
Abstract
The aim of this study was to estimate the genetic parameters and to predict experimental breeding values (EBVs) for saturated (SFA), unsaturated (UFA), monounsaturated (MUFA) and polyunsaturated (PUFA) fatty acids, the ratio of fatty acids, and the productive traits in Italian Brown Swiss (BSW) and Holstein Friesian (HOL) cattle. Test-day yields from 235,658 HOL and 21,723 BSW cows were extracted from the Italian HOL and BSW Associations databases from November 2009 to October 2012 out of 3310 herds. The milk samples collected within the routine milk recording scheme were processed with the Milkoscan™ FT 6500 Plus (Foss, Hillerød, Denmark) for the identification of SFA, UFA, MUFA and PUFA composition in milk. Genetic parameters for fatty acids and productive traits were estimated on 1,765,552 records in HOL and 255,592 records in BSW. Heritability values estimated for SFA, UFA, MUFA and PUFA ranged from 0.06 to 0.18 for the BSW breed and from 0.10 to 0.29 for HOL. The genetic trends for the fatty acids were consistent between traits and breeds. Pearson’s and Spearman’s correlations among EBVs for SFA, UFA, MUFA and PUFA and official EBVs for fat percentage were in the range 0.32 to 0.54 for BSW and 0.44 to 0.64 for HOL. The prediction of specific EBVs for milk fatty acids and for the ratio among them may be useful to identify the best bulls to be selected with the aim to improve milk quality in terms of fat content and fatty acid ratios, achieving healthier dairy productions for consumers.
Introduction
Industry and consumers’ attention is turning towards the use of novel tools to assess the quality and nutritional properties of food products. Consistently with consumers’ efforts to improve the healthfulness of their diets and in order to satisfy their demands, the benchmark of milk quality now includes also the specific composition of the lipid and protein fractions in milk.
Bovine milk contains about 4% of lipids, comprising fatty acid with chains of different length (4 to 24 carbon atoms), and saturation, that is the presence or absence of double bonds in the carboxylic chain. More than half of the milk fats (almost 70%) are saturated fatty acids (SFA) while the remaining part is composed by unsaturated fatty acids (UFA), of which 80% are monounsaturated (MUFA) and 20% are polyunsaturated (PUFA) fatty acids (Lindmark Månsson, Citation2008). Among milk fatty acids, SFA are considered harmful for human health, since they are involved in the development of heart diseases, weight gain and obesity (Haug et al., Citation2007). Conversely, MUFA and PUFA have a blood cholesterol lowering effect in humans (Livingstone et al., Citation2012). The impact of cow nutrition on milk fat composition is well described in the literature (Butler et al., Citation2008; Wiking et al., Citation2010), but a lower number of studies is available on the genetic basis of fatty acid variation. At present, milk components included in the dairy cattle selection objectives are limited to fat, protein, and lactose and the genetic variability of a large part of the milk component is still unknown, especially for those where an expensive analysis is needed to measure these specific elements.
The availability of accurate methods to quantify the milk fatty acid composition on a large-scale may be helpful to: i) commercialise higher-value milk with improved nutritional characteristics through a specific food chain production with specific nutritional claims as the Bleu-Blanc-Coeur (http://www.bleu-blanccoeur.com/) in France; ii) develop a payment system for milk according to customer requirements; iii) for farmers, adapt the feeding systems of their animals accordingly. Such tools may be also useful in long-term breeding strategies to improve specific fatty acid components according to market requirements. Midinfrared (MIR) spectrometry is a powerful and quick methodology for the quantification of the major components of bovine milk such as protein and fat percentages (Soyeurt et al., Citation2010). Milk constituents are identified and quantified based on the absorption of a specific infrared radiation frequency typical of each molecule. Quantification of milk components is based on calibration equations specifically developed for each trait of dairy cattle milk. This technology is faster and cheaper than the classical reference chemical analysis. Moreover, it is commonly used to predict fatty acids composition of bovine milk (De Marchi et al., Citation2011; Soyeurt et al., Citation2006) and other components, validating its potential for the measurement of fatty acids in dairy products. Only few studies have determined the genetic variability of milk fatty acids in large data sets in order to assess the feasibility of selection programmes including fatty acid composition as selection objective (Bobe et al., Citation2008; Gion et al., Citation2011; Soyeurt et al., Citation2007b). In these studies emphasis is also given to the ratios among fatty acids, since the altering of the SFA to UFA ratio (SFA:UFA) causes changes in oxidative stability of milk fat and of its sensorial properties, with consequences on butter texture and whipping properties of cream (Hillbrick and Augustin, Citation2002; Gonzalez et al., Citation2003; Henning et al., Citation2006). Soyeurt et al. (Citation2007a) showed that SFA:UFA varied throughout the lactation with a rapid increase of the values of SFA:UFA until 100 days-in-milk (DIM), when the ratio was near 2 and a decrease to 1.5 at 365 DIM. The importance of the fatty acid ratio in milk can be also highlighted by studies in human nutrition reporting that diets with low PUFA to SFA ratio (PUFA:SFA) were correlated with fatal heart attacks in males, type 2 diabetes (Soinio et al., Citation2003) and lower extremity arterial disease in males (Zheng et al., Citation1997).
The aims of this study were: i) the estimation of the genetic parameters for SFA, UFA, MUFA, PUFA, ratio of fatty acids and productive traits; ii) a pilot prediction of breeding values (EBVs) for SFA, UFA, MUFA, PUFA and productive traits in Italian Holstein Friesian (HOL) and Italian Brown Swiss (BSW) dairy cattle.
Materials and methods
Data
Test-day (TD) yields from 235,658 HOL and 21,723 BSW cows were extracted from the Italian Holstein Friesian Association (ANAFI) and the Italian Brown Swiss Association (ANARB) database. Data collection spanned from November 2009 to October 2012 in 3310 herds (1099 and 2211 for BSW and HOL, respectively) collected within the national routine milk recording system. The TD collected were 1,765,552 and 255,592 for the HOL and the BSW breed, respectively. Data recording was limited to the Lombardy region in the North of Italy. Milk samples were processed at the Association of Lombardy Region Breeders (ARAL) with the Milkoscan™ FT 6500 Plus instrument (Foss, Hillerød, Denmark) for the identification of SFA, UFA, MUFA and PUFA composition in milk through MIR spectrometry. The calibration curves of the Milkoscan™ FT 6500 Plus (Foss) for the identification and quantification of specific groups of milk fatty acids were developed by ARAL in collaboration with Foss. The curves were built comparing the fatty acid concentrations of 80 randomly chosen milk samples analysed both with the MIR spectrometry and in gas chromatography (GC). The milk fatty acid concentration was obtained with the GC Thermo Finningan TRACE GC equipped with a DB-WAXETR (Agilent Technologies, Santa Clara, CA, USA) fused silica capillary column (30 m x 0.32 x 0.25 μm). The methodologies used in the stages of preparation, extraction and separation for the gas chromatographic determination of the fatty acid composition of milk fat were carried out following the standards, respectively: ISO 14156/FIL 172; ISO 15884/IDF 182; ISO 23065/IDF 211 and ISO 15885 182/IDF 184 (ISO, Citation2001, Citation2002a, Citation2002b, Citation2009). The conditions for the chromatographic analyses were as follows: carrier gas, helium, flame-ionisation detector at 250°C and the injected volume was 0.5 μL. In order to determine the quantity of milk samples fatty acids, the F.A.M.E. Mix C4-C24 (Sigma Aldrich, St. Louis, MO, USA) was used. The traits considered were: the content of the four categories of fatty acids (SFA, UFA, MUFA, PUFA), milk yield (MY), protein yield and contents (PY and P%, respectively), fat yield and contents (FY and F%, respectively), casein content (C%), urea content (UR), lactose content (L%), milk somatic cell count (SCC), and the ratios among the contents of specific categories of fatty acids (UFA:SFA; MUFA:SFA; PUFA:SFA). Only lactations with DIM between 5 and 400 and with at least 3 TD records were included in the analysis and TD records were edited to remove outliers by discarding those records with individual fatty acids content more than 2 SD from the mean. Means and standard deviations were calculated separately for the two breeds. The maximum number of individual TD records per lactation resulted to be 13 for BSW and 15 for HOL. The two final datasets, one for each breed, included 154,467 TD records for 16,680 BSW cows reared in 982 herds, and 1,171,022 TD records for 170,914 HOL cows reared in 1121 herds. A summary of the data is shown in . Genealogical information was extracted from the herd books of each breed including all known ancestors for a total of 47,994 individuals for BSW and 273,290 individuals for HOL.
Genetic parameters estimates
Environmental factors included in the model for the variance component estimation were previously tested for their significance with the GLM procedure of SAS 9.3 (SAS, Citation2011) separately for both BSW and HOL breeds. The estimation of the (co)variance components and the prediction of the EBVs for productive traits and acidic profile traits were obtained using the VCE 6.0 package (Groeneveld et al., Citation2010), based on a REML algorithm. Each breed was considered separately also in the variance component estimation analysis. Data were analysed in a series of 16 single-trait TD repeatability animal models applied to the following traits: MY, PY, P%, FY, F%, C%, UR, L%, SCC, SFA (%), UFA (%), MUFA (%), PUFA (%), SFA:UFA, SFA:MUFA and SFA:PUFA. To reduce computer memory requirements, the estimation of the genetic correlations among traits was performed through a bivariate analysis. The model used for the estimation of the (co)variance components included the fixed effects of herd (982 levels for BSW and 1121 for HOL), month*year of calving (44 levels for BSW and 43 for HOL), age at calving within parity (17 levels), DIM (30 days classes resulting in13 levels), and season (4 levels). Additive genetic effect (47,994 animals for the BSW and 273,290 for the HOL) and the permanent environmental effect (16,680 and 107,914 animals with production for BSW and HOL, respectively) were considered as random. Experimental EBVs for each trait were predicted in both breeds with the same TD repeatability animal model used for the variance component estimations. The REMLF90 software (Misztal et al., Citation2002) was used to estimate the prediction error variance (PEV) of EBVs. The reliabilities of the EBVs (RELEBV) were calculated according to Mrode (Citation2005), as: RELEBV=1-(PEV/σ2a). The correlations among the EBVs here predicted (adjusted for RELEBV) and the official EBVs for fat % available for the breeds were calculated. The official EBVs for fat were provided from ANAFI and ANARB.
Results and discussion
A linear relationship was found between milk fatty acids concentration obtained using the Milkoscan™ FT 6500 Plus (Foss) and the gas chromatography, achieving correlations between the two methodologies equal to 99% for the SFA and 94% for the UFA ( and ). The descriptive statistics of productive traits and milk fatty acids content for the BSW and the HOL breeds are reported in . Holstein cows had the greatest milk production, whereas BSW cows on average showed higher milk fat and protein contents than HOL cows. Daily MY was 21.52 kg/d in BSW and 30.03 kg/d in HOL, with contents of 3.64 and 3.40 of P% and 3.99 and 3.71 of F% in BSW and HOL, respectively. These values are in agreement with the average yields of the cows registered in the national herd books and reported in the Italian Breeders Association (AIA) bulletins (AIA, Citation2012). The greatest differences in milk fat composition between the breeds was observed for SFA content with values of 2.67 for BSW and 2.48 for HOL, while the lowest differences were in the group of PUFA content, with values of 0.17 for BSW and 0.16 for HOL. Concerning the saturation of milk fat, SFA were the most represented fatty acids in milk, followed by MUFA and PUFA. The fatty acids profiles of both breeds agreed with the results reported in other studies (Soyeurt et al., Citation2007a, Citation2007b; De Marchi et al., Citation2011) and, as reported by Grummer (Citation1991), SFA, UFA, MUFA and PUFA represented respectively about the 70, 30, 22 and 5% of the total fat for both the breeds. The values of the ratios among specific fatty acids were in agreement with results reported by Soyeurt et al. (Citation2008). The heritability values here estimated for all the traits are reported in . Heritability values estimated for SFA, UFA, MUFA and PUFA, ranged from 0.06 to 0.18 for the BSW breed and from 0.10 to 0.29 for HOL. In both breeds the lowest values of heritability, among fatty acids, was for MUFA (with values of 0.06 for BSW and 0.10 for HOL), while the highest was for the SFA (with values of 0.18 and 0.29 for BSW and HOL, respectively). Heritability estimates of specific fatty acid were in agreement with those from Bobe et al. (Citation2008), Soyeurt et al. (Citation2007a) and Bastin et al. (Citation2013) who reported heritability estimates for daily yields of SFA and with higher values than those estimates for MUFA and PUFA. Estimated heritability values for productive traits obtained in this study were 0.06 (MY), 0.14 (F%), 0.25 (P%) for BSW and 0.10 (MY), 0.24 (F%) and 0.33 (P%) for HOL. In general, estimated values for heritability of productive traits were smaller than values reported in the literature, but this may be due to the statistical model definitions, the breeds involved, the number of available records per animal, the data structure and the editing procedures (Miglior et al., Citation2007; Stoop et al., Citation2007; Cecchinato et al., Citation2011; Samoré et al., Citation2012). As reported by Clement et al. (Citation2001), in fact, an important source of difference may lie in the choice of the genetic model used to analyse data, and the data structure may be a second important source of difference likely to affect the estimation of variance components. The estimated heritability values for L%, U%, C% and SCC () turned out to be similar to those obtained in the BSW and HOL breeds by Samorè et al. (Citation2012). The estimated heritability values of the ratios UFA:SFA, MUFA:SFA and PUFA:SFA () resulted smaller in both breeds than those reported by Soyeurt et al. (Citation2008), but their results were referred to daily heritability values estimates, while in the present study the heritability values were related to the whole lactation. In general, the values of repeatability estimated for all the traits included in this study, ranged from 0.11 to 0.46 for BSW breed and from 0.17 to 0.53 for the HOL cows. The genetic correlation values estimated for all the traits in the two breeds are reported in and .
Genetic correlations between fatty acids and F% were strong and positive, varying from 0.57 to 0.97 for BSW, and from 0.67 to 0.98 for HOL ( and ). Thus, an increase in fat content would increase all fatty acids content in milk. The highest genetic correlation of F% was with SFA (with genetic correlation estimated values of 0.98 and 0.97 in the HOL and BSW breed, respectively). The genetic correlation estimated values among fatty acids and P% were all positive and ranged from 0.38 to 0.61 for BSW, and from 0.45 to 0.64 for HOL. The strongest correlations of fatty acids with P% resulted for PUFA, both in BSW (0.61) and in HOL breed (0.64). Positive correlations between protein and fatty acids contents reflected the general high genetic correlation between fat and protein contents (0.62 for BSW and 0.50 for HOL; data not shown). In both breeds, the genetic correlations estimates between the four categories of fatty acids and L% were positive, while the estimated genetic correlations with SCC were negative. The UFA was highly correlated with the MUFA and PUFA (with estimated genetic correlations of 0.90 and 0.61 for BSW and 0.95 and 0.72 for HOL), probably because of the similar underlying metabolic processes. Genetic correlations estimates between MY and the production traits, except for FY and PY, were negative, confirming the results of several studies (Soyeurt et al., Citation2007b; Samorè et al., Citation2012; Miglior et al., Citation2007). Negative genetic correlations between MY and milk constituents contents (fat, protein, lactose and urea) are related to dilution as the relative amount of milk components tends to decline physiologically when milk production increases (Soyeurt et al., Citation2007b). Genetic correlations estimates between the ratio UFA:SFA, MY and P% were slightly lower than those reported by Soyeurt et al. (Citation2008) for both breeds, while correlations between the ratio UFA:SFA and F% were much more similar to results obtained by the same authors. The gative genetic correlation estimates between MUFA:SFA and MY (-0.17 for BSW, -0.16 for HOL) indicated that cows genetically predisposed to have high milk production, may produce milk with more MUFA content or less SFA than less productive cows. The phenotypic correlation estimates (data not shown) of the traits analysed in the present study were in the same direction as the genetic estimates. Experimental EBVs were predicted in both breeds using the same TD repeatability animal model used for the variance component estimations. In HOL the model differed from the random regression TD model officially used by ANAFI that accounts for the genetic variability over DIM and parities (Muir et al., Citation2007). In general, the correlations between official EBVs for F% calculated by ANARB and ANAFI and those estimated in the present study were positive (0.56 and 0.64 for BSW and HOL, respectively). The genetic trends for the different specific fatty acids groups were almost overlapping with very few differences between traits and breeds. Correlations (Pearson’s and Spearman’s) between EBVs for SFA, UFA, MUFA and PUFA and official EBV for F% were in the range 0.32 to 0.54 for BSW, while the same correlations calculated for HOL were in the range 0.44 to 0.64. These correlations, though not high and despite the great differences in the dataset sizes and in their structures, and in the statistical model definition used in case of HOL, indicated that even if the data used in the study are only a subsample of the overall Italian genetic evaluation the ranking of individuals the EBVs here found follow the same trend of the overall populations in both breeds.
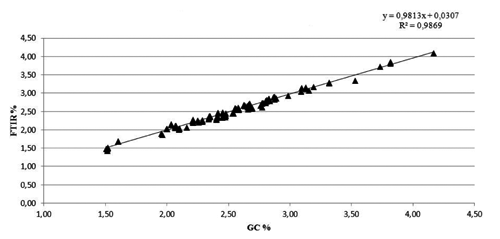
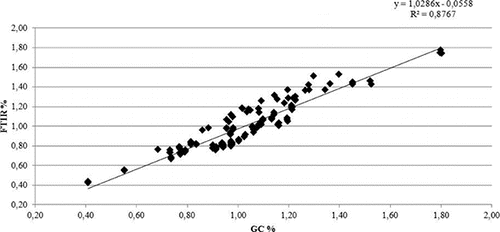
Table 1. Number of records, lactations, cows and the average number of test-day records per parity per cow for the Italian Brown Swiss and the Holstein Friesian cattle.
Table 2. Mean and standard deviation of milk fatty acids and other productive traits in Italian Brown Swiss and Holstein Friesian cows.
Table 3. Heritability and repeatability values within parity estimated with the test-day repeatability model in Italian Brown Swiss and Holstein Friesian cows.
Table 4. Genetic correlation values estimated for all the traits and fatty acids in the two groups of cows: Italian Brown Swiss and Holstein Friesian.
Table 5. Genetic correlation values estimated for all the traits and fatty acids ratios in the two groups of cows: Italian Brown Swiss and Holstein Friesian.
Conclusions
Thanks to this study, the genetic parameters for SFA, UFA, MUFA and PUFA are available and these traits may be integrated in the selection index of the Italian BSW and HOL breeds. Furthermore, the implementation of routinely collecting MIR spectra to determine detailed milk fatty acids, jointly with the increased interest on nutritional and technological properties of milk, would allow to develop genetic evaluations for these traits in the HOL and BSW populations. The genetic variability for fatty acids contents in milk found in both breeds resulted in heritability estimated with moderate values in both dairy cattle populations, indicating that fatty acid profile may be changed by genetic selection. From the results obtained in this study the selection for milk fatty acids seems promising. The prediction of specific EBVs for milk fatty acids and their ratio would be useful to identify the best bulls to be selected to improve the milk quality for fat content and fatty acid ratios, achieving, in this way, healthier dairy productions for the consumers. The genetic correlations among fatty acids were high, suggesting that the selection for a specific group of fatty acid would affect the whole fatty acid profile in milk.
However, the economic advantage of a breeding goal that includes fatty acids is still to be assessed. At present in Italy there is no a definite market of milk with increased content of specific fatty acids, but examples can be found in the French market (http://www.bleu-blanc-coeur.com/) and farmers from Lombardy region are envisaging such possibility. With current market conditions the decision to include fatty acid traits in the economic selection indexes of the two breeds seems not advisable given that this will affect genetic gain of other traits, like milk production, without certain economic advantages. Nevertheless, future changes in selection aims, according to markets looking for milk with the presence of specific components or nutritional properties, would lead to the revision of the selection indexes of the two breeds with specific emphasis also to different fatty acids. At present niche markets requesting milk with specific nutritional properties may advise farmers to develop specific milk production where the analyses on fatty acid content may guarantee the added value.
Acknowledgements
This study was part of the LattOmega project supported by Lombardy region, Italy (Project n.1328).
Related Research Data
References
- AIA, 2012. Bollettino dei controlli della produttività del latte in Italia. Statistiche ufficiali. Associazione Italiana Allevatori Ed., Roma, Italy.
- BastinC. SoyeurtH. GenglerN., 2013. Genetic parameters of milk production traits and fatty acid contents in milk for Holstein cows in parity 1-3. J. Anim. Breed. Genet. 130:118-127.
- BobeG. Minick BormannJ.A. LindbergG.L. FreemanA.E. BeitzD.C., 2008. Short communication: estimates of genetic variation of milk fatty acids in US Holstein cows. J. Dairy Sci. 91:1209-1213.
- ButlerG. NielsenJ.H. SlotsT. SealC. EyreM.D. SandersonR. LeifertC., 2008. Fatty acids and fat-soluble antioxidant concentration in milk from high- and low-input conventional and organic systems: seasonal variation. J. Sci. Food Agr. 88:1430-1441.
- CecchinatoA. PenasaM. De MarchiM. GalloL. BittanteG. CarnierP., 2011. Genetic parameters of coagulation properties, milk yield, quality, and acidity estimated using coagulating and noncoagulating milk information in Brown Swiss and Holstein-Friesian cows. J. Dairy Sci. 94:4205-4213.
- ClementV. BibeB. VerrierE. ElsenJ.M. ManfrediE. BouixJ. HanocqE., 2001. Simulation analysis to test the influence of model adequacy and data structure on the estimation of genetic parameters for traits with direct and maternal effects. Genet. Sel. Evol. 33:369-395.
- De MarchiM. PenasaM. CecchinatoA. MeleM. SecchiariP. BittanteG., 2011. Effectiveness of mid-infrared spectroscopy to predict fatty acid composition of Brown Swiss bovine milk. Animal 5:1653-1658.
- GionA. LarroqueH. BrochardM. LahalleF. BoichardD., 2011. Genetic parameter estimation for milk fatty acids in three French dairy cattle breeds. Proc. 2011 Interbull Meet., Stavanger, Norway, 44:185-189.
- GonzalezS. DuncanS.E. O’KeefeS.F. SumnerS.S. HerbeinJ.H., 2003. Oxidation and textural characteristics of butter and ice cream with modified fatty acid profiles. J. Dairy Sci. 86:70-77.
- GroeneveldE. KovacM. MielenzN., 2010. VCE user’s guide and reference manual, ver. 6.0. Institute of Animal Science Federal Agricultural Research Center Publ., Neustadt, Germany.
- GrummerR.R., 1991. Effect of feed on the composition of milk fat. J. Dairy Sci. 74:3244-3257.
- HaugA. HostmarkA. HarstadO., 2007. Bovine milk in human nutrition: a review. Lipids Health Dis. 6:25.
- HenningD.R. BaerR.J. HassanA.N. DaveR., 2006. Major advances in concentrated and dry milk products, cheese, and milk fat-based spreads. J. Dairy Sci. 89:1179-1188.
- HillbrickG. AugustinM.A., 2002. Milk fat characteristics and functionality: opportunities for improvement. Aust. J. Dairy Technol. 57:45-51.
- ISO, 2001. Milk and milk products. Extraction methods for lipids and liposoluble compounds. ISO Norm 14156:2001 (IDF 172:2001). International Standardization Organization ed., Geneva, Switzerland.
- ISO, 2002a. Milk fat. Determination of the fatty acid composition by gas-liquid chromatography. ISO Norm 15885:2002 (IDF 184:2002). International Standardization Organization ed., Geneva, Switzerland.
- ISO, 2002b. Milk fat. Preparation of fatty acid methyl esters. ISO Norm 15884:2002 (IDF 182: 2002). International Standardization Organization ed., Geneva, Switzerland.
- ISO, 2009. Milk fat from enriched dairy products. Determination of omega-3 and omega-6 fatty acid content by gas-liquid chromatography. ISO Norm 23065:2009 (IDF 211: 2009). International Standardization Organization ed., Geneva, Switzerland.
- Lindmark ManssonH., 2008. Fatty acids in bovine milk fat. Food Nutr. Res. 52:10.3402/fnr.v52i0.1821.
- LivingstoneK.M. LovegroveJ.A. GivensD.I., 2012. The impact of substituting SFA in dairy products with MUFA or PUFA on CVD risk: evidence from human intervention studies. Nutr. Res. Rev. 25:193-206.
- MigliorF. SewalemA. JamrozikJ. BohmanovaJ. LefebvreD.M. MooreR.K., 2007. Genetic analysis of milk urea nitrogen and lactose and their relationships with other production traits in Canadian Holstein cattle. J. Dairy Sci. 90:2468-2479.
- MisztalI. TsurutaS. StrabelT. AuvrayB. DruetT. LeeD.H., 2002. BLUPF90 and related programs (BGF90). Available from: http://nce.ads.uga.edu/wiki/lib/exe/fetch.php?media=28-07.pdf
- MrodeR.A., 2005. Linear model for the prediction of animal breeding values. CABI Publ., Oxfordshire, UK.
- MuirB.L. KistemakerG. CanavesiF., 2007. Genetic parameters for a multiple-trait multiple-lactation random regression test day model in Italian Holsteins. J. Dairy Sci. 90:1564-1574.
- SamoreA.B. CanavesiF. RossoniA. BagnatoA., 2012. Genetics of casein content in Brown Swiss and Italian Holstein dairy cattle breeds. Ital. J. Anim. Sci. 11:e36.
- SAS, 2011. SAS/STAT user’s guide, version 9.3. SAS Inst. Inc., Cary, NC, USA.
- SoinioM. LaaksoM. LehtoS. HakalaP. RonnemaaT., 2003. Dietary fat predicts coronary heart disease events in subjects with type 2 diabetes. Diabetes Care 26:619-624.
- SoyeurtH. DardenneP. BertozziC. GenglerN., 2007a. Genetic parameters of butter hardness estimated by test-day model. Available from: https://journal.interbull.org/index.php/ib/article/view/984/975
- SoyeurtH. DardenneP. DeharengF. BastinC. GenglerN., 2008. Genetic parameters of saturated and monounsaturated fatty acid content and the ratio of saturated to unsaturated fatty acids in bovine milk. J. Dairy Sci. 91:3611-3626.
- SoyeurtH. DardenneP. DeharengF. LognayG. VeselkoD. MarlierM. BertozziC. MayeresP. GenglerN., 2006. Estimating fatty acid content in cow milk using mid-infrared spectrometry. J. Dairy Sci. 89:3690-3695.
- SoyeurtH. GillonA. VanderickS. MayeresP. BertozziC. GenglerN., 2007b. Estimation of heritability and genetic correlations for the major fatty acids in bovine milk. J. Dairy Sci. 90:4435-4442.
- SoyeurtH. MisztalI. GenglerN., 2010. Genetic variability of milk components based on mid-infrared spectral data. J. Dairy Sci. 93:1722-1728.
- StoopW.M. BovenhuisH. van ArendonkJ.A.M., 2007. Genetic parameters for milk urea nitrogen in relation to milk production traits. J. Dairy Sci. 90:1981-1986.
- WikingL. TheilP.K. NielsenH.J. SørensenM.T., 2010. Effect of grazing fresh legumes or feeding silage on fatty acids and enzymes involved in the synthesis of milk fat in dairy cows. J. Dairy Res. 77:337-342.
- ZhengZ.J. FolsomA.R. ShaharE. McGovernP.G. EckfeldtJ.H., 1997. Association of plasma fatty acid composition with lower extremity arterial disease. Nutr. Metab. Cardiovas. 7:360-370.