159–195.
- Hyman AJ, Tumova S, Beech DJ. Piezo1 channels in vascular development and the sensing of shear stress. Curr Top Membr. 2017;79:37–57.
- Li J, Hou B, Beech DJ. Endothelial Piezo1: life depends on it. Channels (Austin). 2015;9(1):1–2. doi:10.4161/19336950.2014.986623
- Ridone P, Vassalli M, Martinac B. Piezo1 mechanosensitive channels: what are they and why are they important. Biophys Rev. 2019;11(5):795–805. doi:10.1007/s12551-019-00584-5
- Retailleau K, Duprat F, Arhatte M, et al. Piezo1 in smooth muscle cells is involved in hypertension-dependent arterial remodeling. Cell Rep. 2015;13(6):1161–1171. doi:10.1016/j.celrep.2015.09.072
- Wang S, Chennupati R, Kaur H, Iring A, Wettschureck N, Offermanns S. Endothelial cation channel PIEZO1 controls blood pressure by mediating flow-induced ATP release. J Clin Invest. 2016;126(12):4527–4536. doi:10.1172/JCI87343
- Burke SD, Jordan J, Harrison DG, Karumanchi SA. Solving baroreceptor mystery: role of PIEZO ion channels. J Am Soc Nephrol. 2019;30(6):911–913. doi:10.1681/ASN.2019020160
- Alper SL. Genetic diseases of PIEZO1 and PIEZO2 dysfunction. Curr Topics Membranes. 2017;79:97–134. doi:10.1016/bs.ctm.2017.01.001
- Martin-Almedina S, Mansour S, Ostergaard P. Human phenotypes caused by PIEZO1 mutations; one gene, two overlapping phenotypes? J Physiol. 2018;596(6):985–992. doi:10.1113/JP275718
- Zarychanski R, Schulz VP, Houston BL, et al. Mutations in the mechanotransduction protein PIEZO1 are associated with hereditary xerocytosis. Blood. 2012;120(9):1908–1915. doi:10.1182/blood-2012-04-422253
- Xiao B. Levering mechanically activated piezo channels for potential pharmacological intervention. Annu Rev Pharmacol Toxicol. 2020;60:195–218. doi:10.1146/annurev-pharmtox-010919-023703
- Fang XZ, Zhou T, Xu JQ, et al. Structure, kinetic properties and biological function of mechanosensitive Piezo channels. Cell Biosci. 2021;11(1):13. doi:10.1186/s13578-020-00522-z
- De Felice D, Alaimo A. Mechanosensitive piezo channels in cancer: focus on altered calcium signaling in cancer cells and in tumor progression. Cancers (Basel). 2020;12(7):1780. doi:10.3390/cancers12071780
- Chang JE, Buechler MB, Gressier E, Turley SJ, Carroll MC. Mechanosensing by Peyer’s patch stroma regulates lymphocyte migration and mucosal antibody responses. Nat Immunol. 2019;20(11):1506–1516. doi:10.1038/s41590-019-0505-z
- Song J, Liu L, Lv L, et al. Fluid shear stress induces Runx-2 expression via upregulation of PIEZO1 in MC3T3-E1 cells. Cell Biol Int. 2020;44(7):1491–1502. doi:10.1002/cbin.11344
- Sun Y, Li M, Liu G, et al. The function of Piezo1 in colon cancer metastasis and its potential regulatory mechanism. J Cancer Res Clin Oncol. 2020;146(5):1139–1152. doi:10.1007/s00432-020-03179-w
- Velasco-Estevez M, Gadalla KKE, Linan-Barba N, Cobb S, Dev KK, Sheridan GK. Inhibition of Piezo1 attenuates demyelination in the central nervous system. Glia. 2020;68(2):356–375. doi:10.1002/glia.23722
- Zhong M, Komarova Y, Rehman J, Malik AB. Mechanosensing Piezo channels in tissue homeostasis including their role in lungs. Pulm Circ. 2018;8(2):2045894018767393. doi:10.1177/2045894018767393
- Kang H, Hong Z, Zhong M, et al. Piezo1 mediates angiogenesis through activation of MT1-MMP signaling. Am J Physiol Cell Physiol. 2019;316(1):C92–C103. doi:10.1152/ajpcell.00346.2018
- Rode B, Shi J, Endesh N, et al. Piezo1 channels sense whole body physical activity to reset cardiovascular homeostasis and enhance performance. Nat Commun. 2017;8(1):1–11 doi:10.1038/s41467-017-00429-3.
- Lim CG, Jang J, Kim C. Cellular machinery for sensing mechanical force. BMB Rep. 2018;51(12):623–629. doi:10.5483/BMBRep.2018.51.12.237
- Douguet D, Patel A, Xu A, Vanhoutte PM, Honore E. Piezo ion channels in cardiovascular mechanobiology. Trends Pharmacol Sci. 2019;40(12):956–970. doi:10.1016/j.tips.2019.10.002
- Zhao Q, Zhou H, Chi S, et al. Structure and mechanogating mechanism of the Piezo1 channel. Nature. 2018;554(7693):487–492. doi:10.1038/nature25743
- Jiang Y, Yang X, Jiang J, Xiao B. Structural designs and mechanogating mechanisms of the mechanosensitive piezo channels. Trends Biochem Sci. 2021;46(6):472–488. doi:10.1016/j.tibs.2021.01.008
- Zheng W, Gracheva EO, Bagriantsev SN. A hydrophobic gate in the inner pore helix is the major determinant of inactivation in mechanosensitive Piezo channels. Elife. 2019;8:e44003. doi:10.7554/eLife.44003
- Liang X, Howard J. Structural biology: piezo senses tension through curvature. Curr Biol. 2018;28(8):R357–R359. doi:10.1016/j.cub.2018.02.078
- Kamajaya A, Kaiser JT, Lee J, Reid M, Rees DC. The structure of a conserved piezo channel domain reveals a topologically distinct beta sandwich fold. Structure. 2014;22(10):1520–1527. doi:10.1016/j.str.2014.08.009
- Wu J, Lewis AH, Grandl J. Touch, tension, and transduction - the function and regulation of piezo ion channels. Trends Biochem Sci. 2017;42(1):57–71. doi:10.1016/j.tibs.2016.09.004
- Gottlieb PA. A tour de force: the discovery, properties, and function of piezo channels. Curr Top Membr. 2017;79:1–36.
- Li J, Hou B, Tumova S, et al. Piezo1 integration of vascular architecture with physiological force. Nature. 2014;515(7526):279–282. doi:10.1038/nature13701
- Ranade SS, Qiu Z, Woo SH, et al. Piezo1, a mechanically activated ion channel, is required for vascular development in mice. Proc Natl Acad Sci U S A. 2014;111(28):10347–10352. doi:10.1073/pnas.1409233111
- Coste B, Mathur J, Schmidt M, et al. Piezo1 and Piezo2 are essential components of distinct mechanically activated cation channels. Science. 2010;330(6000):55–60. doi:10.1126/science.1193270
- Liu X, Sun A, Fan Y, Deng X. Physiological significance of helical flow in the arterial system and its potential clinical applications. Ann Biomed Eng. 2015;43(1):3–15. doi:10.1007/s10439-014-1097-2
- Baratchi S, Chen YC, Peter K. Helical flow: a means to identify unstable plaques and a new direction for the design of vascular grafts and stents. Atherosclerosis. 2020;300:34–36. doi:10.1016/j.atherosclerosis.2020.03.002
- De Nisco G, Hoogendoorn A, Chiastra C, et al. The impact of helical flow on coronary atherosclerotic plaque development. Atherosclerosis. 2020;300:39–46. doi:10.1016/j.atherosclerosis.2020.01.027
- Zhang L, Yu J, Wei W. Advance in targeted immunotherapy for graft-versus-host disease. Front Immunol. 2018;9:1087. doi:10.3389/fimmu.2018.01087
- Chen Z, Fan Y, Deng X, Xu Z. Swirling flow can suppress flow disturbances in endovascular stents: a numerical study. ASAIO J. 2009;55(6):543–549. doi:10.1097/MAT.0b013e3181b78e46
- Beech DJ. Endothelial Piezo1 channels as sensors of exercise. J Physiol. 2018;596(6):979–984. doi:10.1113/JP274396
- Chatzizisis YS, Giannoglou GD, Parcharidis GE, Louridas GE. Is left coronary system more susceptible to atherosclerosis than right? A pathophysiological insight. Int J Cardiol. 2007;116(1):7–13. doi:10.1016/j.ijcard.2006.03.029
- Blythe NM, Muraki K, Ludlow MJ, et al. Mechanically activated Piezo1 channels of cardiac fibroblasts stimulate p38 mitogen-activated protein kinase activity and interleukin-6 secretion. J Biol Chem. 2019;294(46):17395–17408. doi:10.1074/jbc.RA119.009167
- Ando J, Yamamoto K. Flow detection and calcium signalling in vascular endothelial cells. Cardiovasc Res. 2013;99(2):260–268. doi:10.1093/cvr/cvt084
- Beech DJ, Xiao B. Piezo channel mechanisms in health and disease. J Physiol. 2018;596(6):965–967. doi:10.1113/JP274395
- Fels B, Kusche-Vihrog K. It takes more than two to tango: mechanosignaling of the endothelial surface. Pflugers Arch. 2020;472(4):419–433. doi:10.1007/s00424-020-02369-2
- Brown BM, Nguyen HM, Wulff H. Recent advances in our understanding of the structure and function of more unusual cation channels. F1000Res. 2019;8:123. doi:10.12688/f1000research.17163.1
- Nagel T, Resnick N, Dewey CF, Gimbrone MA. Vascular endothelial cells respond to spatial gradients in fluid shear stress by enhanced activation of transcription factors. Arterioscler Thromb Vasc Biol. 1999;19(8):1825–1834. doi:10.1161/01.ATV.19.8.1825
- Nigro P, Abe J, Berk BC. Flow shear stress and atherosclerosis: a matter of site specificity. Antioxid Redox Signal. 2011;15(5):1405–1414. doi:10.1089/ars.2010.3679
- Hansson GK, Libby P, Schonbeck U, Yan ZQ. Innate and adaptive immunity in the pathogenesis of atherosclerosis. Circ Res. 2002;91(4):281–291. doi:10.1161/01.RES.0000029784.15893.10
- Libby P, Hansson GK. Inflammation and immunity in diseases of the arterial tree: players and layers. Circ Res. 2015;116(2):307–311. doi:10.1161/CIRCRESAHA.116.301313
- Libby P, Loscalzo J, Ridker PM, et al. Inflammation, immunity, and infection in atherothrombosis: JACC review topic of the week. J Am Coll Cardiol. 2018;72(17):2071–2081. doi:10.1016/j.jacc.2018.08.1043
- Solis AG, Bielecki P, Steach HR, et al. Mechanosensation of cyclical force by PIEZO1 is essential for innate immunity. Nature. 2019;573(7772):69–74. doi:10.1038/s41586-019-1485-8
- Walmsley SR. Pressure regulate immune cell function. Nature. 2019;573(7772):41–42. doi:10.1038/d41586-019-02339-4
- Williams ER. PIEZO1 promotes inflammation. Sci Signal. 2019;12(598):eaaz4154.
- Liu CSC, Raychaudhuri D, Paul B, et al. Cutting edge: piezo1 mechanosensors optimize human T cell activation. J Immunol. 2018;200(4):1255–1260. doi:10.4049/jimmunol.1701118
- Bentzon JF, Otsuka F, Virmani R, Falk E. Mechanisms of plaque formation and rupture. Circ Res. 2014;114(12):1852–1866. doi:10.1161/CIRCRESAHA.114.302721
- Libby P, Ridker PM, Maseri A. Inflammation and atherosclerosis. Circulation. 2002;105(9):1135–1143. doi:10.1161/hc0902.104353
- Chatterjee S. Endothelial mechanotransduction, redox signaling and the regulation of vascular inflammatory pathways. Front Physiol. 2018;9:524. doi:10.3389/fphys.2018.00524
- Mantella LE, Quan A, Verma S. Variability in vascular smooth muscle cell stretch-induced responses in 2D culture. Vasc Cell. 2015;7:7. doi:10.1186/s13221-015-0032-0
- Jufri NF, Mohamedali A, Avolio A, Baker MS. Mechanical stretch: physiological and pathological implications for human vascular endothelial cells. Vasc Cell. 2015;7:8. doi:10.1186/s13221-015-0033-z
- Basatemur GL, Jorgensen HF, Clarke MCH, Bennett MR, Mallat Z. Vascular smooth muscle cells in atherosclerosis. Nat Rev Cardiol. 2019;16(12):727–744. doi:10.1038/s41569-019-0227-9
- Chen Y, Waqar AB, Nishijima K, et al. Macrophage-derived MMP-9 enhances the progression of atherosclerotic lesions and vascular calcification in transgenic rabbits. J Cell Mol Med. 2020;24(7):4261–4274. doi:10.1111/jcmm.15087
- Lichtman AH, Binder CJ, Tsimikas S, Witztum JL. Adaptive immunity in atherogenesis: new insights and therapeutic approaches. J Clin Invest. 2013;123(1):27–36. doi:10.1172/JCI63108
- Heo KS, Berk BC, Abe J. Disturbed flow-induced endothelial proatherogenic signaling via regulating post-translational modifications and epigenetic events. Antioxid Redox Signal. 2016;25(7):435–450. doi:10.1089/ars.2015.6556
- Chistiakov DA, Orekhov AN, Bobryshev YV. Effects of shear stress on endothelial cells: go with the flow. Acta Physiologica. 2017;219(2):382–408. doi:10.1111/apha.12725
- Chiu JJ, Chien S. Effects of disturbed flow on vascular endothelium: pathophysiological basis and clinical perspectives. Physiol Rev. 2011;91(1):327–387. doi:10.1152/physrev.00047.2009
- Tovar-Lopez F, Thurgood P, Gilliam C, et al. A microfluidic system for studying the effects of disturbed flow on endothelial cells. Front Bioeng Biotechnol. 2019;7:81. Doi:10.3389/fbioe.2019.00081
- Baratchi S, Khoshmanesh K, Woodman OL, Potocnik S, Peter K, McIntyre P. Molecular sensors of blood flow in endothelial cells. Trends Mol Med. 2017;23(9):850–868. doi:10.1016/j.molmed.2017.07.007
- Lee DY, Chiu JJ. Atherosclerosis and flow: roles of epigenetic modulation in vascular endothelium. J Biomed Sci. 2019;26(1):56. doi:10.1186/s12929-019-0551-8
- Wang S, Iring A, Strilic B, et al. P2Y(2) and Gq/G(1)(1) control blood pressure by mediating endothelial mechanotransduction. J Clin Invest. 2015;125(8):3077–3086. doi:10.1172/JCI81067
- Tanaka K, Joshi D, Timalsina S, Schwartz MA. Early events in endothelial flow sensing. Cytoskeleton. 2021. doi:10.1002/cm.21652
- Nakayama A, Albarran-Juarez J, Liang G, et al. Disturbed flow-induced Gs-mediated signaling protects against endothelial inflammation and atherosclerosis. JCI Insight. 2020;5(23). doi:10.1172/jci.insight.140485.
- Givens C, Tzima E. Endothelial mechanosignaling: does one sensor fit all? Antioxid Redox Signal. 2016;25(7):373–388. doi:10.1089/ars.2015.6493
- Song L, Leung C, Schindler C. Lymphocytes are important in early atherosclerosis. J Clin Invest. 2001;108(2):251–259. doi:10.1172/JCI200111380
- Hansson GK. Inflammation, atherosclerosis, and coronary artery disease. N Engl J Med. 2005;352(16):1685–1695. doi:10.1056/NEJMra043430
- Pageon SV, Govendir MA, Kempe D, Biro M. Mechanoimmunology: molecular-scale forces govern immune cell functions. Mol Biol Cell. 2018;29(16):1919–1926. doi:10.1091/mbc.E18-02-0120
- Liu CSC, Ganguly D. Mechanical cues for T cell activation: role of piezo1 mechanosensors. Crit Rev Immunol. 2019;39(1):15–38. doi:10.1615/CritRevImmunol.2019029595
- Gistera A, Robertson AK, Andersson J, et al. Transforming growth factor-beta signaling in T cells promotes stabilization of atherosclerotic plaques through an interleukin-17-dependent pathway. Sci Transl Med. 2013;5(196):196ra100. doi:10.1126/scitranslmed.3006133
- Jin Y, Fu J. Novel insights into the NLRP 3 inflammasome in atherosclerosis. J Am Heart Assoc. 2019;8(12):e012219. doi:10.1161/JAHA.119.012219
- Chen X, Guo X, Ge Q, Zhao Y, Mu H, Zhang J. ER stress activates the NLRP3 inflammasome: a novel mechanism of atherosclerosis. Oxid Med Cell Longev. 2019;2019:1–18.
- Ingber DE. From mechanobiology to developmentally inspired engineering. Philos Trans R Soc Lond B Biol Sci. 2018;373(1759):20170323. doi:10.1098/rstb.2017.0323
- Faucherre A, Moha Ou Maati H, Nasr N, et al. Piezo1 is required for outflow tract and aortic valve development. J Mol Cell Cardiol. 2020;143:51–62. doi:10.1016/j.yjmcc.2020.03.013
- Asmussen A, Fink K, Busch H-J, et al. Inflammasome and toll-like receptor signaling in human monocytes after successful cardiopulmonary resuscitation. Critical Care. 2016;20(1):1–5. doi:10.1186/s13054-016-1340-3.
- Arbore G, Kemper C. A novel “complement-metabolism-inflammasome axis” as a key regulator of immune cell effector function. Eur J Immunol. 2016;46(7):1563–1573. doi:10.1002/eji.201546131
- Hoseini Z, Sepahvand F, Rashidi B, Sahebkar A, Masoudifar A, Mirzaei H. NLRP3 inflammasome: its regulation and involvement in atherosclerosis. J Cell Physiol. 2018;233(3):2116–2132. doi:10.1002/jcp.25930
- Luo H, Liu H, Bian W, Chen B, Yang D, Yang M. Yoda1 activates piezo1 in vitro to simulate the upregulation of piezo1 in the infected brain: piezo1 participates in the immune activation of microglia. 2021.
- Koushki K, Shahbaz SK, Mashayekhi K, et al. Anti-inflammatory action of statins in cardiovascular disease: the role of inflammasome and toll-like receptor pathways. Clin Rev Allergy Immunol. 2021;60(2):175–199.
- Latz E, Duewell P. NLRP3 inflammasome activation in inflammaging. Semin Immunol. 2018;40:61–73.
- Liu Y, Dai Y, Li Q, et al. Beta-amyloid activates NLRP3 inflammasome via TLR4 in mouse microglia. Neurosci Lett. 2020;736:135279. doi:10.1016/j.neulet.2020.135279
- Wu J, Goyal R, Grandl J. Localized force application reveals mechanically sensitive domains of Piezo1. Nat Commun. 2016;7:12939. doi:10.1038/ncomms12939
- Chen X, Wanggou S, Bodalia A, et al. A feedforward mechanism mediated by mechanosensitive ion channel PIEZO1 and tissue mechanics promotes glioma aggression. Neuron. 2018;100(4):799–815 e797. doi:10.1016/j.neuron.2018.09.046
- Maruyama K, Nemoto E, Yamada S. Mechanical regulation of macrophage function - cyclic tensile force inhibits NLRP3 inflammasome-dependent IL-1beta secretion in murine macrophages. Inflamm Regen. 2019;39:3. doi:10.1186/s41232-019-0092-2
- Sun Y, Leng P, Song M, et al. Piezo1 activates the NLRP3 inflammasome in nucleus pulposus cell-mediated by Ca(2+)/NF-kappaB pathway. Int Immunopharmacol. 2020;85:106681. doi:10.1016/j.intimp.2020.106681
- Wu J, Yan Z, Schwartz DE, Yu J, Malik AB, Hu G. Activation of NLRP3 inflammasome in alveolar macrophages contributes to mechanical stretch-induced lung inflammation and injury. J Immunol. 2013;190(7):3590–3599. doi:10.4049/jimmunol.1200860
- Stojadinovic O, Minkiewicz J, Sawaya A, et al. Deep tissue injury in development of pressure ulcers: a decrease of inflammasome activation and changes in human skin morphology in response to aging and mechanical load. PLoS One. 2013;8(8):e69223. doi:10.1371/journal.pone.0069223
- Maruyama K, Sakisaka Y, Suto M, et al. Cyclic stretch negatively regulates IL-1beta secretion through the inhibition of NLRP3 inflammasome activation by attenuating the AMP kinase pathway. Front Physiol. 2018;9:802. doi:10.3389/fphys.2018.00802
- Hu N, Zhang Y. TLR4 knockout attenuated high fat diet-induced cardiac dysfunction via NF-kappaB/JNK-dependent activation of autophagy. Biochim Biophys Acta Mol Basis Dis. 2017;1863(8):2001–2011. doi:10.1016/j.bbadis.2017.01.010
- Lacroix JJ, Botello-Smith WM, Luo Y. Probing the gating mechanism of the mechanosensitive channel Piezo1 with the small molecule Yoda1. Nat Commun. 2018;9(1):2029. doi:10.1038/s41467-018-04405-3
- Botello-Smith WM, Jiang W, Zhang H, et al. A mechanism for the activation of the mechanosensitive Piezo1 channel by the small molecule Yoda1. Nat Commun. 2019;10(1):4503. doi:10.1038/s41467-019-12501-1
- Goldblatt ZE, Cirka HA, Billiar KL. Mechanical regulation of apoptosis in the cardiovascular system. Ann Biomed Eng. 2021;49(1):75–97. doi:10.1007/s10439-020-02659-x
- Chan DD, Van Dyke WS, Bahls M, et al. Mechanostasis in apoptosis and medicine. Prog Biophys Mol Biol. 2011;106(3):517–524. doi:10.1016/j.pbiomolbio.2011.08.002
- Muluh TA, Chen Z, Li Y, et al. Enhancing cancer immunotherapy treatment goals by using nanoparticle delivery system. Int J Nanomedicine. 2021;16:2389–2404. doi:10.2147/IJN.S295300
- Gnanasambandam R, Bae C, Gottlieb PA, Sachs F. Ionic selectivity and permeation properties of human PIEZO1 channels. PLoS One. 2015;10(5):e0125503. doi:10.1371/journal.pone.0125503
- Mendoza MC, Er EE, Blenis J. The Ras-ERK and PI3K-mTOR pathways: cross-talk and compensation. Trends Biochem Sci. 2011;36(6):320–328. doi:10.1016/j.tibs.2011.03.006
- Saxton RA, Sabatini DM. mTOR signaling in growth, metabolism, and disease. Cell. 2017;168(6):960–976. doi:10.1016/j.cell.2017.02.004
- Aminzadeh A, Salarinejad A. Effects of myricetin against cadmium-induced neurotoxicity in PC12 cells. Toxicol Res (Camb). 2021;10(1):84–90. doi:10.1093/toxres/tfaa104
- Zhang R, Zhang N, Zhang H, et al. Celastrol prevents cadmium-induced neuronal cell death by blocking reactive oxygen species-mediated mammalian target of rapamycin pathway. Br J Pharmacol. 2017;174(1):82–100. doi:10.1111/bph.13655
- Syeda R, Xu J, Dubin AE, et al. Chemical activation of the mechanotransduction channel Piezo1. Elife. 2015;4:e07369. Doi:10.7554/eLife.07369
- Wang Y, Chi S, Guo H, et al. A lever-like transduction pathway for long-distance chemical- and mechano-gating of the mechanosensitive Piezo1 channel. Nat Commun. 2018;9(1):1300. doi:10.1038/s41467-018-03570-9
- Bae C, Sachs F, Gottlieb PA. The mechanosensitive ion channel Piezo1 is inhibited by the peptide GsMTx4. Biochemistry. 2011;50(29):6295–6300. doi:10.1021/bi200770q
- Beqja D, Haidar S, Nikolaev M, Shen Y, Denholm B. Transgenic tarantula toxin: a novel tool to study mechanosensitive ion channels in Drosophila. J Insect Physiol. 2020;127:104116. doi:10.1016/j.jinsphys.2020.104116
- Gnanasambandam R, Ghatak C, Yasmann A, et al. GsMTx4: mechanism of inhibiting mechanosensitive ion channels. Biophys J. 2017;112(1):31–45. doi:10.1016/j.bpj.2016.11.013
- Suchyna TM. Piezo channels and GsMTx4: two milestones in our understanding of excitatory mechanosensitive channels and their role in pathology. Prog Biophys Mol Biol. 2017;130(Pt B):244–253. doi:10.1016/j.pbiomolbio.2017.07.011
- Coste B, Xiao B, Santos JS, et al. Piezo proteins are pore-forming subunits of mechanically activated channels. Nature. 2012;483(7388):176–181. doi:10.1038/nature10812
- Liu S, Pan X, Cheng W, et al. Tubeimoside I antagonizes yoda1-evoked piezo1 channel activation. Front Pharmacol. 2020;11:768. doi:10.3389/fphar.2020.00768
- Zhao Y, Ren P, Li Q, et al. Low shear stress upregulates CX3CR1 expression by inducing VCAM-1 via the NF-kappaB pathway in vascular endothelial cells. Cell Biochem Biophys. 2020;78(3):383–389. doi:10.1007/s12013-020-00931-4
- Fan Y, Xu Z, Jiang W, Deng X, Wang K, Sun A. An S-type bypass can improve the hemodynamics in the bypassed arteries and suppress intimal hyperplasia along the host artery floor. J Biomech. 2008;41(11):2498–2505. doi:10.1016/j.jbiomech.2008.05.008
- Caro CG, Seneviratne A, Heraty KB, et al. Intimal hyperplasia following implantation of helical-centreline and straight-centreline stents in common carotid arteries in healthy pigs: influence of intraluminal flow. J R Soc Interface. 2013;10(89):20130578. doi:10.1098/rsif.2013.0578
- Zhan F, Fan Y, Deng X. Swirling flow created in a glass tube suppressed platelet adhesion to the surface of the tube: its implication in the design of small-caliber arterial grafts. Thromb Res. 2010;125(5):413–418. doi:10.1016/j.thromres.2009.02.011
- Doty DB, Flores JH, Doty JR, Millar RC. Mitral valve replacement with homograft. Semin Thorac Cardiovasc Surg. 1999;11(4 Suppl 1):191–193.
- Chen Z, Zhan F, Fan Y, Deng X. A novel way to reduce thrombus build-up in vena cava filters. Catheter Cardiovasc Interv. 2011;78(5):792–798. doi:10.1002/ccd.23107