Abstract
The involvement of stemness factors in cancer initiation and progression has drawn much attention recently, especially after the finding that introducing four stemness factors in somatic cells is able to reprogram the cells back to an embryonic stem cell-like state. Following accumulating data revealing abnormal elevated expression levels of key stemness factors, like Nanog, Oct4, and Sox2, in several types of cancer stem cells; the importance and therapeutic potential of targeting these stemness regulators in cancers has turned to research focus. Nanog determines cell fate in both embryonic and cancer stem cells; activating Nanog at an inappropriate time would result in cancer stem cells rather than normal pluripotent stem cells or differentiated somatic cells. Upregulated Nanog is correlated with poor survival outcome of patients with various types of cancer. The discoveries of downstream regulatory pathways directly or indirectly mediated by Nanog indicate that Nanog regulates several aspects of cancer development such as tumor cell proliferation, self-renewal, motility, epithelial-mesenchymal transition, immune evasion, and drug-resistance, which are all defined features for cancer stem cells. The current review paper illustrates the central role of Nanog in the regulatory networks of cancer malignant development and stemness acquirement, as well as in the communication between cancer cells and the surrounding stroma. Though a more defined model is needed to test the therapeutic efficacy of targeting Nanog as a cancer treatment method, current animal experiments using siNanog or shNanog have shown the promising therapeutic potential of Nanog targeting in several types of cancer.
Keywords:
Cancer stem cells
Human tissues, under physiological conditions, are able to maintain their mass and architecture over time through a tightly regulated process of renovation. This process is sustained by a minor population of long-lived cells with unlimited expansion potential, known as stem cells, which are defined as cells that have the ability to perpetuate themselves through self-renewal and to generate mature cells of a particular tissue through differentiation.
Tumors can be considered as aberrant organs that are initiated by tumorigenic cancer cells that acquired the capacity for indefinite proliferation through accumulating mutations. Like their normal tissue counterparts, tumors are composed of heterogeneous populations of cells that differ in their degree of accumulated mutations and state of differentiation. Evidence was outlined in supporting the hierarchical organization of normal human breast tissue and the breast stem or luminal progenitor cells as targets for transformation.Citation1 The idea that cancer is raised from a subpopulation of tumor cells with stem cell properties, which are latterly termed cancer stem cells (CSCs), was proposed around 150 years ago.Citation1,Citation2 This unique subpopulation of cancer cells are capable of propagating the tumor as well as developing high sustainability against conventional therapeutic drugs, and, therefore, account for intratumoral heterogeneity and malignant behaviors like metastasis and recurrence.Citation2 Though the identity of CSCs is still argued, the definition of CSCs is in concord based on the functional characteristics of these cells: the ability to initiate tumors in immunocompromised or syngeneic mice, self-renewal capacity measured by tumor formation in secondary mice in vivo and sphere formation in vitro, and the capacity to differentiate into the non-self-renewable cancer cells with diversity.Citation1
Identification of CSCs
During the last decades, the existence and the identity of CSCs have been identified in hematopoietic tumors as well as a range of solid tumors including breast, brain, lung, colon, prostate, head and neck, and others.Citation3–Citation11 This rare subpopulation of cells with chemo- or radioresistant properties in each malignancy has greater potential of tumor initiation and accelerated regrowth after a therapeutic treatment.Citation9,Citation11–Citation20 Breast cancer was the first solid tumor from which CSC was isolated: the breast CSC was characterized as a minor population (less than 5%) of cells expressing a high level of CD44 and a low level of CD24 cell surface markers as well as being positive for epithelial cell surface antigen.Citation13,Citation21 As few as 100 of the cells with this phenotype were able to form tumors in immunocompromised mice.Citation13 Technically, CSCs can be isolated through three distinct methodologies based on the defined properties of CSCs. First of all, CSCs can be sorted by flow cytometry according to the expression pattern of surface markers such as CD24, CD44, and CD133.Citation4,Citation7,Citation22–Citation24 For example, colon, lung, and hematopoietic CSCs are isolated by cell sorting with the expression of CD133,Citation7,Citation8,Citation22,Citation25,Citation26 though the function of CD133 is still unclear. These cells grow indefinitely as spheres in vitro and were tumorigenic in vivo.Citation8 Second, with Hoechst 33342 staining, CSCs are enriched in the side populations of cancer cells, excluding intracellular Hoechst 33342 in vitro,Citation27,Citation28 and can be isolated by flow cytometry. Expression of ATP-binding cassette subfamily G member 2 (ABCG2), an ATPase transporter, in CSC is found closely associated with its specific exclusion capacity, rendering ABCG2 a CSC marker in some cases.Citation29 Third, CSCs are enriched in a population of cells that are capable of forming sphere bodies under defined serum-free cultivation medium plus necessary growth factors according to individual solid tumor or cancer.Citation14,Citation30 This cultivation condition also helps the CSC to maintain its undifferentiated state.Citation14,Citation30
CSCs as therapeutic target
The increasing interests in the CSC model are dramatically altering the current research directions in cancer treatments and drug developments. The majority of cancer cells within a tumor are nontumorigenic; therapies targeting these cells would cause tumor regression. However, if therapies fail to target the tumorigenic CSCs, then these cells would persist after therapies and be able to regenerate the tumor, resulting in tumor relapse or recurrence. Therefore, eradication of cancers requires the elimination of CSCs. Advanced strategies that specifically target CSCs without harming normal cells are urgently needed to advance current therapeutic treatments. The origin of CSCs is still debated as they may be derived from adult stem cells, which accumulate oncogenic mutations to functionally divert from normal stem cells, or from terminally differentiated cancer cells that undergo a reprogramming-like process to acquire stem-like properties. It is, however, generally accepted that several embryonic stem cell-specific signalings are reactivated in CSCs in several cancer models. The embryonic stem cell-specific transcriptional factor, Nanog, for instance, was shown to highly express in hepatic, colorectal, and brain CSCs.Citation31–Citation33 CSCs in brain tumors were demonstrated to express various neural stem/progenitor-specific proteins such as sex determine region Y-box 2 (Sox2), octamer-binding transcription factor 4 (Oct4 [also known as POU5F1]), nestin, and Musashi.Citation15 Our previous report demonstrated high expression of Oct4 and Nanog in malignant lung adenocarcinoma cells, and forced overexpression of Oct4 and Nanog transformed the lung cancer cells into a CSC-like state.Citation34 Accumulating reports have recently placed interests on identifying CSC molecules, including transcription factors, cell surface markers, stemness-related signaling pathways, and microRNAs (miRNAs), in the hope of identifying targets for potential therapeutic development that can completely eliminate CSCs and prevent the recurrence of tumors.
The Nanog transcriptional factor
Nanog is a homeodomain-containing transcription factor and, along with the POU domain-containing Oct4 and high mobility group domain-containing Sox2, is part of the key set of transcription factors that are involved in the maintenance of pluripotency and selfrenewal in undifferentiated embryonic stem (ES) cells.Citation35–Citation41 Nanog protein is encoded by the only open reading frame of the 2184-nucleotide NANOG cDNA.Citation35 Apart from the embryonic NANOG gene, there are eleven Nanog pseudogenes that have been reported in the human genome to date,Citation42 but only the Nanog homeobox pseudogene 8 (NANOGP8) has a complete open reading frame to transcribe and translate a functional Nanog protein.Citation42–Citation44 The protein derived from NANOGP8 is almost identical to that from NANOG with only one amino acid alteration (from Gln-253 in NANOG to His-253 in NANOGP8).Citation5 Comprehensive expression patterns of NANOG and NANOGP8 in human cancers have not been fully addressed. Although it is presumed that embryonic NANOG is an important regulator of pluripotency while NANOGP8 plays a role in tumorigenesis,Citation5 the defined line between the roles of the two genes is becoming blurred. A functional and molecular analysis in prostate cancer tissue revealed that Nanog mRNA in cancer cells is derived predominantly from NANOGP8, and the majority of Nanog protein is detected in the nucleus of cancer cells.Citation45 On the other hand, Nanog was found to be expressed from both NANOG and NANOGP8 in colorectal cancers.Citation5
Human Nanog protein consists of 305 amino acidsCitation35,Citation36 and can be divided into N-terminal (amino acid 1–95), homeobox domain (amino acid 96–155), and C-terminal (amino acid 156–305) regions.Citation35,Citation46 The N-terminus is rich in serine, threonine, and proline, providing a structural motif for the transcriptional activity of Nanog. This region is tightly regulated through phosphorylation or other post-translational modifications.Citation46,Citation47 The C-terminus contains two potent transactivation subdomains.Citation46,Citation47 The homeobox domain in the central region contains a DNA-binding motif; its N- and C-terminal regions are shown to contain nuclear localization sequencesCitation47 and its middle region is reported to harbor potent nuclear export motif,Citation48 allowing the Nanog protein to transport in and out of the nucleus. Nanog is also one of the key transcription factors that could reprogram a human somatic fibroblast into an embryonic stem cell-like pluripotent cell, termed inducible pluripotent stem cell.Citation49 A recent study revealed that a short sequence in the well conserved homeobox domain of Nanog was sufficient to induce pluripotency in Nanog-deficient somatic cells,Citation50 indicating a crucial role of the homeobox domain in mediating the reprogramming ability of Nanog and that the transcriptional activity of Nanog might be dispensable.
Roles of Nanog in embryonic stem cells
Nanog mRNA is present in pluripotent mouse and human stem cell lines, and absent from differentiated cells.Citation36 Functional screening for the selfrenewal determinants in ES cells revealed that Nanog expression is crucial to maintain ES cell identity.Citation36 Hyslop et al reported that small interfering RNA (siRNA)-mediated downregulation of Nanog in human embryonic stem cells induces a pro-extraembryonic lineage differentiation, evidenced by the upregulated endoderm- and trophectoderm-associated genes,Citation37 suggesting that Nanog acts as a gatekeeper of pluripotency in human embryonic development.
The expression of Nanog has been shown to be regulated by Oct4/Sox2 heterodimers, in which Oct4/Sox2 binds to the octamer/sox elements within the Nanog proximal promoter region and induces Nanog transcription.Citation51,Citation52 Although the Oct4/Sox2 complex may seem to locate in a higher position in the hierarchical regulatory network of ES cells, Nanog possesses unique properties that divert from the other key stemness transcriptional factors. The leukemia inhibitory factor (LIF) has been utilized to maintain the symmetrical selfrenewal of mouse ES cells.Citation53 Through activating its downstream effector, signal transducer and activator of transcription 3 (STAT3), the LIF/STAT3 pathway has been proved to be indispensable in the maintenance of pluripotent state of mouse ES cells. Several key transcriptional factors, including Oct4 and Sox2, are shown to be functionally dependent on LIF-STAT3. Constitutively activated Oct4 from an exogenous promoter in ES cells still required LIF for inducing selfrenewal in ES cells.Citation54 In 2003, two independent groups, Chambers et al and Mitsui et al, applied different strategies to screen for critical factors that can maintain ES cell pluripotency independently of the LIF-STAT3 pathway and found Nanog.Citation35,Citation36 Their reports demonstrated that Nanog overexpression relieves ES selfrenewal from dependence on the activity of LIF-STAT3 pathway. Moreover, Chambers’ report showed that Nanog is expressed in Oct4-deficient embryos, and Nanog overexpression does not revert the differentiation program of ES cells triggered by Oct4 downregulation.Citation36 These results suggest that Nanog is not just a downstream of Oct4, and Nanog and Oct4 work in concert to support stem cell potency and selfrenewal.
Nanog, tumorigenicity, and cancer stemness
Expression of Nanog has been detected by several groups in germ cell tumors as well as other tumors, including breast, cervix, oral, kidney, prostate, lung, gastric, brain, and ovarian cancer.Citation10,Citation34,Citation55–Citation65 Strong expression of Nanog is shown as an indicator of a poor prognosis for ovarian serous carcinoma, colorectal, and breast cancer patients.Citation66–Citation68 In oral squamous cell and lung adenocarcinoma, higher expression of Nanog, along with Oct4, was associated with advanced cancer stage and shorter patient survival rate.Citation34,Citation63
The expression level of Nanog has been shown to be higher in cancer stem cells than non-stemness cancer cells in several types of cancers.Citation4,Citation5,Citation10,Citation31,Citation32,Citation65,Citation69,Citation70 In some cases, Nanog-positive cancer stem cells only comprise less than 2% of the total cancer cell population.Citation5 The difference may be due to the fact that several key factors in the stemness network, such as Oct4 and Sox2, are also increased in CSC, which increases stemness-related genes like Nanog; or, it is possibly because of the increased activity or expression of upstream regulators of Nanog, such as Hedgehog,Citation71 Ezh2,Citation72 and several miRNAs. This question, as well as the causal relationship between elevated stemness pathways and CSC properties, is still under investigation in this field. Moreover, a direct regulatory link between Nanog and CSC surface markers is yet to be clarified. There are, however, several correlation studies demonstrating that cancer cells expressing high levels of the CSC surface marker would possess higher levels of stemness genes like Nanog. For example, CD133+ or CD44+ cancer cells express significant higher level of Nanog in comparison to CD133− or CD44+ ones, respectively.Citation73–Citation75 On the other hand, Nanog induction in prostate cancer cell lines results in upregulation of CD133 and ALDH1.Citation70
Functional studies have demonstrated that Nanog is not only a CSC marker, but also promotes CSC-like characteristics in several cancers. For example, ectopic overexpression of the NANOGP8 gene in prostate cancer cells enhanced clonal growth and tumor regenerative capacity,Citation70 and the activation of embryonic NANOG gene drives a subpopulation of colorectal cancer cells to adopt a stem-like phenotype.Citation5 Overexpression of Nanog also increased drug-resistance capacity in breast cancer cell lines.Citation70 The Nanog protein can bind to the promoter region of cyclin D1 and regulate cell cycle and proliferation.Citation76,Citation77 Moreover, Nanog positively regulates cancer cell motility and tumor metastasis capability. Nanog was found highly expressed in ovarian cancer cell lines with metastasis-associated property and in clinical samples of metastatic foci. Knockdown of Nanog impeded cell proliferation, migration, and invasion.Citation78
Recently, Noh et al reported that Nanog activates the Akt signaling pathway through T-cell leukemia/lymphoma protein 1 A (Tcl1a), and hyperactivation of the Nanog/Tcl1a/Akt signaling axis is conserved across multiple types of human cancer.Citation79 They demonstrated that the Nanog-mediated activation of Akt pathway renders cancer cells adapted to host immune system and leads to a successful escape from the immune-mediated clearance. Therefore, the failure of cancer vaccination may be due to a Nanog-dependent evolution of tumor cells toward an immune-resistant and stem-like phenotype.Citation80 Apart from cancer cells, Gu et al showed that the cancer stromal cells that express high levels of cytoplasmic Nanog may promote human cervical cancer progression,Citation81 suggesting a role of Nanog in regulating the cross talk between cancer cells and cancer associated stromal cells.
Upstream regulators of Nanog in cancers
In ES cells, Nanog is involved in a complicated stemness regulatory network in cooperation with other key transcriptional factors, such as Oct4, Sox2, and Lin 28, to precisely balance between pluripotency and differentiation tendency. Kalmar et al reported that Nanog presents a transcriptional fluctuation between low and high expression level in ES cells.Citation82 They proposed that the dynamic distribution of Nanog is crucial for ES cells to maintain the undifferentiated state and yet possess the potential to be differentiated without marking definitive commitment if a proper differentiation signaling strikes the cells. The tightly regulated Nanog seems to be a key switch that determines the fate of ES cells.Citation83 In cancer cells, Nanog is usually overexpressed, especially in the malignant, high grade, poorly differentiated ones, and the fluctuation of Nanog expression in cancer cells has so far not been reported. It seems that deregulated and abnormal Nanog expression drives the cells to a reprogramming-like process but fails to keep cells in the track that leads to an ordinary stemness state, resulting in cells going on a diverted route that ends with upregulation of several downstream signalings involving in tumorigenesis.
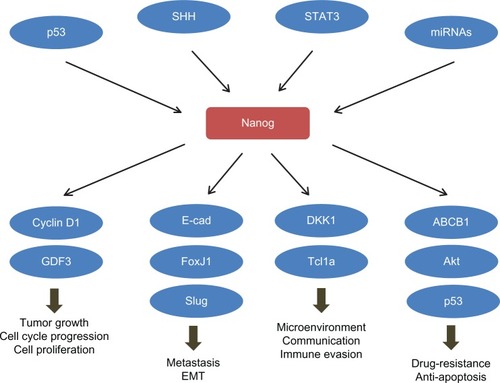
Despite the Oct4 and Sox2 stemness factors that are involved in ES regulatory networks, many regulators, including microRNAs, transcription factors, and kinases, have been reported to mediate the silencing or overexpression of Nanog and thus regulate the stemness and malignant transformation, as well as CSC-like phenotypes in cancer cells (). To date, most of the regulatory mechanisms on Nanog are focused on the transcription level, though accumulating data have also emphasized the importance of post-transcriptional and translational regulation of Nanog.
Table 1 Upstream regulators of Nanog
p53
p53 is a stress-activated transcription factor that prevents the proliferation of genetically damaged cells. Given that p53 is a key mediator regulating programmed cell death (apoptosis) and cell-cycle related pathways, such as activating cell-cycle checkpoint and promoting cell senescence, it has been well accepted as a major player for tumor suppression. Loss or inactivation of p53 facilitates the clonal expansion of aberrant cells and leads to genomic instability. During the last decade, a direct involvement of p53 in the stemness regulatory network has emerged and draws much attention in the fields of both stem cell and cancer research. In mouse ES cells, p53 directly binds to Nanog promoter through two consensus p53-binding motifs and suppresses Nanog transcription in response to DNA damage.Citation84 The DNA damage-induced and p53-mediated suppression of Nanog, which depends on the phosphorylation at serine 315 of p53, results in reduced selfrenewal and enhanced differentiation tendency of mouse ES cells.Citation85 Through inducing differentiation via Nanog suppression, p53 protects the genomic intactness and maintains genetic stability in ES cells. The p53-Nanog regulatory signaling was also implicated in cancer cells, especially in brain CSCs. Nanog was shown to induce the CSC-like properties in primary p53-defecient mature mouse astrocytes, but not in the astrocytes with intact p53,Citation86 suggesting p53 as the gatekeeper, not only preventing normal cells from tumor transformation, but also preventing cancer cells from further acquirement of CSC properties. These findings also echo the discovery that, during the reprogramming process, suppression of p53 is essential to enhance the generation of induced pluripotent stem (iPS) cells.Citation87 Hong et al reported that loss of p53 facilitates cellular immortalization, and suggested that the p53-p21 pathway serves as a barrier in iPS cell generation as well as tumorigenicity.Citation87 Given that expression of Nanog improves the cloning efficiency of human ES cells, it could therefore increase the survival rate of early reprogrammed cells.Citation88 The suppressive effect of p53 on reprogramming and cancer-stemness could partially count on its negative regulation of Nanog. Since approximately >50% of cancers harbored p53 mutation or deficiency, blockade of Nanog, and, therefore, Nanog mediated regulatory circuits, would be a potential method to prevent tumor malignancy.
Hedgehog
Nanog and hedgehog are both essential regulators of stemness properties by promoting selfrenewal in ES cells during the early stage of embryonic development as well as in cancer progression. The sonic hedgehog and its downstream transcription factor, GLI family zinc finger 1 (Gli1), have been shown to promote CSC survival, tumor growth, and the expression of stem-like signature in human glioma cells,Citation89 suggesting a direct regulatory role of hedgehog-Gli1 in the core stemness network. Indeed, Po et alCitation71 later on reported a direct regulatory pathway between hedgehog signaling and Nanog. They showed that the downstream effectors of hedgehog activity, Gli1 and GLI family zinc finger 2, directly bind to the cis-regulatory sequences of the NANOG gene in both mouse and human neural stem cells, and thus activate Nanog transcription, which is essential for hedgehog-induced selfrenewal in neural stem cells.Citation71 They suggested a model in which Nanog is a critical mediator of hedgehog effect on neural stem cells. This regulation does not go through p53; on the other hand, since p53 is known to suppress sonic hedgehog signaling, they proposed that p53 might inhibit Nanog transcription partially through the suppression of hedgehog-Gli pathway.Citation71 Moreover, Zbinden et al demonstrated a positive regulatory loop between Nanog and sonic hedgehog signaling; knockdown of Nanog decreased the endogenous activity of Gli1 whereas the Nanog mRNA levels were modulated by hedgehog pathway.Citation32 This positive feedback loop is negatively regulated by p53, and suppression of Nanog enhanced p53 and repressed Gli1, suggesting that Nanog is situated in a neat balance between p53 and hedgehog signaling. Deregulation of the balance between p53 and hedgehog signaling would result in aberrant expression of Nanog, which further feedback to both p53 and hedgehog signalings and resulted in either acquirement of stem-like properties of cells or shrinking of the CSC population.
STAT3
STAT3 is one of the major effectors of the LIF pathway, which is essential for the maintenance of the pluripotency in ES cells. A microarray analysis in mouse ES cells showed that 14 out of the 22 STAT3 target genes that contribute to the maintenance of an undifferentiated state were also regulated by Nanog, suggesting a functional redundancy and cooperation between STAT3 and Nanog.Citation90 In some cases, Nanog and STAT3 were found to bind to and synergistically activate STAT3-dependent promoters.Citation91 The direct interaction between Nanog and STAT3 was found by Bourguignon et al;Citation92 they showed the Nanog-STAT3 complex in head and neck squamous cell carcinoma cells. The complex translocates to the nucleus and transcriptionally activates their common targets.Citation92 On the other hand, LIF-induced phosphorylation of STAT3 directs the binding of STAT3 to the enhancer of NANOG gene and, thus, results in upregulation of Nanog expression in ES cells.Citation93 Moreover, it is also suggested that STAT3 regulates Nanog expression through methylation, as inhibiting the Janus kinase/STAT3 activity blocks demethylation of Nanog during the reprogramming process, along with upregulation of DNA methyltransferase 1 and histone deacetylases.Citation94 This indicates that, apart from cooperative partner, STAT3 may also regulate Nanog expression through epigenetic modification. The fact that LIF/STAT3 pathway promotes Nanog expression in ES and cancer cells suggests Nanog as an important mediator for LIF-dependent maintenance of pluripotency, which may explain why Nanog-maintained pluripotency of cultured ES cells does not require the addition of LIF. Moreover, through activating STAT3, molecules like CD24Citation4 and E-cadherin,Citation95 are also shown to modulate Nanog expression, both in ES cells and in cancer cells.
microRNAs
miRNAs are short RNAs that direct the degradation of messenger RNA or disruption of messenger RNA translation in a sequence-dependent manner. Through targeting the core stemness factors, such as Oct4, Nanog, and Sox2, the up- and downregulation of miRNAs, such as miR134, miR296, and miR470 play roles in modulating the selfrenewal, pluripotency, and differentiation of ES cells.Citation96 There are increasing reports identifying miRNAs that target Nanog and mediate the malignant progression in cancer cells or CSCs. miR134 was first found directly targeting Nanog in mouse ES cells, through which it promotes mouse ES cells differentiation toward ectodermal lineages.Citation97 The same phenomenon was also found in glioblastoma cells; overexpression of miR134 reduced the proliferation, invasiveness, and migration capability, and promoted apoptosis of glioblastoma cell lines through directly suppressing Nanog expression.Citation98 Enhanced expression of miR214 has been found in several human malignanciesCitation99,Citation100 and was associated with late-stage and high-grade ovarian cancer.Citation101 Xu et alCitation101 reported that Nanog level was induced by overexpressed miR214 in p53-wildtype ovarian cancer cell lines, but not in p53-mutant ones, linking miR214 with the p53/Nanog pathway. They further showed p53 as a direct target of miR214, and demonstrated that, through suppressing p53 and consequently elevating Nanog, miR214 increased ovarian cancer cell stemness.Citation101 In glioblastoma, miR302-367 cluster was found upregulated when suppressing the stemness of CSC. This miRNA cluster was shown to suppress the stemness properties (like stemness signature, selfrenewal, and cell infiltration) of glioblastoma CSC through disrupting the sonic hedgehog-Gli1-Nanog signaling pathway.Citation102 These reports emphasize a role of Nanog in miRNAs-mediated regulation of CSC properties, and therefore suggest Nanog as a potential target to suppress the increased cancer stemness due to deregulated miRNAs.
Downstream targets and cellular functions of Nanog in cancer
Nanog is involved in several cellular functions that positively affect tumor development and progression, such as cell proliferation, cell cycle progression, motility, apoptosis, and the cross-communication between cancer cells and their surrounding stromal cells. In some cases, ectopic overexpression of Nanog is sufficient to induce malignant transformation.Citation103,Citation104 Investigators have endeavored to discover through which mechanisms Nanog regulates tumorigenesis. The identified downstream targets of Nanog are listed in .
Table 2 Downstream modulators of Nanog
Cell proliferation and cell cycle
Nanog is well-known for being involved in the regulation of selfrenewal in ES cells. In cancer cells, aberrant expression of Nanog has been linked to increased proliferation rate in vitro and tumor growth in vivo. We previously showed that overexpression of Nanog, along with Oct4, increased clonogenic growth and spheroid body formation of lung adenocarcinoma cells.Citation34 Knockdown of Nanog/Oct4 in lung cancer,Citation34 or Nanog alone in breast cancer cells,Citation76 suppressed clonogenic growth and proliferation. A global gene expression profile screening in Nanog siRNA-transfected embryonal carcinoma cells suggested the involvement of Nanog in cell cycle-signaling pathway.Citation77 This group reported that with Nanog knockdown alone, several cell cycle-related genes (such as Cyclins D1, D2, D3, and E1 as well as cyclin-dependent kinase 1 and 6) as well as p53-related signaling pathway (such as Bcl6 and Atf3) were downregulated, suggesting a role of Nanog in cell cycle and survival.Citation77 In breast cancer cells, Han et al found that knockdown of Nanog results in inhibited cell proliferation, G0/G1 arrest of cell cycle, and suppressed expression of cyclin D1 and oncogene c-Myc.Citation76 They further demonstrated that Nanog directly binds to the minimal promoter region of cyclin D1, transcriptionally regulates its expression, and thus drives the cell cycle moving forward and accelerates cell proliferation.Citation76 Growth differentiation factor-3, a member of the transforming growth factor β superfamily, is another direct transcriptional target of Nanog that mediates Nanog-dependent regulation of growth and transformation in embryonic carcinoma.Citation105 However, the existence of this regulation, as well as its physiological effects, in other cancer models are still elusive. Further clarification of the mechanism through which Nanog regulates cell proliferation is still needed to shed light on the role of Nanog in tumor growth.
Migration and invasion
Overexpression of Nanog is associated with tumor metastasis in several types of cancer.Citation59,Citation61,Citation66–Citation68,Citation73,Citation76,Citation106 In ovarian cancer patient samples, increased nuclear Nanog expression was significantly associated with high-grade, serious histological subtypes, and poor disease-free survival.Citation78 The correlation between Nanog overexpression and advanced stage of cancer or metastatic incidence indicates a crucial role of Nanog in tumor progression. Stable knockdown of Nanog in ovarian cancer cell lines resulted in increased E-cadherin, FOXO1, FOXO3a, FOXJ1, and FOXB1 mRNA levels, whereas ectopic Nanog overexpression decreased them.Citation78 It is further claimed that Nanog-mediated cell migration and invasion involved its regulation of FOXJ1 and E-cadherin,Citation78 though whether Nanog directly regulates their expression is still unclear. We previously demonstrated that co-overexpression of Nanog and Oct4 in lung adenocarcinoma cells induces the level of Slug through a transcriptional regulation, leading to enhanced cell motility and tumor metastasis.Citation34 We found that Nanog alone is able to induce Slug transcription, an important transcription factor of epithelial-mesenchymal transition and tumor invasion.
Chemoresistance
The chemoresistant property of CSC has been associated with the expression of several stemness factors such as Oct4 and Nanog.Citation5,Citation31,Citation34,Citation70,Citation107 In human esophageal cancer, overexpression of Nanog promoted cisplatin resistance and decreased the proportion of cells undergoing apoptosis process.Citation107 We showed in lung adenocarcinoma cells that ectopic overexpression of Nanog and Oct4 enhanced the mRNA level of ATP-binding cassette sub-family B member 1 (ABCB1) – a membrane-bound transporter involved in the ATP-dependent exclusion of cellular toxic agents – leading to a high tolerance of lung adenocarcinoma cells to cisplatin treatment.Citation34 Similar phenomena were also found in oral squamous cell carcinoma, where increased Nanog and Oct4 expression was correlated with a cisplatin-resistant phenotype as well as cancer recurrence;Citation108 the chemoresistant cells also expressed high level of the drug-resistance-related gene ABCG2. In fact, Bourguignon et al reported a Nanog-STAT3-ABCB1 signaling pathway in breast and ovarian cancer that mediates the resistance against several chemotherapeutic drugs.Citation109 In their report, they showed that Nanog forms a complex with the STAT3 transcription factor in the nucleus and leads to STAT3-specific transcriptional activation and ABCB1 gene expression.Citation109 These data revealed a direct regulatory link between Nanog and the drug-resistance mechanism in cancer cells. Moreover, there are some traits of indication that Nanog may also regulate p53-related signaling and negatively affect the pro-apoptosis mechanisms.Citation77 On the other hand, an earlier report suggested that p53-dependent signaling pathway may directly suppress Nanog expression in DNA-damaged ES cells in order to maintain the gene stability.Citation110 Given that Nanog is involved in p53-dependent pro-apoptosis pathway and ABC genes expression, it is not surprising why CSCs are sustainable to apoptosis and drug treatments.
Immune tolerance
Immune-related cancer therapy, such as cancer vaccination, was once thought to be a specific method targeting cancer cells. However, positive and promising outcomes in clinical trials of this approach are so far still lacking. This may partially, if not entirely, be due to the capability of cancer cells to adapt themselves to the host immune system in order to avoid the killing and apoptosis mediated by the CD8+ cytotoxic T lymphocytes.Citation79,Citation80 Noh et al found that the application of cancer vaccination resulted in increased Nanog expression and stemness properties in tumor cells.Citation80 Though it was uncertain whether the increase was due to an immune selection or induction, the elevated Nanog drove tumor cells toward an immune-resistant state, rendering cells tolerant to the killing by cytotoxic T lymphocytes.Citation80 The same group further demonstrated that the Nanog-dependent immune resistance in tumor cells is mediated through a Tcl1a/Akt pathway.Citation79 The hyperactive Nanog/Tcl1a/Akt pathway not only increased stem-like phenotype in tumor cells but also promoted an immune evasion. These reports reveal an important role of Nanog in regulating the interaction between cancer cells and host immune cells, in which Nanog promotes the disguise of cancer cells to avoid the attack from immune system. Interestingly, delivery of siRNA against Nanog in these tumor cells reversed the immune-resistant phenotype and strongly retarded tumor growth. Therefore, it is suggested that targeting Nanog is a potential strategy for achieving immune-based therapy.
Cross talk between Nanog and oncogenic pathways
ESRRB
The orphan nuclear receptor estrogen-related receptor β (Esrrb) that modulates the estrogen signaling pathway is a vital component of the core pluripotency network in embryonic stem cells (ESCs). Previously, Esrrb was reported to initiate the expression of Nanog in ESCs. Van den Berg et al demonstrated that, through direct interaction with Oct4, Esrrb was directed to the Oct4 responsive element within Nanog promoter and induce the expression of Nanog, leading to increased self-renewal in ESC.Citation111 Moreover, the Esrrb is not only an upstream regulator of Nanog but also a direct target of Nanog. Nanog binds to Esrrb locus, recruits RNA polymerase II to the Esrrb promoter, and increases Esrrb gene expression.Citation112 Nanog and Esrrb proteins also interact directly in ESCs, and share overlap downstream targets.Citation112 Furthermore, Esrrb, like Nanog, presented a mosaic pattern in ESCs, and the mosaic patterns of the two molecules largely overlap.Citation111 Nanog and Esrrb may act to reinforce expression of the reciprocal genes through a positive feedback loop. Fluctuation of the Nanog-Esrrb feedback loop confers ESCs’ pluripotency and differentiation potential without being committed to specific lineages. Although the mutual dependence between Nanog and Esrrb was mainly discovered in ESCs, it is possible that the feedback circuit could exist in cancer or CSCs, as Nanog and accumulating members of estrogen-related receptors are implicated in cancer malignancies. Investigations into the involvement of the Nanog-Esrrb circuit in CSC properties may shed some light on how Nanog mediates selfrenewal in CSCs.
Receptor tyrosine kinases and Akt-dependent pathways
Deregulation of receptor tyrosine kinases is a common phenomenon in cancer cells. The epithelial growth factor receptor (EGFR) is frequently found overexpressed or mutated in several types of cancer including lung, breast, and brain. In some reports, EGFR seems to be involved in the acquisition of stemness properties in cancer cells.Citation113–Citation115 Previous reports, and our unpublished data, showed that activation of EGFR pathway, either through EGF treatment or ectopic overexpression of active EGFR, resulted in enhanced Nanog expression both at its protein and mRNA levels.Citation116,Citation117 This data may explain how EGFR regulates stem-like properties in cancers. Shan et al demonstrated that Nanog regulates selfrenewal of CSCs through the insulin-like growth factor receptor (IGF-1R) pathway in human hepatocellular carcinoma, and suppression of IGF-1R decreased the expression of Nanog in return.Citation33 The role of the phosphatidylinositide 3-kinase (PI3K) pathway in regulating Nanog expression is well known in ES cells.Citation118,Citation119 In lung adenocarcinoma cells, the IGF-1R activation leads to transcriptional activation of Nanog via the PI3K/Akt/GSK3/β-catenin cascade.Citation120 In mouse ES cells, loss of phosphatase and tensin homolog, a tumor suppressor gene acting through inhibiting the PI3K pathway, causes tumor initiation following differentiation because of failed repression of Nanog.Citation121 These data demonstrated a closely related regulation between stemness and oncogenic pathways and the crucial role of Nanog in this cross talk, rendering Nanog as a prospective target of cancer therapy.
FAK signaling
Nanog also cross talks with the focal adhesion kinase (FAK), which is known to play a significant role in tumor survival. Nanog directly binds to the promoter region of FAK and enhances its transcription, whereas FAK protein interacts with Nanog and mediates Nanog phosphorylation.Citation122 Phosphorylated Nanog is distributed in the nucleus and is essential to mediate tumor invasion in several cancer cells.Citation122
Nanog and cancer-stroma communication
There has been an accumulating interest in focusing on the role of the microenvironment surrounding cancer cells in regulating cancer progression and CSC property acquisition. We previously showed that the Nanog expression level in lung adenocarcinoma cells could be suppressed by cocultured bone marrow-derived mesenchymal stem cell (MSC) or its conditioned medium. This suppression is, at least in part, due to the secreted oncostatin M from MSC paracrinally inhibiting Nanog, Snail, and Slug in lung adenocarcinoma cells through a STAT1 (signal transducer and activator of transcription 1)-dependent pathway.Citation123 The suppressed Nanog, Snail, and Slug led to reduced cell motility in vitro and tumor metastasis in vivo. These data suggests that Nanog in cancer cells is able to receive extracellular signaling from cancer microenvironment.
Apart from promoting cancer stemness in cancer cells, Nanog was also found to mediate MSC and affect the MSC-dependent suppressive effect on tumor growth.Citation124 Zhu et alCitation124 showed that MSC inhibits cancer cell proliferation by secreting dickkopf-1. They further showed that Nanog binds to the promoter of dickkopf-1 in MSC and drives its transcription as well as increases its secretion. Therefore, MSCs with a high expression level of Nanog may possess antitumor ability. It has been suggested that the pluripotent status of MSC mediates its antitumor effect; MSC suppressed tumor cells only when they were precursors of matrix cells.Citation125 In agreement, the expression level of Nanog may determine the antitumor capacity of MSCs. Furthermore, Nanog was also reported to regulate the expression and secretion of TGF-β1, TGF-β2, and TGF-β3 in MSC.Citation126 These data suggest a role of Nanog in regulating the composition of paracrine factors in cancer microenvironments. Caution should be taken in terms of which cell is targeted when suppressing Nanog for cancer therapy.
Targeting Nanog in cancer cells
Targeting gene therapy is a promising therapeutic method to specifically fix the genetic disorder in diseases such as cancers. Through delivery of exogenous DNA or RNA sequence into host cells, we are able to enhance or suppress the expression of a selected gene to counteract the deregulated signaling pathway. Theoretically, there are several advantages to choose Nanog as a therapeutic target. First of all, Nanog cross talks with several well-known oncogenic pathways, which makes it a highly potential candidate to be targeted. Second, Nanog is supposedly not expressed in ordinary differentiated somatic cells; therefore, off-target effects of Nanog-targeting should be limited. Finally, according to the cellular functions mediated by Nanog downstream effectors, specific suppression of Nanog inhibits CSC functions, reduces tumor growth and metastasis, prevents the resistance to chemotherapeutic drugs, and enhances the immune surveillance of the host and apoptosis of the cancer.
There is an accumulation of evidence demonstrating the therapeutic potential of targeting Nanog in several cancer cells. For example, using RNAi-mediated Nanog knockdown, Jeter et al observed a reduced growth of subcutaneous xenograft tumors in vivo.Citation45 They pre-transfected shNanog in prostate, colon, and breast cancer cell lines before injecting the cells in NOD/SKID mice and found that the shNanog-transfected cells presented reduced long-term clonogenic growth, proliferation, and tumor size in mice. Similar phenomena were also observed in other reports with different cancer models, though how Nanog regulated cell proliferation and tumor growth had not been clear until cell cycle-related molecules, such as cyclin D1 and cyclin-dependent kinases, were identified as Nanog transcriptional targets in CSCs recently.Citation76,Citation77 Apart from reducing tumor growth, shNanog was also found to suppress tumor metastasis in a mouse model. Zhang et al showed that colorectal cancer cells transduced with shNanog failed to form grossly visible or microscopic hepatic liver colonies in mice, compared with parental cells.Citation31 They claimed that shRNA-mediated inhibition of Nanog gene expression is associated with both inhibition of experimental tumor growth and metastasis, as well as malignant phenotype and cancer stemness.
Targeting Nanog in cancer cells showed a synergistic therapeutic effect with chemotherapeutic drugs such as cisplatin. siRNA-mediated knockdown of Nanog increased the sensitivity of esophageal cancer cells to cisplatin treatment and induced cell apoptosis.Citation127 Although this effect still lacks in vivo evidence, it is not surprising that inhibition of Nanog would decrease drug resistance of cancers since Nanog has been linked to chemoresistance mechanisms through ABCB1 and p53.Citation32,Citation34,Citation85,Citation109 Furthermore, inhibition of Nanog in mice repressed the immune evasion of the tumor cells and increased the therapeutic effect of cancer vaccination. Delivery of siNanog into tumor-bearing mice increased tumor vulnerability to immune surveillance and suppressed tumor growth.Citation80 It seems that combining Nanog knockdown with current therapeutic methods, like cisplatin and cancer vaccination, would largely improve the therapeutic outcome in the mouse model.
There are still issues, however, that need to be clarified before further development of siNanog or shNanog as a therapeutic method. First, although experimental evidence has shown a promising therapeutic efficacy of Nanog knockdown in controlling tumor growth, metastasis, drug-resistance, and immune evasion in animal models, its efficacy in a model that is more close to clinical operations is still under investigation. For example, delivering siNanog or shNanog in tumor-bearing animals with appropriate delivery methods and reasonable doses would reveal how realistic it is to use Nanog-targeting as a cancer treatment. Second, the optimal delivery methods for siNanog or shNanog need to be identified. This would affect the side effects they may cause and the stability of delivered genes in cells. It has to be noted that adeno-associated virusCitation128 and nanoparticlesCitation129,Citation130 have been implicated as therapeutic delivery agents with promising efficacy. Moreover, conjugating nuclear localization sequence to the double-stranded DNA encoding for siNanog or shNanog could possibly enhance their nuclear delivery, as reported previously.Citation131 As well as choosing an appropriate delivery method, modification of the RNAi construct structure might also be a way to improve delivery efficacy. Third, being able to direct the Nanog RNAi to the cancer site would be a challenge. Whether to combine specific cancer or CSC markers as well as the selection of markers will need further evaluation. Finally, Nanog-targeting may be operated alone or in combination with current treatment methods such as chemotherapy and cancer vaccination. The synergistic effects still need to be validated in vivo.
Conclusion
Nanog determines the cell-fate both in ESC and CSC. Activating Nanog at an inappropriate time would cause CSC rather than ESC-like iPS. Failure to repress Nanog during the differentiation process in mouse pluripotent stem cells results in tumor initiation. Moreover, Nanog cross talks with several signal pathways both pro-oncogenic, such as PI3K and STAT3, and tumor-suppressive, such as p53 and PTEN (). Nanog also mediates the communication between cancer cells and their surrounding stromal and immune cells; Nanog expression level in both cancer and stromal cells determines the direction of the paracrine effects between them. These crucial roles of Nanog make it a center of cancer signaling networks () and a potentially ideal target for cancer treatment. In order to target Nanog in CSCs, developments of appropriate targeting methods, and an understanding of whether to combine with tissue- or CSC-specific markers, would be needed to avoid off-target side effects. It can be foreseen that the selection of suitable markers would be a challenge and it is necessary to investigate this selection. Current attempts of targeting Nanog in prostate, breast, and colorectal cancers have shown promising therapeutic effects in mouse models. In vivo data with experimental platforms that are more close to clinical operation are expected to support the therapeutic potential of Nanog in cancer treatment.
Figure 1 Nanog plays a central role in the cancer signaling network.
Acknowledgments
We thank Dr Hui-Wen Chen, professor in the Graduate Institute of Toxicology, National Taiwan University, for advice and suggestions during the preparation of the paper. This research was supported by the Department of Health (DOH101-TD-C-111-007) and the National Science Council (NSC101-2321-B-010-011, NSC102-2325-B-010-010), Executive Yuan, Taiwan, Republic of China.
Disclosure
The authors report no conflicts of interest in this work.
References
- WichaMSLiuSDontuGCancer stem cells: an old idea – a paradigm shiftCancer Res200666418831890 discussion 1895–188616488983
- VisvaderJELindemanGJCancer stem cells in solid tumours: accumulating evidence and unresolved questionsNat Rev Cancer200881075576818784658
- JiangFQiuQKhannaAAldehyde dehydrogenase 1 is a tumor stem cell-associated marker in lung cancerMol Cancer Res20097333033819276181
- LeeTKCastilhoACheungVCTangKHMaSNgIOCD24(+) liver tumor-initiating cells drive self-renewal and tumor initiation through STAT3-mediated NANOG regulationCell Stem Cell201191506321726833
- IbrahimEEBabaei-JadidiRSaadeddinAEmbryonic NANOG activity defines colorectal cancer stem cells and modulates through AP1- and TCF-dependent mechanismsStem Cells201230102076208722851508
- BaoSWuQMcLendonREGlioma stem cells promote radioresistance by preferential activation of the DNA damage responseNature2006444712075676017051156
- BertoliniGRozLPeregoPHighly tumorigenic lung cancer CD133+ cells display stem-like features and are spared by cisplatin treatmentProc Natl Acad Sci U S A200910638162811628619805294
- EramoALottiFSetteGIdentification and expansion of the tumorigenic lung cancer stem cell populationCell Death Differ200815350451418049477
- PrinceMESivanandanRKaczorowskiAIdentification of a subpopulation of cells with cancer stem cell properties in head and neck squamous cell carcinomaProc Natl Acad Sci U S A2007104397397817210912
- BussolatiBBrunoSGrangeCFerrandoUCamussiGIdentification of a tumor-initiating stem cell population in human renal carcinomasFASEB J200822103696370518614581
- SinghSKHawkinsCClarkeIDIdentification of human brain tumour initiating cellsNature2004432701539640115549107
- BonnetDDickJEHuman acute myeloid leukemia is organized as a hierarchy that originates from a primitive hematopoietic cellNat Med1997377307379212098
- Al-HajjMWichaMSBenito-HernandezAMorrisonSJClarkeMFProspective identification of tumorigenic breast cancer cellsProc Natl Acad Sci U S A200310073983398812629218
- GalliRBindaEOrfanelliUIsolation and characterization of tumorigenic, stem-like neural precursors from human glioblastomaCancer Res200464197011702115466194
- HemmatiHDNakanoILazareffJACancerous stem cells can arise from pediatric brain tumorsProc Natl Acad Sci U S A200310025151781518314645703
- LapidotTSirardCVormoorJA cell initiating human acute myeloid leukaemia after transplantation into SCID miceNature199436764646456487509044
- O’BrienCAPollettAGallingerSDickJEA human colon cancer cell capable of initiating tumour growth in immunodeficient miceNature2007445712310611017122772
- Ricci-VitianiLLombardiDGPilozziEIdentification and expansion of human colon-cancer-initiating cellsNature2007445712311111517122771
- SchattonTMurphyGFFrankNYIdentification of cells initiating human melanomasNature2008451717634534918202660
- SinghSKClarkeIDTerasakiMIdentification of a cancer stem cell in human brain tumorsCancer Res200363185821582814522905
- FillmoreCMKuperwasserCHuman breast cancer cell lines contain stem-like cells that self-renew, give rise to phenotypically diverse progeny and survive chemotherapyBreast Cancer Res2008102R2518366788
- WakamatsuYSakamotoNOoHZExpression of cancer stem cell markers ALDH1, CD44 and CD133 in primary tumor and lymph node metastasis of gastric cancerPathol Int201262211211922243781
- TirinoVCamerlingoRFrancoRThe role of CD133 in the identification and characterisation of tumour-initiating cells in non-small-cell lung cancerEur J Cardiothorac Surg200936344645319464919
- SalvenPMustjokiSAlitaloRAlitaloKRafiiSVEGFR-3 and CD133 identify a population of CD34+ lymphatic/vascular endothelial precursor cellsBlood2003101116817212393704
- YinAHMiragliaSZanjaniEDAC133, a novel marker for human hematopoietic stem and progenitor cellsBlood19979012500250129389720
- SaigusaSTanakaKToiyamaYCorrelation of CD133, OCT4, and SOX2 in rectal cancer and their association with distant recurrence after chemoradiotherapyAnn Surg Oncol200916123488349819657699
- DasBTsuchidaRMalkinDKorenGBaruchelSYegerHHypoxia enhances tumor stemness by increasing the invasive and tumorigenic side population fractionStem Cells20082671818183018467664
- HoMMNgAVLamSHungJYSide population in human lung cancer cell lines and tumors is enriched with stem-like cancer cellsCancer Res200767104827483317510412
- JiangYHeYLiHExpressions of putative cancer stem cell markers ABCB1, ABCG2, and CD133 are correlated with the degree of differentiation of gastric cancerGastric cancer201215444045022395309
- FangDNguyenTKLeishearKA tumorigenic subpopulation with stem cell properties in melanomasCancer Res200565209328933716230395
- ZhangJEspinozaLAKindersRJNANOG modulates stemness in human colorectal cancerOncogene Epub10222012
- ZbindenMDuquetALorente-TrigosANgwabytSNBorgesIRuiz i AltabaANANOG regulates glioma stem cells and is essential in vivo acting in a cross-functional network with GLI1 and p53EMBO J201029152659267420581802
- ShanJShenJLiuLNanog regulates self-renewal of cancer stem cells through the insulin-like growth factor pathway in human hepatocellular carcinomaHepatology20125631004101422473773
- ChiouSHWangMLChouYTCoexpression of oct4 and nanog enhances malignancy in lung adenocarcinoma by inducing cancer stem cell-like properties and epithelial-mesenchymal transdifferentiationCancer Res20107024104331044421159654
- MitsuiKTokuzawaYItohHThe homeoprotein Nanog is required for maintenance of pluripotency in mouse epiblast and ES cellsCell2003113563164212787504
- ChambersIColbyDRobertsonMFunctional expression cloning of Nanog, a pluripotency sustaining factor in embryonic stem cellsCell2003113564365512787505
- HyslopLStojkovicMArmstrongLDownregulation of NANOG induces differentiation of human embryonic stem cells to extraembryonic lineagesStem Cells20052381035104315983365
- WangJRaoSChuJA protein interaction network for pluripotency of embryonic stem cellsNature2006444711736436817093407
- KashyapVRezendeNCScotlandKBRegulation of stem cell pluripotency and differentiation involves a mutual regulatory circuit of the NANOG, OCT4, and SOX2 pluripotency transcription factors with polycomb repressive complexes and stem cell microRNAsStem Cells Dev20091871093110819480567
- WangJLevasseurDNOrkinSHRequirement of Nanog dimerization for stem cell self-renewal and pluripotencyProc Natl Acad Sci U S A2008105176326633118436640
- PanGThomsonJANanog and transcriptional networks in embryonic stem cell pluripotencyCell Res2007171424917211451
- BoothHAHollandPWEleven daughters of NANOGGenomics200484222923815233988
- ZhangJWangXLiMNANOGP8 is a retrogene expressed in cancersFEBS J200627381723173016623708
- FairbanksDJFairbanksADOgdenTHParkerGJMaughanPJNANOGP8: evolution of a human-specific retro-oncogeneG3 (Bethesda)20122111447145723173096
- JeterCRBadeauxMChoyGFunctional evidence that the self-renewal gene NANOG regulates human tumor developmentStem Cells2009275993100519415763
- OhJHDoHJYangHMIdentification of a putative transactivation domain in human NanogExp Mol Med200537325025416000880
- DoHJLimHYKimJHSongHChungHMAn intact homeobox domain is required for complete nuclear localization of human NanogBiochem Biophys Res Commun2007353377077517196939
- ParkSWDoHJHuhSHIdentification of a putative nuclear export signal motif in human NANOG homeobox domainBiochem Biophys Res Commun2012421348448922516749
- ParkIHZhaoRWestJAReprogramming of human somatic cells to pluripotency with defined factorsNature2008451717514114618157115
- TheunissenTWCostaYRadzisheuskayaAReprogramming capacity of Nanog is functionally conserved in vertebrates and resides in a unique homeodomainDevelopment2011138224853486522028025
- KurodaTTadaMKubotaHOctamer and Sox elements are required for transcriptional cis regulation of Nanog gene expressionMol Cell Biol20052562475248515743839
- DoHJLeeWYLimHYTwo potent transactivation domains in the C-terminal region of human NANOG mediate transcriptional activation in human embryonic carcinoma cellsJ Cell Biochem200910661079108919229867
- WilliamsRLHiltonDJPeaseSMyeloid leukaemia inhibitory factor maintains the developmental potential of embryonic stem cellsNature198833662006846873143916
- NiwaHMiyazakiJSmithAGQuantitative expression of Oct-3/4 defines differentiation, dedifferentiation or self-renewal of ES cellsNat Genet200024437237610742100
- AmsterdamARaananCSchreiberLFreyhanOSchechtmanLGivolDLocalization of the stem cell markers LGR5 and Nanog in the normal and the cancerous human ovary and their inter-relationshipActa Histochem2013115433033823092806
- EzehUITurekPJReijoRAClarkATHuman embryonic stem cell genes OCT4, NANOG, STELLAR, and GDF3 are expressed in both seminoma and breast carcinomaCancer2005104102255226516228988
- HartAHHartleyLParkerKThe pluripotency homeobox gene NANOG is expressed in human germ cell tumorsCancer2005104102092209816206293
- Hoei-HansenCEAlmstrupKNielsenJEStem cell pluripotency factor NANOG is expressed in human fetal gonocytes, testicular carcinoma in situ and germ cell tumoursHistopathology2005471485615982323
- LinTDingYQLiJMOverexpression of Nanog protein is associated with poor prognosis in gastric adenocarcinomaMed Oncol201229287888521336986
- GuoYLiuSWangPExpression profile of embryonic stem cell-associated genes Oct4, Sox2 and Nanog in human gliomasHistopathology201159476377522014056
- PanYJiaoJZhouCChengQHuYChenHNanog is highly expressed in ovarian serous cystadenocarcinoma and correlated with clinical stage and pathological gradePathobiology201077628328821266826
- WenJParkJYParkKHOct4 and Nanog expression is associated with early stages of pancreatic carcinogenesisPancreas201039562262620173672
- ChiouSHYuCCHuangCYPositive correlations of Oct-4 and Nanog in oral cancer stem-like cells and high-grade oral squamous cell carcinomaClin Cancer Res200814134085409518593985
- GillisAJStoopHBiermannKExpression and interdependencies of pluripotency factors LIN28, OCT3/4, NANOG and SOX2 in human testicular germ cells and tumours of the testisInt J Androl2011344 Pt 2e160e17421631526
- ZhouXZhouYPHuangGRExpression of the stem cell marker, Nanog, in human endometrial adenocarcinomaInt J Gynecol Pathol201130326227021464727
- NagataTShimadaYSekineSPrognostic significance of NANOG and KLF4 for breast cancerBreast cancer2012
- LeeMNamEJKimSWKimSKimJHKimYTPrognostic impact of the cancer stem cell-related marker NANOG in ovarian serous carcinomaInt J Gynecol Cancer20122291489149623095773
- MengHMZhengPWangXYOverexpression of nanog predicts tumor progression and poor prognosis in colorectal cancerCancer Biol Ther201094
- NiuCSLiDXLiuYHFuXMTangSFLiJExpression of NANOG in human gliomas and its relationship with undifferentiated glioma cellsOncol Rep201126359360121573506
- JeterCRLiuBLiuXNANOG promotes cancer stem cell characteristics and prostate cancer resistance to androgen deprivationOncogene201130363833384521499299
- PoAFerrettiEMieleEHedgehog controls neural stem cells through p53-independent regulation of NanogEMBO J201029152646265820581804
- VillasanteAPiazzollaDLiHGomez-LopezGDjabaliMSerranoMEpigenetic regulation of Nanog expression by Ezh2 in pluripotent stem cellsCell Cycle20111091488149821490431
- XuFDaiCZhangRZhaoYPengSJiaCNanog: a potential biomarker for liver metastasis of colorectal cancerDig Dis Sci20125792340234622562535
- HeAQiWHuangYCD133 expression predicts lung metastasis and poor prognosis in osteosarcoma patients: A clinical and experimental studyExp Ther Med20124343544123181114
- LeungELFiscusRRTungJWNon-small cell lung cancer cells expressing CD44 are enriched for stem cell-like propertiesPLoS One2010511e1406221124918
- HanJZhangFYuMRNA interference-mediated silencing of NANOG reduces cell proliferation and induces G0/G1 cell cycle arrest in breast cancer cellsCancer Lett20123211808822381696
- ChoiSCChoiJHParkCYAhnCMHongSJLimDSNanog regulates molecules involved in stemness and cell cycle-signaling pathway for maintenance of pluripotency of P19 embryonal carcinoma stem cellsJ Cell Physiol2012227113678369222378194
- SiuMKWongESKongDSStem cell transcription factor NANOG controls cell migration and invasion via dysregulation of E-cadherin and FoxJ1 and contributes to adverse clinical outcome in ovarian cancersOncogene201332303500350922945654
- NohKHKimBWSongKHNanog signaling in cancer promotes stem-like phenotype and immune evasionJ Clin Invest2012122114077409323093782
- NohKHLeeYHJeonJHCancer vaccination drives Nanog-dependent evolution of tumor cells toward an immune-resistant and stem-like phenotypeCancer Res20127271717172722337995
- GuTTLiuSYZhengPSCytoplasmic NANOG-positive stromal cells promote human cervical cancer progressionAm J Pathol2012181265266122683467
- KalmarTLimCHaywardPRegulated fluctuations in nanog expression mediate cell fate decisions in embryonic stem cellsPLoS Biol200977e100014919582141
- MacArthurBDSevillaALenzMNanog-dependent feedback loops regulate murine embryonic stem cell heterogeneityNat Cell Biol201214111139114723103910
- GolubovskayaVMFAK and Nanog cross talk with p53 in cancer stem cellsAnticancer Agents Med Chem201313457658022934707
- LinTChaoCSaitoSp53 induces differentiation of mouse embryonic stem cells by suppressing Nanog expressionNat Cell Biol20057216517115619621
- MoonJHKwonSJunEKNanog-induced dedifferentiation of p53-deficient mouse astrocytes into brain cancer stem-like cellsBiochem Biophys Res Commun2011412117518121810410
- HongHTakahashiKIchisakaTSuppression of induced pluripotent stem cell generation by the p53-p21 pathwayNature200946072591132113519668191
- YuJVodyanikMASmuga-OttoKInduced pluripotent stem cell lines derived from human somatic cellsScience200731858581917192018029452
- ClementVSanchezPde TriboletNRadovanovicIRuiz i AltabaAHEDGEHOG-GLI1 signaling regulates human glioma growth, cancer stem cell self-renewal, and tumorigenicityCurr Biol200717216517217196391
- BourillotPYAksoyISchreiberVNovel STAT3 target genes exert distinct roles in the inhibition of mesoderm and endoderm differentiation in cooperation with NanogStem Cells20092781760177119544440
- TorresJWattFMNanog maintains pluripotency of mouse embryonic stem cells by inhibiting NFkappaB and cooperating with Stat3Nat Cell Biol200810219420118223644
- BourguignonLYEarleCWongGSpevakCCKruegerKStem cell marker (Nanog) and Stat-3 signaling promote MicroRNA-21 expression and chemoresistance in hyaluronan/CD44-activated head and neck squamous cell carcinoma cellsOncogene201231214916021685938
- SuzukiARayaAKawakamiYNanog binds to Smad1 and blocks bone morphogenetic protein-induced differentiation of embryonic stem cellsProc Natl Acad Sci U S A200610327102941029916801560
- TangYLuoYJiangZJak/Stat3 signaling promotes somatic cell reprogramming by epigenetic regulationStem Cells201230122645265622968989
- HawkinsKMohametLRitsonSMerryCLWardCME-cadherin and, in its absence, N-cadherin promotes Nanog expression in mouse embryonic stem cells via STAT3 phosphorylationStem Cells20123091842185122696497
- TayYZhangJThomsonAMLimBRigoutsosIMicroRNAs to Nanog, Oct4 and Sox2 coding regions modulate embryonic stem cell differentiationNature200845572161124112818806776
- TayYMTamWLAngYSMicroRNA-134 modulates the differentiation of mouse embryonic stem cells, where it causes post-transcriptional attenuation of Nanog and LRH1Stem Cells2008261172917916804
- NiuCSYangYChengCDMiR-134 regulates the proliferation and invasion of glioblastoma cells by reducing Nanog expressionInt J Oncol20134251533154023467648
- PennaEOrsoFCiminoDmicroRNA-214 contributes to melanoma tumour progression through suppression of TFAP2CEMBO J201130101990200721468029
- YangHKongWHeLMicroRNA expression profiling in human ovarian cancer: miR-214 induces cell survival and cisplatin resistance by targeting PTENCancer Res200868242543318199536
- XuCXXuMTanLMicroRNA miR-214 regulates ovarian cancer cell stemness by targeting p53/NanogJ Biol Chem201228742349703497822927443
- FarehMTurchiLVirolleVThe miR 302-367 cluster drastically affects self-renewal and infiltration properties of glioma-initiating cells through CXCR4 repression and consequent disruption of the SHH-GLI-NANOG networkCell Death Differ201219223224421720384
- PiestunDKochupurakkalBSJacob-HirschJNanog transforms NIH3T3 cells and targets cell-type restricted genesBiochem Biophys Res Commun2006343127928516540082
- LinYLHanZBXiongFYMalignant transformation of 293 cells induced by ectopic expression of human NanogMol Cell Biochem20113511–210911621246261
- ParkSWLimHYDoHJRegulation of human growth and differentiation factor 3 gene expression by NANOG in human embryonic carcinoma NCCIT cellsFEBS Lett2012586193529353522963770
- YinXLiYWZhangBHCoexpression of stemness factors Oct4 and Nanog predict liver resectionAnn Surg Oncol20121992877288722461131
- YangLZhangXZhangMIncreased nanog expression promotes tumor development and Cisplatin resistance in human esophageal cancer cellsCell Physiol Biochem201230494395223221432
- TsaiLLYuCCChangYCYuCHChouMYMarkedly increased Oct4 and Nanog expression correlates with cisplatin resistance in oral squamous cell carcinomaJ Oral Pathol Med201140862162821342274
- BourguignonLYPeyrollierKXiaWGiladEHyaluronan-CD44 interaction activates stem cell marker Nanog, Stat-3-mediated MDR1 gene expression, and ankyrin-regulated multidrug efflux in breast and ovarian tumor cellsJ Biol Chem200828325176351765118441325
- XuYA new role for p53 in maintaining genetic stability in embryonic stem cellsCell Cycle20054336336415701975
- van den BergDLZhangWYatesAEstrogen-related receptor beta interacts with Oct4 to positively regulate Nanog gene expressionMol Cell Biol200828195986599518662995
- FestucciaNOsornoRHalbritterFEsrrb is a direct Nanog target gene that can substitute for Nanog function in pluripotent cellsCell Stem Cell201211447749023040477
- AbholdELKiangARahimyEEGFR kinase promotes acquisition of stem cell-like properties: a potential therapeutic target in head and neck squamous cell carcinoma stem cellsPLoS One201272e3245922384257
- YangJLiaoDChenCTumor-associated macrophages regulate murine breast cancer stem cells through a novel paracrine EGFR/Stat3/Sox-2 signaling pathwayStem Cells201331224825823169551
- YiDKuoSZZhengHActivation of PDGFR and EGFR promotes the acquisition of a stem cell-like phenotype in schwannomasOtol Neurot201233916401647
- MaLZhangGMiaoXBCancer stem-like cell properties are regulated by EGFR/AKT/beta-catenin signaling and preferentially inhibited by gefitinib in nasopharyngeal carcinomaFEBS J201328092027204123461856
- HyderAEhnertSHinzHNusslerAKFandrichFUngefrorenHEGF and HB-EGF enhance the proliferation of programmable cells of monocytic origin (PCMO) through activation of MEK/ERK signaling and improve differentiation of PCMO-derived hepatocyte-like cellsCell Commun Signal20121012322873932
- WatanabeSUmeharaHMurayamaKOkabeMKimuraTNakanoTActivation of Akt signaling is sufficient to maintain pluripotency in mouse and primate embryonic stem cellsOncogene200625192697270716407845
- StormMPBoneHKBeckCGRegulation of Nanog expression by phosphoinositide 3-kinase-dependent signaling in murine embryonic stem cellsJ Biol Chem200728296265627317204467
- XuCXieDYuSCbeta-Catenin/POU5F1/SOX2 transcription factor complex mediates IGF-I receptor signaling and predicts poor prognosis in lung adenocarcinomaCancer Res201373103181318923539445
- LindgrenAGNatsuharaKTianELoss of Pten causes tumor initiation following differentiation of murine pluripotent stem cells due to failed repression of NanogPLoS One201161e1647821304588
- HoBOlsonGFigelSGelmanICanceWGGolubovskayaVMNanog increases focal adhesion kinase (FAK) promoter activity and expression and directly binds to FAK protein to be phosphorylatedJ Biol Chem201228722186561867322493428
- WangMLPanCMChiouSHOncostatin m modulates the mesenchymal-epithelial transition of lung adenocarcinoma cells by a mesenchymal stem cell-mediated paracrine effectCancer Res201272226051606423139208
- ZhuYSunZHanQHuman mesenchymal stem cells inhibit cancer cell proliferation by secreting DKK-1Leukemia200923592593319148141
- OrimoAGuptaPBSgroiDCStromal fibroblasts present in invasive human breast carcinomas promote tumor growth and angiogenesis through elevated SDF-1/CXCL12 secretionCell2005121333534815882617
- SunZHanQZhuYNANOG has a role in mesenchymal stem cells’ immunomodulatory effectStem Cells Dev20112091521152821235326
- DuYShiLWangTLiuZWangZNanog siRNA plus Cisplatin may enhance the sensitivity of chemotherapy in esophageal cancerJ Cancer Res Clin Oncol2012138101759176722714588
- ShaoWPaulAAbbasiSA novel polyethyleneimine-coated adeno-associated virus-like particle formulation for efficient siRNA delivery in breast cancer therapy: preparation and in vitro analysisInt J Nanomedicine201271575158622619514
- ChiouGYCherngJYHsuHSCationic polyurethanes-short branch PEI-mediated delivery of Mir145 inhibited epithelial-mesenchymal transdifferentiation and cancer stem-like properties and in lung adenocarcinomaJ Control Release2012159224025022285547
- PengCHCherngJYChiouGYDelivery of Oct4 and SirT1 with cationic polyurethanes-short branch PEI to aged retinal pigment epitheliumBiomaterials201132349077908821890195
- LoWLChienYChiouGYNuclear localization signal-enhanced RNA interference of EZH2 and Oct4 in the eradication of head and neck squamous cell carcinoma-derived cancer stem cellsBiomaterials201233143693370922361100
- MachidaKTsukamotoHMkrtchyanHToll-like receptor 4 mediates synergism between alcohol and HCV in hepatic oncogenesis involving stem cell marker NanogProc Natl Acad Sci U S A200910651548155319171902
- HeddlestonJMLiZMcLendonREHjelmelandABRichJNThe hypoxic microenvironment maintains glioblastoma stem cells and promotes reprogramming towards a cancer stem cell phenotypeCell Cycle20098203274328419770585
- KimJSKimBSKimJParkCSChungIYThe phosphoinositide-3-kinase/Akt pathway mediates the transient increase in Nanog expression during differentiation of F9 cellsArch Pharm Res20103371117112520661723
- TsaiCCSuPFHuangYFYewTLHungSCOct4 and Nanog directly regulate Dnmt1 to maintain self-renewal and undifferentiated state in mesenchymal stem cellsMol Cell201247216918222795133