
Abstract
Current treatment modalities for pancreatic carcinoma afford only modest survival benefits. TRAIL, as a potent and specific inducer of apoptosis in cancer cells, would be a promising new treatment option. However, since not all pancreatic cancer cells respond to TRAIL, further improvements and optimizations are still needed. One strategy to improve the effectiveness of TRAIL-based therapies is to specifically target one of the 2 cell death inducing TRAIL-receptors, TRAIL-R1 or TRAIL-R2 to overcome resistance. To this end, we designed constructs expressing soluble TRAIL (sTRAIL) variants that were rendered specific for either TRAIL-R1 or TRAIL-R2 by amino acid changes in the TRAIL ectodomain. When we expressed these constructs, including wild-type sTRAIL (sTRAILwt), TRAIL-R1 (sTRAILDR4) and TRAIL-R2 (sTRAILDR5) specific variants, in 293 producer cells we found all to be readily expressed and secreted into the supernatant. These supernatants were subsequently transferred onto target cancer cells and apoptosis measured. We found that the TRAIL-R1 specific variant had higher apoptosis-inducing activity in human pancreatic carcinoma Colo357 cells as well as PancTu1 cells that were additionally sensitized by targeting of XIAP. Finally, we tested TRAIL-R1 specific recombinant TRAIL protein (rTRAILDR4) on Colo357 xenografts in nude mice and found them to be more efficacious than rTRAILwt. Our results demonstrate the benefits of synthetic biological approaches and show that TRAIL-R1 specific variants can potentially enhance the therapeutic efficacy of TRAIL-based therapies in pancreatic cancer, suggesting that they can possibly become part of individualized and tumor specific combination treatments in the future.
Abbreviations
| AML | = | Acute myeloid leukemia |
| ANOVA | = | Analysis of variance between groups |
| Bcl-xL | = | B-cell lymphoma-extra large |
| BSA | = | Bovine Serum Albumin |
| CMV | = | Cytomegalie virus |
| CuZnSOD | = | Copper-Zinc Superoxide Dismutase |
| DMEM | = | Dulbecco's modified Eagle's medium |
| DNA | = | Deoxyribonucleic acid |
| EGFP | = | Enhanced green fluorescent protein |
| ELISA | = | Enzyme-linked immunosorbent assay |
| FACS | = | Fluorescence-activated cell sorting |
| FADD | = | Fas-associated protein with death domain |
| FBS | = | Fetal bovine serum |
| FLIP | = | FLICE-like inhibitory protein |
| FIB | = | Fibrillin |
| Furin CS | = | Furin cleavage site |
| IFN-g | = | Interferon-gamma |
| ILZ | = | Isoleucine zipper |
| MSC | = | Mesenchymal stem cell |
| NF-κB | = | Nuclear factor kappa-light-chain-enhancer of activated B cells |
| OPG | = | Osteoprogerin |
| PCR | = | Polymerase chain reaction |
| PBS | = | Phosphate buffered saline |
| RANKL | = | Receptor activator of nuclear factor kappa-B ligand |
| RNAi | = | RNA interference |
| RPMI 1640 medium | = | Roswell Park Memorial Institute 1640 medium |
| SDS | = | Sodium dodecyl sulphate |
| SDS-PAGE | = | SDS-Polyacrylamide gel electrophoresis |
| SEM | = | Standard error of the mean |
| sh-sequence | = | short-hairpin sequence |
| TNF | = | Tumor necrosis factor |
| TRAIL | = | TNF-related apoptosis-inducing ligand |
| rTRAIL | = | recombinant TNF-related apoptosis-inducing ligand |
| sTRAIL | = | soluble TNF-related apoptosis-inducing ligand |
| TRAIL-R1/DR4 | = | TRAIL-receptor 1/Death – receptor 4 |
| TRAIL-R2/DR5 | = | TRAIL-receptor 2/ Death – receptor 5 |
| TRAIL-R3/DcR1 | = | TRAIL-receptor 3/Decoy-receptor 1 |
| TRAIL-R4/DcR2 | = | TRAIL-receptor 4/Decoy-receptor 2 |
| XIAP | = | X-linked Inhibitor of apoptosis protein |
Introduction
The TNF-related apoptosis-inducing ligand (TRAIL/Apo-2L) was identified in the 1990s based on its sequence homology to other members of the TNF superfamily and is a type II transmembrane protein that can rapidly induce apoptosis.Citation1-4 It has been mostly used as a recombinant and soluble protein produced in bacteria and is made up of the TRAIL ectodomain that corresponds to amino acid 114 to 281 of the TRAIL amino acid sequence. It is usually referred to as recombinant TRAIL (rTRAIL) or simply TRAIL.Citation5-7 An important and unique characteristic of TRAIL is its ability to selectively trigger receptor-mediated apoptosis in cancer cells but not in normal cells.Citation8 For this reason, TRAIL has been extensively studied as an anti-cancer reagent. TRAIL interacts with an intricate receptor system consisting of 2 apoptosis-inducing agonistic death receptors, death receptor 4 (DR4/TRAIL-R1) and death receptor 5 (DR5/TRAIL-R2), and 3 antagonistic or decoy receptors, decoy receptor 1 (DcR1/TRAIL-R3), decoy receptor 2 (DcR2/ TRAIL-R4), and the soluble receptor osteoprotegerin (OPG).Citation9 TRAIL binds as homotrimer to DR4 and DR5, which results in trimerization of the receptors and this leads to the recruitment of Fas-associated death domain (FADD), which in turn allows binding and activation of the initiator caspase, caspase-8. Mature caspase-8 molecules are capable to activate downstream effector caspases, which then execute apoptosis.Citation10-12 DcR1 lacks a death domain and DcR2 contains a truncated death domain, so the binding of TRAIL to these receptors does not induce apoptosis but could instead prevent apoptosis by sequestering available TRAIL or by interfering with TRAIL-R1 or TRAIL-R2 signaling complexes.Citation13-15 OPG is a secreted protein that functions as a decoy receptor for RANKL,Citation16 but may also act as decoy receptor for TRAIL and protect against apoptosis.Citation17 The resistance of normal, non-transformed cells to TRAIL-induced apoptosis is likely regulated at more levels than just presence of decoy receptors, but decoy receptors may be responsible for the unresponsiveness of some cancer cells.Citation18 Consequently, one potential way of improving the therapeutic efficacy of TRAIL is to generate agents that act in a TRAIL-receptor specific way, preferring one of the 2 death receptors and potentially avoiding the decoy receptors. To this end, agonistic antibodies directed against one of the 2 apoptosis-inducing TRAIL-receptors have been developed and tested in various experimental systems as well as in the clinic.Citation19-22 In addition, engineered variants of TRAIL, containing specific amino acid changes leading to specific targeting of TRAIL-R1 or TRAIL-R2 have been developed and have shown enhanced anti-tumor properties in-vitro and in-vivo.Citation23-31 This is of particular interest in the case of pancreatic cancer as previous studies have shown that pancreatic tumor cells preferentially use TRAIL-R1 to execute TRAIL-induced apoptosis.Citation32,33 While these studies were carried out with agonistic antibodies against TRAIL-R1, TRAIL variants that possess specificity for TRAIL-R1 might hold important advantages such as the fact that they are smaller than antibodies and might therefore better able to reach and infiltrate growing tumors. Moreover, such proteins can be further engineered and optimized to increase activity, specificity and stability, and are potentially easier to produce and to test. Therefore, we generated TRAIL-receptor specific TRAIL variants based on earlier published amino acid sequence changes Citation24,26,Citation28-30 and tested them on pancreatic cancer cells in-vitro and in-vivo. We utilized rTRAIL variant proteins as well as a gene expression system that we described previously and gives rise to secreted and soluble TRAIL (sTRAIL).Citation27,34,35 This system, in principle, also allows for high-throughput analyses of TRAIL variants as new mutants would not have to be expressed in bacteria and the protein purified before they can be tested on tumor cells and can also be integrated into various gene and cellular delivery vehicles. Here, we show that both rTRAIL and sTRAIL variants directed against TRAIL-R1 possess higher apoptosis inducing activity in pancreatic cancer cells as compared to TRAIL-R2 specific variants and wild-type rTRAIL and sTRAIL. Thus, our study highlights that TRAIL-R1 specific variants constitute a potential improvement to conventional TRAIL therapies and might be able to overcome apoptosis resistance particularly in pancreatic cancers.
Results
TRAIL-receptor specific sTRAIL variants are expressed and secreted
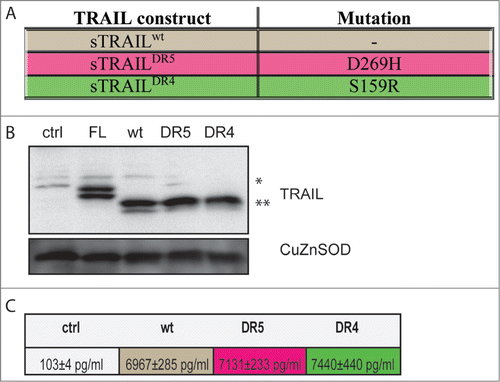
DNA fragments encoding the signal peptide of the human fibrillin-1 gene (hFIB) gene, the Furin cleavage site (Furin CS), an Isoleucine zipper (ILZ) and the ectodomain of TRAIL (aa114-aa281; sTRAIL) were ligated together (Fig. S1). The hFIB signaling peptide leads to effective secretion of sTRAIL while the Furin CS mediates the removal of the signal peptide, while the ILZ forces the formation of trimeric sTRAIL, which has higher activity than monomeric sTRAIL. In addition to the common wild-type TRAIL construct (sTRAILwt) containing the consensus TRAIL cDNA sequence, we engineered one TRAIL-R1 (DR4) specific sTRAIL variant, termed sTRAILDR4 and one specific for TRAIL-R2 (DR5), labeled sTRAILDR5 (Fig. S2). Both sTRAILDR4 and sTRAILDR5 contained only one amino acid exchange, namely S159R and D269H, respectively ().Citation27 These 2 expression cassettes together with sTRAILwt were cloned into pCDNA3.1 and along an EGFP control plasmid were transfected into 293 cells. Subsequent Western blot analyses demonstrated that the 2 sTRAIL variants were expressed () and ELISA measurements of the 293 cell supernatants revealed that sTRAIL variants were indeed secreted (). The results demonstrated that the different sTRAIL variants were expressed and secreted to approximately the same levels as sTRAILwt and that our expression system can be used as an alternative to recombinant proteins as part of gene and cellular therapeutic approaches. In addition, the system serves as an easy test system for novel TRAIL variants or other therapeutic proteins without the need for laborious and time-consuming recombinant protein production and purification.
Figure 1. Generation and expression of sTRAIL specific variants. (A) Table showing the mutations in sTRAILwt leading to sTRAILDR5 (TRAIL-R2 specific) and sTRAILDR4 (TRAIL-R1 specific) variants. The TRAIL receptor specific variants were generated by site-directed mutagenesis inserting a D269H amino acid change (sTRAILDR5) and an amino acid change (S159R) to generate sTRAILDR4 in the sTRAIL ectodomain. (B) All 3 sTRAIL expression constructs were transiently transfected into 293 cells. After 24 h whole cell lysates were analyzed by Western blotting for TRAIL revealing similar expression levels for all 3 constructs. As control, expression from a conventional construct encoding membrane-bound, full-length TRAIL (FL) without hFIB, Furin CS or ILZ segments was also analyzed. EGFP transfected cells (ctrl) served as additional control. CuZnSOD was used as a loading control (C): Results of ELISA analyses for TRAIL showing the levels of secreted sTRAILwt (brown), sTRAILDR5 (red) and sTRAILDR4 (green) into the supernatant of transfected 293 cells. Results for cells transfected with the EGFP control plasmid (ctrl; gray) are also shown.
Apoptosis induction by sTRAILDR4 in pancreatic cancer cells
Using TRAIL-receptor specific antibodies we and others have recently shown that TRAIL-induced apoptosis is preferentially mediated via TRAIL-R1 in pancreatic carcinoma cells.Citation32,33 Therefore, we set out to test our sTRAILDR4 variant in pancreatic tumor cells and to compare it to sTRAILDR5 and sTRAILwt. For this, we transfected 293 cells with the 4 constructs expressing EGFP (ctrl), sTRAILwt, sTRAILDR5 and sTRAILDR4, respectively, harvested the supernatant media after 48 h, measured the sTRAIL concentration by ELISA and applied 8 ng/ml, 2 ng/ml, 0.5 ng/ml and 0.25 ng/ml onto Colo357 pancreatic cancer cells. After 24 h exposure to the sTRAIL variants we measured apoptosis. We found that at 8 ng/ml the 3 sTRAIL types (sTRAILwt, sTRAILDR5 and sTRAILDR4) had very similar apoptosis-inducing activities (). Even though there was a trend toward increased cell death with sTRAILDR4, the difference was not significant. Added onto Colo357 cells at 2 ng/ml, sTRAILDR4 triggered relatively more apoptosis than sTRAILDR5, but still provided no significant advantage over sTRAILwt (). However, at lower concentrations of sTRAILDR4 (0.5 ng/ml and 0.25 ng/ml) it showed significantly higher levels of apoptosis as compared to sTRAILwt and sTRAILDR5 ().
Figure 2. TRAIL-R1 (A–D) Supernatants from 293 cells that were transfected with EGFP (ctrl; gray), sTRAILwt (brown), sTRAILDR5 (red), and sTRAILDR4 (green) constructs were then transferred onto Colo357 pancreatic cancer cells at 8 ng/ml (A), 2 ng/ml (B), 0.5 ng/ml (C) and 0.25 ng/ml (D). After 24 h apoptosis was measured. (E) HeLa cells were incubated with the same EGFP (ctrl; gray) sTRAILwt (brown), sTRAILDR5 (red), and sTRAILDR4 (green) 293 cell supernatants at 2 ng/ml of TRAIL for 24 h before apoptosis was measured. (F) HL-60 cells were also incubated with EGFP (ctrl; gray) sTRAILwt (brown), sTRAILDR5 (red), and sTRAILDR4 (green) 293 cell supernatants (8 ng/ml of sTRAIL). 24 h later apoptosis was measured.
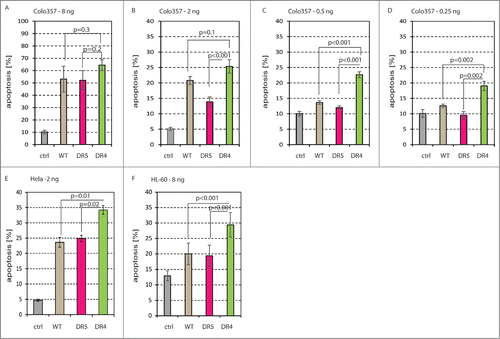
Moreover, in HeLa cervical cancer cells and HL-60 acute myeloid leukemia (AML) cells, we also found a preference to undergo apoptosis via TRAIL-R1, confirming results with purified rTRAIL variant protein ().Citation30,36
Effects of rTRAILDR4 on pancreatic cancer cells in-vitro
Next, we tested rTRAILDR4 and rTRAILwt protein on Colo357 cells. These proteins were generated and purified as described previously.Citation37 We applied the 2 proteins at a concentration of 100 ng/ml for 24 h before we measured apoptosis. The results reveal that rTRAILDR4 led to significantly more apoptosis than rTRAILwt in Colo357 cells corroborating our result with the sTRAIL variants ().
Figure 3. Apoptosis analyses of pancreatic carcinoma cells treated with rTRAILwt and rTRAILDR4.(A) Colo357 cells were treated with PBS (gray) as control, 100 ng/ml rTRAILwt (rwt; brown) and rTRAILDR4 (rDR4; green) for 24 h before apoptosis was measured. The apoptosis was significantly higher with rTRAILDR4 than rTRAILwt. (B) PancTu1.shctrl (filled) and PancTu1.shXIAP (dashed) cells were treated with PBS (gray) as control, 100 ng/ml rTRAILwt (rwt; brown) and rTRAILDR4 (rDR4; green) for 24 h before apoptosis was measured. The results show that PancTu1.shctrl cells are TRAIL resistant and that silencing of XIAP (in PancTu1.shXIAP) sensitizes the cells. In PancTu1.shXIAP rTRAILDR4 induces significantly more apoptosis than rTRAILwt. In the insert the knock-down of XIAP is shown by Western blot of protein lysates from PancTu1.shXIAP (lane 1) and PancTu1.shctrl (lane 2) cells. The CuZnSOD blot serves as loading control.
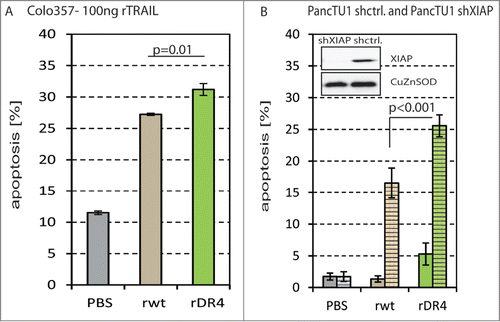
As not all pancreatic cancer cells are principally TRAIL sensitive and we and others have recently reported that TRAIL-induced apoptosis is preferentially mediated via TRAIL-R1 in pancreatic carcinoma cells and profoundly enhanced by XIAP inhibitors,Citation32,33 we tested the utility of TRAIL-R1 specific variants in conjunction with XIAP inhibition. When we treated XIAP-silenced PancTu1 (PancTu1.shXIAP) cells with rTRAILwt and rTRAILDR4, we could measure apoptosis in all cases, but rTRAILDR4 gave rise to significantly higher cell death rates than rTRAILwt (). These findings are in line with our previous findings Citation32,33 and underline that pancreatic cancer cells undergo apoptosis preferentially via TRAIL-R1 even in cases where the TRAIL treatment has to be combined with targeting of other molecules such as XIAP. In summary, TRAIL receptor specific variants offer a substantial improvement over normal TRAIL as they can better target TRAIL resistant cancer cells and act more specifically and effectively in combined treatment approaches.
The effect of rTRAILDR4 in pancreatic cancer cell-derived xenografts
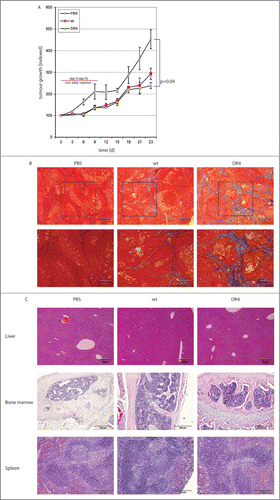
After having demonstrated the effectiveness and specificity of our TRAILDR4 specific variant we tested and compared rTRAILDR4 and rTRAILwt in Colo357 xenografts. First, we established tumors in immune-deficient nu/nu mice by subcutaneously injecting 5 × 105 Colo357 cells into the flank. After 10 d when the tumors where palpable, the animals were injected intraperiteonally with 5 mg/kg rTRAILwt and rTRAILDR4 proteins, respectively. The injections were repeated daily over a period of 10 d. During this time and 13 additional days following the conclusion of the treatment schedule the sizes of the tumors were followed. The results of this study show that both rTRAILwt and rTRAILDR4 had a significant tumor regressive effect compared to the carrier (PBS buffer). In addition, this effect was slightly larger in the rTRAILDR4 mice than in rTRAILwt treated animals (). Histological analyses of these tumors confirmed the better anti-neoplastic activity of rTRAILDR4 over rTRAILwt on Colo357 xenografts (), as we detected increased connective tissue formation in tumors treated with rTRAILDR4 by trichrome staining. These results are indicative of higher levels of apoptosis in cancer cells in the rTRAILDR4 treated xenografts, since dead tumor cells are being replaced by scar tissue. Moreover, tissue sections from liver, bone marrow and spleen from rTRAILwt and rTRAILDR4 injected animals showed no abnormalities demonstrating a lack of toxic side effects from these treatments (). Taken together, TRAIL-R1 specific variants have enhanced apoptosis-inducing effects in pancreatic cancer cells in-vitro and show a trend toward greater therapeutic efficacy in-vivo and warrant further investigation with the goal to optimize TRAIL-based therapies in the future.
Figure 4. Treatment with rTRAIL variants lead to remission of Colo357 xenografts. (A) Immune-deficient mice were injected with5 × 106 Colo357 cells. After 10 d ( = day 0 of treatment regimen) 5 mg/kg rTRAILDR4 (green triangles; n = 3) were injected intraperitoneally. Treatments were repeated daily over 10 d. Animals in 2 additional groups received the same schedule with rTRAILwt (red squares; n = 3) and PBS (gray diamonds; n = 3), respectively. The tumor growth was followed over 23 d in total and the values are depicted in the graph. (B) Trichrome staining of tumor sections from mice treated with PBS, rTRAILwt and rTRAILDR4. (C) H&E staining of liver, bone marrow and spleen sections from mice treated with PBS, rTRAILwt and rTRAILDR4, respectively.
Discussion
While TRAIL is generally a potent inducer of apoptosis in various tumor cells, several other cancer cells are resistant owing to a variety of molecular mechanisms. The resistance to TRAIL can occur at multiple steps in the TRAIL signaling pathway.Citation38 Various studies Citation39,40 found an NF-κB mediated survival mechanism involving NF-κB target genes such as XIAP, c-Flip and/or Bcl-xL.Citation41 In fact, in many cancer cells XIAP was found to be over-expressed in comparison to normal cells and several studies have confirmed the involvement of this anti-apoptotic protein in TRAIL resistance as inhibition of XIAP either chemically or by RNAi yielded an increase in cell death after TRAIL treatment in different types of cancer.Citation42-45 Similarly, Bcl-xLCitation46 and c-FlipCitation47-49 have been shown to confer TRAIL-resistance to tumor cells. However, we found that in pancreatic cancer cells XIAP had a dominant role and that Bcl-xL and c-Flip had either a modest or no function in apoptosis resistance.Citation35,50 Noteworthy, silencing of XIAP by RNAi resulted in substantial and significant TRAIL sensitization, but only additional pro-apoptotic signals such as adenoviral-mediated overexpression of TRAIL-R1 or IFN-γ-induced increases in caspase-8 levels resulted in very high cell death rates of 70–90%,Citation50 clearly indicating that both excessive anti-apoptotic factors and limiting pro-apoptotic functions contribute to apoptosis resistance. Thus, in the present study we turned to variants of TRAIL that are specific for TRAIL-R1 or TRAIL-R2 to augment the pro-apoptotic responses in cancer cells by circumventing the inhibitory functions of the 2 decoy receptors, TRAIL-R3 and TRAIL-R4. In addition, the functional unit of TRAIL-receptors is a trimer, and in this context it is thought that in principle TRAIL-R1 or TRAIL-R2 homotrimers possess higher apoptosis-inducing activities than heterodimers containing both TRAIL-R1 and TRAIL-R2 or even decoy receptor molecules.Citation27,Citation51-53 Therefore, TRAIL-receptor specific variants could possess more pro-apoptotic activity, as they would tend to assemble homotrimers. However, as cancer cells often prefer to trigger apoptosis either via TRAIL-R1 or TRAIL-R2,Citation27 it is important to determine the preference and then use TRAIL variants in an individualized manner. Pancreatic cancers have been reported to mainly utilize TRAIL-R1.Citation32,33 Therefore, we tested a TRAIL-R1 specific variant on 2 different pancreatic cancer cell lines along with a TRAIL-R2 specific variant and wild-type TRAIL. We found that the TRAIL-R1 variant had superior apoptosis-inducing activities when compared to the other forms of TRAIL. In these studies we used expression constructs that contained the sequence for the ectodomain of TRAIL including specific single point mutations that serve to change the specificity toward one of the 2 TRAIL death receptors. These TRAIL ectodomain encoding cDNA segments were ligated to an ILZ stretch that enforces the trimerization of TRAIL molecules turning it into a more active protein. The secretion of this fusion protein consisting of sTRAIL and the ILZ domain is facilitated by an exogenous signal peptide and a cleavage site for the ubiquitous protease Furin.Citation34 Such expression cassettes can be integrated into viral vector systems for cancer gene therapeutic approaches Citation54 or they can be used in connection with cellular delivery vehicles such as mesenchymal stem cells (MSCs) that can infiltrate tumor tissues where they produce and secrete sTRAIL, which in turn acts on the neighboring cancer cells.Citation35,55,56 In the present study we initially transfected our TRAIL constructs into 293 cells that produced and secreted the different sTRAIL variants. After harvesting of the sTRAIL containing supernatants, measuring and normalizing of TRAIL concentrations, the supernatants were transferred onto cancer cells and apoptosis measured. We could show that these expressed and secreted variants behaved similarly to the equivalent recombinant proteins and can therefore be used inside of gene and cell therapeutic vectors providing the same specificity for TRAIL-R1 and TRAIL-R2, respectively.
In addition, this system allows for faster and more effective testing of potential further sTRAIL variants sparing the expensive and time-consuming need for purification of recombinant proteins from bacterial lysates. Numerous new TRAIL variants could be generated by PCR-based random mutagenesis,Citation57 cloned into our expression plasmid backbone and directly tested for their activity and specificity on cancer cells after transfection into 293 cells. Thus, our system could greatly enhance the search for optimized TRAIL variants and possibly other therapeutic proteins.
Having demonstrated the general utility of our TRAIL variants and in particular the superior apoptosis-inducing activity of the TRAIL-R1 variant on Colo357 pancreatic cancer cells, we turned to an in-vivo model. To this end, we examined the effects of recombinant wild-type and TRAIL-R1 specific TRAIL protein in Colo357 xenografts. The results showed that the rTRAILDR4 protein displayed significantly better anti-tumor efficacy when compared to the control cohort (PBS). Even though, the difference between rTRAILDR4 and rTRAILwt failed to reach significance in this study, we observed an increase in scar tissue formation in tumors treated with rTRAILDR4 as compared to wild-type TRAIL, which shows that more tumor tissue is replaced by connective tissue indicative of higher cancer cell killing. Taken together, these results suggest a higher effectivity of rTRAILDR4 and further highlight the potential utility of TRAIL-R1 specific variants in pancreatic cancer.
However, the overall efficacy of the rTRAILDR4 protein still leaves room for improvements such as integration of an ILZ domain or additional changes to the protein sequence to make the protein more stable and/or more active in-vivo. Our expression system as discussed above is well suited to identify such novel variants. Moreover, cellular delivery vehicles such as MSCs could be used to enhance availability of TRAIL in tumor tissues and TRAIL could be combined with conventional chemotherapeutic drugs for increased anti-tumor effects.Citation3,35,55,58
In summary, our results demonstrate that TRAIL-receptor specific TRAIL variants are a significant improvement and represent a significant step forward to increase the potential for clinical tests and use of MSC-based delivery of TRAIL as part of a tumor-specific and patient-tailored therapy. In particular aggressive pancreatic cancer that is difficult to treat successfully is a possible target for such novel treatment approaches.
Materials and Methods
Reagents and cell culture
All chemicals, unless otherwise stated, were purchased from Sigma (St. Louis). Antibodies used in this study were sheep anti-CuZnSOD (The Binding Site) and rabbit anti-TRAIL (Peprotech) mouse anti-XIAP (BD Bioscience). Peroxidase-conjugated secondary antibodies were anti-mouse, anti-rabbit and anti-sheep (Santa Cruz).
Human cervix carcinoma HeLa cells (ATCC), human pancreatic cancer PancTu1 cells (gift from Simone Fulda) as well as human embryonic kidney 293 cells (Life Technologies) were grown in Dulbecco's modified eagle medium (DMEM) supplemented with 10% FBS, 100 U/ml penicillin and 100 μg/ml streptomycin. HL-60 and Colo357 (gifts from Eva Szegezdi) were grown in RPMI-1640 medium supplemented with 10% FBS, 100 U/ml penicillin and 100 μg/ml streptomycin.
Generation of TRAIL constructs
Generation of sTRAIL constructs and site-directed mutagenesis have been previously described.Citation27 Further information and details can be found in the Supplementary Material and Methods section.
Apoptosis assay
Apoptosis was measured according to Nicoletti et al.Citation59 and has been described previously.Citation60
Transfection of 293 cells
For the transfection of 293 cells (5 × 105 cells/well) we used the calcium phosphate transfection method which has been previously described.Citation27 Supernatants of transfected 293 cells were cleared by centrifugation and diluted to determine the TRAIL concentration by ELISA.
Animal studies
Ten week old female CD1 nu/nu mice (Charles River, Wilmington, MA) were injected with 5 × 106 Colo357 cells in 200 μl PBS. After 10 d and when tumors were palpable the animals were intraperitoneally daily injected with purified rTRAIL (wt and DR4) at a concentration of 5 mg/kg over a period of 10 d. The growth of the tumors was followed over 23 d. The tumor volume was calculated using 3 different diameters and the formula: π/6*(d1*d2*d3). The animal studies were performed according to national laws and covered by license.
Histological analyses
Tumor, liver, bone marrow and spleen samples taken during necropsy were fixed in 10% neutral-buffered formalin, paraffin embedded and 4-μm sections were stained with hematoxylin and eosin (H&E). Bone marrow samples were decalcified before processing. Additionally Masson's Trichrome staining was carried out on tumor sections as follows. Slides were rinsed and placed in Bouin's solution overnight at room temperature. The next day, slides were washed, stained with Weigert Hematoxylin for 10 min, rinsed in distilled water and stained with Biebrich scarlet-acid fuchsin for 5 min followed with immersion in phosphomolybdic/phosphotungstic acid solution for 15 min and stained with aniline blue for 5 min. Slides were then rinsed, dehydrated with 95% and absolute alcohol and mounted.
Western blot
Western blots were carried out as described previously.Citation61 Briefly, cells were harvested and lyzed in cell lysis buffer. Proteins were separated by SDS-PAGE and transferred onto PVDF membranes (GE Healthcare Biosciences). Primary and secondary antibodies were diluted in TBS, 0.1% Tween and 3% BSA. Bands were visualized with ECL Western blotting Substrate (Pierce).
TRAIL enzyme-linked immunosorbent assay (ELISA)
To examine secreted sTRAIL, the culture supernatants of transfected 293 cells were cleared by centrifugation at 2,000 g for 20 min. The supernatants were diluted 20-fold and a commercial TRAIL ELISA kit (R&D Systems) was used following the manufacturer's instructions.
RNAi knock-down constructs and stable cell line generation
The following small hairpin (sh) RNA motif was used to target: XIAP (5′-GTGGTAGTCCTGTTTCAGC-3′) and has been previously described.Citation39 Sense and antisense oligos containing the sh-sequence and 5′ overhang representing a restricted BbsI site and EcoRI site on the 3′ side were hybridized to generate double-stranded DNA fragments. These fragments were then cloned into a modified pU6.ENTR plasmid (Life technologies). The resulting pU6.ENTR plasmid was used to generate the pBlockIt.shXIAP plasmid using the pBlockIt vector (Life Technologies) and LR Clonase II. This was used to generate stable XIAP knockdown clones of PancTu1 cells. For this, pBlockit.shXIAP plasmids were FuGene-transfected (Roche) into PancTu1 cells. Three days later, the transfected cells were split into Blasticidin containing selection medium. Arising clones were picked, transferred to 24 well-plates and tested for XIAP silencing, respectively. Clones that did not show a knock-down were used as controls labeled PancTu1.shctrl. These control clones were tested and shown to behave like parental PancTu1 cells.
Statistical analysis
If not stated, 3 independent experiments were performed. Experimental values are expressed as mean value ± standard error (SEM). For significance analyses, analysis of variance (ANOVA) between groups was used and P < 0.05 was considered significant and P < 0.01 as highly significant.
Disclosure of Potential Conflicts of Interest
WJQ and RHC are co-founders of Triskel Therapeutics, holding patents on receptor-specific rTRAIL variants. The other authors declare no potential conflict of interest.
972183_Supplementary_Materials.zip
Download Zip (15.8 KB)Acknowledgments
We thank the members of the TRIDENT consortium for helpful discussions and input and are grateful to Jérôme Courtête for his assistance during the initial stages of the project.
Funding
The work was supported by an EU-FP6-STREP (TRIDENT) award. The work was also supported by an EU-FP6 Marie-Curie Excellence Team Award (MIST) and by an EU-RTN Award (ApopTrain) (to RMZ).
Supplemental Material
Supplemental data for this article can be accessed on the publisher's website.
References
- Wiley SR, Schooley K, Smolak PJ, Din WS, Huang CP, Nicholl JK, Sutherland GR, Smith TD, Rauch C, Smith CA, et al. Identification and characterization of a new member of the TNF family that induces apoptosis. Immunity 1995; 3:673-82; PMID:8777713; http://dx.doi.org/10.1016/1074-7613(95)90057-8
- Mohr A, Henderson G, Dudus L, Herr I, Kuerschner T, Debatin KM, Weiher H, Fisher KJ, Zwacka RM. AAV-encoded expression of TRAIL in experimental human colorectal cancer leads to tumor regression. Gene Ther 2004; 11:534-43; PMID:14999225; http://dx.doi.org/10.1038/sj.gt.3302154
- Mohr A, Lyons M, Deedigan L, Harte T, Shaw G, Howard L, Barry F, O'Brien T, Zwacka R. Mesenchymal stem cells expressing TRAIL lead to tumour growth inhibition in an experimental lung cancer model. J Cell Mol Med 2008; 12:2628-43; PMID:18373740; http://dx.doi.org/10.1111/j.1582-4934.2008.00317.x
- Testa U. TRAILTRAIL-R in hematologic malignancies. J Cell Biochem 2010; 110:21-34; PMID:20336667
- Pitti RM, Marsters SA, Ruppert S, Donahue CJ, Moore A, Ashkenazi A. Induction of apoptosis by Apo-2 ligand, a new member of the tumor necrosis factor cytokine family. J Biol Chem 1996; 271:12687-90; PMID:8663110; http://dx.doi.org/10.1074/jbc.271.22.12687
- Cha SS, Kim MS, Choi YH, Sung BJ, Shin NK, Shin HC, Sung YC, Oh BH. 2.8 A resolution crystal structure of human TRAIL, a cytokine with selective antitumor activity. Immunity 1999; 11:253-61; PMID:10485660; http://dx.doi.org/10.1016/S1074-7613(00)80100-4
- Hymowitz SG, O’Connell MP, Ultsch MH, Hurst A, Totpal K, Ashkenazi A, de Vos AM, Kelley RF. A unique zinc-binding site revealed by a high-resolution X-ray structure of homotrimeric Apo2LTRAIL. Biochem (Mosc) 2000; 39:633-40; PMID:10651627; http://dx.doi.org/10.1021/bi992242l
- Ashkenazi A, Pai RC, Fong S, Leung S, Lawrence DA, Marsters SA, Blackie C, Chang L, McMurtrey AE, Hebert A, et al. Safety and antitumor activity of recombinant soluble Apo2 ligand. J Clin Invest 1999; 104:155-62; PMID:10411544; http://dx.doi.org/10.1172/JCI6926
- LeBlanc HN, Ashkenazi A. Apo2LTRAIL and its death and decoy receptors. Cell Death Differ 2003; 10:66-75; PMID:12655296; http://dx.doi.org/10.1038/sj.cdd.4401187
- Bodmer JL, Holler N, Reynard S, Vinciguerra P, Schneider P, Juo P, Blenis J, Tschopp J. TRAIL receptor-2 signals apoptosis through FADD and caspase-8. Nat Cell Biol 2000; 2:241-3; PMID:10783243; http://dx.doi.org/10.1038/35008667
- Kischkel FC, Lawrence DA, Chuntharapai A, Schow P, Kim KJ, Ashkenazi A. Apo2LTRAIL-dependent recruitment of endogenous FADD and caspase-8 to death receptors 4 and 5. Immunity 2000; 12:611-20; PMID:10894161; http://dx.doi.org/10.1016/S1074-7613(00)80212-5
- Sprick MR, Weigand MA, Rieser E, Rauch CT, Juo P, Blenis J, Krammer PH, Walczak H. FADDMORT1 and caspase-8 are recruited to TRAIL receptors 1 and 2 and are essential for apoptosis mediated by TRAIL receptor 2. Immunity 2000; 12:599-609; PMID:10894160; http://dx.doi.org/10.1016/S1074-7613(00)80211-3
- Sheridan JP, Marsters SA, Pitti RM, Gurney A, Skubatch M, Baldwin D, Ramakrishnan L, Gray CL, Baker K, Wood WI, et al. Control of TRAIL-induced apoptosis by a family of signaling and decoy receptors. Science 1997; 277:818-21; PMID:9242611; http://dx.doi.org/10.1126/science.277.5327.818
- Kimberley FC, Screaton GR. Following a TRAIL: update on a ligand and its five receptors. Cell Res 2004; 14:359-72; PMID:15538968; http://dx.doi.org/10.1038/sj.cr.7290236
- Merino D, Lalaoui N, Morizot A, Schneider P, Solary E, Micheau O. Differential inhibition of TRAIL-mediated DR5-DISC formation by decoy receptors 1 and 2. Mol Cell Biol 2006; 26:7046-55; PMID:16980609; http://dx.doi.org/10.1128/MCB.00520-06
- Leibbrandt A, Penninger JM. RANKLRANK as key factors for osteoclast development and bone loss in arthropathies. Adv Exp Med Biol 2009; 649:100-13; PMID:19731623; http://dx.doi.org/10.1007/978-1-4419-0298-6_7
- Emery JG, McDonnell P, Burke MB, Deen KC, Lyn S, Silverman C, Dul E, Appelbaum ER, Eichman C, DiPrinzio R, et al. Osteoprotegerin is a receptor for the cytotoxic ligand TRAIL. J Biol Chem 1998; 273:14363-7; PMID:9603945; http://dx.doi.org/10.1074/jbc.273.23.14363
- Vindrieux D, Reveiller M, Chantepie J, Yakoub S, Deschildre C, Ruffion A, Devonec M, Benahmed M, Grataroli R. Down-regulation of DcR2 sensitizes androgen-dependent prostate cancer LNCaP cells to TRAIL-induced apoptosis. Cancer Cell Int 2011; 11:42; PMID:22136382; http://dx.doi.org/10.1186/1475-2867-11-42
- Camidge DR, Herbst RS, Gordon MS, Eckhardt SG, Kurzrock R, Durbin B, Ing J, Tohnya TM, Sager J, Ashkenazi A, et al. A phase I safety and pharmacokinetic study of the death receptor 5 agonistic antibody PRO95780 in patients with advanced malignancies. Clin Cancer Res 2010; 16:1256-63; PMID:20145186; http://dx.doi.org/10.1158/1078-0432.CCR-09-1267
- Chuntharapai A, Dodge K, Grimmer K, Schroeder K, Marsters SA, Koeppen H, Ashkenazi A, Kim KJ. Isotype-dependent inhibition of tumor growth in vivo by monoclonal antibodies to death receptor 4. J Immunol 2001; 166:4891-8; PMID:11290766; http://dx.doi.org/10.4049/jimmunol.166.8.4891
- Ichikawa K, Liu W, Zhao L, Wang Z, Liu D, Ohtsuka T, Zhang H, Mountz JD, Koopman WJ, Kimberly RP, et al. Tumoricidal activity of a novel anti-human DR5 monoclonal antibody without hepatocyte cytotoxicity. Nat Med 2001; 7:954-60; PMID:11479629; http://dx.doi.org/10.1038/91000
- Trarbach T, Moehler M, Heinemann V, Kohne CH, Przyborek M, Schulz C, Sneller V, Gallant G, Kanzler S. Phase II trial of mapatumumab, a fully human agonistic monoclonal antibody that targets and activates the tumour necrosis factor apoptosis-inducing ligand receptor-1 (TRAIL-R1), in patients with refractory colorectal cancer. Br J Cancer 2010; 102:506-12; PMID:20068564; http://dx.doi.org/10.1038/sj.bjc.6605507
- MacFarlane M, Kohlhaas SL, Sutcliffe MJ, Dyer MJ, Cohen GM. TRAIL receptor-selective mutants signal to apoptosis via TRAIL-R1 in primary lymphoid malignancies. Cancer Res 2005; 65:11265-70; PMID:16357130; http://dx.doi.org/10.1158/0008-5472.CAN-05-2801
- van der Sloot AM, Tur V, Szegezdi E, Mullally MM, Cool RH, Samali A, Serrano L, Quax WJ. Designed tumor necrosis factor-related apoptosis-inducing ligand variants initiating apoptosis exclusively via the DR5 receptor. Proc Natl Acad Sci U S A 2006; 103:8634-9; PMID:16731632; http://dx.doi.org/10.1073/pnas.0510187103
- Duiker EW, de Vries EG, Mahalingam D, Meersma GJ, Boersma-van Ek W, Hollema H, Lub-de Hooge MN, van Dam GM, Cool RH, Quax WJ, et al. Enhanced antitumor efficacy of a DR5-specific TRAIL variant over recombinant human TRAIL in a bioluminescent ovarian cancer xenograft model. Clin Cancer Res 2009; 15:2048-57; PMID:19276284; http://dx.doi.org/10.1158/1078-0432.CCR-08-1535
- Reis CR, van der Sloot AM, Natoni A, Szegezdi E, Setroikromo R, Meijer M, Sjollema K, Stricher F, Cool RH, Samali A, et al. Rapid and efficient cancer cell killing mediated by high-affinity death receptor homotrimerizing TRAIL variants. Cell Death Dis 2010; 1:e83; PMID:21368856; http://dx.doi.org/10.1038/cddis.2010.61
- Yu R, Deedigan L, Albarenque SM, Mohr A, Zwacka RM. Delivery of sTRAIL variants by MSCs in combination with cytotoxic drug treatment leads to p53-independent enhanced antitumor effects. Cell Death Dis 2013; 4:e503; PMID:23429289; http://dx.doi.org/10.1038/cddis.2013.19
- Tur V, van der Sloot AM, Reis CR, Szegezdi E, Cool RH, Samali A, Serrano L, Quax WJ. DR4-selective tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) variants obtained by structure-based design. J Biol Chem 2008; 283:20560-8; PMID:18474604; http://dx.doi.org/10.1074/jbc.M800457200
- Reis CR, van der Sloot AM, Szegezdi E, Natoni A, Tur V, Cool RH, Samali A, Serrano L, Quax WJ. Enhancement of antitumor properties of rhTRAIL by affinity increase toward its death receptors. Biochem (Mosc) 2009; 48:2180-91; PMID:19236007; http://dx.doi.org/10.1021/bi801927x
- Szegezdi E, Reis CR, van der Sloot AM, Natoni A, O’Reilly A, Reeve J, Cool RH, O'Dwyer M, Knapper S, Serrano L, et al. Targeting AML through DR4 with a novel variant of rhTRAIL. J Cell Mol Med 2011; 15:2216-31; PMID:21070598; http://dx.doi.org/10.1111/j.1582-4934.2010.01211.x
- Szegezdi E, van der Sloot AM, Mahalingam D, O’Leary L, Cool RH, Munoz IG, Montoya G, Quax WJ, de Jong S, Samali A, et al. Kinetics in signal transduction pathways involving promiscuous oligomerizing receptors can be determined by receptor specificity: apoptosis induction by TRAIL. Mol Cell Proteomics 2012; 11:M111 013730; PMID:22213832; http://dx.doi.org/10.1074/mcp.M111.013730
- Lemke J, Noack A, Adam D, Tchikov V, Bertsch U, Roder C, Schütze S, Wajant H, Kalthoff H, Trauzold A. TRAIL signaling is mediated by DR4 in pancreatic tumor cells despite the expression of functional DR5. J Mol Med 2010; 88:729-40; PMID:20354842; http://dx.doi.org/10.1007/s00109-010-0619-0
- Stadel D, Mohr A, Ref C, MacFarlane M, Zhou S, Humphreys R, Bachem M, Cohen G, Möller P, Zwacka RM, et al. TRAIL-induced apoptosis is preferentially mediated via TRAIL receptor 1 in pancreatic carcinoma cells and profoundly enhanced by XIAP inhibitors. Clin Cancer Res 2010; 16:5734-49; PMID:20940278; http://dx.doi.org/10.1158/1078-0432.CCR-10-0985
- Kim MH, Billiar TR, Seol DW. The secretable form of trimeric TRAIL, a potent inducer of apoptosis. Biochem Biophys Res Commun 2004; 321:930-5; PMID:15358116; http://dx.doi.org/10.1016/j.bbrc.2004.07.046
- Mohr A, Albarenque SM, Deedigan L, Yu R, Reidy M, Fulda S, Zwacka RM. Targeting of XIAP combined with systemic mesenchymal stem cell-mediated delivery of sTRAIL ligand inhibits metastatic growth of pancreatic carcinoma cells. Stem Cells 2010; 28:2109-20; PMID:20882532; http://dx.doi.org/10.1002/stem.533
- Wajant H, Moosmayer D, Wuest T, Bartke T, Gerlach E, Schonherr U, Peters N, Scheurich P, Pfizenmaier K. Differential activation of TRAIL-R1 and -2 by soluble and membrane TRAIL allows selective surface antigen-directed activation of TRAIL-R2 by a soluble TRAIL derivative. Oncogene 2001; 20:4101-6; PMID:11494138; http://dx.doi.org/10.1038/sj.onc.1204558
- Reis CR, van Assen AH, Quax WJ, Cool RH. Unraveling the binding mechanism of trivalent tumor necrosis factor ligands and their receptors. Mol Cell Proteomics 2011; 10:M110 002808; PMID:20852190; http://dx.doi.org/10.1074/mcp.M110.002808
- Fulda S, Debatin KM. Modulation of TRAIL signaling for cancer therapy. Vitam Horm 2004; 67:275-90; PMID:15110182; http://dx.doi.org/10.1016/S0083-6729(04)67015-4
- Braeuer SJ, Buneker C, Mohr A, Zwacka RM. Constitutively activated nuclear factor-kappaB, but not induced NF-kappaB, leads to TRAIL resistance by up-regulation of X-linked inhibitor of apoptosis protein in human cancer cells. Mol Cancer Res 2006; 4:715-28; PMID:17050666; http://dx.doi.org/10.1158/1541-7786.MCR-05-0231
- Khanbolooki S, Nawrocki ST, Arumugam T, Andtbacka R, Pino MS, Kurzrock R, Logsdon CD, Abbruzzese JL, McConkey DJ. Nuclear factor-kappaB maintains TRAIL resistance in human pancreatic cancer cells. Mol Cancer Ther 2006; 5:2251-60; PMID:16985059; http://dx.doi.org/10.1158/1535-7163.MCT-06-0075
- Zhang L, Fang B. Mechanisms of resistance to TRAIL-induced apoptosis in cancer. Cancer Gene Ther 2005; 12:228-37; PMID:15550937; http://dx.doi.org/10.1038/sj.cgt.7700792
- Chawla-Sarkar M, Bae SI, Reu FJ, Jacobs BS, Lindner DJ, Borden EC. Downregulation of Bcl-2, FLIP or IAPs (XIAP and survivin) by siRNAs sensitizes resistant melanoma cells to Apo2LTRAIL-induced apoptosis. Cell Death Differ 2004; 11:915-23; PMID:15118763; http://dx.doi.org/10.1038/sj.cdd.4401416
- Vogler M, Durr K, Jovanovic M, Debatin KM, Fulda S. Regulation of TRAIL-induced apoptosis by XIAP in pancreatic carcinoma cells. Oncogene 2007; 26:248-57; PMID:16832350; http://dx.doi.org/10.1038/sj.onc.1209776
- Vogler M, Walczak H, Stadel D, Haas TL, Genze F, Jovanovic M, Bhanot U, Hasel C, Möller P, Gschwend JE, et al. Small molecule XIAP inhibitors enhance TRAIL-induced apoptosis and antitumor activity in preclinical models of pancreatic carcinoma. Cancer Res 2009; 69:2425-34; PMID:19258513; http://dx.doi.org/10.1158/0008-5472.CAN-08-2436
- Vogler M, Walczak H, Stadel D, Haas TL, Genze F, Jovanovic M, Gschwend JE, Simmet T, Debatin KM, Fulda S. Targeting XIAP bypasses Bcl-2-mediated resistance to TRAIL and cooperates with TRAIL to suppress pancreatic cancer growth in vitro and in vivo. Cancer Res 2008; 68:7956-65; PMID:18829553; http://dx.doi.org/10.1158/0008-5472.CAN-08-1296
- Hinz S, Trauzold A, Boenicke L, Sandberg C, Beckmann S, Bayer E, Walczak H, Kalthoff H, Ungefroren H. Bcl-XL protects pancreatic adenocarcinoma cells against CD95- and TRAIL-receptor-mediated apoptosis. Oncogene 2000; 19:5477-86; PMID:11114725; http://dx.doi.org/10.1038/sj.onc.1203936
- Leverkus M, Neumann M, Mengling T, Rauch CT, Brocker EB, Krammer PH, Walczak H. Regulation of tumor necrosis factor-related apoptosis-inducing ligand sensitivity in primary and transformed human keratinocytes. Cancer Res 2000; 60:553-9; PMID:10676636
- Siegmund D, Hadwiger P, Pfizenmaier K, Vornlocher HP, Wajant H. Selective inhibition of FLICE-like inhibitory protein expression with small interfering RNA oligonucleotides is sufficient to sensitize tumor cells for TRAIL-induced apoptosis. Mol Med 2002; 8:725-32; PMID:12520089
- Ganten TM, Haas TL, Sykora J, Stahl H, Sprick MR, Fas SC, Krueger A, Weigand MA, Grosse-Wilde A, Stremmel W, et al. Enhanced caspase-8 recruitment to and activation at the DISC is critical for sensitisation of human hepatocellular carcinoma cells to TRAIL-induced apoptosis by chemotherapeutic drugs. Cell Death Differ 2004; 11 Suppl 1:S86-96; PMID:15105837; http://dx.doi.org/10.1038/sj.cdd.4401437
- Buneker CK, Yu R, Deedigan L, Mohr A, Zwacka RM. IFN-gamma combined with targeting of XIAP leads to increased apoptosis-sensitisation of TRAIL resistant pancreatic carcinoma cells. Cancer Lett 2012; 316:168-77; PMID:22104728; http://dx.doi.org/10.1016/j.canlet.2011.10.035
- Leverkus M, Sprick MR, Wachter T, Denk A, Brocker EB, Walczak H, Neumann M. TRAIL-induced apoptosis and gene induction in HaCaT keratinocytes: differential contribution of TRAIL receptors 1 and 2. J Invest Dermatol 2003; 121:149-55; PMID:12839575; http://dx.doi.org/10.1046/j.1523-1747.2003.12332.x
- Kelley RF, Totpal K, Lindstrom SH, Mathieu M, Billeci K, DeForge L, Pai R, Hymowitz SG, Ashkenazi A. Receptor-selective mutants of apoptosis-inducing ligand 2tumor necrosis factor-related apoptosis-inducing ligand reveal a greater contribution of death receptor (DR) 5 than DR4 to apoptosis signaling. J Biol Chem 2005; 280:2205-12; PMID:15520016; http://dx.doi.org/10.1074/jbc.M410660200
- van Geelen CM, Pennarun B, Le PT, de Vries EG, de Jong S. Modulation of TRAIL resistance in colon carcinoma cells: different contributions of DR4 and DR5. BMC Cancer 2011; 11:39; PMID:21272366; http://dx.doi.org/10.1186/1471-2407-11-39
- Kim CY, Jeong M, Mushiake H, Kim BM, Kim WB, Ko JP, Kim MH, Kim M, Kim TH, Robbins PD, et al. Cancer gene therapy using a novel secretable trimeric TRAIL. Gene Ther 2006; 13:330-8; PMID:16195699; http://dx.doi.org/10.1038/sj.gt.3302658
- Kim SM, Lim JY, Park SI, Jeong CH, Oh JH, Jeong M, Oh W, Park SH, Sung YC, Jeun SS. Gene therapy using TRAIL-secreting human umbilical cord blood-derived mesenchymal stem cells against intracranial glioma. Cancer Res 2008; 68:9614-23; PMID:19047138; http://dx.doi.org/10.1158/0008-5472.CAN-08-0451
- Menon LG, Kelly K, Yang HW, Kim SK, Black PM, Carroll RS. Human bone marrow-derived mesenchymal stromal cells expressing S-TRAIL as a cellular delivery vehicle for human glioma therapy. Stem Cells 2009; 27:2320-30; PMID:19544410; http://dx.doi.org/10.1002/stem.136
- McCullum EO, Williams BA, Zhang J, Chaput JC. Random mutagenesis by error-prone PCR. Methods Mol Biol 2010; 634:103-9; PMID:20676978; http://dx.doi.org/10.1007/978-1-60761-652-8_7
- Loebinger MR, Eddaoudi A, Davies D, Janes SM. Mesenchymal stem cell delivery of TRAIL can eliminate metastatic cancer. Cancer Res 2009; 69:4134-42; PMID:19435900; http://dx.doi.org/10.1158/0008-5472.CAN-08-4698
- Nicoletti I, Migliorati G, Pagliacci MC, Grignani F, Riccardi C. A rapid and simple method for measuring thymocyte apoptosis by propidium iodide staining and flow cytometry. J Immunol Methods 1991; 139:271-9; PMID:1710634; http://dx.doi.org/10.1016/0022-1759(91)90198-O
- Mohr A, Buneker C, Gough RP, Zwacka RM. MnSOD protects colorectal cancer cells from TRAIL-induced apoptosis by inhibition of SmacDIABLO release. Oncogene 2008; 27:763-74; PMID:17653087; http://dx.doi.org/10.1038/sj.onc.1210673
- Zwacka RM, Stark L, Dunlop MG. NF-kappaB kinetics predetermine TNF-alpha sensitivity of colorectal cancer cells. J Gene Med 2000; 2:334-43; PMID:11045427; http://dx.doi.org/10.1002/1521-2254(200009/10)2:5%3c334::AID-JGM129%3e3.0.CO;2-Q