
Abstract
In this study, we aimed to investigate the effect of some coumarin and benzoxazinone derivatives on the activity of human PON1. Human serum paraoxonase 1 was purified from fresh human serum blood by two-step procedures that are ammonium sulfate precipitation (60–80%) and then hydrophobic interaction chromatography (Sepharose 4B, L-tyrosine and 1-napthylamine). The enzyme was purified 232-fold with a final specific activity of 27.1 U/mg. In vitro effects of some previously synthesized ionic coumarin or benzoxazinone derivatives (1–21) on purified PON1 activity were investigated. Compound 14 (1-(2,3,4,5,6)-pentamethylbenzyl-3-(6,8-dimethyl-2H-chromen-2-one-4-yl))benzimidazolium chloride was found out as the strongest inhibitor (IC50 = 7.84 μM) for PON1 among the compounds. Kinetic investigation and molecular docking study were evaluated for one of the most active compounds (compound 12) and obtained data showed that this compound is competitive inhibitor of PON1 and interact with Leu262 and Ser263 in the active site of PON1. Moreover, coumarin derivatives were found out as the more potent inhibitors for PON1 than benzoxazinone derivatives.
Introduction
The paraoxonase (PON) enzyme family has three members, PON1, PON2 and PON3, that PON1 is the best studied member of this family within mammalian enzymes. The PON1 is the calcium-dependent enzyme that has 355 amino acids with a mass of 43 kDaCitation1,Citation2. “PON” gets name from one of the most commonly used in vitro substrates, paraoxonCitation3. It also hydrolyzes aromatic carboxyl esters such, lactones, thiolactones and cyclic carbonate estersCitation4–7. Furthermore, PON1 hydrolyzes some organophosphate-derived nerve agents such as sarin and soman and pesticide diazoxonCitation1; however, physiological substrate and biological function of this enzyme have not known yet. Beside hydrolysis of organophosphates, antiatherogenic properties of PON1 were reported. Some in vitro studies showed that PON1 inhibits oxidation of low-density lipoprotein (LDL) and high-density lipoprotein (HDL) and reduced levels of oxidized lipids are involved in the initiation of atherosclerosisCitation8,Citation9. Moreover, decreased PON1 levels and activities were reported on the atherosclerotic patientsCitation10.
In light of these knowledge, decreased activity of PON1 was acknowledged as a risk of atherosclerosis and organophosphate toxicity, and therefore, the factors affecting PON1 have to be understood clearly. In literature, some crucial studies were carried out in order to understand the biological function and substrate, mechanisms of action of PON1Citation11,Citation12. A small number of inhibition studies on PON1 were reported in literature with various commercially available drugs, some metal ions, coumarin derivatives and pesticidesCitation13–18. Because of all these reasons, PONs have become the subject of intensive research area.
Coumarins are member of a class of compounds known as benzopyrones. In recent year’s research, the coumarin chemistry have attracted much attention due to diverse biological and pharmacological properties of coumarin derivatives, such as anticoagulantCitation19, anticancerCitation20–22, anti-HIVCitation23,Citation24, anti-inflammatoryCitation25, antimicrobialCitation26,Citation27 and some different biological activitiesCitation28–32. Most recently, coumarins were reported as “suicide” inhibitors of human carbonic anhydrases (hCA)Citation33. As shown, coumarin derivatives are promising candidates for the treatment of some important disease.
The other class of compounds, which was used in this work, is benzoxazinone. This compound is a heterocyclic compound that contains the 1,4-oxazinone ring fused to benzene. Various biological properties of benzoxazinone derivatives are known in the literature and they have an important role in the design and development of new drugsCitation34–38.
As mentioned previously, the activity of PON1 is vital for human and we aimed to examine the in vitro effects of 21 coumarin and benzoxazinone derivatives on the activity of purified human serum PON1. We chose these class compounds because of their important biological activities. Moreover, in our previous study, we showed that some coumarin compounds have significant inhibitory properties on the activity of PON1Citation39. Another reason of carrying out this study is biological interference of coumarin compounds with coagulation, thrombotic events and antiatherogenic properties of PON1. The compounds that were used in this study had remained from our previous studyCitation40. For one of the strongest inhibitors, we performed kinetic and molecular docking studies to obtain insights into binding mode of these compounds with key residues of the active site of PON1.
Materials and methods
The materials used include Sepharose 4B, L-tyrosine, 1-napthylamine, paraoxon and protein assay reagents were obtained from Sigma Chem. Co (Istanbul, Turkey). Thirteen coumarin and eight benzoxazinone derivatives were ready from our previous study and detailedCitation40.
Paraoxonase enzyme assay
Paraoxonase enzyme activity toward paraoxon was quantified spectrophotometrically by the method described by Gan et al.Citation41. The enzyme assay was based on the estimating of p-nitrophenol at 412 nm. The molar extinction coefficient of p-nitrophenol (ɛ = 17 100 M −1 cm−1 at pH 8) was used to calculate enzyme activity. The reaction was followed for 2 min at 37 °C by monitoring the appearance of p-nitrophenol at 412 nm in automated recording spectrophotometer (Biotek, Winooski, VT). About 2 mM of final substrate concentration was used during enzyme assay, and all measurements were taken in duplicate and corrected for the nonenzymatic hydrolysis.
Purification of paraoxonase from human serum by hydrophobic interaction chromatography
Human serum was isolated from 40 mL fresh human blood and put into a dry tube. The blood samples were centrifuged at 3000 rpm for 15 min and the serum was removed. Firstly, serum paraoxonase was isolated by ammonium sulfate precipitation (60–80%). The precipitate was collected by centrifugation at 15 000 rpm for 20 min and redissolved in 100 mM Tris HCl buffer (pH 8.0). Then, we synthesized the hydrophobic gel, including Sepharose 4B, L-tyrosine and 1-napthylamine, for the purification of human serum paraoxonaseCitation42. The purified enzyme had a specific activity of 11.76 U/mg. The column was equilibrated with 0.1 M of a Na2HPO4 buffer (pH 8.00) including 1 M ammonium sulfate. The paraoxonase was eluted with an ammonium sulfate gradient using 0.1 M Na2HPO4 buffer with and without ammonium sulfate (pH 8.00). The purified PON1 enzyme was stored in the presence of 2 mM calcium chloride in order to maintain activity.
Total protein determination
The absorbance at 280 nm was used to monitor the protein in the column effluents and ammonium sulfate precipitation. Quantitative protein determination was achieved by absorbance measurements at 595 nm according to BradfordCitation43, with bovine serum albumin as a standard.
SDS polyacrylamide gel electrophoresis
SDS polyacrylamide gel electrophoresis was performed after purification of the enzyme. It was carried out in 10% and 3% acrylamide concentration for the running and stacking gel, respectively, containing 0.1% SDS according to LaemmliCitation44. A 20 mg sample was applied to the electrophoresis medium. Gel was stained overnight in 0.1% Coomassie Brilliant Blue R-250 in 50% methanol and 10% acetic acid, then destained by frequently changing the same solvent, without dye. The electrophoretic pattern was photographed with the system of produce as an image of the gel.
In vitro kinetic studies and calculation of IC50 and ki values
For the inhibition studies of coumarin derivatives, different concentrations of coumarin derivatives were added to the reaction medium. PON1 activity with coumarin derivatives was assayed by following the hydration of paraoxon. Activity percentage values of PON for five different concentrations of each coumarin derivatives were determined by regression analysis using the Microsoft Office 2000 Excel. PON1 enzyme activity without a coumarin derivative was considered as 100% activity. The inhibitor concentration causing up to 50% inhibition (IC50 values) for coumarin derivatives were determined from the graphs. So as to determinate Ki, Km and Vmax values of the PON1 enzyme using paraoxon as a substrate was measured at eight different substrate concentrations (0.5, 0.6, 0.7, 0.8, 0.9, 1.0, 1.1 and 1.2 mM) at pH 8.00 and 37 °C in the presence of two different coumarin concentrations of compound 12 (28.5 μM and 19 μM) as knowing highest and lowest final concentrations. Km and Vmax values were calculated 62.07 μM, 6.66 μmol/min, respectively, through Lineweaver–Burke graphs. The coumarin compound named 12 inhibited PON1 enzyme activity in a competitive inhibition type with Ki of 13.72 μM value. As a result of in vitro experiments and by the means of Lineweaver–Burke graphs competitive inhibition type was controlled with docking for confirmation.
Explanation of using compound 12 instead of compound 14 for determination of Ki value
Although there were more effective inhibitors, compound 12 was chosen for estimation Ki, Km and Vmax values. The only reason was run short of these inhibitors. So that compound 12 was used for the most potent inhibitor in depleted compounds after inhibition studies (estimation for IC50 values).
Molecular docking
Enzyme setup. Determination of the consistent receptor was based on previous studyCitation12. Macromolecule file (PDB code: 1V04) was modified using the ADT package version 1.5.6rc3 (Ankara, Turkey). All water molecules were deleted and polar hydrogens were added. Subsequently, Gasteiger charges were calculated and the generated pdbqt files were saved.
Ligand Energy minimization of compound 12 was carried out using GAMESS module for ChemOffice version Ultra 8.0.3 (Ankara, Turkey). All data were saved as pdb with the aid of Molegro Molecular Viewer version 2.5 (Ankara, Turkey). Further modification of these partial charges of pdb files was carried out through the ADT package so that the charges of the nonpolar hydrogen atoms allocated to the atom to which the hydrogen is attached. These modified pdb files saved as pdbqt files. Appropriate grid box points were determined by centering on ligand separately for each compound.
Results and discussion
As mentioned in “Introduction” section, a few in vitro inhibition studies on PON1 were reported in literature so far. In a study, Erzengin and coworkers studied inhibition effects of some coumarin derivatives and 6,7-dihydroxy-3-(4-methylphenyl)-2H-chromen-2-one inhibited PON1 with IC50 value of 3 μMCitation14. According to our literature survey, this compound is the most active inhibitor of human PON1. In addition, inhibition effects of some anesthetics, calcium channel blockers, some metal ions and some pesticides on PON1 were reported and IC50 values of 85–1678, 121–255, 4–317 and 709–35011 μM were determined, respectivelyCitation15–18.
In this study, the effects of 21 ionic coumarin or benzoxazinone derivatives on PON1 were investigated. These compounds were ready from our previous study. The compounds had been synthesized by the interaction of N-alkyl imidazolium or benzimidazolium derivatives with some 4-chloromethylene substituted coumarin derivatives or 6-chloroacetylene substituted benzoxazinone derivatives. The synthesized compounds had been obtained in good yields between 43–92%. Characterization of the compounds had been elucidated by nuclear magnetic resonance (NMR), infrared (IR) and elemental analyses in satisfactory mannerCitation40. For evaluating the PON1 activity, all compounds were subjected to PON1 inhibition assay with paraoxon as substrate. The IC50 values and structures of compounds were given in . The results showed that all compounds (1–21) inhibited PON1 enzyme activity with different sensitivity. We determined IC50 values in the range of 7.84–543.70 μM. Among them, 13 compounds (1,3,4,6,10–16,20,21) performed high inhibition activity and IC50 values of these compounds are ranging between 7.84–36.57 μM. Compound 14 was found out as the strongest inhibitor of PON1 with 7.84 μM value of IC50. The other eight compounds showed relatively lower inhibitory activity against PON1. Moreover, Ki value and inhibition type of compound 12 (1-benzyl-3–(6,8-dimethyl-2H-chromene-2-one-4-yl))benzimidazolium chloride were investigated and results demonstrated that this compound inhibited enzyme activity in a competitive manner with Ki value of 13.72 μM (). As mentioned earlier, inhibitory activities of some biologically active compounds and metal ions on PON1 were studied but, studies about interactions of previously unused compounds with PON1 are scarce. In literature, it is known that coumarin derivatives have high inhibitory properties against PON1 but, mechanism of action is still unknown. We may say that, the compounds which reported in this study have good inhibitory properties when results compared with literature.
Figure 1. Inhibition of paraoxonase by compound 12. The slope of Lineweaver–Burk plots indicates competitive inhibition for paraoxon.
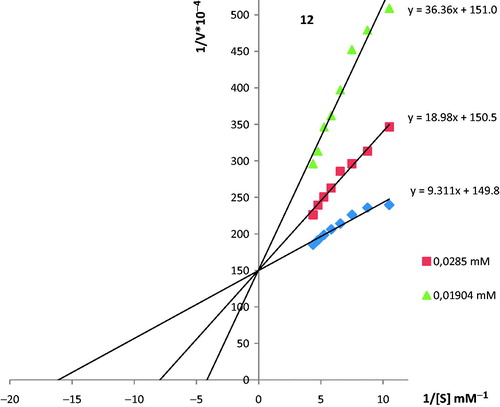
Table 1. Structures and IC50 values of compounds. These compounds were ready from our previous studyCitation35.
In a study, it was shown that simple lactone derivatives were hydrolyzed by PON1. In the same study, authors reported that lactam derivatives and a coumarin were not hydrolyzed by PON1Citation11. In our study, tested compounds are bearing several types of substituent, such as hydroxyl, azolium and alkyl or aryl groups, so according to the results of our and previous studies, compounds 1–21 are not suitable substrates for PON1 and some of them inhibited PON1 effectively.
Structure–activity relationship
We can classify the compounds tested in this study in two different ways; (i) imidazolium- or benzimidazolium-based compounds, (ii) coumarin- or benzoxazinone-based compounds. From results of enzyme inhibition assay, two opinions about structure–activity relationship were emerged.
In the tested compounds, some compounds are bearing same groups in their structures apart from coumarin or benzoxazinone scaffolds and we can use these compounds for comparison. When we compared the compounds 10–13 with 17–20, each of these compounds are bearing methyl, allyl, benzyl and naphthyl groups as substituent and 6,8-dimethyl coumarin derivatives (10–13) have much lower IC50 values than benzoxazinone derivatives (17–20). In addition, this situation can be seen in the imidazolium salts. When compounds 1–6 compared with compounds 7–9, it is clear that coumarin-based imidazolium salts are more active than benzoxazinone corresponding. Therefore, it must be noted that coumarin derivatives showed higher inhibitory activity than benzoxazinone derivatives. If we compare coumarin benzimidazolium salts, there are some minor differences due to electronic effects. If compounds 15 and 16 compared, dihydroxy-substituted 16 is more active than monohydroxy-substituted 15. Moreover, when compounds 14 and 15 compared which are monohydroxy coumarin substituted, pentamethyl substituted 14 is more active than naphthalene substituted 15. If we want to observe the difference between imidazole and benzimidazole scaffold, we may compare the compounds 1 and 10. These compounds are bearing same groups on nitrogen atom. IC50 values showed that benzimidazole derivative 10 is more active than imidazole derivative 1.
When imidazolium-based compounds 2,5 compared with 1,3,4,6, an interesting difference can be seen. Compounds 2,5 performed lower activity than other imidazolium coumarin derivatives. In these compounds, methylation of position-2 decreased inhibition activity and possible explanation for the lower activity associated with the absence of acidic imidazolium proton. This fragment of compounds is the most hydrophilic and polar part of compounds and absence of acidic proton may cause reduced interactions between inhibitors and amino acid residues.
Molecular docking studies
Kinetic data demonstrated that compound 12 is competitive inhibitor of PON1 and in order to obtain more insights into the binding mode, molecular docking studies were also performed for compound 12. Docking scores were obtained using Lamarckian Genetic Algorithm and scoring function of AutoDock 4. Then, interactions were checked with the aid of ADT and Discovery Studio 4.0 Client (Ankara, Turkey). Docking scores and binding interactions of compound 12 with PON1 (PDB code: 1V04) are presented in . Final images of compound 12 for binding interactions are shown in .
Figure 2. Docking of compound 12 within the active site of PON1 (A) Discovery Studio 4.0 Client images and (B) ADT images.
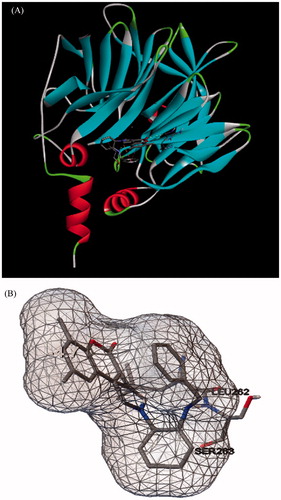
Table 2. Kinetic and molecular docking data of compound 12.
As seen from the data presented in , there are two amino acid residues participating in close van der Waals (<4Å) contacts with the inhibitor compound 12 when bound to the active site of PON1. According to results of molecular docking analysis, compound 12 interacts with Leu262 and Ser263 in the active site of PON1.
In 2004, Harel et al reported that the crystal structure of recombinant PON1 (rePON1) obtained from Escherichia coli (E. coli). In the same study, the authors reported that rePON1 and human PON1 (hPON1) have some differences in the sequence of amino acids, but they also reported that sequence variations between rePON1 and hPON1 are in the areas where their active site and overall structures are not affected. One of these differences is in the residue 263. It was shown that in the crystal structure of rePON1 residue 263 is serine and in hPON1 aspartic acid takes place of serineCitation12. Although molecular docking studies report the interaction of compound 12 with serine, in hPON1, most probably it interacts with aspartic acid. Aspartic acid has a carboxyl group and this amino acid is negatively charged at pH 7.4 and this may cause more effective interaction with positively charged compound 12. Moreover, decrease in the inhibition activity of compounds 2 and 5, which were pointed out above may attributable with the absence of interaction of acidic proton with negatively charged aspartic acid. In addition, results showed that compound 12 did not interact with Ca2+ which is responsible for catalytic activity.
Conclusion
PON1 has important detoxification role in metabolism and it is thought that decreased activity of PON1 may cause atherosclerosis and organophosphate toxicity so all factors affecting PON1 have to be well determined. Enzyme inhibition is an important issue for drug design and biochemical applicationsCitation45–47. In this study, we have showed that human PON1 was inhibited effectively by some coumarin or benzoxazinone derivatives. Coumarin derivative compound 12 has been showed as competitive inhibitor of PON1. Molecular docking data showed that compound 12 inhibits PON1 by not-interacting with active site metal Ca2+ ions and it interacts with Leu262 and Ser263 in the active site of PON1. It is known that coumarins are toxic for humans in certain levels, but their some important biological activities make them attractive for use in future. Inhibition of PON1 by coumarin derivatives provides a justification to further consideration of limitation dosage of coumarin as drug and as flavor cause of risk assessment, but more detailed investigations is necessary about mechanisms of inhibition of PON1 by coumarin and benzoxazinone derivatives.
Declaration of interest
This work was supported by the Research Fund of the Balikesir University (project no. 2014–54).
Related Research Data
References
- Draganov DI, La Du BN. Pharmacogenetics of paraoxonases: a brief review. Naunyn Schmiedebergs Arch Pharmacol 2004;369:78–88
- Goswani B, Tayal D, Gupta N, Mallika V. Paraoxonase: a multifaceted biomolecule. Clin Chim Acta 2009;410:1–12
- Van Himbergen TM, Van Tits LJH, Roest M, Stalenhoef AFH. The story of PON1: how an organophosphate-hydrolysing enzyme is becoming a player in cardiovascular medicine. Neth J Med 2006;64:34–8
- Aharoni A, Gaidukov L, Khersonsky O, et al. The ‘evolvability' of promiscuous protein functions. Nat Genet 2005;37:73–6
- Khersonsky O, Tawfik DS. Structure-reactivity studies of serum paraoxonase PON1 suggest that its native activity is lactonase. Biochemistry 2005;44:6371–8
- Rochua D, Chobriere E, Massono P. Human paraoxonase: a promising approach for pre-treatment and therapy of organophosphorus poisoning. Toxicology 2007;233:459
- Alici HA, Ekinci D, Beydemir S. Intravenous anaesthetics inhibit human serum paraoxonase (PON1) activity in vitro and in vivo. Clin Biochem 2008;41:1384–90
- Mackness MI, Arrol S, Durrington PN. Paraoxonase prevents accumulation of lipoperoxides in low-density-lipoprotein. FEBS Lett 1991;286:152–4
- Reddy ST, Wadleigh DJ, Grijalva V, et al. Human paraoxonase-3 is an HDL-associated enzyme with biological activity similar to paraoxonase-1 protein but it is not regulated by oxidizing lipids. Arterioscler Thromb Vasc Biol 2001;21:542–7
- Rozenberg O, Aviram M. S-Glutathionation regulates HDL-associated paraoxonase1 (PON1) activity. Biochem Biophys Res Comm 2006;351:492–8
- Billecke S, Draganov D, Counsell R, et al. Human serum paraoxonase (PON1) isozymes Q and R hydrolyze lactones and cyclic carbonate esters. Drug Metab Dispos 2000;28:1335–41
- Harel M, Aharoni A, Gaidukov L, et al. Structure and evolution of the serum paraoxonase family of detoxifying and anti-atherosclerotic enzymes. Nat Struct Mol Biol 2004;11:412–19
- Kockar F, Sinan S, Yildirim H, Arslan O. Differential effects of some antibiotics on paraoxonase enzyme activity on human hepatoma cells (Hep62) in vitro. J. Enzyme Inhib Med Chem 2010;25:715–19
- Erzengin M, Basaran I, Cakir U, et al. In vitro inhibition effects of some dihydroxycoumarin compounds on purified human serum paraoxonase I (PON1). Appl Biochem Biotechnol 2012;168:1540–8
- Senturk M, Ekinci D, Alici HA, Beydemir S. Paraoxonase-1, an organophosphate detoxifier and cardioprotective enzyme is inhibited by anasthetics: an in vitro and in vivo insight. Pest Biochem Physiol 2011;11:206–11
- Turkes C, Soyut H, Beydemir S. Effect of calcium channel blockers on paraoxonase-1 (PON1) activity and oxidative stress. Pharmacol Rep 2014;66:74–80
- Pla A, Rodrigo L, Hernandez AF, et al. Effect of metal ions and calcium on purified PON1 and PON3 from rat liver. Chem Biol Interact 2007;167:63–70
- Gencer N, Arslan O. In vitro effects of some pesticides on PON1Q192 and PON1R192 isoenzymes from human serum. Fresen Environ Bull 2011;20:590–6
- Gebauer M. Synthesis and structure-activity relationship of novel warfarin derivatives. Bioorg Med Chem 2007;25:2414–20
- Lacy A, O’Kennedy R. Studies on coumarins and coumarin-related compounds to determine their therapeutic role in the treatment of cancer. Curr Pharm Des 2004;10:3797–811
- Kontogiorgis CA, Detsi A, Hadjipavlou-Litina D. Coumarin-based drugs: a patent review. Expert Opin Ther Patents 2012;22:437–54
- Saidu NEB, Valente S, Bana E, et al. Coumarin polysulfides inhibit cell growth and induce apoptosis in HCT116 colon cancer cells. Bioorg Med Chem 2012;20:1584–93
- Zhao H, Neamati N, Hong H, et al. Coumarin-based inhibitors of HIV integrase. J Med Chem 1997;40:242–9
- Yu D, Suzuki M, Xie L, et al. Recent progress in the development of coumarin derivatives as potent anti-HIV agents. Med Res Rev 2003;23:322–45
- Kontogiorgis CA, Hadjipavlou-Litina D. Synthesis and anti-inflammatory activity of coumarin derivatives. J Med Chem 2005;48:6400–8
- Smyth T, Ramaehandran VN, Smyth WF. A study of the antimicrobial activity of selected naturally occurring and synthetic coumarins. Int J Antimicrob Agents 2009;33:421–6
- Li Z, Li J, Qu D, et al. Synthesis and pharmacological evaluations of 4-hydroxycoumarin derivatives as a new class of anti-Staphylococcus aureus agents. J Pharm Pharmacol 2014;67:573–82
- Khan KM, Saify ZS, Hayat S, et al. Synthesis, antioxidant and insectisidal activities of some coumarin derivatives. J Chem Soc Pak 2002;24:226–31
- Khan KM, Saify ZS, Khan MZ, et al. Synthesis of coumarin derivatives with cytotoxic, antibacterial and antifungal activities. J Enzyme Inhib Med Chem 2004;19:373–9
- Haq Z, Lodhi MA, Nawaz SA, et al. 3S-QSAR CoMFA studies on bis-coumarin analogues as urease inhibitor: a strategic design in anti-urease agents. Bioorg Med Chem 2008;16:3456–61
- Khan KM, Rahim F, Wadood A, et al. Synthesis and molecular docking studies of potent α-glucosidase inhibitors based on biscoumarin skeleton. Eur J Med Chem 2014;81:245–52
- Khan KM, Fakhri MI, Shaikh NN, et al. Glucuronidase inhibitory studies on coumarin derivatives. Med Chem 2014;10:778–82
- Maresca A, Temperini C, Vu H, et al. Non-zinc mediated inhibition of carbonic anhydrases: coumarins are a new class of suicide inhibitors. J Am Chem Soc 2009;131:3057–62
- Kajino M, Shibouta Y, Nishikawa K, Megura K. Synthesis and biological activities of new 2-substitue 1,4-benzoxazinone derivatives. Chem Pharm Bull 1991;38:2896–908
- Katsura Y, Nishino S, Takasugi H. Studies on antiulcer drugs. I. Synthesis and antiulcer activities of imidazo[1,2-alpha]pyridinyl-2-oxobenzoxazolidines-3-oxo-2H-1,4-benzoxazines and related compounds. Chem Pharm Bull 1991;39:2937–43
- Turk CF, Krapcho J, Miche IM, Weinryb I. Synthesis and central nervous system activity of 2-arylidene-4-aminoalkyl-2H-1,4-benzoxazin-3(4H)-ones and related compounds. J Med Chem 1997;20:729–32
- Fringuelli R, Giocche N, Milanese L, et al. Bulky 1,4-benzoxazine derivatives with antifungal activity. Bioorg Med Chem 2009;17:3838–46
- Li X, Liu N, Zhang H, et al. Synthesis and SAR studies of benzoxazine MenB inhibitors: novel antibacterial agent against Mycobacterium tuberculosis. Bioorg Med Chem Lett 2010;2:6306–9
- Gokce B, Gencer N, Arslan O, et al. In vitro inhibiton effect of some coumarin compounds on purified human serum paraoxonase 1 (PON1). J Enzyme Inhib Med Chem 2015. [Epub ahead of print]. Doi: 10.3109/14756366.2015.1043297
- Karatas MO, Uslu H, Sari S, et al. Coumarin or benzoxazionone based novel carbonic anhydrase inhibitors: synthesis, molecular docking and anticonvulsant studies. J Enzyme Inhib Med Chem 2015. [Epub ahead of print]. Doi: 10.3109/14756366.2015.1063624
- Gan KN, Smolen A, Eckerson HW, La Du BN. Purification of human serum paraoxonase/arylesterase. Evidence for one esterase catalyzing both activities. Drug Metab Dispos 1991;19:100–6
- Sinan S, Kockar F, Arslan O. Novel purification strategy for human PON1 and inhibition of the activity by cephalosporin and aminoglikozide derived antibiotics. Biochimie 2006;88:565–74
- Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 1976;72:248–54
- Laemmli DK. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970;227:680–5
- Karatas MO, Alici B, Cakir U, et al. Synthesis and carbonic anhydrase inhibitory properties of novel coumarin derivatives. J Enzyme Inhib Med Chem 2013;28:299–304
- Arslan M, Gencer N, Arslan O, et al. In vitro efficacy of some cattle drugs on bovine serum paraoxonase 1 (PON1) activity. J Enzyme Inhib Med Chem 2012;27:722–9
- Sayin D, Cakir DT, Gencer N, et al. Effects of some metals on paraoxonase activity from shark Scyliorhinus canicula. J Enzyme Inhib Med Chem 2012;27:595–8