
Abstract
Aim
Disruption of the blood–brain barrier (BBB) is a critical pathological feature after stroke. Although tissue kallikrein (TK) has used in the treatment of stroke in China, the role of TK in modulating BBB permeability is not clear.
Methods
We investigated the effect of different doses of TK on BBB by in vivo assessments of Evans blue (EB) and sodium-fluorescein isothiocyanate (FITC) leakage and in vitro assessments of the integrity of BBB and monolayers of microvascular endothelial cells (BMVECs). The expression of zonula occludens-1 (ZO-1) and bradykinin receptor-mediated signaling in BMVECs was detected.
Results
A significant increase in BBB permeability was observed in the mice treated with high dose of TK. However, standard and medium doses of TK could only enable sodium-FITC to enter the brain. The result of in vitro study indicated that high-doses of TK, but not standard and medium-dose of TK, reduced normal BBB integrity accompanied by a decreased expression of zonula occludens-1 (ZO-1), upregulated the mRNA levels of bradykinin 2 receptor (B2R) and endothelial nitric oxide synthase (eNOS) and the abundance of B2R. Moreover, standard-dose of TK exacerbated lipopolysaccharide-induced BBB hyperpermeability, upregulated the mRNA levels of bradykinin 1 receptor (B1R) and inducible nitric oxide synthase (iNOS), increased the abundance of B1R and reduced the abundance of ZO-1; these effects were inhibited by TK inhibitor.
Conclusion
TK can disrupt tight junctions and increase normal BBB permeability via B2R-dependent eNOS signaling pathway, aggravate impairment of BBB via B1R-dependent iNOS signaling pathway, and consequently serve as a useful adjunctive treatment for enhancing the efficacy of other neurotherapeutics.
Graphical Abstract
Introduction
Human urinary kallidinogenase (HUK) is tissue kallikrein (TK) extracted from human urine and used in the treatment of cerebral stroke in China.Citation1 TK, a key enzyme in the tissue kallikrein-kinin system (KKS), cleaves low molecular kininogen to release kinin and bradykinin. Bradykinin acts on bradykinin 1 receptor (B1R) and bradykinin 2 receptor (B2R) to release nitric oxide (NO) and thus, trigger a wide range of biological effects, including lowering blood pressure, inhibiting oxidative stress, and increasing vascular permeability.Citation2,Citation3 Previously, our clinical investigation indicated that reduced TK level was associated with first-ever stroke and predicted higher stroke recurrences and shorter event-free survival time.Citation4 Numerous basic and clinical studies also suggest that TK improves the long-term prognosis of cerebral stroke through bradykinin released by KKS.Citation5–Citation7 However, disruption of the blood–brain barrier (BBB) plays an important role in the development of neurological dysfunction in stroke.Citation8 The role of TK in regulating BBB permeability is yet unknown. We, therefore, suppose that TK modulates BBB permeability via promoting bradykinin formation.
Unlike high doses of BK, which might open the whole brain barrier, low doses of BK can selectively increase the BBB permeability of brain tumors through B2R without affecting the normal brain.Citation9,Citation10 Typically, B2Rs are constitutively expressed and mediate endothelial nitric oxide synthase (eNOS) activation, which leads to a transient output of NO in endothelial cells, whereas B1R/inducible nitric oxide synthase (iNOS) activation is induced in lipopolysaccharide (LPS)-treated endothelial cells and leads to very high and prolonged (~90 min) NO production.Citation11 Under pathological conditions such as inflammation, ischemia-reperfusion (I/R), and trauma, BK was thought to mediate the increase in BBB permeability.Citation12–Citation14 Moreover, inhibition of B1R was proved to mitigate the destruction of BBB in central system diseases, reduce BBB permeability and relieve cerebral edema.Citation15,Citation16 However, whether TK, as a key enzyme regulating the release of BK, disrupts BBB integrity, and if so, whether its role is mediated by B2R/eNOS or B1R/iNOS signaling remains unclear.
Investigating the role and underlying molecular mechanisms of TK in regulating BBB permeability is substantially significant in the treatment of neurological disease. Herein, a BBB model that mimics the in vivo environment, was established by co-cultured murine brain microvascular endothelial cells (BMECs) with astrocytes in a transwell system. Then we investigated the potential role of TK on BBB permeability by combining the in vivo assessments of dye extravasation and brain edema in mice and the in vitro assessments of BBB integrity and the expression of tight junction protein zonula occludens-1 (ZO-1).
Materials and Methods
TK Treatment in the in vivo Study
To investigate the effect of TK on BBB, 48 C57BL/6 male mice were divided equally into four groups: control (intravenous injection of 0.9% saline solution), standard-dose TK treatment, medium-dose TK treatment, and high-dose TK treatment. We took the TK concentration which is employed in clinical and basic studiesCitation17 as standard-dose TK treatment (intravenous injection of HUK with 25×10−3 PNA U/kg), and then increased TK concentration on multiple. Finally, the TK concentration which can increase the permeability of BBB to EB was regarded as high-dose TK treatment (intravenous injection of HUK with 400×10−3 PNA U/kg), and the intermediate concentration was used as the medium-dose TK treatment (intravenous injection of HUK with 100×10−3 PNA U/kg). All drugs were administered 24 h before the mice were treated. Animal care and experimental procedures were carried out in accordance with the guidelines provided by the Institutional Animal Care and Use Committee of Tongji Hospital, Tongji Medical College, Huazhong University of Science and Technology (Wuhan, China). This project was conducted at Tongji Hospital, Huazhong University of Science and Technology with approvals granted by the Animal Ethics Committee of Huazhong University of Science and Technology.
In vivo BBB Permeability Assay in Mice
Both Evans blue (EB) and fluorescein isothiocyanate (FITC) assay were used for the assessment of the BBB integrity.Citation18–Citation20 The mice were injected with 2% of EB or sodium-FITC (1mg/mL in saline) 200μL through the tail vein 2 hours before sacrificing them; then, the dyes were cleared from the vessels via saline infusion. Subsequently, the animals were sacrificed, and the brain tissues were removed. Then, 0.5 mL formamide was added to the brain homogenate to dissolve EB dye. After incubation at 55°C for 48 h, the absorbance of the supernatant obtained by centrifugation of the samples at 12000g for 30 min at room temperature was determined at 632nm. For mice that were injected with sodium-FITC, a total of 1mL 50mM of Tris buffer solution was added to the homogenate of brain tissue, and the supernatant was harvested by centrifugation at 3000 rpm for 30 min. We proceeded to collect the supernatant, which was then mixed with methanol (1:1) and centrifuged at 3000rpm for 30min. Finally, the content of Evans blue and sodium-FITC was calculated according to the standard curve.
Brain Water Content Detection
The brain water content was measured according to the methods described previously.Citation21 The whole excised brain was weighed immediately to determine the wet weight and then dried at 80°C for 72 h to obtain the dry weight. The formula for calculating brain water content was as follows: [(wet weight − dry weight)/wet weight] × 100%.
In vitro Model of the Blood–Brain Barrier
We established an in vitro BBB model of mouse BMECs co-cultured with astrocytes using Transwell (Corning, New York, NY, USA) co-culture system.Citation22 Astrocytes were isolated and purified by differential adherence method.Citation23 Meanwhile, we isolated and purified BMVECs using PECAM-1-coated Dynabeads (Invitrogen, Carlsbad, CA, USA).Citation24–Citation26 Then, astrocytes (1×106/mL) were added to the underside of the Transwell and incubated at 37°C and 5% CO2 for 24 hours. When the astrocytes reached 60% confluence under an inverted microscope, BMVECs (1×107/mL) were seeded on top of the Transwell at 37°C and 5% CO2. Finally, we observed the cells under an inverted microscope until the astrocytes and BMVECs reached a high-density co-culture.
Detection of BBB and BMVECs Permeability in vitro Study
We set up four groups of BBB in vitro models: Control (PBS), standard-doses of TK (0.5 mM HUK), medium-doses of TK (1 mM HUK) and high-doses of TK (HUK 2 mM). Next, a FITC-dextran (Dextran-blue-3KD and Dextran-red-40KD)Citation27 Transwell assay was used to assess the in vitro BBB permeability. Furthermore, measurements of transendothelial electrical resistance (TER) across the BMVECs were performed using the electrical cell-substrate impedance sensing system (ECIS) (Applied BioPhysics, Troy, NY, USA).Citation28,Citation29 To measure impedance, cells were grown on 8W10E+ Arrays (Applied BioPhysics, Inc., Troy, NY, USA). The arrays were treated with 10 mM L-cysteine (cat#C7352-25G; Sigma-Aldrich, St. Louis, MO, USA) followed by coating with type-I collagen (cat#. A1048301, Thermo Fisher Scientific, Waltham, MA, USA). BMVECs were seeded onto the arrays at a density of 60,000 cells/cm2 in 400μL of L-DMEM growth media. ECIS was conducted using the multiple frequency/time (MFT) option to record the impedance measurements over a broad spectrum of frequencies.
Quantitative Real-Time PCR (qPCR)
Total RNA was extracted from cells using the RNeasy Mini Kit (Qiagen, Valencia, CA, USA), according to the manufacturer’s instructions. Then, cDNA was obtained by reverse transcription kit (Invitrogen, Grand Island, NY, USA). Quantitative real-time PCR (qPCR) was conducted on 7300 Real-Time PCR System (Applied Biosystems, Foster City, CA, USA). All qPCR data of the target gene were normalized against that of 18S as an internal reference. The fold-changes in the transcripts were determined using the ΔΔCt method.
Western Blot and Immunofluorescence Staining
Total protein lysates were separated by 12% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to polyvinyl difluoride membranes (PVDF; Millipore, Bedford, MA, USA). The membrane was blocked with 5% skimmed milk at 37 °C for 1 h and probed with anti-ZO-1 (1:1000; Invitrogen), anti B1R (1:1000; MyBioSource, Inc. San Diego, CA), anti-B2R (1:500; MyBioSource, Inc. San Diego, CA) and GAPDH (1:1000; Invitrogen) antibodies for 16 hours at 4°C. Subsequently, the membranes were incubated with the corresponding goat anti-rabbit and anti-mouse secondary antibodies (1:1000) at ambient temperature for 2 h. The intensity of the immunoreactive bands was calculated by ImageJ software (NIH, Bethesda, MD, USA).
Immobilized cell samples were permeabilized with 0.1% Triton X-100 for 15 min, and the nonspecific-binding sites were blocked with PBS containing 1% bovine serum albumin BSA and 10% secondary antibody of the same origin. Then, the samples were incubated with primary antibodies, such as von Willebrand Factor (vWF, 1:500; Dako, Carpinteria, CA, USA), glial fibrillary acidic protein (GFAP, 1:50 dilution; Invitrogen), or ZO-1 (1:50; Invitrogen) overnight at 4 °C, followed by incubation with fluorescent-labeled secondary antibody (1:200; Invitrogen). Vectashield-containing 4ʹ,6-diamidino-2-phenylindole (DAPI; Vector Laboratories, Burlingame, CA, USA) was added to the slides for the detection of nuclei and sealed before capturing the images under a fluorescence microscope (Olympus, FV500-IX71, Tokyo, Japan).
Statistical Analysis
All data are shown as the mean ± standard deviation. Data were determined to be normal by the Shapiro–Wilks test. Independent-sample t-test and one-way ANOVA were employed to assess the differences among groups. A two-tailed p-value < 0.05 was considered statistically significant. Data were analyzed using SPSS version 17.0 (Armonk, NY, USA) and GraphPad Prism version 8.0.0 (San Diego, CA, USA).
Results
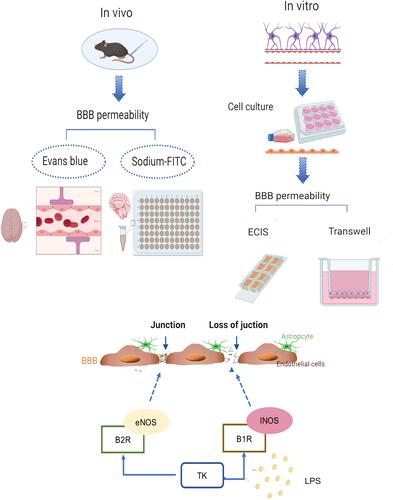
High Dose of TK Promotes BBB Permeability in Mice
To determine the effect of different doses of TK on BBB, mice were systemically injected with EB dye (macromolecular substances) or sodium-FITC dye (small molecules), and extravasation of the dyes into the brain parenchyma was evaluated. Little evidence of Evans blue extravasation was observed in the brains of control, standard-dose, and medium-dose TK treatment groups, while high-dose TK treatment visibly increased the amount of Evans blue in mouse brains (). Furthermore, both EB (147.400±8.853 μg/g of tissue vs 85.520±7.589 μg/g of tissue; P < 0.0001; ) and sodium-FITC (108.1±7.693 μg/g of tissue vs 53.910±9.344 μg/g of tissue; P < 0.0001; ) levels and water content (85.47±0.82% vs 80.33±0.55%; P < 0.0001; ) were significantly higher in the brain of mice treated with high-dose of TK than that of the control. However, EB levels and water content in the brain showed no difference among the groups of control, standard-dose and medium-dose TK treatment though sodium-FITC levels in the groups of standard-dose and medium-dose treatment were apparently increased (68.990±7.448μg/g of tissue vs 96.86±5.391 μg/g of tissue vs 53.910±9.344 μg/g of tissue; P < 0.0001; ). Collectively, these observations suggest that standard and medium-dose of TK may partially open BBB by increasing the permeability of BBB to small molecules, while high-dose of TK may disrupt the integrity of BBB and enable small molecules and large molecules to enter the brain.
Figure 1 Tissue kallikrein (TK) increases normal blood–brain barrier (BBB) permeability in mice. The mice were divided into four groups: Control (PBS); TK-S (standard dose of TK); TK-M (medium dose of TK); TK-H (high dose of TK). (A) The content of Evans blue was qualitatively analyzed by brain images. (B) The content of Evans blue (mg/g of tissue) was quantitatively detected in the supernatant of the brain homogenate. ****P<0.0001. (C) Brain water content. ****P<0.0001. (D) The content of sodium-FITC in the supernatant of brain homogenate was quantitated (mg/g of tissue). ****P<0.0001; &&&&P<0.0001, one-way analysis of variance among the four groups.
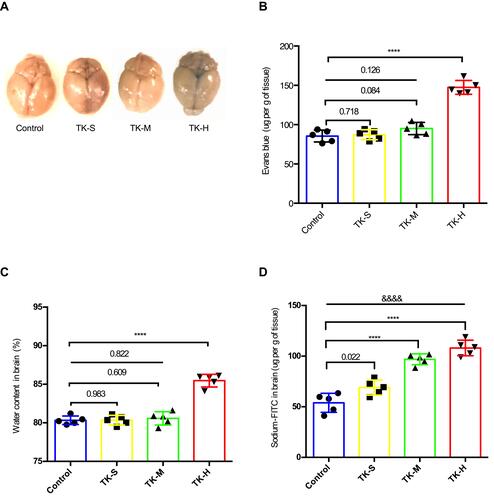
TK Regulates the Integrity and Permeability of the BBB in vitro
To more accurately mimic in vivo conditions, an in vitro model of BBB was established with cocultures of freshly isolated BMVECs and primary mouse astrocytes. The primary cultured astrocytes adhered to the wall on day 7 () and were stained red () by GFAP antibody immunofluorescence. BMVECs began to fuse between cells on day 3 after the second sorting () and were stained green using vWF antibody by immunofluorescence staining (). Then, the BBB model in vitro () was established successfully in transwell co-culture system. Data of detecting the BBB permeability present no difference in the fluorescence absorbance of dextran-blue-40KD among the groups of control, standard and medium doses of TK (), whereas the fluorescence absorbance of dextran-blue-3KD significantly increased when BBB model was treated with high-dose of TK (), suggesting high dose of TK could increase normal BBB permeability and selectively enable small molecular substances to cross the BBB. Using LPS treating BBB model in vitro,Citation30,Citation31 a significant increase of fluorescence absorbance of dextran-blue-3KD () and dextran-red-40KD () was found, indicating that LPS induces hyper permeability of BBB. Furthermore, standard-dose of TK exacerbates the LPS-induced hyperpermeability of BBB, which could be antagonized by TK-specific blocker—kallistatin ( and ). These suggest that the permeability of BBB could be regulated by TK.
Figure 2 Blood–brain barrier (BBB) model in vitro. (A) Image of astrocytes. (B) Image of endothelial cells. (C) GFAP immunofluorescence staining of astrocyte. (D) vWF immunofluorescence staining of mice BMVECs. (E) Schematic of BBB model in vitro.
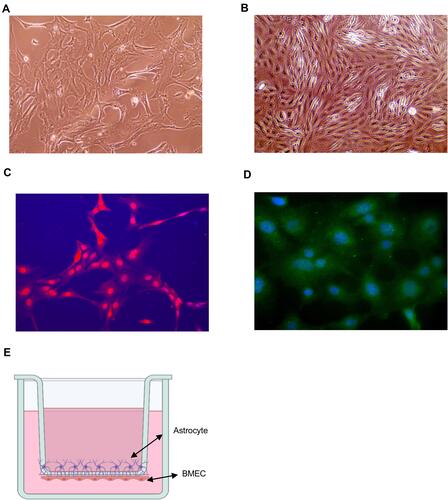
Figure 3 Effect of tissue kallikrein (TK) on blood–brain barrier (BBB) permeability in vitro. BMVECs were grown with primary mouse astrocytes in transwell chambers and treated with PBS, standard dose of TK (TK-S), medium dose of TK (TK-M), and high dose of TK (TK-H), LPS, LPS+ TK-S, and LPS+kallistatin. Then (A and C) red (40 kDa) or (B and D) blue (3 kDa) FITC-dextran was added to the chamber, and permeability was evaluated after 24 h by measuring the intensity of dextran fluorescence. **P<0.010; ***P<0.001; ****P<0.0001.
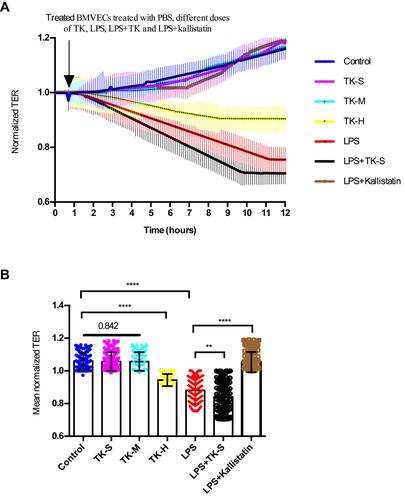
The potential role of TK in BBB permeability was also evaluated in vitro by using ECIS to measure TER in monolayer BMVECs. A slightly decreased in resistance measurements was found in BMVECs treated with high dose of TK, whereas no difference was showed between the groups with control, standard and medium doses of TK ( and ). It was consistent with the results of Transwell and indicated that TK regulated the integrity of BMVEC monolayers as resistance measurements in BMVECs declined significantly in response to LPS, and measurements were apparently lower when the cells were cotreated with standard of TK and increased significantly when LPS-treated cells were cotreated with TK inhibitor kallistatin ( and ).
Figure 4 Effect of tissue kallikrein (TK) on the integrity of brain microvascular endothelial cell (BMVEC) monolayers. Transcellular electrical resistance (TER) measurements were recorded in monolayers of BMVECs during treatment with the indicated combinations of PBS, standard dose of TK (TK-S), medium dose of TK (TK-M), high dose of TK (TK-H), LPS, LPS+ TK-S and LPS+kallistatin. (A) Real time electric cell–substrate impedance sensing (ECIS) measurements were conducted using normalized TER to record BMVECs s permeability from 3 independent experiments. The lines denote the mean normalized resistance TERs ± standard deviation of the mean (SD). (B) Mean normalized TERs were measured from 3 independent experiments using ECIS. **P<0.010; ****P<0.0001.
High Dose of TK Upregulates Normal BBB via B2R/eNOS Signal Pathway
Bradykinin is the major end-product of KKS activation, which affects the integrity of BBB by regulating the tight junction (TJ) protein ZO-1 in BMVECs;Citation32,Citation33 thus, we investigated whether TK may increase normal BBB permeability by disrupting expression of ZO-1. The protein expression of ZO-1 did not change when BMVECs were treated with standard dose of TK though the abundance of B2R and the mRNA levels of B2R and eNOS were increased (– and and ). Also, ZO-1 fluorescence was clearly distinguished at the cell borders in BMVECs in both control and standard-dose TK treatment groups (). Notably, treating BMVECs with high dose of TK caused a loss of ZO-1 and changed the immunofluorescence staining pattern of the cell boundary, increased the abundance of B2R and the mRNA levels of B2R and endothelial nitric oxide synthase (eNOS); nevertheless, the abundance of B1R and the mRNA levels of B1R and iNOS did not change (– and and –). Collectively, these suggest that high dose of TK upregulates the permeability of normal BBB by reducing the ZO-1 level via B2R/eNOS signal pathway.
Figure 5 Tissue kallikrein (TK) regulates the abundance of ZO-1 in brain microvascular endothelial cells (BMVECs). (A–H) The abundance of ZO-1, bradykinin 1 receptor (B1R) and bradykinin 2 receptor (B2R) were evaluated by Western blot in BMVECs. BMVECs had been treated with PBS, standard dose of TK (TK-S), high dose of TK (TK-H), LPS, LPS+TK-S, and LPS+kallistatin. GAPDH abundance was also evaluated to confirm equal loading, and then (B and F) ZO-1, (C and G) B1R, (D and H) B2R were quantified via normalization to that of GAPDH. ***p<0.001; ****p<0.0001. (I) Immunofluorescence staining was used to detect the expression of ZO-1 in endothelial cells. BMVECs were incubated with ZO-1 primary antibodies and fluorescent secondary antibodies, and nuclei were counterstained with DAPI.
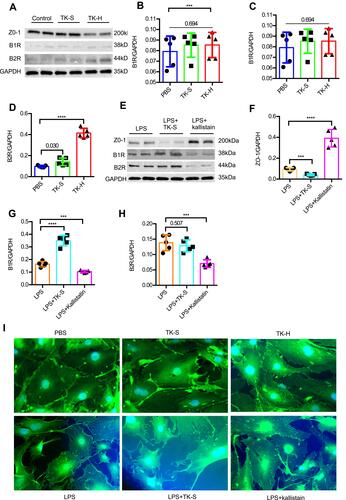
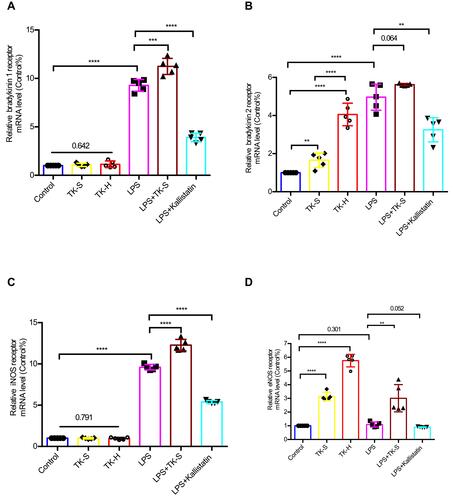
Figure 6 The mRNA levels of bradykinin 1 receptor (B1R), bradykinin 2 receptor (B2R), inducible nitric oxide synthase (iNOS), and endothelial nitric oxide synthase (eNOS) were evaluated via qPCR in brain microvascular endothelial cells (BMVECs). BMVECs were treated with PBS, TK-S, TK-H, LPS, LPS+TK-S, and LPS+kallistatin. B1R (A), B2R (B), iNOS (C) and eNOS (D) mRNA abundance were evaluated via q-PCR in BMVECs. **P<0.010; ***P<0.001; ****P<0.0001.
Disruption of BBB Integrity is Regulated by TK via B1R/iNOS Signal Pathway
Using LPS inducing the impairment of BBB, ZO-1 immunofluorescence lost its normally smooth nature at the BMVECs borders () and the mRNA levels of B1R, B2R, and iNOS in BMVECs were significantly increased, while that of eNOS is unaltered (–). Moreover, standard dose of TK appeared to strengthen LPS-induced changes in mRNA, reduce protein level of ZO-1, upregulate the abundance of B1R and destroyed the cell boundary immunofluorescence staining of ZO-1 in BMVECs (– and –). Simultaneously, kallistatin, a specific inhibitor of TK, not only reversed an increase in BBB permeability induced by LPS but also upregulated the expression of ZO-1 protein, reversed the disrupt cell boundary staining of ZO-, reduced the abundance of B1R and B2R and downregulated the mRNA of B1R and iNOS in BMVECs (– and –). Thus, LPS-induced disruption of BBB integrity was regulated by TK via B1R/iNOS signal pathway.
Discussion
To the best of our knowledge, this study presented here is the first to observe the effect of different doses of TK on the permeability of BBB. Notably, high dose of TK, but not standard and medium dose of TK, was proved to reduce BBB integrity in vivo and the integrity of BMVEC monolayers in vitro. LPS-induced hyperpermeability of BBB in vitro was exacerbated by standard-dose of TK and reversed by a specific inhibitor of TK, indicating that TK may be used to treat neurological diseases by targeting BBB permeability. Furthermore, the results from our experiments in vitro suggest that an increase of normal BBB permeability mediated by high-doses of TK may be at least partially through downregulation of ZO-1 level via B2R/eNOS signal pathway, whereas exacerbate exogenous factor-induced hyperpermeability of BBB may be regulated by TK through further disrupting of ZO-1 via B1R/iNOS signal pathway.
TK was demonstrated to play protective effects in stroke. It is implicated that opening collateral circulation improves cerebral blood perfusion and might also increase the permeability of BBB, thereby making it easy for TK to reach the ischemic injury site and protect the blood vessels of the brain.Citation5–Citation7,Citation34 Moreover, BBB impairment is a key pathological manifestation of ischemic stroke.Citation35 BK was proved to mediate the increase in BBB permeability under inflammation, I/R, and other conditions.Citation12–Citation14 Therefore, the role of TK, a key enzyme that affects BK release, in normal and impairment BBB is deserved investigating. EB dye is a macromolecular substance, while sodium-FITC dye is a small molecule. In the present study, both EB and sodium-FITC levels and water content were significantly higher than controls in the brain of mice treated with high-dose of TK, indicating that high dose of TK could increase the permeability of BBB to small molecules and macromolecules and lead to brain edema. However, only sodium-FITC level was increased in the brain of mice treated with standard and medium dose of TK, suggesting that standard and medium doses of TK could increase the permeability of BBB to small molecules without edema. We, therefore, suppose that standard and medium dose of TK selectively increased the permeability of BBB and allowed small molecules enter brain, whereas a high dose of TK increased BBB leakage with both of small and large molecules and lead brain swelling. Moreover, a high dose of TK selectively increased the permeability of BBB to small molecules but not large molecules using the in vitro model of BBB. It is suggested that an increase in the permeability of endothelial cells might be mediated by B2R under normal conditions,Citation36 whereas B2R is barely expressed in normal BMVECs.Citation37 TK was injected through the tail vein in the current study, and B2R was widely distributed in vascular endothelial cells of different organs.Citation38 Then, TK releases NO through B2R that in turn increases the permeability of BBB.Citation12 Therefore, standard and medium dose of TK can selectively open BBB without leading to brain edema, though they cannot increase the permeability of BBB in vitro. These suggest that standard and medium doses of TK may target the arterial expansion of ischemic brain tissue by increasing the permeability of mouse BBB to small molecular substances, thereby improving cerebral blood perfusion and neurological function without cerebral edema.
BMVECs are the main components of BBB which is the primary regulator of exchange between the peripheral blood and the brain. KKS is also a crucial regulatory system for endothelial cells. The expression level of ZO-1 on BMVECs is related to the permeability of BBB.Citation39 The present study of in vitro combined with other studies referring to KKSCitation40 indicates that in normal conditions, standard dose TK exerts cerebrovascular protection through B2R/eNOS signal pathway without affecting the permeability of BBB, while a sufficiently high dose of TK reduces the expression of ZO-1 through B2R/eNOS signal pathway and increases the permeability of BBB.
Downstream of TK, BK mediates biological effects via B1R and B2R, respectively. Since B1R is barely expressed in normal blood vessels, current studies propose that the protective effect of TK on cardiovascular and cerebrovascular systems is exerted via B2R,Citation3,Citation40,Citation41 while the effects of vascular injury and increased BBB might be effectuated by B1R.Citation42 In acute cerebral infarction and other ischemia and cell injury, the endothelial cells of blood vessels at the ischemic site were thought to be induced to generate B1R. Then BK bind to B1 receptor to increase the permeability and target arterial expansion of ischemic brain tissue, thereby improving the perfusion of cerebral blood and neurological function.Citation5,Citation43 Therefore, when exogenous injury leads to increased permeability of BBB, TK may release BK through B1R to increase the permeability of BBB, thus protecting the brain. This is consistent with the current results that the mRNA levels of B1R and iNOS are significantly increased when the hyperpermeability of BBB is induced by LPS. Moreover, standard and medium doses of TK further increase the mRNA levels of B1R and iNOS and reduce the protein level of ZO-1, thereby enhancing the LPS-induced hyperpermeability of BBB. However, these can be reversed by kallistatin, a specific inhibitor of TK. Therefore, we speculate that TK might be an important regulator targeting on the hyperpermeability of BBB via B1R/iNOS signal pathway.
BBB, the key surface through which systematically administered drugs access the central nervous system (CNS), is the primary regulator of exchange between the peripheral circulation and the brain.Citation44 The success of CNS therapeutic development is often dependent upon techniques for modulating BBB permeability to ensure optimal CNS penetration.Citation27 Thus, our results suggest that TK may be a useful adjunctive treatment for enhancing the efficacy of some neurotherapeutics by enabling patients to be treated with lower doses of the primary treatment.Citation27,Citation45 Moreover, hyperpermeability of BBB enables blood-borne immune cells to enter the brain and provoke a neuroinflammatory response.Citation46 It is proved to be associated with brain inflammatory conditions such as stroke, Alzheimer’s disease, and multiple sclerosis.Citation47 These suggest that modulating BBB permeability targeting on TK may have important implications for the treatment of neuroinflammatory conditions and other CNS disorders.
Nevertheless, the present study has several limitations in this study. Firstly, the effects of different doses of TK on the permeability of BBB in vivo were only limited to normal, and no in vivo study clarified whether TK could affect stroke-induced disruption of BBB integrity. An in vivo study will carry out in the future to address this issue and reveal the therapeutic effect of TK on stroke at different timing points. Secondly, we established the in vitro BBB model of mouse BMECs co-cultured with astrocytes and therefore cannot provide data on peripheral mononuclear cell infiltration and staining of the tight junction and cannot explain the effect of TK on basal laminae and pericytes. Thirdly, we used LPS to induce BBB integrity disruption and investigated the role of TK on the LPS-induced hyperpermeability of BBB. Due to the complex role and mechanism of LPS in opening BBB, we could not eliminate the interference of these confounding factors. Therefore, future studies will address these limitations by conducting experiments in EC-specific TK-knock out mice, by investigating the effect and mechanism of TK on BBB permeability after BBB is opened by inflammation, I/R, and other factors in vivo and in vitro. Moreover, whether different doses of TK exert protective effects on cerebral vessels and tissues while affecting BBB is also worth investigating in the future.
Conclusions
In summary, our observations suggest that TK may selectively increase the permeability of normal BBB by upregulating the B2R-dependent eNOS signaling and subsequently reduce inter-EC junctions, exacerbate the impairment of BBB through disrupt ZO-1 via B1R/iNOS signaling. Since the success of CNS therapeutic development is often dependent upon techniques for modulating BBB permeability, the present study likely has important implications for the treatment of stroke and other CNS disorders.
Abbreviations
HUK, human urinary kallidinogenase; TK, tissue kallikrein; KKS, kallikrein-kinin system; BBB, blood–brain barrier; B1R, bradykinin 1 receptor; B2R, bradykinin 2 receptor; NO, nitric oxide; eNOS, endothelial nitric oxide synthase; iNOS, inducible nitric oxide synthase; LPS, lipopolysaccharide; I/R, ischemia-reperfusion; BMEC, brain microvascular endothelial cell; ZO-1, zonula occludens-1; EB, Evans blue; FITC, fluorescein isothiocyanate; TER, transendothelial electrical resistance; ECIS, electrical cell-substrate impedance sensing system; MFT, multiple frequency/time; qPCR, quantitative real-time PCR; SDS-PAGE, sodium dodecyl sulfate-polyacrylamide gel electrophoresis; PVDF, polyvinyl difluoride membranes; vWF, von Willebrand Factor; GFAP, glial fibrillary acidic protein; DAPI, Vectashield-containing 4ʹ,6-diamidino-2-phenylindole.
Data Sharing Statement
The data sets used and analyzed during the current study are available from the corresponding author on reasonable request.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Acknowledgments
TK was provided by Techpool Bio-Pharma Co., Ltd. (Guangzhou, China).
Disclosure
The authors declare no conflicts of interest.
Additional information
Funding
References
- Dong Y, Qu J, Zhang Z, Wang C, Dong Q. Human urinary kallidinogenase in treating acute ischemic stroke patients: analyses of pooled data from a randomized double-blind placebo-controlled phase IIb and Phase III clinical trial. Neurol Res. 2020;42(4):286–290. doi:10.1080/01616412.2020.1711648
- Hillmeister P, Persson PB. The Kallikrein-kinin system. Acta Physiol. 2012;206(4):215–219. doi:10.1111/apha.12007
- Chao J, Bledsoe G, Yin H, Chao L. The tissue kallikrein-kinin system protects against cardiovascular and renal diseases and ischemic stroke independently of blood pressure reduction. Biol Chem. 2006;387(6):665–675. doi:10.1515/BC.2006.085
- Zhang Q, Ding H, Yan J, et al. Plasma tissue kallikrein level is negatively associated with incident and recurrent stroke: a multicenter case-control study in China. Ann Neurol. 2011;70(2):265–273. doi:10.1002/ana.22404
- Han L, Li J, Chen Y, et al. Human urinary kallidinogenase promotes angiogenesis and cerebral perfusion in experimental stroke. PLoS One. 2015;10(7):e0134543. doi:10.1371/journal.pone.0134543
- Li J, Chen Y, Zhang X, Zhang B, Zhang M, Xu Y. Human urinary kallidinogenase improves outcome of stroke patients by shortening mean transit time of perfusion magnetic resonance imaging. J Stroke Cerebrovasc Dis. 2015;24(8):1730–1737. doi:10.1016/j.jstrokecerebrovasdis.2015.03.032
- Chen ZB, Huang DQ, Niu FN, Zhang X, Li EG, Xu Y. Human urinary kallidinogenase suppresses cerebral inflammation in experimental stroke and downregulates nuclear factor-kappaB. J Cereb Blood Flow Metab. 2010;30(7):1356–1365. doi:10.1038/jcbfm.2010.19
- Shi Y, Zhang L, Pu H, et al. Rapid endothelial cytoskeletal reorganization enables early blood-brain barrier disruption and long-term ischaemic reperfusion brain injury. Nat Commun. 2016;7:10523. doi:10.1038/ncomms10523
- Xia CY, Zhang Z, Xue YX, Wang P, Liu YH. Mechanisms of the increase in the permeability of the blood-tumor barrier obtained by combining low-frequency ultrasound irradiation with small-dose bradykinin. J Neurooncol. 2009;94(1):41–50. doi:10.1007/s11060-009-9812-9
- Zhang H, Gu YT, Xue YX. Bradykinin-induced blood-brain tumor barrier permeability increase is mediated by adenosine 5ʹ-triphosphate-sensitive potassium channel. Brain Res. 2007;1144:33–41. doi:10.1016/j.brainres.2007.01.133
- Kuhr F, Lowry J, Zhang Y, Brovkovych V, Skidgel RA. Differential regulation of inducible and endothelial nitric oxide synthase by kinin B1 and B2 receptors. Neuropeptides. 2010;44(2):145–154. doi:10.1016/j.npep.2009.12.004
- Bartus RT, Elliott PJ, Dean RL, et al. Controlled modulation of BBB permeability using the bradykinin agonist, RMP-7. Exp Neurol. 1996;142(1):14–28. doi:10.1006/exnr.1996.0175
- Schilling L, Wahl M. Mediators of cerebral edema. Adv Exp Med Biol. 1999;474:123–141.
- Raslan F, Schwarz T, Meuth SG, et al. Inhibition of bradykinin receptor B1 protects mice from focal brain injury by reducing blood-brain barrier leakage and inflammation. J Cereb Blood Flow Metab. 2010;30(8):1477–1486. doi:10.1038/jcbfm.2010.28
- Gobel K, Pankratz S, Schneider-Hohendorf T, et al. Blockade of the kinin receptor B1 protects from autoimmune CNS disease by reducing leukocyte trafficking. J Autoimmun. 2011;36(2):106–114. doi:10.1016/j.jaut.2010.11.004
- Tang M, Liu P, Li X, Wang JW, Zhu XC, He FP. Protective action of B1R antagonist against cerebral ischemia-reperfusion injury through suppressing miR-200c expression of microglia-derived microvesicles. Neurol Res. 2017;39(7):612–620. doi:10.1080/01616412.2016.1275096
- Wei Z, Lyu Y, Yang X, Chen X, Zhong P, Wu D. Therapeutic values of human urinary kallidinogenase on cerebrovascular diseases. Front Neurol. 2018;9:403. doi:10.3389/fneur.2018.00403
- Kumar M, Tyagi N, Moshal KS, et al. GABAA receptor agonist mitigates homocysteine-induced cerebrovascular remodeling in knockout mice. Brain Res. 2008;1221:147–153. doi:10.1016/j.brainres.2008.04.021
- Gurses C, Ekizoglu O, Orhan N, et al. Levetiracetam decreases the seizure activity and blood-brain barrier permeability in pentylenetetrazole-kindled rats with cortical dysplasia. Brain Res. 2009;1281:71–83. doi:10.1016/j.brainres.2009.05.033
- Guo C, Wang H, Liang W, et al. Bilobalide reversibly modulates blood-brain barrier permeability through promoting adenosine A1 receptor-mediated phosphorylation of actin-binding proteins. Biochem Biophys Res Commun. 2020;526(4):1077–1084. doi:10.1016/j.bbrc.2020.03.186
- Abbott NJ, Patabendige AA, Dolman DE, Yusof SR, Begley DJ. Structure and function of the blood-brain barrier. Neurobiol Dis. 2010;37(1):13–25. doi:10.1016/j.nbd.2009.07.030
- Vernon H, Clark K, Bressler JP. In vitro models to study the blood brain barrier. Methods Mol Biol. 2011;758:153–168.
- Weinstein DE. Isolation and purification of primary rodent astrocytes. Curr Protoc Neurosci. 2001;1:3–5.
- Sobczak M, Dargatz J, Chrzanowska-Wodnicka M. Isolation and culture of pulmonary endothelial cells from neonatal mice. J Vis Exp. 2010;14:e2316.
- Aneja RK, Alcamo AM, Cummings J, et al. Lack of benefit on brain edema, blood-brain barrier permeability, or cognitive outcome in global inducible high mobility group box 1 knockout mice despite tissue sparing after experimental traumatic brain injury. J Neurotrauma. 2019;36(2):360–369. doi:10.1089/neu.2018.5664
- Daneman R, Barres BA. The blood-brain barrier–lessons from moody flies. Cell. 2005;123(1):9–12. doi:10.1016/j.cell.2005.09.017
- Ben-Zvi A, Lacoste B, Kur E, et al. Mfsd2a is critical for the formation and function of the blood-brain barrier. Nature. 2014;509(7501):507–511. doi:10.1038/nature13324
- Szulcek R, Bogaard HJ, van Nieuw Amerongen GP. Electric cell-substrate impedance sensing for the quantification of endothelial proliferation, barrier function, and motility. J Vis Exp. 2014;85:51300.
- Ke Y, Oskolkova OV, Sarich N, et al. Effects of prostaglandin lipid mediators on agonist-induced lung endothelial permeability and inflammation. Am J Physiol Lung Cell Mol Physiol. 2017;313(4):L710–L721. doi:10.1152/ajplung.00519.2016
- Haileselassie B, Joshi AU, Minhas PS, Mukherjee R, Andreasson KI, Mochly-Rosen D. Mitochondrial dysfunction mediated through dynamin-related protein 1 (Drp1) propagates impairment in blood brain barrier in septic encephalopathy. J Neuroinflammation. 2020;17(1):36. doi:10.1186/s12974-019-1689-8
- Fujimoto T, Morofuji Y, Kovac A, et al. Pitavastatin ameliorates lipopolysaccharide-induced blood-brain barrier dysfunction. Biomedicines. 2021;9(7):837. doi:10.3390/biomedicines9070837
- Fanning AS, Jameson BJ, Jesaitis LA, Anderson JM. The tight junction protein ZO-1 establishes a link between the transmembrane protein occludin and the actin cytoskeleton. J Biol Chem. 1998;273(45):29745–29753. doi:10.1074/jbc.273.45.29745
- Silva LS, Pinheiro AS, Teixeira DE, et al. Kinins released by erythrocytic stages of Plasmodium falciparum enhance adhesion of infected erythrocytes to endothelial cells and increase blood brain barrier permeability via activation of bradykinin receptors. Front Med. 2019;6:75. doi:10.3389/fmed.2019.00075
- Miao J, Deng F, Zhang Y, Xie HY, Feng JC. Exogenous human urinary kallidinogenase increases cerebral blood flow in patients with acute ischemic stroke. Neurosciences. 2016;21(2):126–130. doi:10.17712/nsj.2016.2.20150581
- Abdullahi W, Tripathi D, Ronaldson PT. Blood-brain barrier dysfunction in ischemic stroke: targeting tight junctions and transporters for vascular protection. Am J Physiol Cell Physiol. 2018;315(3):C343–C356. doi:10.1152/ajpcell.00095.2018
- Sikpa D, Whittingstall L, Savard M, et al. Pharmacological modulation of blood-brain barrier permeability by kinin analogs in normal and pathologic conditions. Pharmaceuticals. 2020;13(10):279. doi:10.3390/ph13100279
- Liu Y, Hashizume K, Chen Z, et al. Correlation between bradykinin-induced blood-tumor barrier permeability and B2 receptor expression in experimental brain tumors. Neurol Res. 2001;23(4):379–387. doi:10.1179/016164101101198596
- Regoli D, Gobeil F. Kinins and peptide receptors. Biol Chem. 2016;397(4):297–304. doi:10.1515/hsz-2015-0240
- Nico B, Quondamatteo F, Herken R, et al. Developmental expression of ZO-1 antigen in the mouse blood-brain barrier. Brain Res Dev Brain Res. 1999;114(2):161–169. doi:10.1016/S0165-3806(99)00008-5
- Chao J, Shen B, Gao L, Xia CF, Bledsoe G, Chao L. Tissue kallikrein in cardiovascular, cerebrovascular and renal diseases and skin wound healing. Biol Chem. 2010;391(4):345–355. doi:10.1515/bc.2010.042
- Shi R, Yuan K, Hu B, et al. Tissue kallikrein alleviates cerebral ischemia-reperfusion injury by activating the B2R-ERK1/2-CREB-Bcl-2 signaling pathway in diabetic rats. Oxid Med Cell Longev. 2016;2016:1843201. doi:10.1155/2016/1843201
- Sang H, Qiu Z, Cai J, et al. Early increased bradykinin 1 receptor contributes to hemorrhagic transformation after ischemic stroke in type 1 diabetic rats. Transl Stroke Res. 2017;8(6):597–611. doi:10.1007/s12975-017-0552-4
- Wagner S, Kalb P, Lukosava M, Hilgenfeldt U, Schwaninger M. Activation of the tissue kallikrein-kinin system in stroke. J Neurol Sci. 2002;202(1–2):75–76. doi:10.1016/S0022-510X(02)00208-3
- Zlokovic BV. The blood-brain barrier in health and chronic neurodegenerative disorders. Neuron. 2008;57(2):178–201. doi:10.1016/j.neuron.2008.01.003
- Nguyen LN, Ma D, Shui G, et al. Mfsd2a is a transporter for the essential omega-3 fatty acid docosahexaenoic acid. Nature. 2014;509(7501):503–506. doi:10.1038/nature13241
- Chen H, Konofagou EE. The size of blood-brain barrier opening induced by focused ultrasound is dictated by the acoustic pressure. J Cereb Blood Flow Metab. 2014;34(7):1197–1204. doi:10.1038/jcbfm.2014.71
- Patel JP, Frey BN. Disruption in the blood-brain barrier: the missing link between brain and body inflammation in bipolar disorder? Neural Plast. 2015;2015:708306. doi:10.1155/2015/708306