Abstract
The overall impacts of winter feeding, including the effects of both forage and reindeer (Rangifer tarandus), were studied in an experiment in the Hammastunturi herding cooperative (68°N), Finland, with 300 reindeer in a sub-xeric heath forest (35 ha) during March/April 2009 and 2010. The feeding practices on the 50 plots were: (1) feeding with grass silage+hay with leftovers cleared in the spring; (2) feeding with grass silage+hay with leftovers not cleared; and (3) feeding with pellets. In addition, (4) unfenced and (5) fenced control plots were included, on which the reindeer were not fed. No invasive plant species introduced through grass silage+hay were observed on the plots. The coverage of Deschampsia flexuosa increased on grass silage+hay plots after the first winter, and both coverage and height of the grass increased after the second winter. The coverage of Dicranum sp. and Pleurozium schreberi was lower on plots where reindeer were fed with grass silage (leftover silage not cleared) vs. controls. Some dwarf shrubs, e.g., Calluna vulgaris, showed a similar response after the second winter. The N content of Empetrum nigrum was higher on grass silage plots (leftover silage not cleared) after the first winter vs. controls. After two winters, the N content of both E. nigrum and Vaccinium myrtillus was increased. Of the soil variables studied, C/N decreased on grass silage+hay plots vs. fenced controls. We conclude that even a moderate level of feeding may cause changes that can lead to a gradual shift from nutrient-poor heath forest towards a more nutrient-rich type.
To access the supplementary material for this article, please see the supplementary files under Article Tools online.
In northern Fennoscandia and north-west Russia, reindeer herding has been affected in recent decades by such socio-economic changes as mechanization, supplementary winter feeding, regulations regarding meat production and parasite medication (Jernsletten & Klokov Citation2002; Forbes et al. Citation2006; Tyler et al. Citation2007). In addition, the quality and extent of pasture have declined considerably owing to, among other factors, competing forms of land use, such as forestry, industrial development, construction, mining and tourism, as well as overgrazing caused by the increased number of animals (Kumpula et al. Citation2009; Mattila & Mikkola Citation2009). Furthermore, pressures exist to increase supplementary winter feeding: higher reindeer densities and inadequate pasture rotation systems have impaired the quality of winter pastures, and the predicted fluctuations in winter weather and snow conditions may well reduce the availability of natural forage (Putkonen & Roe Citation2003; Nieminen Citation2010; Callaghan et al. Citation2011; Hansen et al. Citation2011).
A changing climate can have both positive and negative impacts on the availability and quality of reindeer forage (Weladji & Holland Citation2006; Tyler et al. Citation2007; Moen Citation2008; Turunen et al. Citation2009; Tømmervik et al. Citation2012). It has been predicted that the northern latitudes will become warmer and wetter, especially during autumn, winter and spring. Increasing winter temperatures and temperature fluctuations will lead to more frequent snowmelt, rain-on-snow events and freeze—thaw cycles, which in turn causes compaction of the snow pack, formation of ice layers above and within the snow cover, and ice encasement (Putkonen & Roe Citation2003; Solomon et al. Citation2007; Callaghan et al. Citation2011). Icing and variations in the thickness of the snow cover may worsen the conditions under which reindeer dig for forage and thereby increase their energy expenditure. In fact, decreased availability of winter forage may be one reason for the deterioration observed in the condition of animals and for low calving percentages (Helle & Kojola Citation2008; Hansen et al. Citation2011; Vuojala-Magga et al. Citation2011).
In Finland, supplementary winter feed consists primarily of hay, grass silage, grain-based commercial pellets or a combination of these. During the period 2007–08, 18 million kg of pellets and over 40 million kg of hay+grass silage (calculated as dry hay) were used for feeding (Nieminen Citation2010). In the central and northern parts of the reindeer herding area, field feeding is more common than pen feeding, with more than 60% of all animals being fed in the field. Field feeding is always based partly on natural pastures; in pens reindeer get all or at least most of the forage from the herder. Supplementary feeding has made it possible to keep the number and condition of reindeer stable despite difficult winter weather and snow conditions (Åhman & Danell Citation2006; Nieminen Citation2010). A female reindeer in good condition after the winter will normally give birth to a healthy calf and be able to take care of it well (Eloranta & Nieminen Citation1986). The calves of reindeer fed with supplementary forage are heavier than those of reindeer that forage only on natural pastures (Säkkinen et al. Citation1999; Maijala & Nieminen Citation2001). Yet, winter feeding may increase the risk of diseases (Oksanen et al. Citation1997; Tryland et al. Citation2001; Aschfalk et al. Citation2003). It may also intensify the widely reported direct impacts of reindeer grazing on and trampling winter pasture vegetation—reduced lichen cover in particular—as well as the indirect impacts on soil processes, for example, C and N mineralization and organic matter decomposition, which greatly depend on the site type and grazing intensity (Väre et al. Citation1995; Olofsson et al. Citation2001; Olofsson & Oksanen Citation2002, Citation2005; Olofsson et al. Citation2004; Pajunen et al. Citation2008; Moen et al. Citation2009; Kumpula et al. Citation2011).
To our knowledge, the ecological impacts of winter feeding of reindeer on vegetation and soil have not been studied earlier. The aim of this investigation was therefore to study the overall impacts of supplementary winter feeding on vegetation and soil in a sub-xeric heath type forest in northern Finland (68°N). Our main focus was to study the potential changes in species coverage in the field- and ground-layer vegetation as well as the N content of dwarf shrubs and soil. In this study, “the overall impacts of winter feeding” include the effect of both forage (grass silage, hay, pellets) and reindeer activity in the form of grazing, trampling, digging and droppings. We proceeded on the basis of three hypotheses. First, winter feeding causes changes in the species coverage of the ground and field layer of the sub-xeric heath forest. Secondly, winter feeding results in increased N content in the dwarf shrubs Vaccinium myrtillus and Empetrum nigrum and in the N content and pH of soil below the humus layer. And thirdly, pellet feeding causes fewer significant effects on vegetation and soil than feeding with grass silage+hay and that grass silage+hay feeding causes fewer effects on plots where uneaten forage was cleared, when compared to uncleared plots. We were also interested in determining when the overall impacts of winter feeding on vegetation and soil become visible. The research utilized an extensive controlled field experiment simulating winter feeding of reindeer over two winters (2008–10) in the Hammastunturi herding cooperative in Finland. The study was planned and conducted in cooperation with the local herders.
Material and methods
Study site
The experiment was conducted approximately 15 km from the village of Kuttura. The village is located in the Hammastunturi herding cooperative (68° 21'N, 26° 50'E), one of 13 Sami reindeer herding cooperatives in northern Finland (). The area is characterized by boreal coniferous forests, sub-Arctic mountain birch woodlands, mires and fells (Oksanen & Virtanen Citation1995). The 35 ha site was selected for this study in cooperation with the local herders. It has a “cold, snowy, forest climate” (Köppen climate type: Dfc) that is characterized by high annual variability. The mean annual temperature, measured from 1970 to 2000, was −0.61°C, with a mean temperature in July of 14.1°C and in January of −13.0°C. Annual precipitation was 426 mm; about half of this is snow, which accumulates to a depth of 65 cm and melts on average by 17 May (data obtained from the Finnish Meteorological Institute).
Fig. 1 Location of the reindeer winter feeding experiment in the Hammastunturi herding cooperative (circle) and of the weather station in Saariselkä (star). In the upper map, the reindeer herding area in Finland is indicated in black.
The largest permissible number of reindeer in the cooperative is 5500 animals. There are 2.5 reindeer/km2. Winter pastures in Hammastunturi contain 12.2 ha of terricolous lichen pasture/reindeer and 16.0 ha of arboreal lichen pasture/reindeer. Some of the forests in the cooperative are protected, and some are used for commercial forestry. The total coverage of infrastructure is 0.8% (17 km2), and its impact area is 6.7% (146 km2) of the land area of the cooperative (Kumpula et al. Citation2009).
Since the middle of the 1960s, the calving percentage (number of calves born per 100 females as calculated during the summer or autumn calf markings) has ranged between 6 and 75%. It reached its minimum in 1973 (6%) and 1974 (18%) due to a sharp decline in the availability of winter forage, caused by difficult snow conditions. In the 2000s, the calving percentage ranged between 44 and 60% (Reindeer Herders’ Association Citation2011; Vuojala-Magga et al. Citation2011).
Winter herding in Hammastunturi entails herding and feeding in the field or in calving fences. Hay, grass silage or pellets are used as feed in winter principally for moving reindeer from one place to another and keeping the herd together. Calving fences, which facilitate safe calving, are used from March/April to the end of May, depending on the annual snow conditions. Herders began using the fences in the mid-1990s because many calves were born on snow or in water holes due to late snowmelt in the spring. In fence calving, female reindeer are collected and kept in large fenced forest areas, which allows herders to ensure that the animals are fit and well. Towards the end of the 2000s, snow began melting earlier and some of the siidas (extended family or kin group consisting of economically independent households) have changed from the calving fence system to the spring herding system, in which reindeer are fed in the forest if snow is deep or hard (Vuojala-Magga et al. Citation2011).
Feeding experiment
The feeding experiment, which simulated supplementary winter feeding of reindeer, was conducted from 2 March to 19 April 2009 (49 days) and from 1 March to 30 April 2010 (61 days). The experimental site (35 ha) is a poorly regenerated pine-dominated sub-xeric heath forest (Empetrum–Myrtillus type [EMT]), with Scots pine seed trees left after cuttings in the 1950–1960s. It is a typical forestry and winter pasture area in the region. The criteria for selecting the site were that it had to provide easy access by snowmobile for herders in winter for daily feeding and by car for researchers in summer for conducting the field measurements and sampling.
In June 2008, 50 experimental plots (Ø 5 m, 19.6 m2) were established on the experimental site (35 ha), and each plot was allocated randomly for one of the following feeding practices: (1) feeding with grass silage+hay with leftovers cleared from the plot in the spring; (2) feeding with grass silage+hay with leftovers not cleared; and (3) feeding with pellets. The experiment also included (4) unfenced and (5) fenced control plots, on which the reindeer were not given supplementary forage. The fences were approximately 2 m high and made of wire mesh. Reindeer had no access to the fenced plots. Each feeding practice was replicated 10 times (total number of plots: 50). The plots were arranged to form an oval; the distance between the closest plots was 30 m and 1 km between the farthest.
Our approach was practical rather than theoretical, which is why no fenced plots with pellets or grass silage+hay were included in the experiment (reindeer herders fed the reindeer during the experiment). As all of the feeding practices in the experiment (except fenced and unfenced control plots) encompassed not only the impact of forage (grass silage+hay, pellets), but also the effect of reindeer on the pastures in the form of grazing, trampling, digging and droppings, we studied the overall effect of winter feeding on various vegetation and soil variables. Another reason for this approach was our expectation that the effects of reindeer on pasture would not be equal in the case of all of the feeding practices. For example, the attractiveness of different types of forage to reindeer varies, with the animals preferring pellets to grass silage+hay.
The forage used in the experiment was a mixture of grass silage preserved with molasses syrup and hay (hereafter “grass silage+hay”); it was produced 50–100 km from the site. The main forage species were Festuca pratensis Hudson, Deschampsia cespitosa (L.) Beauv., Phleum pratense L. and Taraxacum officinale. The forage also contained Trifolium repens L., Rumex acetosella L., Ranunculus acris L., Stellaria media (L.) Vill., Raphanus raphanistrum L., Achillea millefolium L., Rumex acetosa L., Veronica chamaedrys L. and Epilobium angustifolium L. The herders gave reindeer roughly 3 kg of forage/plot/day, which equals 150 g forage/m2/day. The animals were also fed with commercial reindeer pellets (Tähti-Poro Artic and Tähti-Poro 2 Balans, Kinnusen Mylly Ltd., Utajärvi, Finland), which contained oat, wheat, barley and molasses. The herders gave reindeer roughly 1 kg of pellets/plot/day, which equals 50 g pellets/m2/day. The results of the forage and pellet analyses are presented in . According to the reindeer herders participating in this study, the reindeer dug approximately half of their daily forage from the natural pasture.
Table 1 The results of the analysis of grass silage produced in summer 2008 and 2009 and pellets used in the reindeer feeding experiment. The analysis was carried out by Viljavuuspalvelu Ltd. (Mikkeli, Finland).
There were between 100 and 200 reindeer at the experimental site and an additional 150 animals in the surroundings. There was no need to fence the site because the reindeer stayed there to feed. The herders fed the animals daily with grass silage+hay and pellets on the clean snow of the plots, transporting the forage by snowmobile on a track made for that purpose. The reindeer in the surroundings were also fed. Herders distributed about one-third of the supplementary forage (grass silage+hay and pellets) to the plots and spread the remaining two-thirds in the area between the plots and in the surroundings of the experimental site. Accordingly, all reindeer—a maximum of 300 animals—received supplemental forage. On 15 June 2009 and on 26 May 2010, the leftover grass silage+hay, containing mostly dry straw, was removed using a rake from 10 of the 20 plots on which the herders had fed reindeer with grass silage+hay).
Measurements
Vegetation surveys
Vegetation surveys were conducted before the experiment was established (1–13 July 2008) and after the first (8–20 July 2009) and second (4–12 July 2010) winters. The surveys were conducted on two subplots (1 m2) per plot (altogether 2×50 plots=100 subplots) using a point-counting method (Molau & Molgaard Citation1996). In addition to determining the coverage of vascular plants, lichens, mosses and hepatics, the survey recorded coverage of reindeer faeces (see, e.g., Campbell et al. Citation2004; Skarin Citation2007; Moen et al. Citation2009), uneaten forage and bare soil. In addition, the heights of dwarf shrubs and Deschampsia flexuosa were measured. To avoid pseudoreplication, mean values for each plot were calculated on the basis of the respective subplots.
Plant and soil analysis.
Since the chemical composition of plants was expected to reflect eutrophication caused by experimental reindeer feeding before any changes in soil and species coverage, the evergreen dwarf shrub E. nigrum L. (current-year shoots) and the deciduous V. myrtillus L. (current leaves) were chosen as indicator species. Samples were collected from the 50 plots on 25 July 2008, 21 July 2009 and 23 July 2010 for the measurement of C, H and N content (LECO-1000, Leco Corporation, St. Joseph, MI, USA). The measurements were conducted from dry ash samples using a thermal gravimetric analyser with inductively coupled plasma emission spectrometry (TJA Iris Advantage, Thermo Jarrell Ash Corporation, Franklin, MA, USA).
Soil samples for the measurement of pH and C, H and N content were taken from below the humus layer on each experimental plot; pH was measured in the field (Model Testo 206, Lenzkirch, Germany). Soil samples were dried at 40°C, pre-treated with a 2 mm sieve, and analysed for C, H and N content (% dry weight) using the LECO-1000 instrument.
Statistical analysis
We used an analysis of covariance to analyse the effect of reindeer winter feeding on the N concentration and C/N of E. nigrum and V. myrtillus, the coverage (%) of species in the ground and field layer, as well as the pH, N content and C/N of the soil. The data from 2009 (after the first winter) and 2010 (after the second winter) were analysed separately, because we were interested in finding out when the overall impacts of winter feeding on vegetation and soil first become visible. The explanatory variables used in the yearly model were feeding practice (five levels) and the starting values from 2008 (before the experiment was established), the latter being used as a covariate. In addition, we studied the relationship between the coverage of faeces and the coverage and height of D. flexuosa by adding the coverage of faeces into the model and examining only grazed plots (four feeding practice levels). There was no need to make transformations for the response variables, because the diagnostic residual plots were checked and found to be satisfactory.
The tests of the effects of feeding practice and year 2008 (covariate) are presented in tables for 2010 only. The multiple comparisons of the covariate-adjusted feeding practice means were analysed using the Tukey–Kramer method (overall significance level: P=0.05). The feeding practice means in 2008 and the covariate-adjusted means in 2009 and 2010 (with the multiple comparisons) are presented in the figures for the species whose coverage in 2010 exceeded 5%. The MIXED procedure of the SAS statistical software package (version 9.2) was used in the modelling analyses.
Results
Reindeer activity and leftover forage
Mean coverage of faeces, which indicates reindeer activity on the plots, was higher after the first winter (2009) than after the second (2010; a). After both years, the mean coverage of faeces was highest on the plots where reindeer were fed with pellets (9% in 2009 and 5% in 2010), ranging between 2 and 5% for other feeding practices. There were no faeces on the fenced control plots.
Fig. 2 The mean (with standard error) coverage (%) of (a) faeces and (b) uneaten grass silage+hay in the feeding experiment after the first winter (2009) and the second (2010), n=10 plots per feeding practice.
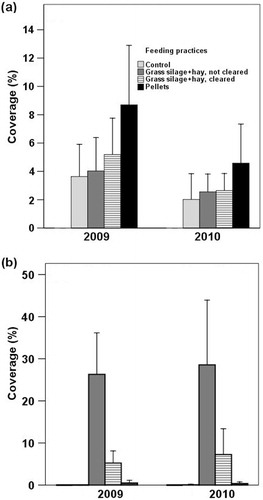
The uneaten grass silage+hay on the plots mainly contained straw. The mean coverage of leftovers on uncleared plots was 26% after the first year (2009) and 29% after the second year (2010) as against 5% and 7% on the cleared plots, respectively (b). There were no leftovers on control plots.
Vegetation surveys
In 2008, before any feeding practices were introduced, the field layer of the pine-dominated sub-xeric heath forest site was characterized by evergreen dwarf shrubs (61%), deciduous dwarf shrubs (14%) and graminoids (14%). The most common evergreen shrub species were E. nigrum (39%), Calluna vulgaris (16%) and Vaccinium vitis-idaea L. (6%) and the most common deciduous shrub was V. myrtillus L. (14%; a–d). Other shrubs, among them V. uliginosum L., Ledum palustre L., Diphasiastrum complanatum (L.) Holub. and Lycopodium clavatum L. were rare (<1%). The only graminoid was D. flexuosa (14%; e). The mean percentage of the site covered by non-vegetated bare soil and litter was 9%. The ground layer exhibited mosses (73%), lichens (21%) and bare soil (6%; f–i). The most common moss and hepatic species were Pleurozium schreberi (34%), Dicranum spp. (26%), Barbilophozia spp. (7%), Hylocomium splendens (5%) and Polytrichum spp. (<1%). Of the lichens present, the most common species was Cladonia rangifera (6%), a type of winter forage preferred by reindeer.
Fig. 3 The mean (with standard error) coverage (%) of (a) Empetrum nigrum, (b) Calluna vulgaris, (c) Vaccinium myrtillus, (d) Vaccinium vitis-idaea and (e) Deschampsia flexuosa in the field layer, and (f) all mosses and hepatics, (g) Pleurozium schreberi, (h) Dicranum sp. and (i) all lichens in the ground layer of a sub-xeric heath forest before the feeding experiment was started (2008), after the first winter (2009) and the second (2010). The figures for 2008 are starting means, and those for 2009 and 2010 covariate-adjusted means (2008 values as a covariate) for feeding practices. The different letters denote differences between the feeding practices at significance level P=0.05, n=10 plots per feeding practice.
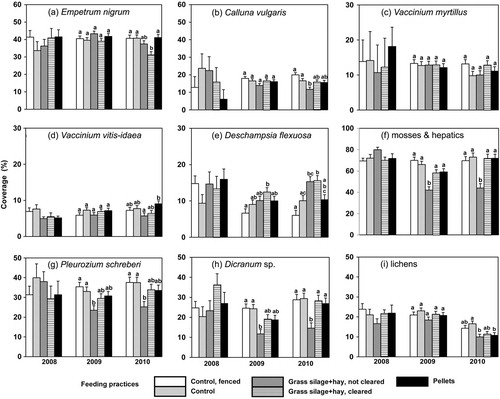
No invasive plant species introduced through grass silage+hay could be observed on the plots after two winters of simulated feeding. The overall effect of feeding, including the effect of forage as well as reindeer on the pasture, did not result in significant differences in the mean coverage of dwarf shrub species after the first winter (2009; a–d). After the second winter (2010), however, there were significant differences between the feeding practices in the mean coverage of the dwarf shrubs E. nigrum, C. vulgaris and V. vitis-idaea. The coverage was lower or tended to be lower on plots where reindeer were fed with grass silage+hay (leftovers not cleared) compared to the control plots (a, b, d, ). In contrast, there was a significant effect of feeding practice on the coverage of the graminoid D. flexuosa after the first winter, in 2009 (feeding practice: F=3.09, P=0.025; e): its coverage increased on plots where reindeer were fed with grass silage+hay (cleared) when compared to fenced controls. After the second winter (2010), the height of D. flexuosa increased significantly on these plots regardless of whether leftovers were cleared when compared to fenced or not fenced controls (feeding practice: F=8.33, P<0.0001; data not shown in the figures). The models, in which both the starting values from 2008 and the feeding practices were taken into consideration, indicated that there was a significant positive linear relationship on herded plots (fenced control plots excluded) between the coverage of faeces and the coverage and height of D. flexuosa. In 2009, the regression coefficient of the model for coverage of D. flexuosa was 0.32 (P=0.030) and for height 0.55 (P=0.032). In 2010, the corresponding regression coefficients were increased to 0.59 (P=0.021) and 0.96 (P=0.039), respectively.
Table 2 F-tests of the effects of feeding practice (10 plots per feeding practice) and the year 2008 (data measured before the experiment was established) on the species coverage in the ground and field layer of the sub-xeric heath type forest after two winters of experimental feeding of reindeer in 2010.
There was a significant effect of feeding practice on the coverage of low-growing mosses and hepatics from the first winter, in 2009 (F=10.96, P<0.0001) onwards (f–h, ), because leftover forage covered them to a considerable extent. For example, the mean coverage of Dicranum spp. after the second winter (2010) was 28% on cleared grass silage+hay plots, but only 15% on uncleared ones (h, ). There were no significant differences in coverage of lichens between the feeding practices after the first year (2009). After the second year (2010), however, the uncleared plots where reindeer were fed with grass silage+hay or pellets had a significantly lower coverage of lichens when compared to unfenced control plots (i, ).
Plant and soil analysis
After the first winter (2009), there was a significant effect in N content of E. nigrum due to feeding practices (F=4.68, P=0.003). The mean N content of E. nigrum was 7–9% (a), and the C/N ratio was 8–9% higher on the plots with grass silage+hay feeding (leftovers not cleared) compared to control plots (fenced/unfenced). After the second winter (2010), the differences due to feeding practices were more pronounced: the mean N content of E. nigrum was 11–14% higher on uncleared plots and 9–13% higher on cleared plots vs. control plots (unfenced/fenced). Feeding reindeer with pellets did not increase the N content of E. nigrum leaves during the experiment (a, ).
Fig. 4 Mean (with standard error) N concentration (%) in leaves of (a) Empetrum nigrum and (b) Vaccinium vitis-idaea, and (c) C/N of soil on the plots in the sub-xeric heath forest before the feeding experiment was started (2008), after the first winter (2009) and after the second (2010). The figures for 2008 are starting means, and those for 2009 and 2010 covariate-adjusted means (2008 values as a covariate) for feeding practices. The different letters denote differences between the feeding practices at significance level P=0.05, n=10 plots per feeding practice.
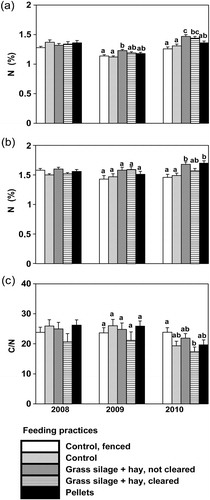
Table 3 F-tests of the effects of feeding practice (10 plots per feeding practice) and year 2008 (data measured before the experiment was established) on N concentration (%) and C/N in E. nigrum, V. myrtillus and soil, and soil pH after two winters of experimental feeding of reindeer in 2010, as well as the multiple comparisons (Tukey–Kramer method) of the covariate-adjusted feeding practice means.
There were no significant differences among the feeding practices in the N content of V. myrtillus after the first winter (2009), but after the second winter (2010), the N content of V. myrtillus was 13–15% higher on uncleared grass silage+hay plots and 14–16% higher on pellet plots when compared to controls (unfenced/fenced); b, ).
No significant effects of feeding practice on pH or on C and N content measured from the soil below the humus layer could be observed during the experiment. The only exception was soil C/N, which exhibited significant effects due to feeding practices after the second winter: the ratio was 27% lower on grass silage+hay plots (leftovers cleared) when compared to fenced control plots (c, ).
Discussion
No invasive species introduced from the grass silage or hay could be observed on the plots during our 2-year experiment. This most probably shows that the grass silage+hay, whose main components are Festuca pratensis Deschampsia cespitosa, Phleum pratense and Taraxacum officinale, was cut before seeding, that the method for producing the silage decreased seed survival or the seeds failed to germinate for some other reason, or that the seedlings had lost their viability due to winter frosts or unsuitable soil. Another reason for the lack of invasive species might be the rather moderate level of supplementary grass silage/hay feeding (ca. 150 g per m2 per day) in combination with the short duration of the experiment (110 days in total during two winters in 2009–10). Our earlier research on three intensively used reindeer winter feeding pens (1–360 reindeer in 0.1–100 ha pens for 4–10 winters) in northern Finland indicates that lichens, mosses and dwarf shrubs become replaced by faster-growing graminoids and herbs; these include N-favouring pioneer species such as Epilobium angustifolium and weeds such as Stellaria media that are introduced either through reindeer forage (grass silage+hay) or from the surroundings of the winter feeding pens (Turunen et al. Citation2010; Turunen & Vuojala-Magga Citation2011). The lack of invasive species in the present study may also be due to the resistance of the sub-xeric heath forest to the overall impacts of forage and reindeer; the appearance of invasive species is dependent on the resistance of the habitat type and/or its ability to resist or recover from erosion caused by grazing, trampling and digging. The habitat types that have the weakest resistance include moist mires and xeric heath forests, whereas mesic and sub-xeric heath forests—the site types in the present study—are more resistant (Kellomäki & Saastamoinen Citation1975; Ukkola Citation1995).
Our experiment showed a significant overall impact of winter feeding on the field and ground-layer vegetation of a sub-xeric pine-dominated heath forest in northern Finland. Since N is the limiting factor of plant growth in nutrient-poor heath forests (Vitousek & Howarth Citation1991), both the fast-growing, N-demanding graminoid D. flexuosa and the slow-growing evergreen dwarf shrub E. nigrum and deciduous dwarf shrub V. myrtillus were able to utilize the increased availability of N compounds. In D. flexuosa, this could be observed already after the first winter both as coverage and height increasing with the coverage of reindeer faeces and as increased coverage where grass silage+hay had been used. In E. nigrum, N content increased on grass silage+hay plots (leftover not cleared in the spring) after the first winter, and in V. myrtillus after the second winter. Earlier studies have indicated that the impacts of reindeer grazing on vegetation and soil depend greatly on reindeer population density, season of grazing and the type and successional state of the site (Väre et al. Citation1995; Väre et al. Citation1996; Suominen & Olofsson Citation2000; Moen et al. Citation2009; Stark et al. Citation2010). In oligotrophic climax-stage Cladonia stellaris-rich pine-dominated forests in northern Finland, heavy grazing has been found to result in development of the understorey vegetation in two directions: towards first Cladonia stage, or towards bryophyte-rich sites having mesotrophic features such as elevated soil N content and pH (Väre et al. Citation1995; Väre et al. Citation1996). The studies of heathlands and tundra in northern Fennoscandia have shown that reindeer may increase the abundance of graminoids and productivity of vegetation through several mechanisms: favouring plants with high-quality litter (Olofsson & Oksanen Citation2002), producing rapidly recycled urine and faeces (van der Wal et al. Citation2004) and increasing soil temperatures (Olofsson et al. Citation2004). In general, moderate grazing has been found to increase diversity on heathlands, whereas diversity has remained unaltered or slightly lower on moderately grazed grass- and herb-dominated meadows (Suominen & Olofsson Citation2000).
Our results show that both reindeer droppings (faeces and urine) and uneaten forage left on the pasture contributed to the increased availability of N compounds in the sub-xeric forest studied. Reindeer faeces and urine provide highly decomposable resources that are rich in labile nutrients, such as ammonium and nitrate, and can stimulate soil microbial biomass and activity, net C and N mineralization, as well as plant N acquisition and growth (Suominen & Olofsson Citation2000; Stark et al. Citation2003; van der Wal et al. Citation2004). The origin of the N compounds derived from the uneaten leftover forage may be natural, that is, a species of forage plant, or artificial, for example, fertilizers, preservatives and/or additives used during the production of grass silage. Proteolysis (breakdown of proteins) in the grass silage causes ammonium-N to be formed, the content of which increases particularly when the sugar content of grass silage is low (McDonald et al. Citation1991; Aagnes & Mathiesen Citation1995; Heiskari & Nieminen Citation2004). The N compounds, particularly ammonium-N, may thus have been absorbed by the vegetation and soil (or adsorbed onto them) under the forage leftover, leading to increased tissue N content in E. nigrum and V. myrtillus leaves. The release of nutrients from reindeer faeces and uneaten forage left on the pasture is greatly dependent on soil moisture (see, e.g., van der Wal et al. Citation2004), most probably being greatest immediately after snowmelt (17 May in the Ivalo—Kuttura area in an average year (data obtained from the Finnish Meteorological Institute) and after persistent rains.
Of the feeding practices applied, feeding with grass silage+hay (leftover not cleared in the spring) most often caused the earliest and most significant overall impacts on vegetation. The low-growing lichens and mosses, such as Dicranum spp., and dwarf shrubs observed in this study are easily covered by reindeer forage leftovers on pasture. Besides being an additional N source, uneaten forage left in thick layers on pasture may directly decrease photosynthesis and thus suppress the growth of ground- and field-layer vegetation. In the spring, a thick layer of uneaten forage with a high straw content may insulate the soil from heat and lead to later melting of soil frost and lower soil temperatures (see Peth & Horn Citation2006). Conversely, forage leftovers may even out the temperature variation of the soil and slow evaporation from the underlying soil and the vegetation, thereby inhibiting the exchange of other gases also.
The present study shows that the overall impact of feeding reindeer with pellets on vegetation and soil below the humus layer was less significant than that caused by feeding with grass silage+hay. On the plots where reindeer were fed with pellets, the species coverage of dwarf shrubs and mosses and hepatics remained unchanged, and N contents in E. nigrum leaves were lower than on the plots where reindeer were fed with grass silage+hay. Reindeer prefer pellets to hay or grass silage, and leftovers are not common. Therefore, the processing of crude hay—into hay pellets, for example—and/or the use of commercial grain-based reindeer pellets could minimize the amount of leftover forage and thus environmental impacts such as eutrophication of vegetation and soil and the spread of invasive species.
Our conclusion is that even a rather moderate level of feeding (ca. 150 g of grass silage+hay and 50 g pellets/m2/day for 49 days in 2009 and 61 days in 2010) may cause changes in vegetation and soil that in the longer term can lead to a gradual shift from a nutrient-poor forest (sub-xeric heath forest type) towards a more nutrient-rich type (mesic heath forest type) (see Raunio et al. Citation2008). This would result in a higher coverage of the N-demanding graminoids that naturally grow in the forest, such as D. flexuosa, or of those derived from the supplementary winter forage of reindeer. It is also likely that the coverage of E. angustifolium, a herb that commonly appears in northern Fennoscandic pine-dominated heath forests after clear-cutting and soil preparation, would increase. Finally, it can be speculated that the impact of winter feeding on forest ecosystems will increase in the future due to predicted climate change. For example, in the summer, increased availability of water and nutrients, especially N compounds in the soil, may improve germination of the seeds of invasive species and the survival and growth of their seedlings (Walck et al. Citation2011). Some of the seeds introduced in reindeer forage, those of graminoids and weeds, for example, may end up in the soil seed bank, where they may retain their germination capacity for decades and germinate only when conditions—for instance, temperature and water availability—become more favourable.
Supplementary Material
Download PDF (146.2 KB)Acknowledgements
We thank reindeer herders Jaana Magga, Jarmo Haataja and Mauno Magga of the Hammastunturi herding cooperative for participating in this study. Special thanks are due to forest engineer Jouni Puoskari at the Rovaniemi Research Unit of the Finnish Forest Research Institute, and MSc Anna Hyyryläinen for their technical assistance during the course of this study. We are also grateful to Kyösti Tikkanen and Laura Räsänen for assisting in the vegetation surveys in summer 2010. Thanks are also due to Richard Foley for checking the language of the manuscript. This research is part of the Reindeer Forage and Supplementary Feeding in Changing Climate project (2008–10), which was funded by the Ministry of Agriculture and Forestry (Development Fund of Agriculture and Forestry; project no. 2941/503/2007), Kinnusen Mylly Ltd. and the Arctic Centre of the University of Lapland.
References
- Aagnes T.H. Mathiesen S.D. Round baled grass silage as food for reindeer in winter. Rangifer. 1995; 15: 27–35. 10.3402/polar.v32i0.18610.
- Åhman B. & Danell Ö. 2006. Can supplementary feeding improve productivity in reindeer husbandry?. In: L.Bartoš, et al..: Advances in deer biology. Deer in a changing world. Proceedings of the 6th International Deer Biology Congress: Prague, Czech Republic. , 2006. Pp. 30–32. Prague: Research Institute of Animal Production.
- Aschfalk A. Josefsen T.D. Steingass H. Müller W. Goethe R. Crowding and winter emergency feeding as predisposing factors for kerato-conjunctivitis in semi-domesticated reindeer in Norway. Deutsche Tierarztliche Wochenschrifte. 2003; 110: 295–298.
- Callaghan T.V. Johansson M. Brown R.D. Groisman P.Y. Labba N. Radionov V. Barry R.G. Bulygina O.N. Essery R.L.H. Frolov D.M. Golubev V.N. Grenfell T.C. Petrushina M.N. Razuvaev V.N. Robinson D.A. Romanov P. Shindell D. Shmakin A.B. Sokratov S.A. Warren S. Yang D. The changing face of Arctic snow cover: a synthesis of observed and projected changes. Ambio. 2011; 40: 17–31. 10.3402/polar.v32i0.18610.
- Campbell D. Swanson G.M. Sales J. Comparing the precision and cost effectiveness of faecal pellet group count methods. Journal of Applied Ecology. 2004; 41: 1185–1196. 10.3402/polar.v32i0.18610.
- Eloranta E. & Nieminen M. 1986. Calving of the experimental reindeer herd in Kaamanen during 1970–85. Rangifer 1. (Special Issue), 115–121.
- Forbes B.C., Bölter M., Müller-Wille L., Hukkinen J., Müller F., Gunslay N. & Konstantinov Y. 2006. Reindeer management in northernmost Europe. Linking practical and scientific knowledge in social–ecological systems. Ecological Studies 184. Berlin: Springer.
- Hansen B.B. Aanes R. Herfindal I. Kohler J. Saether B.-E. Climate, icing, and wild Arctic reindeer: past relationships and future prospects. Ecology. 2011; 92: 1917–1923. 10.3402/polar.v32i0.18610.
- Heiskari U. & Nieminen M. 2004. Erilaiset nurmirehut porojen talviruokinnassa. (Different grass fodders in the winter feeding of reindeer.) Kala-ja Riistaraportteja 314. Helsinki: Finnish Game and Fisheries Research Institute.
- Helle T. Kojola I. Demographics in an alpine reindeer herd: effects of density and winter weather. Ecography. 2008; 31: 221–230. 10.3402/polar.v32i0.18610.
- Jernsletten J.L.L. Klokov K. Sustainable reindeer husbandry. Centre for Sami Studies, University of Tromsø. TromsøNorway, 2002
- Kellomäki S. Saastamoinen V.L. Trampling tolerance of forest vegetation. Acta Forestalia Fennica. 1975; 147: 1–22.
- Kumpula J. Stark S. Holand Ø;.Seasonal grazing effects by semi-domesticated reindeer on subarctic mountain birch forests. Polar Biology. 2011; 34: 441–453. 10.3402/polar.v32i0.18610.
- Kumpula J, Tanskanen A, Colpaert A, Anttonen M, Törmänen H, Siitari J. & Siitari S. 2009. Poronhoitoalueen pohjoisosan talvilaitumet vuosina 2005–2008—laidunten tilan muutokset 1990—luvun puolivälin jälkeen. (Winter pastures in the northern part of the reindeer management area in Finland during 2005–2008—changes in the state of pastures after the mid-1990s.) Riista-ja kalatalous—Tutkimuksia 3. Helsinki: Finnish Game and Fisheries Research Institute.
- Maijala V. Nieminen M. Winter feeding of reindeer—experiences and results from Finland. Rangifer Report. 2001; 5: 15–16.
- Mattila E. & Mikkola K. 2009. Poronhoitoalueen etelä- ja keskiosien talvilaitumet. Tila paliskunnissa 2000-luvun alkuvuosina ja eräiden ravintokasvien esiintymisrunsauden muutokset merkkipiireissä 1970-luvulta lähtien. (Winter pastures in southern and central parts of the reindeer herding area in Finland. State of the herding districts in the beginning of the 2000s and changes in abundancy of some forage plants in reindeer marking districts from 1970s onwards.) Metlan työraportteja 115. Vantaa: Finnish Forest Research Institute.
- McDonald P. Henderson A.R. Heron S.J.E. The biochemistry of silage. Chalcombe. MarlowUK, 1991
- Moen J. Climate change: effects on the ecological basis of reindeer husbandry in Sweden. Ambio. 2008; 37: 304–311. 10.3402/polar.v32i0.18610.
- Moen J. Boogerd C. Skarin A. Variations in mountain vegetation use by reindeer (Rangifer tarandus) affects dry heath but not grass heath. Journal of Vegetation Science. 2009; 20: 805–813. 10.3402/polar.v32i0.18610.
- Molau U. Mølgaard P. ITEX manual. International Tundra Experiment. Danish Polar Center. Copenhagen, 1996
- Nieminen M. Why supplementary feeding in Finland?. Rangifer Report. 2010; 14: 40–41.
- Oksanen A. Åsbakk K. Nieminen M. Norberg H. Näreaho H. Antibodies against Toxoplasma gondii in Fennoscandian reindeer—association with the degree of domestication. Parasitology International. 1997; 46: 255–261. 10.3402/polar.v32i0.18610.
- Oksanen L. Virtanen R. Topographic, altitudinal and regional patterns in continental and suboceanic heath vegetation of northern Fennoscandia. Acta Botanica Fennica. 1995; 153: 1–80.
- Olofsson J. Kitti H. Rautiainen P. Stark S. Oksanen L. Effects of summer grazing by reindeer on composition of vegetation, productivity and nitrogen cycling. Ecography. 2001; 24: 13–24. 10.3402/polar.v32i0.18610.
- Olofsson J. Oksanen L. Role of litter decomposition for the increased primary production in areas of heavily grazed by reindeer: a litter bag experiment. Oikos. 2002; 96: 507–515. 10.3402/polar.v32i0.18610.
- Olofsson J. Oksanen L. Effects of reindeer density on vascular plant diversity on north Scandinavian mountains. Rangifer. 2005; 25: 5–18. 10.3402/polar.v32i0.18610.
- Olofsson J. Stark S. Oksanen L. Reindeer influence on ecosystem processes in the tundra. Oikos. 2004; 105: 386–396. 10.3402/polar.v32i0.18610.
- Pajunen A. Virtanen R. Roininen H. The effects of reindeer grazing on the composition and species richness of vegetation in forest-tundra ecotone. Polar Biology. 2008; 31: 1233–1244. 10.3402/polar.v32i0.18610.
- Peth S. & Horn R. 2006Consequences of grazing on soil physical and mechanical properties in forest and tundra environments. In: B.C.Forbes, et al..: Reindeer management in northernmost Europe. Linking practical and scientific knowledge in social–ecological systems. Ecological Studies 184. Pp. 218–291. Berlin: Springer.
- Putkonen J. & Roe G. 2003. Rain-on-snow events impact soil temperatures and affect ungulate survival. Geophysical Research Letters 30. article no. 1188. 10.3402/polar.v32i0.18610.
- Raunio A. Schulman A. Kontula T. Suomen luontotyyppien uhanalaisuus. Osa 2: Luontotyyppien kuvaukset. (Assessment of threatened habitat types in Finland. Part 2: Habitat type descriptions.) Suomen ympäristö 8/2008. Finnish Environment Institute. Luonto. Helsinki, 2008
- Reindeer Herders’ Association. 2011. Annual statistics of reindeer numbers for the period 1965–2011. (See Supplementary File for details about this source.)RovaniemiFinland: Reindeer Herders’ Association.
- Säkkinen H. Timisjärvi J. Eloranta E. Heiskari U. Nieminen M. Pukka M. Nutrition-induced changes in blood chemical parameters of pregnant reindeer hinds. (Rangifer tarandus tarandus.). Small Ruminant Research. 1999; 32: 211–221. 10.3402/polar.v32i0.18610.
- Skarin A. Habitat use by semi-domesticated reindeer, estimated with pellet-group counts. Rangifer. 2007; 27: 121–132. 10.3402/polar.v32i0.18610.
- Solomon S., Qin D., Manning M., Chen Z., Marquis M., Averyt K.B., Tignor M. & Miller H.L. Jr. 2007. Climate change. The physical science basis: contribution of Working Group I to the fourth assessment report of the Intergovernmental Panel on Climate Change. Cambridge: Cambridge University Press.
- Stark S. Männistö M. Smolander A. Multiple effects of reindeer grazing on the soil processes in nutrient-poor northern boreal forests. Soil Biology and Biochemistry. 2010; 42: 2068–2077. 10.3402/polar.v32i0.18610.
- Stark S. Tuomi J. Strömmer R. Helle T. Non-parallel changes in soil microbial carbon and nitrogen dynamics due to reindeer grazing in northern boreal forests. Ecography. 2003; 26: 51–59. 10.3402/polar.v32i0.18610.
- Suominen O. Olofsson J. Impacts of semi-domesticated reindeer on structure of tundra and forest communities in Fennoscandia: a review. Annales Zoologi Fennici. 2000; 37: 233–249.
- TØmmervik H, Bjerke J.W, Gaare E, Johansen B. & Thannheiser D. 2012. Rapid recovery of recently overexploited winter grazing pastures for reindeer in northern Norway. Fungal Ecology. 5, 3–15. 10.3402/polar.v32i0.18610.
- Tryland M. Josefsen T. Oksanen A. Aschfalk A. Contagious ecthyma in Norwegian semi-domesticated reindeer (Rangifer tarandus tarandus). Veterinary Record. 2001; 149: 394–395. 10.3402/polar.v32i0.18610.
- Turunen M. Oksanen P. Markkula I. Vuojala-Magga T. Sutinen M.-L. Maijala J. Tuomaala R. Vegetation changes in reindeer winter corrals in Finnish Lapland. Rangifer Report. 2010; 14: 110–111.
- Turunen M. Soppela P. Kinnunen H. Sutinen M.-L.&;Martz F. Does climate change influence the availability and quality of reindeer forage plants? A review. Polar Biology. 2009; 32: 813–832. 10.3402/polar.v32i0.18610.
- Turunen M. Vuojala-Magga T. Poron ravinto ja lisäruokinta muuttuvassa ilmastossa. (Reindeer forage and supplementary feeding in a changing climate.) Arktisen keskuksen tiedotteita 56. Arctic Centre, University of Lapland. RovaniemiFinland, 2011
- Tyler N.J.C. Turi J.M. Sundset M.A. Stom Bull K. Sara M.N. Reinert E. Oskal N. Nellemann C. McCarthy J.J. Mathiesen S.D. Martello M.L. Magga O.H. Hovelsrud G.K. Hanssen-Bauer I. Eira N.I. Eira I.M.G. Corell R.W. Saami reindeer pastoralism under climate change: applying a generalized framework for vulnerability studies to a sub-Arctic social-ecological system. Global Environmental Change. 2007; 17: 191–206. 10.3402/polar.v32i0.18610.
- Ukkola R. Trampling tolerance of plants and ground cover in Finnish Lapland, with an example from the Pyhätunturi National Park. Zeszyty Naukowe Uniwersytetu Jagiellonskiego. 1995; 98: 91–110.
- van der Wal R. Bardgett R.D. Harrison K.A. Stien A. Vertebrate herbivores and ecosystem control: cascading effects of faeces on tundra ecosystems. Ecography. 2004; 27: 242–252. 10.3402/polar.v32i0.18610.
- Väre H. Ohtonen R. Mikkola K. The effect and extent of heavy grazing by reindeer in oligotrophic pine heaths in northeastern Fennoscandia. Ecography. 1996; 19: 245–253.
- Väre H. Ohtonen R. Oksanen L. Effects of reindeer grazing on understorey vegetation in dry Pinus sylvestris forests. Journal of Vegetation Science. 1995; 6: 523–530. 10.3402/polar.v32i0.18610.
- Vitousek P.M. Howarth R.W. Nitrogen limitation on land and in the sea: how can it occur?. Biogeochemistry. 1991; 13: 87–115. 10.3402/polar.v32i0.18610.
- Vuojala-Magga T. Turunen M. Ryyppö T. Tennberg M. Resonance strategies of Sami reindeer herders in northernmost Finland during climatically extreme years. Arctic. 2011; 64: 227–241.
- Walck J.L. Hidayati S.N. Dixon K.W. Thompson K. Poschlod P. Climate change and plant regeneration from seed. Global Change Biology. 2011; 17: 2145–2161. 10.3402/polar.v32i0.18610.
- Weladji R.B. Holand O. Influences of large-scale climatic variability on reindeer population dynamics: implications for reindeer husbandry in Norway. Climate Research. 2006; 32: 119–127. 10.3402/polar.v32i0.18610.