
Abstract
Protonography, a sodium dodecyl sulfate – polyacrylamide gel electrophoresis (SDS-PAGE) technique derived from zymography was recently reported by our group to be an effective, cheap and reproducible technique for evidencing catalytically active α-carbonic anhydrase (CA, EC 4.2.1.1) isoforms, such as the bovine red blood cell isoform bCA or the bacterial enzyme from Vibrio cholerae, VchCA. CA activity was also observed on the protonogram of a cellular extract of Escherichia coli, evidencing the presence of one or more β-class such enzymes. Here we show that protonography can also be applied to the recently discovered η-CA family using the Plasmodium falciparum enzyme PfCA as an example. The protonogram of PfCA clearly showed catalytically active η-CA with a specific band at 22.0 kDa, which was quite distinct from the band of the red blood cell bovine enzyme bCA, which was observed at 28.8 kDa. The different migration pattern of α- and η-CAs might be a useful tool to detect Plasmodium falciparum in infected human red blood cells by an easy, routine inexpensive technique.
Introduction
Carbonic anhydrases (CAs, CA, EC 4.2.1.1) catalyze a simple reaction in all life domains: the carbon dioxide hydration to bicarbonate and protons: Citation1–4. These enzymes are thus involved in many physiologic processes, such as photosynthesis, respiration, CO2 transport, as well as metabolism of xenobiotics (e.g. cyanate in Escherichia coli)Citation1–7. Six different, genetically distinct CA families are known to date, the α-, β-, γ-, δ-, ζ- and η-CAsCitation8–11. The α-, β-, δ-and η-CAs use Zn(II) ions at the active site, the γ-CAs are probably Fe(II) enzymes (but they are active also with bound Zn(II) or Co(II) ions), whereas the ζ-class uses Cd(II) or Zn(II) to perform the physiologic reaction catalysisCitation5,Citation11–28. Many representatives of all these enzyme classes have been crystallized and characterized in detail, except δ- and η-CAs. The three-dimensional analysis of CAs present a substantial variability between the different classes: the overall shape of the molecules, the protein folding patterns as well as the oligomeric organization of the six genetic CA families are very much differentCitation5,Citation11–28. For example, α-CAs are normally monomers and rarely dimmers; β-CAs are dimers, tetramers or octamers; γ-CAs are trimers, whereas the δ- and ζ-CAs are less well understood. The only ζ-CA crystallized so far has three slightly different active sites on the same polypeptide chain, whereas X-ray crystal structures of δ- and η-CAs are not available so farCitation12–14,Citation29–35.
Many classes of CA inhibitors (CAIs) are known to date: the metal complexing anions and the unsubstituted sulfonamides which bind to the Zn(II) ion of the enzyme either by substituting the non-protein zinc ligand or add to the metal coordination sphere, generating trigonal–bipyramidal species are the classical, are the most frequently investigated onesCitation15,Citation36–52. In addition to them, coumarins, phenols, dithiocarbamates and xanthates as well as polyamines, are new classes of CAIs reported in the last years by these groupsCitation9,Citation53–63.
The η-CA class, recently discovered in our laboratories, was identified in the genome of the protozoan parasite Plasmodium falciparum, the causative agent of the most lethal type of human malaria, and in that of other Plasmodia. The enzyme identified in P. falciparum was indicated with the cryptonym PfCACitation11. We demonstrated that PfCA has significant CO2 hydrase activity, with a kcat of 1.4 × 105 s−1 and a kcat/KM of 5.4 × 106 M−1 s−1. Moreover, through an extensive inhibitory study we detected numerous effective inhibitors such as inorganic and complex anions and other molecules known to interact with zinc proteins, including sulfamide, sulfamic acid, phenylboronic/arsonic acids and sulfonamidesCitation11.
Recently, our group reported a new technique to assay CA activity, named “protonography”, similar to zymography and derived from it. However, unlike zymography, protonography allows the detection of the activity of α-CAs on a sodium dodecyl sulfate – polyacrylamide gel electrophoresis (SDS-PAGE) gel, as showed in our original workCitation27. The technique is based on monitoring the pH variation in the gel (protonogram) as a consequence of the CA-catalyzed conversion of CO2 to bicarbonate and protons. The protonogram is then stained with bromothymol blue, which is an indicator of pH variations. This dye appears blue in its deprotonated form, while its color changes to yellow in the protonated form. Thus, the production of hydrogen ions during the CO2 hydration reaction, due to the hydratase activity of the enzyme lowers the pH of the solution/gel until the color transition point of the dye is reached (pH 6.8). The yellow band on the gel corresponds to the CA position on the protonogramCitation27. The time required for the color change is inversely related to the quantity of carbonic anhydrase, which is responsible for dropping the pH from 8.2 to the transition point of the dye in a control buffer. The success of this technique is that the hydratase activity of the α-CAs was reversibly inhibited by SDS during electrophoresis and recovered by incubating the gel in aqueous Triton X-100.
In the present paper, we demonstrate that the protonography technique can be used to detect the η-CA activity of PfCA on a SDS-PAGE gel also, allowing us to prove that this technique may be extended to enzymes belonging to other CA classes than the α one.
Materials and methods
Mammalian bCA
Bovine α-CA (bCA) was supplied by Sigma-Aldrich (St. Louis, MO) as a crude extract from bovine erythrocytes (lyophilized powder).
Expression and purification of PfCA
A truncated form of the P. falciparum CA gene (PlasmoDB: PF3D7_1140000) encoding for the η-CA, PfCA (amino acid residues from 211 to 445)Citation64 and contained into the expression vector pET-43.1a, was kindly donated by Prof. Sally-Ann Poulsen, Eskitis Institute, Griffith University, Brisbane, Australia. Arctic Express competent cells (Agilent Technologies, La Jolla, CA) were transformed with pET43.1a/PfCA expression vector. Several transformant colonies were picked and grown overnight in LB containing 20 μg/ml of gentamicin and 100 μg/ml of ampicillin for selecting the expression plasmid at 37 °C. Then, cells were grown without antibiotic selection at 30 °C and induced at an A600 of 0.6–1 with 1 mM isopropylthiogalactopyranoside (IPTG). Zn(SO4) was added after 30 min. After additional growth for 16 h at 20 °C, cells were harvested, resuspended in 10 mM buffer Tris/HCl, pH 8.3 and disrupted by sonication at 4 °C. Following centrifugation, soluble protein extract was loaded onto a His-select HF nickel affinity gel column (GE Healthcare), then washed with five column volumes with His binding buffer (20 mM NaH2PO4, 500 mM NaCl, 20 mM Imidazole, pH 8). The His-tagged protein was eluted from the column with elution buffer (20 mM NaH2PO4, 500 mM NaCl, 250 mM Imidazole, pH 8). The purified recombinant PfCA was dialyzed over-night against 10 mM buffer Tris/HCl, pH 8.3.
CA assay
An applied photophysics stopped-flow instrument was used for assaying the CA catalyzed CO2 hydration activityCitation65. Phenol red (at 0.2 mM) was used as indicator, working at the absorbance maximum of 557 nm with 10 mM Hepes (pH 7.5) as buffer and 0.1 M NaClO4 (for maintaining constant ionic strength), at 20 °C, following the CA-catalyzed CO2 hydration reaction for a period of 10–100 s (the uncatalyzed reaction needs around 60–100 s in the assay conditions, whereas the catalyzed ones are of around 6–10 s). The CO2 concentrations ranged from 1.7 to 17 mM for the determination of the kinetic parameters. The uncatalyzed rates were determined in the same manner and subtracted from the total observed rates. Enzyme concentrations in the assay system were 9.1 nM for bCA and 8.6 nM for VchCA.
Protonography
SDS-PAGE was performed as described by LaemmliCitation66. Briefly, wells of 12% SDS-gel were loaded with bCA or PfCA mixed with Laemmli loading buffer without 2-mercaptoethanol and without boiling the samples, in order to avoid protein denaturation. The gel was run at 180 V until the dye front ran off the gelCitation27. Following the electrophoresis, the 12% SDS-gel was subject to protonography to detect the bCA and PfCA hydratase activity on the gel as described by Capasso and coworkersCitation27.
Results and discussion
Recombinant PfCA was recovered in the soluble fraction of the Arctic Express cell extract after sonication and centrifugation as describes in the “Experimental section”. PfCA was produced as fusion proteins containing an N-terminal His-Tag coding sequence. The recombinant fusion protein was purified by affinity chromatography on a column of HisTrap FF, prepacked with precharged Ni Sepharose 6 Fast Flow (GE Healthcare, Umeå, Sweden). Chromatography was performed on FPLC System. The (His)6-tagged PfCA protein eluted at 250 mM imidazole was recovered at approximately 90% purity.
A phylogenetic tree was constructed for showing that the Plasmodia CAs form a distinct group noticeably different from α-CAs (). The CA class, organism, accession numbers and cryptonyms of the amino acid sequences used to construct the phylogenetic tree are reported in .
Figure 1. Phylogenetic tree constructed using amino acid sequences of the α- and η-CAs from selected prokaryotic and eukaryotic species. The tree was generated using the program PhyML 3.0 (Montpellier (France) Bioinformatic platform). Class, organisms, accession numbers and cryptonyms of the sequences used in the alignment have been indicated in .
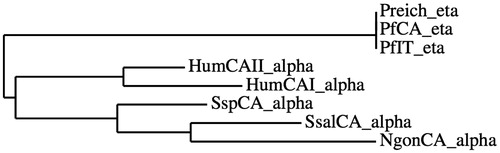
Table 1. CA class, organism, accession numbers and cryptonyms of the amino acid sequences used to construct the phylogenetic tree.
A stopped-flow CO2 hydrase assay has been used to measure the catalytic activity of PfCA, which was compared with those of other CAs belonging to a different CA classes. shows the catalytic activity of these enzymes for the CO2 hydration reaction. It may be observed that PfCA has kinetic parameters quite similar to those of the β- and γ-CAs.
Table 2. Kinetic parameters for CO2 hydration reaction catalyzed by PfCA and other CA classes at 20 °C and pH 7.5.
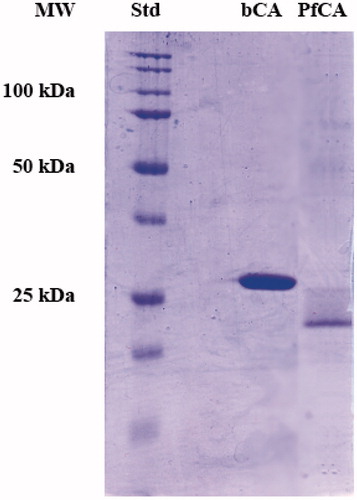
To investigate the hydratase activity of η-CA on the SDS-Page gel, 300 ng of PfCA or bCA were loaded on the gel. PfCA molecular weight estimated by SDS-PAGE carried out without 2-mercaptoethanol and without boiling the samples was 22.0 kDa, while bCA showed a mass of 28.8 kDa (). A subunit molecular mass of 30 kDa was calculated on the basis of PfCA and bCA amino acid sequences. Molecular mass difference between the theoretical molecular weights and those determined by SDS-Page is due to the fact that SDS-Page was carried out without 2-mercaptoethanol and without boiling samples to avoid protein denaturation, which is essential to follow the protonography (as explained in the “Materials and methods” section).
Figure 2. SDS-PAGE of bCA and PfCA. The gel was stained with Coomassie blue and run under denaturing but non-reducing conditions (see text for details). Gels were loaded with 300 ng/well of bCA and PfCA.
The protonography monitors the pH variation in the gel (protonogram) as a function of the catalyzed conversion of CO2 to bicarbonate and protons. The protonogram is stained with bromothymol blue as pH indicator. Thus, it monitors the production of hydrogen ions during the CO2 hydration reaction, due to the bCA or PfCA hydratase activity, which lowered the pH of the solution until the color transition point of the dye has been reached (pH 6.8). The protonogram obtained using the η-CA (PfCA) and the α-CA (bCA) is shown in . The yellow band corresponds to the individual CA position on the gel. The hydratase activity of η-CA can be reversibly inhibited by SDS during electrophoresis and recovered by incubating the gel in aqueous Triton X-100, as already demonstrated for α-CAsCitation27.
Figure 3. Protonogram of bCA and PfCA. The yellow bands on blue background correspond to the bCA and PfCA activity, which determine the drop of pH from 8.2 to the transition point of the dye (pH 6.8). Yellow bands were obtained with an incubation time of 15 s.
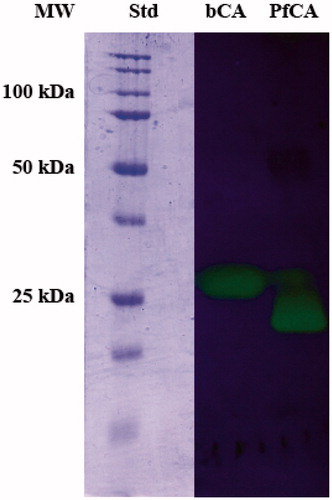
It is interesting to note a difference in the migration of bCA and PfCA on the SDS-PAGE and protonogram carried out without 2-mercaptoethanol and without boiling the samples, although both proteins possess a theoretical molecular weight of 30 kDa. This is probably due to the very different shape of the two proteins which although having the same molecular weight, belong to two different CA classes which is another proof of the relevant difference between the α and η-CA genetic families.
We also hypothesize that the different migration pattern of α-CA (bCA) and η-CA (PfCA) might be a useful tool to detect the presence Plasmodium falciparum, the agent responsible for malaria, in infected human red blood cells. In fact, the η-CA class is produced by the Plasmodium falciparum proliferation and is absent in the non-infected human erythrocytes, which are characterized by the presence of high amounts of α-CAs (hCA I and hCA II). Work is in progress to test this hypothesis.
Conclusions
Protonography, an SDS-PAGE technique similar to zymography was recently reported by our group to be an effective, cheap and reproducible technique for evidencing catalytically active α-CA isoforms, such as the bovine red blood cell isoform bCA or the bacterial enzyme from Vibrio cholerae, VchCACitation27. In the original paper we also observed CA activity on the protonogram of a cellular extract of E. coli, evidencing the presence of one or more β-class such enzymes. Here we show that protonography can be applied to the recently discovered η-CA family, by using the P. falciparum enzyme PfCA as an example. The protonogram of PfCA clearly showed catalytically active η-CA with a specific band at 22.0 kDa, which was quite distinct from the band of the red blood cell bovine enzyme bCA, which was observed at 28.8 kDa. The different migration pattern of α- and η-CA might be a useful tool to detect Plasmodium falciparum in infected human red blood cells.
Declaration of interest
The authors declare no conflict of interest.
References
- Supuran CT. Carbonic anhydrases: novel therapeutic applications for inhibitors and activators. Nat Rev Drug Discov 2008;7:168–81
- Supuran CT. Bacterial carbonic anhydrases as drug targets: toward novel antibiotics? Front Pharmacol 2011;2:34
- Capasso C, Supuran CT. An overview of the alpha-, beta- and gamma-carbonic anhydrases from Bacteria: can bacterial carbonic anhydrases shed new light on evolution of bacteria? J Enzyme Inhib Med Chem 2014. [Epub ahead of print]. DOI: 10.3109/14756366.2014.910202
- Capasso C, Supuran CT. An overview of the selectivity and efficiency of the bacterial carbonic anhydrase inhibitors. Curr Med Chem 2014. [Epub ahead of print]
- Migliardini F, De Luca V, Carginale V, et al. Biomimetic CO2 capture using a highly thermostable bacterial alpha-carbonic anhydrase immobilized on a polyurethane foam. J Enzyme Inhib Med Chem 2014;29:146–50
- Ceruso M, Del Prete S, Alothman Z, et al. Sulfonamides with potent inhibitory action and selectivity against the alpha-carbonic anhydrase from Vibrio cholerae. ACS Med Chem Lett 2014;5:826–30
- Capasso C, Supuran CT. Sulfa and trimethoprim-like drugs – antimetabolites acting as carbonic anhydrase, dihydropteroate synthase and dihydrofolate reductase inhibitors. J Enzyme Inhib Med Chem 2014;29:379–87
- Supuran CT. Carbonic anhydrase inhibitors: an editorial. Expert Opin Ther Pat 2013;23:677–9
- De Simone G, Alterio V, Supuran CT. Exploiting the hydrophobic and hydrophilic binding sites for designing carbonic anhydrase inhibitors. Expert Opin Drug Discov 2013;8:793–810
- Supuran CT. Structure-based drug discovery of carbonic anhydrase inhibitors. J Enzyme Inhib Med Chem 2012;27:759–72
- Del Prete S, Vullo D, Fisher GM, et al. Discovery of a new family of carbonic anhydrases in the malaria pathogen Plasmodium falciparum – the eta-carbonic anhydrases. Bioorg Med Chem Lett 2014;24:4389–96
- Vullo D, Del Prete S, Osman SM, et al. Sulfonamide inhibition studies of the delta-carbonic anhydrase from the diatom Thalassiosira weissflogii. Bioorg Med Chem Lett 2014;24:275–9
- Del Prete S, Vullo D, Scozzafava A, et al. Cloning, characterization and anion inhibition study of the delta-class carbonic anhydrase (TweCA) from the marine diatom Thalassiosira weissflogii. Bioorg Med Chem 2014;22:531–7
- Del Prete S, Vullo D, De Luca V, et al. Biochemical characterization of the delta-carbonic anhydrase from the marine diatom Thalassiosira weissflogii, TweCA. J Enzyme Inhib Med Chem 2014. [Epub ahead of print]. DOI: 10.3109/14756366.2013.868599
- Capasso C, Supuran CT. Anti-infective carbonic anhydrase inhibitors: a patent and literature review. Expert Opin Ther Pat 2013;23:693–704
- Vullo D, Kupriyanova EV, Scozzafava A, et al. Anion inhibition study of the beta-carbonic anhydrase (CahB1) from the cyanobacterium Coleofasciculus chthonoplastes (ex-Microcoleus chthonoplastes). Bioorg Med Chem 2014;22:1667–71
- Vullo D, Flemetakis E, Scozzafava A, et al. Anion inhibition studies of two alpha-carbonic anhydrases from Lotus japonicus, LjCAA1 and LjCAA2. J Inorg Biochem 2014;136:67–72
- Vullo D, Del Prete S, Osman SM, et al. Anion inhibition study of the beta-class carbonic anhydrase (PgiCAb) from the oral pathogen Porphyromonas gingivalis. Bioorg Med Chem Lett 2014;24:4402–6
- Vullo D, Del Prete S, Osman SM, et al. Sulfonamide inhibition studies of the gamma-carbonic anhydrase from the oral pathogen Porphyromonas gingivalis. Bioorg Med Chem Lett 2014;24:240–4
- Rodrigues GC, Feijo DF, Bozza MT, et al. Design, synthesis, and evaluation of hydroxamic acid derivatives as promising agents for the management of Chagas disease. J Med Chem 2014;57:298–308
- Prete SD, Vullo D, Osman SM, et al. Sulfonamide inhibition study of the carbonic anhydrases from the bacterial pathogen Porphyromonas gingivalis: the beta-class (PgiCAb) versus the gamma-class (PgiCA) enzymes. Bioorg Med Chem 2014;22:4537–43
- Nishimori I, Vullo D, Minakuchi T, et al. Anion inhibition studies of two new beta-carbonic anhydrases from the bacterial pathogen Legionella pneumophila. Bioorg Med Chem Lett 2014;24:1127–32
- Nishimori I, Vullo D, Minakuchi T, et al. Sulfonamide inhibition studies of two beta-carbonic anhydrases from the bacterial pathogen Legionella pneumophila. Bioorg Med Chem 2014;22:2939–46
- Del Prete S, Vullo D, De Luca V, et al. Biochemical characterization of recombinant beta-carbonic anhydrase (PgiCAb) identified in the genome of the oral pathogenic bacterium Porphyromonas gingivalis. J Enzyme Inhib Med Chem 2014. [Epub ahead of print]. DOI: 10.3109/14756366.2014.931383
- Del Prete S, De Luca V, Vullo D, et al. Biochemical characterization of the gamma-carbonic anhydrase from the oral pathogen Porphyromonas gingivalis, PgiCA. J Enzyme Inhib Med Chem 2014;29:532–7
- Del Prete S, De Luca V, Scozzafava A, et al. Biochemical properties of a new alpha-carbonic anhydrase from the human pathogenic bacterium, Vibrio cholerae. J Enzyme Inhib Med Chem 2014;29:23–7
- De Luca V, Del Prete S, Supuran CT, Capasso C. Protonography, a new technique for the analysis of carbonic anhydrase activity. J Enzyme Inhib Med Chem 2014. [Epub ahead of print]
- Ceruso M, Del Prete S, AlOthman Z, et al. Synthesis of sulfonamides with effective inhibitory action against Porphyromonas gingivalis gamma-carbonic anhydrase. Bioorg Med Chem Lett 2014;24:4006–10
- Lehneck R, Neumann P, Vullo D, et al. Crystal structures of two tetrameric beta-carbonic anhydrases from the filamentous ascomycete Sordaria macrospora. FEBS J 2014;281:1759–72
- Di Fiore A, Truppo E, Supuran CT, et al. Crystal structure of the C183S/C217S mutant of human CA VII in complex with acetazolamide. Bioorg Med Chem Lett 2010;20:5023–6
- Avvaru BS, Wagner JM, Maresca A, et al. Carbonic anhydrase inhibitors. The X-ray crystal structure of human isoform II in adduct with an adamantyl analogue of acetazolamide resides in a less utilized binding pocket than most hydrophobic inhibitors. Bioorg Med Chem Lett 2010;20:4376–81
- Temperini C, Scozzafava A, Supuran CT. Carbonic anhydrase inhibitors. X-ray crystal studies of the carbonic anhydrase II-trithiocarbonate adduct – an inhibitor mimicking the sulfonamide and urea binding to the enzyme. Bioorg Med Chem Lett 2010;20:474–8
- Alterio V, Hilvo M, Di Fiore A, et al. Crystal structure of the catalytic domain of the tumor-associated human carbonic anhydrase IX. Proc Natl Acad Sci USA 2009;106:16233–8
- Di Fiore A, Monti SM, Hilvo M, et al. Crystal structure of human carbonic anhydrase XIII and its complex with the inhibitor acetazolamide. Proteins 2009;74:164–75
- Temperini C, Scozzafava A, Puccetti L, Supuran CT. Carbonic anhydrase activators: X-ray crystal structure of the adduct of human isozyme II with L-histidine as a platform for the design of stronger activators. Bioorg Med Chem Lett 2005;15:5136–41
- Vullo D, Sai Kumar RS, Scozzafava A, et al. Anion inhibition studies of a beta-carbonic anhydrase from Clostridium perfringens. Bioorg Med Chem Lett 2013;23:6706–10
- Vullo D, Luca VD, Scozzafava A, et al. The alpha-carbonic anhydrase from the thermophilic bacterium Sulfurihydrogenibium yellowstonense YO3AOP1 is highly susceptible to inhibition by sulfonamides. Bioorg Med Chem 2013;21:1534–8
- Vullo D, Leewattanapasuk W, Muhlschlegel FA, et al. Carbonic anhydrase inhibitors: inhibition of the beta-class enzyme from the pathogenic yeast Candida glabrata with sulfonamides, sulfamates and sulfamides. Bioorg Med Chem Lett 2013;23:2647–52
- Vullo D, Isik S, Del Prete S, et al. Anion inhibition studies of the alpha-carbonic anhydrase from the pathogenic bacterium Vibrio cholerae. Bioorg Med Chem Lett 2013;23:1636–8
- Vullo D, De Luca V, Scozzafava A, et al. The extremo-alpha-carbonic anhydrase from the thermophilic bacterium Sulfurihydrogenibium azorense is highly inhibited by sulfonamides. Bioorg Med Chem 2013;21:4521–5
- Syrjanen L, Vermelho AB, Rodrigues Ide A, et al. Cloning, characterization, and inhibition studies of a beta-carbonic anhydrase from Leishmania donovani chagasi, the protozoan parasite responsible for leishmaniasis. J Med Chem 2013;56:7372–81
- Pan P, Vermelho AB, Scozzafava A, et al. Anion inhibition studies of the alpha-carbonic anhydrase from the protozoan pathogen Trypanosoma cruzi, the causative agent of Chagas disease. Bioorg Med Chem 2013;21:4472–6
- Pan P, Vermelho AB, Capaci Rodrigues G, et al. Cloning, characterization, and sulfonamide and thiol inhibition studies of an alpha-carbonic anhydrase from Trypanosoma cruzi, the causative agent of Chagas disease. J Med Chem 2013;56:1761–71
- Nishimori I, Vullo D, Minakuchi T, et al. Restoring catalytic activity to the human carbonic anhydrase (CA) related proteins VIII, X and XI affords isoforms with high catalytic efficiency and susceptibility to anion inhibition. Bioorg Med Chem Lett 2013;23:256–60
- Monti SM, De Simone G, Dathan NA, et al. Kinetic and anion inhibition studies of a beta-carbonic anhydrase (FbiCA 1) from the C4 plant Flaveria bidentis. Bioorg Med Chem Lett 2013;23:1626–30
- Luca VD, Vullo D, Scozzafava A, et al. An alpha-carbonic anhydrase from the thermophilic bacterium Sulphurihydrogenibium azorense is the fastest enzyme known for the CO2 hydration reaction. Bioorg Med Chem 2013;21:1465–9
- Guzel-Akdemir O, Akdemir A, Pan P, et al. A class of sulfonamides with strong inhibitory action against the alpha-carbonic anhydrase from Trypanosoma cruzi. J Med Chem 2013;56:5773–81
- Di Fiore A, Capasso C, De Luca V, et al. X-ray structure of the first `extremo-alpha-carbonic anhydrase', a dimeric enzyme from the thermophilic bacterium Sulfurihydrogenibium yellowstonense YO3AOP1. Acta Crystallogr D Biol Crystallogr 2013;69:1150–9
- Del Prete S, Vullo D, De Luca V, et al. A highly catalytically active gamma-carbonic anhydrase from the pathogenic anaerobe Porphyromonas gingivalis and its inhibition profile with anions and small molecules. Bioorg Med Chem Lett 2013;23:4067–71
- Alafeefy AM, Isik S, Al-Jaber NA, et al. Carbonic anhydrase inhibitors. Benzenesulfonamides incorporating cyanoacrylamide moieties strongly inhibit Saccharomyces cerevisiae beta-carbonic anhydrase. Bioorg Med Chem Lett 2013;23:3570–5
- Akdemir A, Vullo D, De Luca V, et al. The extremo-alpha-carbonic anhydrase (CA) from Sulfurihydrogenibium azorense, the fastest CA known, is highly activated by amino acids and amines. Bioorg Med Chem Lett 2013;23:1087–90
- Akdemir A, Guzel-Akdemir O, Scozzafava A, et al. Inhibition of tumor-associated human carbonic anhydrase isozymes IX and XII by a new class of substituted-phenylacetamido aromatic sulfonamides. Bioorg Med Chem 2013;21:5228–32
- McKenna R, Supuran CT. Carbonic anhydrase inhibitors drug design. Subcell Biochem 2014;75:291–323
- Carta F, Aggarwal M, Maresca A, et al. Dithiocarbamates: a new class of carbonic anhydrase inhibitors. Crystallographic and kinetic investigations. Chem Commun (Camb) 2012;48:1868–70
- Winum JY, Supuran CT. Recent advances in the discovery of zinc-binding motifs for the development of carbonic anhydrase inhibitors. J Enzyme Inhib Med Chem 2014. [Epub ahead of print]
- Carta F, Akdemir A, Scozzafava A, et al. Xanthates and trithiocarbonates strongly inhibit carbonic anhydrases and show antiglaucoma effects in vivo. J Med Chem 2013;56:4691–700
- Davis RA, Vullo D, Supuran CT, Poulsen SA. Natural product polyamines that inhibit human carbonic anhydrases. Biomed Res Int 2014;2014:374079
- Carradori S, De Monte C, D'Ascenzio M, et al. Salen and tetrahydrosalen derivatives act as effective inhibitors of the tumor-associated carbonic anhydrase XII – a new scaffold for designing isoform-selective inhibitors. Bioorg Med Chem Lett 2013;23:6759–63
- Supuran CT. Amide derivatives of benzene-sulfonanilide, pharmaceutical composition thereof and method for cancer treatment using the same (US20120095092). Expert Opin Ther Pat 2012;22:1251–5
- Supuran CT. Carbonic anhydrase inhibitors and activators for novel therapeutic applications. Future Med Chem 2011;3:1165–80
- Durdagi S, Senturk M, Ekinci D, et al. Kinetic and docking studies of phenol-based inhibitors of carbonic anhydrase isoforms I, II, IX and XII evidence a new binding mode within the enzyme active site. Bioorg Med Chem 2011;19:1381–9
- Supuran CT. Carbonic anhydrase inhibition with natural products: novel chemotypes and inhibition mechanisms. Mol Divers 2011;15:305–16
- Carta F, Temperini C, Innocenti A, et al. Polyamines inhibit carbonic anhydrases by anchoring to the zinc-coordinated water molecule. J Med Chem 2010;53:5511–22
- Krungkrai SR, Suraveratum N, Rochanakij S, Krungkrai J. Characterisation of carbonic anhydrase in Plasmodium falciparum. Int J Parasitol 2001;31:661–8
- Khalifah RG. The carbon dioxide hydration activity of carbonic anhydrase. I. Stop-flow kinetic studies on the native human isoenzymes B and C. J Biol Chem 1971;246:2561–73
- Laemmli UK. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970;227:680–5