
Abstract
Whereas no single animal model can reproduce the complexity of periodontitis, different aspects of the disease can be addressed by distinct models. Despite their limitations, animal models are essential for testing the biological significance of in vitro findings and for establishing cause-and-effect relationships relevant to clinical observations, which are typically correlative. We provide evidence that animal-based studies have generated a durable framework for dissecting the mechanistic basis of periodontitis. These studies have solidified the etiologic role of bacteria in initiating the inflammatory response that leads to periodontal bone loss and have identified key mediators (IL-1, TNF, prostaglandins, complement, RANKL) that induce inflammatory breakdown. Moreover, animal studies suggest that dysbiosis, rather than individual bacterial species, are important in initiating periodontal bone loss and have introduced the concept that organisms previously considered commensals can play important roles as accessory pathogens or pathobionts. These studies have also provided insight as to how systemic conditions, such as diabetes or leukocyte adhesion deficiency, contribute to tissue destruction. In addition, animal studies have identified and been useful in testing therapeutic targets.
Introduction
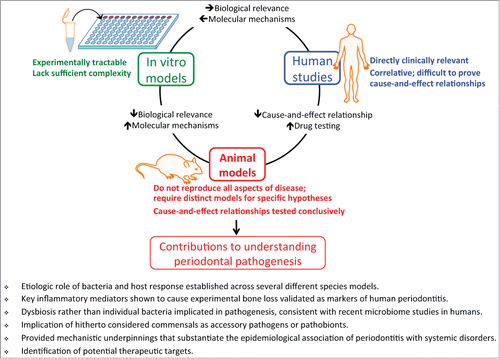
Animal models for the study of human disease have limitations that are inherent in the very definition of the term “model,” i.e., an approximation or simulation of a real system that is under investigation. It is thus obvious that no one single model reproduces all aspects of a human disease. However, the strengths of animal models more than compensate for the simulation. First, cause-and-effect relationships can be tested conclusively in animal models but are difficult to prove in human studies. Moreover, results from animal studies provide initial information on the safety and potential efficacy of novel therapeutic compounds. Furthermore, animal models have less serious limitations than in vitro models, which cannot replicate the complexity of cross-interactions that occur between the immune response, the microbiome, and the host tissue. The appropriateness of a given animal model lies in its capacity to test a specific hypothesis rather than its fidelity to all aspects of disease pathogenesis. Therefore, different models of the same disease can be used to test discrete aspects of its pathogenesis.Citation1 In essence, animal models represent a point on a spectrum of assay systems that span the more experimentally tractable in vitro models, through the biological complexity of animals, to clinically valid human studies ().
Figure 1. Animal models of periodontitis: characteristics and contributions. Animal models are contrasted with in vitro models and human studies in terms of their advantages and disadvantages, followed by a summary of key animal model-based contributions to understanding periodontal disease pathogenesis. It should be noted, however, that animal model-based research benefits from both in vitro models and human studies for obtaining mechanistic insights in finer molecular detail and for determining clinical relevance, respectively. The cycle connecting the 3 experimental systems is meant to demonstrate this interrelationship. For instance, the arrows emanating from “Animal models” and “Human studies” to “In vitro models” indicate the reliance of the former systems on the more tractable in vitro system for dissecting plausible molecular mechanisms. Conversely, the reverse arrows indicate that in vitro model-based mechanisms depend on animal and human systems for testing potential biological relevance. One of the greatest contributions of animal models is the testing of cause-and-effect relationships that cannot be typically addressed in human studies, most of which are correlative. Conversely, candidate drugs identified in animal models require the ultimate test in human clinical trials before they can be validated and enter the clinic.
The most common periodontitis models involve procedures for oral gavage and placement of ligatures. The reader is referred to previous publications for a detailed description of these models and their successful application in a large number of studies.Citation1-3 Briefly, in the oral gavage model, gingival inflammation and bone loss can be induced following oral inoculation with bacteria associated with human periodontitis. In the ligature-induced periodontitis model, the placement of silk ligatures around posterior teeth facilitates local accumulation of indigenous bacteria and enhances bacteria-mediated gingival inflammation and bone loss. Although irrelevant for studying bone loss, the so-called “chamber” and “abscess” models have been used to study specific virulence aspects of periodontal organisms in vivo. In the chamber model, bacteria are injected into the lumen of a subcutaneously implanted titanium-coil chamber and in vivo bacterial interactions with recruited inflammatory cells can be assessed accurately and quantitatively.Citation1,4,5 In the abscess model, bacteria are injected subcutaneously into the dorsum and then scored for impact on systemic health or localized abscess characteristics.Citation6,7
It is now well established that periodontitis is triggered by pathogenic microbial communities forming on subgingival tooth surfaces while the host response is responsible for the tissue damage in periodontitis; moreover, systemic conditions have an impact on periodontal disease by affecting pathologic mechanisms and host immune status.Citation8,9 Animal studies have greatly contributed to these critical principles which have been reproduced across several different animal species models demonstrating a consistency that lends support for the validity of the overall concept, as well as the utility of animal models to study periodontal disease processes. The latter reflects the enduring usefulness of in vivo studies.
We will review studies that have provided a durable framework for understanding periodontal disease, as well as the most common and important issues that have been raised over the years with regard to the relevance of animal models, in particular mice which are widely used in mechanistic studies. Moreover, we will discuss whether these models maintain the potential to generate further knowledge for an in-depth understanding of periodontal disease pathogenesis.
The Role of Bacteria
The concept that bacteria are an important etiologic factor in periodontal tissue destruction first came from studies of gnotobiotic rats and gnotobiotic mice.Citation10,11 The role of bacteria was further demonstrated by topical application of antiseptics or systemic application of antibiotics reducing bone loss in animal models involving dogs, mice, rats and non-human primates.Citation12-15 The role of bacteria is further supported by findings that osteoclastogenesis and alveolar bone resorption are enhanced by the application of bacteria.Citation16,17 Thus loss of function (gnotobiotic animals or use of anti-bacterials) and gain of function (the addition of bacteria) in a number of animal models (non-human primates, dogs, mice and rats) consistently establish a common role of bacteria in initiating the disease process.
More recently, mouse models have additionally facilitated a complete reappraisal of the role of certain organisms, such as Streptococcus gordonii, which have traditionally been considered as oral commensals. In vitro evidence indicates that S. gordonii can contribute to community pathogenicity by providing an attachment substratum for colonization by P. gingivalis.Citation18 Hence co-infection with S. gordonii and P. gingivalis in vivo would be predicted to enhance both P. gingivalis colonization and alveolar bone loss compared to monoinfection with P. gingivalis. Results from the oral gavage model support these predictions. Moreover, blocking attachment of P. gingivalis to S. gordonii ameliorates bone loss, thus opening a new avenue of research into therapeutic agents in periodontal disease.Citation19,20
In the murine abscess model, bacteria are delivered directly into the animal without the need for specialized colonization factors and the alveolar bone is not involved. Despite the limited applicability for periodontitis, the model does allow assessment of an organism's ability to resist immune killing, grow in vivo, and spread systemically. A recent successful use of the abscess model was to establish synergistic interactions between S. gordonii and Aggregatibacter actinomycetemcomitans. In vitro, growth of A. actinomycetemcomitans is enhanced through utilization of L-lactate produced as a metabolic by-product by S. gordonii.Citation21 In order for A. actinomycetemcomitans to cross-feed with S. gordonii, it produces catalase by which it overcomes the adverse effects of hydrogen peroxide released extracellularly by oral streptococci. The murine abscess model not only confirmed the importance of catalase but also established that dispersin B (DspB), an enzyme that dissolves A. actinomycetemcomitans biofilms, is necessary for nutritional synergism between A. actinomycetemcomitans and S. gordonii.Citation22 Specifically, 3D image analysis of the abscess material revealed that DspB is required for an optimal spatial organization of A. actinomycetemcomitans cells at >4 μm from S. gordonii cells, a distance that minimizes exposure to peroxide but allows access to L-lactate. Hence, provided the experimental questions are framed to fit the model system, even a rudimentary model such as abscess formation can provide valuable in vivo verification of processes identified in vitro.
A potential issue regarding the use of mouse models to study periodontal disease pathogenesis is that the periodontitis-associated microbiotas in mice and humans differ considerably. However, this is not a prohibitive factor for using mouse models since periodontitis is fundamentally a dysbiotic inflammatory disease precipitated by disruption of host-microbe homeostasis.Citation9,23 Dysbiosis is not dependent so much on the particular microbial roster but rather on the specific gene combinations or collective virulence activity within the altered microbial community.Citation24,25 This notion is supported by a recent metatranscriptomic study which showed that disease-associated microbial communities exhibit conserved metabolic and virulence gene expression profiles, despite high inter-patient variability in terms of microbial composition.Citation26 Therefore, a conserved periodontitis-associated microbiota is not a requirement for the pathogenesis of human periodontitis. This realization and the fact that periodontitis is not uniquely a human diseaseCitation27 and involves common pathogenic mechanisms among different mammalian species (see above) validates the use of animal models to study periodontitis. In a similar context, intestinal health requires maintaining a balance between the colonic epithelium, the immune system, and the resident microbiota, whereas the breakdown of this homeostatic relationship leads to inflammatory bowel disease (IBD).Citation28 As with periodontitis, this concept confers relevance to the use of mice as models for IBD pathogenesis despite the differences between the mouse and human microbiotas.
Animal models can also provide insights into better understanding of data from human microbiome studies. A recent study in the murine oral gavage model has shown that the oral commensal microbiota is absolutely required for induction of inflammatory bone loss by P. gingivalis, which has traditionally been considered a causative agent in human periodontitis.Citation29 Such commensals can act as pathobionts in a dysbiotic microbial community,Citation30,31 and in human periodontitis are likely represented by hitherto underappreciated species that have now been shown to have as good or better a correlation with disease as P. gingivalis (or other long-established pathogens).Citation32-35 Therefore, a concept first established in mice is consistent with and has explanatory power for results obtained from metagenomic analyses of human periodontitis. Moreover, the commensal-turned-pathobiont concept is supported by a recent metatranscriptomic study, which revealed that a plethora of virulence factors upregulated in the microbiome of periodontitis patients is primarily derived from the previously underappreciated species that were not traditionally associated with periodontitis.Citation36
The Role of the Host Immune Response
A controversy that has flared from time to time in the annals of periodontal research involves the role of the host response in periodontal destruction. That the host response and elements of innate or adaptive immunity can be protective has been shown by several studies. For instance, immunization of gnotobiotic rats against P. gingivalis, protects against bone loss induced by inoculation of this bacteriumCitation37 as does immunization in non-human primates and in mice.Citation38-41 Similar results have been obtained by adoptive transfer of T-helper lymphocytes.Citation42 Moreover, both humans and mice that fail to recruit neutrophils to the periodontal tissue (e.g., due to leukocyte-adhesion deficiency) develop an aggressive form of periodontitis early in life.Citation43 However, animal models also provide conclusive evidence that the host response is intimately involved in the destructive process. Both systemic and topical application of non-steroidal anti-inflammatory drugs that inhibit prostaglandin synthesis reduce periodontal bone loss in spontaneously occurring periodontal disease in dogs and in ligature-induced periodontal disease in non-human primates.Citation44,45 Similarly, application of factors that inhibit cytokines, including tumor necrosis factor [TNF], interleukin [IL]-1, IL-17, complement, and RANKL reduce periodontal tissue destruction whether induced by A. actinomycetemcomitans oral gavage or by ligatures in mice or non-human primates, providing additional evidence that the host response mediates bone loss.Citation46-55 Moreover, such studies have offered promising therapeutic targets for the treatment of human periodontitis. In contrast, application of IL-1 or TNF, or genetic over-expression, enhances bone loss triggered by bacteria.Citation56-58 Likewise, attenuation of the host response by genetic ablation generally lessens bacteria-induced bone loss.Citation59-63 Thus, animal studies consistently demonstrate that bacteria alone are not sufficient to induce periodontal bone loss, a conclusion that would be difficult to make solely from in vitro studies or human studies.
Recent studies have questioned the reliability of murine models for the investigation of human inflammatory disease, a broad conclusion which, if validated, would have a significant impact on the use of mouse models of periodontal disease. Specifically, gene expression profiling of C57BL/6J mice and humans during endotoxemia revealed poor correlation between the human genes and mouse orthologues and vice versa.Citation64 However, this shortcoming in fact does not apply to periodontitis where the same inflammatory mediators (e.g., prostaglandin E2, TNF, IL-1β, and IL-17) mediate inflammatory bone loss in various species including mice, rats, dogs, non-human primates, and humans.Citation43,46,54,55,63,65-68 Moreover, important innate or adaptive immune players implicated in experimental mouse periodontitis have been confirmed in higher animals. For instance the central complement component C3 promotes inflammatory periodontal bone loss in both mice and non-human primates,Citation55 whereas regulatory T cells mediate protection against the same condition in both mice and dogs.Citation67
When mouse models are used in an appropriate context to address specific hypotheses in periodontal disease pathogenesis, the results obtained have been consistent with in vitro observations using human cells. For instance, studies in the oral gavage model have confirmed the capacity of P. gingivalis to inhibit the expression of E-selectin and neutrophil-recruiting chemokines,Citation29 as predicted by the local chemokine paralysis model first established in vitro using endothelial and gingival epithelial cells.Citation69,70 Moreover, consistent with the requirement of intact C5a receptor (C5aR) signaling in human leukocytes for successful evasion of killing by P. gingivalis, the organism fails to colonize the periodontium of C5aR-deficient mice, in contrast to wild-type mice where P. gingivalis can persist and cause disease.Citation29,71,72 Moreover, local treatment of P. gingivalis-colonized mice with a C5aR antagonist essentially eliminates P. gingivalis, reverses its dysbiotic effect, and inhibits development of periodontitis.Citation29,71,73 In line with in vitro evidence that P. gingivalis can escape TLR4 recognition or activation,Citation74 TLR4-deficient neutrophils display normal inflammatory responses to P. gingivalis in the chamber model, comparable with wild-type neutrophils (but not TLR2-deficient neutrophils which exhibit a poor response).Citation5 Furthermore, the lipid A phosphatase activity of P. gingivalis, which is required for modulation of lipid A structure and hence evasion of TLR4,Citation74 was shown to contribute to the capacity of P. gingivalis for oral colonization and enhancement of the commensal bacterial load in a rabbit model of ligature-induced periodontitis.Citation75 These studies also justify the characterization of P. gingivalis as a keystone pathogen, a concept that is relevant also in other inflammatory dysbiotic diseases.Citation76,77 The consistency between in vivo animal and in vitro human experimental systems not only confers biological significance to the in vitro findings but also lends further support and validation of these animal models.
One potential limitation of rodent models is that the cells and effector molecules of the immune system can differ from their human counterparts as is the case with the neutrophil chemokine CXCL8/IL-8. Mice and rats do not produce IL-8, but they do produce functionally equivalent homologs that are controlled by the transcription factor NF-κB.Citation78-80 P. gingivalis can inhibit neutrophil transmigration toward human epithelial cells in vitroCitation81 through production of a serine phosphatase, SerB, that inhibits NF-κB activation by dephosphorylating its p65 subunit.Citation69,70 An important test of the relevance of rodent models then was to assess the functionality of SerB in vivo. In the rat oral gavage model, a SerB-deficient mutant of P. gingivalis incited greater neutrophil infiltration in gingival tissues.Citation82 Thus, even though specific immune effectors may differ between rodents and humans, similarity in the command and control pathways ensures that mice and rats can indeed model the human immune systems in many important ways.
In addition to inducing periodontitis via oral gavage or ligature placement, the disease can develop in mice spontaneously as a result of the aging process, a factor that also contributes to human periodontitis.Citation83 The use of the aging-associated periodontitis model led to the identification of a novel molecule involved in periodontal tissue homeostasis, namely the endothelial cell-derived glycoprotein Del-1.Citation54 Del-1 engages in reciprocal antagonistic interactions with IL-17 in terms of their expression and function in neutrophil recruitment and inflammation. This reciprocal relationship has been confirmed in humans, with Del-1 dominating in healthy gingiva and IL-17 prevailing in inflamed gingiva.Citation54
Importantly, the induction of periodontitis in mice involves more physiological means as compared to other widely used mouse models of other human diseases. For instance, chemically-induced models of IBD have limitations in understanding events that initiate gut inflammation in human IBD.Citation84 Psoriasis, a T-cell-mediated chronic inflammatory skin disease, is generally not seen in animals other than humans, yet, various mouse models including transgenics and knockouts have been developed that mimic psoriasis.Citation85 Despite their serious limitations, these models have established that psoriasis is a T-cell-mediated disease and have been used to dissect novel pathways of disease pathogenesis. In experimental autoimmune encephalomyelitis, a model of human multiple sclerosis, the disease is often induced artificially after injection of autoantigen emulsified in complete Freund's adjuvant. This promotes the induction of CD4+ T cell-mediated autoimmune mechanisms, whereas CD8+ T cells prevail in multiple sclerosis lesions.Citation86 Similarly, collagen-induced arthritis in mice, a commonly used model of rheumatoid arthritis, is elicited by immunization with type II collagen formulated in complete Freund's adjuvant.Citation87 Nevertheless, imperfect as they may be, these models have significantly enhanced our understanding of disease pathogenesis.
Impact of Systemic Disease
It is well documented in human studies that systemic conditions such as diabetes mellitus increase the risk and severity of periodontal disease.Citation88 Animal models have established a mechanistic basis for this phenomenon. Both type 1 and type 2 diabetic mice exhibit a greater inflammatory response than normal mice to the same inoculation of P. gingivalis into connective tissue.Citation89,90 If TNF is blocked in diabetic rats or diabetic mice, much of the diabetes-enhanced bone resorption is reversed, indicating that diabetes-enhanced inflammation, particularly TNF, is problematic.Citation91-93 Interestingly, diabetes appears to cause a particular problem in the resolution of inflammation which leads to dysregulation of a number of pathways that both enhance bone resorption and reduce coupled bone formation.Citation94,95 A number of factors may enhance inflammation in diabetic animals including increased formation of advanced glycation end products (AGEs).Citation8 When AGE signaling is blocked in a periodontal disease model both diabetes-enhanced TNF levels and periodontal bone loss are reduced.Citation96 Therefore, human studies have provided evidence of an association between diabetes, AGEs, inflammation and periodontal disease, but animal studies with the use of specific inhibitors provide conclusive evidence of functional relationships between these parameters and identify specific processes affected. Conversely, the notion that periodontitis exerts an adverse impact on systemic health is substantiated by mechanistic animal studies linking periodontitis or periodontal pathogens to disorders such as atherosclerosis, adverse pregnancy outcomes, and rheumatoid arthritis.Citation97,98
Conclusion
In summary, whereas no one animal model can recapitulate the complexity of periodontal disease, different aspects of the disease can be represented by different models, which have contributed considerably in dissecting the mechanistic underpinning of periodontitis. Of course, the synthesis and integration of findings from all available experimental systems (in vitro, animal, human) are required for better understanding of disease pathogenesis (). A good example of the interconnectivity and relevance of each experimental system is provided by the treatment of periodontitis with local delivery of tetracyclines. Tetracyclines have been shown to inhibit periodontal disease in rats and to alter the subgingival microflora in humans.Citation15,99 However, experiments with germ-free rats demonstrated that tetracyclines can reduce periodontal breakdown in a non-antimicrobial manner involving inhibition of matrix metalloproteinases (MMPs).Citation100 This led to a number of in vitro studies to investigate the precise mechanisms involved and the development of new drugs that inhibit MMP activity.Citation100 MMP-blocking drugs first discovered in rat models of periodontal disease have been subsequently marketed as Periostat® to prevent periodontitis in humansCitation101 and are being further developed for treatments of other tissue-breakdown diseases including cardiovascular disease.Citation102
When using animal models, what matters is not only the species but primarily the ways in which the chosen model is used. For instance, whereas non-human primate models are closer to human periodontitis than any dog, rabbit, or rodent model, no model can be discounted if used appropriately and the data are interpreted within the limitations of the model. It is the opinion of these authors that the dismissal of animal models on the grounds that they do not faithfully represent all aspects of human periodontitis does not constitute serious scientific criticism and, in the absence of better mechanistic alternatives, represents an impediment to scientific progress. Needless to add, however, that it is important to strive to optimize existing models or invent new and improved ones based on new experimental results and constructive criticism.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Funding
The authors’ research is supported by NIH grants; DE015254, DE017138, DE021685, DE024716, AI068730 (GH); DE011111, DE012505, DE016690, DE017921, DE022867, DE023193 (RJL); and DE017732, DE021921 (DTG).
Related Research Data
References
- Graves DT, Fine D, Teng YT, Van Dyke TE, Hajishengallis G. The use of rodent models to investigate host-bacteria interactions related to periodontal diseases. J Clin Periodontol 2008; 35:89-105; PMID:18199146; http://dx.doi.org/10.1111/j.1600-051X.2007.01172.xp
- Baker PJ, Dixon M, Roopenian DC. Genetic control of susceptibility to Porphyromonas gingivalis-induced alveolar bone loss in mice. Infect Immun 2000; 68:5864-8; PMID:10992496
- Abe T, Hajishengallis G. Optimization of the ligature-induced periodontitis model in mice. J Immunol Meth 2013; 394:49-54; PMID:23672778; http://dx.doi.org/10.1016/j.jim.2013.05.002
- Genco CA, Cutler CW, Kapczynski D, Maloney K, Arnold RR. A novel mouse model to study the virulence of and host response to Porphyromonas (Bacteroides) gingivalis. Infect Immun 1991; 59:1255-63; PMID:2004807
- Burns E, Bachrach G, Shapira L, Nussbaum G. Cutting Edge: TLR2 is required for the innate response to Porphyromonas gingivalis: activation leads to bacterial persistence and TLR2 deficiency attenuates induced alveolar bone resorption. J Immunol 2006; 177:8296-300; PMID:17142724
- Singh A, Wyant T, Anaya-Bergman C, Aduse-Opoku J, Brunner J, Laine ML, Curtis MA, Lewis JP. The capsule of Porphyromonas gingivalis leads to a reduction in the host inflammatory response, evasion of phagocytosis, and increase in virulence. Infect Immun 2011; 79:4533-42; PMID:21911459
- Kastelein P, van Steenbergen TJ, Bras JM, de Graaff J. An experimentally induced phlegmonous abscess by a strain of Bacteroides gingivalis in guinea pigs and mice. Antonie Van Leeuwenhoek 1981; 47:1-9; PMID:7247390
- Lalla E, Papapanou PN. Diabetes mellitus and periodontitis: a tale of two common interrelated diseases. Nat Rev Endocrinol 2011; 7:738-48; PMID:21709707; http://dx.doi.org/10.1038/nrendo.2011.106
- Darveau RP. Periodontitis: a polymicrobial disruption of host homeostasis. Nat Rev Microbiol 2010; 8:481-90; PMID:20514045; http://dx.doi.org/; http://dx.doi.org/10.1038/nrmicro2337
- Keyes PH, Jordan HV. Periodontal lesions in the syrian hamster. III. Findings related to an infectious and transmissible component. Arch Oral Biol 1964; 32:377-400; PMID:14179047; http://dx.doi.org/10.1016/0003-9969(64)90024-X
- Rovin S, Costich ER, Gordon HA. The influence of bacteria and irritation in the initiation of periodontal disease in germfree and conventional rats. J Periodontal Res 1966; 1:193-204; PMID:4225530; http://dx.doi.org/10.1111/j.1600-0765.1966.tb01860.x
- Williams RC, Leone CW, Jeffcoat MK, Nitzan D, Goldhaber P. Tetracycline treatment of periodontal disease in the beagle dog. II. The cultivable periodontal pocket flora. J Periodontal Res 1981; 16:666-74; PMID:6460865; http://dx.doi.org/10.1111/j.1600-0765.1981.tb01005.x
- Kenworthy R, Baverel M. Studies of a periodontal tissue lesion in the rat, untreated or treated with chlorhexidine digluconate. J Clin Periodontol 1981; 8:349-58; PMID:6947996; http://dx.doi.org/10.1111/j.1600-051X.1981.tb02044.x
- Luan Q, Desta T, Chehab L, Sanders VJ, Plattner J, Graves DT. Inhibition of experimental periodontitis by a topical boron-based antimicrobial. J Dent Res 2008; 87:148-52; PMID:18218841; http://dx.doi.org/10.1177/154405910808700208
- Weiner GS, DeMarco TJ, Bissada NF. Long term effect of systemic tetracycline administration on the severity of induced periodontitis in the rat. J Periodontol 1979; 50:619-23; PMID:294476; http://dx.doi.org/10.1902/jop.1979.50.12.619
- Samejima Y, Ebisu S, Okada H. Effect of infection with Eikenella corrodens on the progression of ligature-induced periodontitis in rats. J Periodontal Res 1990; 25:308-15; PMID:2145416
- Holt S, Ebersole J, Felton J, Brunsvold M, Kornman K. Implantation of Bacteroides gingivalis in non-human primates initiates progression of periodontitis. Science 1988; 239:55-7; PMID:3336774; http://dx.doi.org/10.1126/science.3336774
- Kuboniwa M, Tribble GD, James CE, Kilic AO, Tao L, Herzberg MC, Shizukuishi S, Lamont RJ. Streptococcus gordonii utilizes several distinct gene functions to recruit Porphyromonas gingivalis into a mixed community. Mol Microbiol 2006; 60:121-39; PMID:16556225; http://dx.doi.org/10.1111/j.1365-2958.2006.05099.x
- Daep CA, Novak EA, Lamont RJ, Demuth DR. Structural dissection and in vivo effectiveness of a peptide inhibitor of Porphyromonas gingivalis adherence to Streptococcus gordonii. Infect Immun 2011; 79:67-74; PMID:21041492; http://dx.doi.org/10.1128/IAI.00361-10
- Wright CJ, Wu H, Melander RJ, Melander C, Lamont RJ. Disruption of heterotypic community development by Porphyromonas gingivalis with small molecule inhibitors. Mol Oral Microbiol 2014; PMID:24899524
- Ramsey MM, Rumbaugh KP, Whiteley M. Metabolite cross-feeding enhances virulence in a model polymicrobial infection. PLOS Pathog 2011; 7:e1002012; PMID:21483753
- Stacy A, Everett J, Jorth P, Trivedi U, Rumbaugh KP, Whiteley M. Bacterial fight-and-flight responses enhance virulence in a polymicrobial infection. Proc Natl Acad Sci U S A 2014; 111:7819-24; PMID:24825893; http://dx.doi.org/10.1073/pnas.1400586111
- Hajishengallis G. Immunomicrobial pathogenesis of periodontitis: keystones, pathobionts, and host response. Trends Immunol 2014; 35:3-11; PMID:24269668; http://dx.doi.org/10.1016/j.it.2013.09.001
- Hajishengallis G, Lamont RJ. Beyond the red complex and into more complexity: The Polymicrobial Synergy and Dysbiosis (PSD) model of periodontal disease etiology. Mol Oral Microbiol 2012; 27:409-19; PMID:23134607; http://dx.doi.org/10.1111/j.2041-1014.2012.00663.x
- Knights D, Lassen KG, Xavier RJ. Advances in inflammatory bowel disease pathogenesis: linking host genetics and the microbiome. Gut 2013; 62:1505-10; PMID:24037875; http://dx.doi.org/10.1136/gutjnl-2012-303954
- Jorth P, Turner KH, Gumus P, Nizam N, Buduneli N, Whiteley M. Metatranscriptomics of the human oral microbiome during health and disease. MBio 2014; 5:e01012-14; PMID:24692635; http://dx.doi.org/10.1128/mBio.01012-14
- Page RC, Schroeder HE. Periodontitis in man and other animals—a comparative review. Basel, Switzerland: Karger; 1982.
- Nell S, Suerbaum S, Josenhans C. The impact of the microbiota on the pathogenesis of IBD: lessons from mouse infection models. Nat Rev Microbiol 2010; 8:564-77; PMID:20622892; http://dx.doi.org/10.1038/nrmicro2403
- Hajishengallis G, Liang S, Payne MA, Hashim A, Jotwani R, Eskan MA, McIntosh ML, Alsam A, Kirkwood KL, Lambris JD, et al. Low-abundance biofilm species orchestrates inflammatory periodontal disease through the commensal microbiota and complement. Cell Host Microbe 2011; 10:497-506; PMID:22036469; http://dx.doi.org/10.1016/j.chom.2011.10.006
- Hajishengallis G, Lamont RJ. Breaking bad: manipulation of the host response by Porphyromonas gingivalis. Eur J Immunol 2014; 44:328-38; PMID:24338806; http://dx.doi.org/10.1002/eji.201344202
- Jiao Y, Darzi Y, Tawaratsumida K, Marchesan JT, Hasegawa M, Moon H, Chen GY, Nunez G, Giannobile WV, Raes J, et al. Induction of bone loss by pathobiont-mediated nod1 signaling in the oral cavity. Cell Host Microbe 2013; 13:595-601; PMID:23684310; http://dx.doi.org/10.1016/j.chom.2013.04.005
- Dewhirst FE, Chen T, Izard J, Paster BJ, Tanner AC, Yu WH, Lakshmanan A, Wade WG. The human oral microbiome. J Bacteriol 2010; 192:5002-17; PMID:20656903; http://dx.doi.org/10.1128/JB.00542-10
- Abusleme L, Dupuy AK, Dutzan N, Silva N, Burleson JA, Strausbaugh LD, Gamonal J, Diaz PI. The subgingival microbiome in health and periodontitis and its relationship with community biomass and inflammation. ISME J 2013; 7:1016-25; PMID:23303375; http://dx.doi.org/10.1038/ismej.2012.174
- Griffen AL, Beall CJ, Campbell JH, Firestone ND, Kumar PS, Yang ZK, Podar M, Leys EJ. Distinct and complex bacterial profiles in human periodontitis and health revealed by 16S pyrosequencing. ISME J 2012; 6:1176-85; PMID:22170420; http://dx.doi.org/10.1038/ismej.2011.191
- Kumar PS, Leys EJ, Bryk JM, Martinez FJ, Moeschberger ML, Griffen AL. Changes in periodontal health status are associated with bacterial community shifts as assessed by quantitative 16S cloning and sequencing. J Clin Microbiol 2006; 44:3665-73; PMID:17021095; http://dx.doi.org/10.1128/JCM.00317-06
- Duran-Pinedo AE, Chen T, Teles R, Starr JR, Wang X, Krishnan K, Frias-Lopez J. Community-wide transcriptome of the oral microbiome in subjects with and without periodontitis. ISME J 2014; 8:1659-72; PMID:24599074; http://dx.doi.org/10.1038/ismej.2014.23
- Klausen B, Evans RT, Ramamurthy NS, Golub LM, Sfintescu C, Lee JY, Bedi G, Zambon JJ, Genco RJ. Periodontal bone level and gingival proteinase activity in gnotobiotic rats immunized with Bacteroides gingivalis. Oral Microbiol Immunol 1991; 6:193-201; PMID:1687484; http://dx.doi.org/10.1111/j.1399-302X.1991.tb00477.x
- Persson GR, Engel D, Whitney C, Darveau R, Weinberg A, Brunsvold M, Page RC. Immunization against Porphyromonas gingivalis inhibits progression of experimental periodontitis in nonhuman primates. Infect Immun 1994; 62:1026-31; PMID:8112836
- Gonzalez D, Tzianabos AO, Genco CA, Gibson FC, 3rd. Immunization with Porphyromonas gingivalis capsular polysaccharide prevents P. gingivalis-elicited oral bone loss in a murine model. Infect Immun 2003; 71:2283-7; PMID:12654858; http://dx.doi.org/10.1128/IAI.71.4.2283-2287.2003
- O'Brien-Simpson NM, Pathirana RD, Paolini RA, Chen YY, Veith PD, Tam V, Ally N, Pike RN, Reynolds EC. An immune response directed to proteinase and adhesin functional epitopes protects against Porphyromonas gingivalis-induced periodontal bone loss. J Immunol 2005; 175:3980-9; PMID:16148146; http://dx.doi.org/10.4049/jimmunol.175.6.3980
- Gibson FC, 3rd, Genco CA. Prevention of Porphyromonas gingivalis-induced oral bone loss following immunization with gingipain R1. Infect Immun 2001; 69:7959-63; PMID:11705986; http://dx.doi.org/10.1128/IAI.69.12.7959-7963.2001
- Yamashita K, Eastcott JW, Taubman MA, Smith DJ, Cox DS. Effect of adoptive transfer of cloned Actinobacillus actinomycetemcomitans-specific T helper cells on periodontal disease. Infect Immun 1991; 59:1529-34; PMID:1825991
- Moutsopoulos NM, Konkel J, Sarmadi M, Eskan MA, Wild T, Dutzan N, Abusleme L, Zenobia C, Hosur KB, Abe T, et al. Defective neutrophil recruitment in leukocyte adhesion deficiency type I disease causes local IL-17–driven inflammatory bone loss. Sci Transl Med 2014; 6:229ra40; PMID:24670684; http://dx.doi.org/10.1126/scitranslmed.3007696
- Williams R, Jeffcoat M, Kaplan M, Goldhaber P, Johnson H, Wechter W. Flurbiprofen: a potent inhibitor of alveolar bone resorption in beagles. Science 1985; 227:640-2; PMID:3969553; http://dx.doi.org/10.1126/science.3969553
- Li KL, Vogel R, Jeffcoat MK, Alfano MC, Smith MA, Collins JG, Offenbacher S. The effect of ketoprofen creams on periodontal disease in rhesus monkeys. J Periodontal Res 1996; 31:525-32; PMID:8971650; http://dx.doi.org/10.1111/j.1600-0765.1996.tb00516.x
- Assuma R, Oates T, Cochran D, Amar S, Graves D. IL-1 and TNF antagonists inhibit the inflammatory response and bone loss in experimental periodontitis. J Immun 1998; 160:403-9; PMID:9551997
- Delima A, Oates T, Assuma R, Schwartz Z, Cochran D, Amar S, Graves D. Soluble antagonists to interleukin-1 (IL-1) and tumor necrosis factor (TNF) inhibits loss of tissue attachment in experimental periodontitis. J Clin Periodontol 2001; 28:233-40; PMID:11284536; http://dx.doi.org/10.1034/j.1600-051x.2001.028003233.x
- Tomofuji T, Ekuni D, Irie K, Azuma T, Endo Y, Tamaki N, Sanbe T, Murakami J, Yamamoto T, Morita M. Preventive effects of a cocoa-enriched diet on gingival oxidative stress in experimental periodontitis. J Periodontol 2009; 80:1799-808; PMID:19905949; http://dx.doi.org/10.1902/jop.2009.090270
- Ekuni D, Tomofuji T, Irie K, Kasuyama K, Umakoshi M, Azuma T, Tamaki N, Sanbe T, Endo Y, Yamamoto T, et al. Effects of periodontitis on aortic insulin resistance in an obese rat model. Lab Invest 2010; 90:348-59; PMID:20065945; http://dx.doi.org/10.1038/labinvest.2009.141
- Endo Y, Tomofuji T, Ekuni D, Irie K, Azuma T, Tamaki N, Yamamoto T, Morita M. Experimental periodontitis induces gene expression of proinflammatory cytokines in liver and white adipose tissues in obesity. J Periodontol 2010; 81:520-6; PMID:20367095; http://dx.doi.org/10.1902/jop.2009.090574
- Teng Y, Nguyen H, Gao X, Kong Y, Gorczynski R, Singh B, Ellen R, Penninger J. Functional human T-cell immunity and osteoprotegerin ligand control alveolar bone destruction in periodontal infection. J Clin Invest, 2000; 106:R59-67; PMID:10995794; http://dx.doi.org/10.1172/JCI10763
- Han X, Kawai T, Eastcott JW, Taubman MA. Bacterial-responsive B lymphocytes induce periodontal bone resorption. J Immunol 2006; 176:625-31; PMID:16365458; http://dx.doi.org/10.4049/jimmunol.176.1.625
- Jin Q, Cirelli JA, Park CH, Sugai JV, Taba M, Jr., Kostenuik PJ, Giannobile WV. RANKL inhibition through osteoprotegerin blocks bone loss in experimental periodontitis. J Periodontol 2007; 78:1300-8; PMID:17608585; http://dx.doi.org/10.1902/jop.2007.070073
- Eskan MA, Jotwani R, Abe T, Chmelar J, Lim JH, Liang S, Ciero PA, Krauss JL, Li F, Rauner M, et al. The leukocyte integrin antagonist Del-1 inhibits IL-17-mediated inflammatory bone loss. Nat Immunol 2012; 13:465-73; PMID:22447028; http://dx.doi.org/10.1038/ni.2260
- Maekawa T, Abe T, Hajishengallis E, Hosur KB, DeAngelis RA, Ricklin D, Lambris JD, Hajishengallis G. Genetic and intervention studies implicating complement C3 as a major target for the treatment of periodontitis. J Immunol 2014; 192 6020-7; PMID:24808362; http://dx.doi.org/10.4049/jimmunol.1400569
- Koide M, Suda S, Saitoh S, Ofuji Y, Suzuki T, Yoshie H, Takai M, Ono Y, Taniguchi Y, Hara K. In vivo administration of IL-1 beta accelerates silk ligature-induced alveolar bone resorption in rats. J Oral Pathol Med 1995; 24:420-34; PMID:8537916; http://dx.doi.org/10.1111/j.1600-0714.1995.tb01212.x
- Gaspersic R, Stiblar-Martincic D, Osredkar J, Skaleric U. Influence of subcutaneous administration of recombinant TNF-alpha on ligature-induced periodontitis in rats. J Periodontal Res 2003; 38:198-203; PMID:12608915; http://dx.doi.org/10.1034/j.1600-0765.2003.01395.x
- Dayan S, Stashenko P, Niederman R, Kupper TS. Oral epithelial overexpression of IL-1alpha causes periodontal disease. J Dent Res 2004; 83:786-90; PMID:15381720; http://dx.doi.org/10.1177/154405910408301010
- Baker P, Evans R, Roopenian D. Oral infection with Porphyromonas gingivalis and induced alveolar bone loss in immunocompetent and severe combined immunodeficient mice. Arch Oral Biol 1994; 39:1035-40; PMID:7717884; http://dx.doi.org/10.1016/0003-9969(94)90055-8
- Baker P, Dixon M, Evans R, Dufour L, Johnson E, Roopenian D. CD4(+) T cells and the proinflammatory cytokines gamma interferon and interleukin-6 contribute to alveolar bone loss in mice. Infect Immun 1999; 67:2804-9; PMID:10338484
- Chiang C, Kyritsis G, Graves D, Amar S. Interleukin-1 and tumor necrosis factor activities partially account for calvarial bone resorption induced by local injection of lipopolysaccharide. Infection and Immunity 1999; 67:4231-6; PMID:10417196
- Graves D, Oskoui M, Volejnikova S, Naguib G, Cai S, Desta T, Kakouras A, Jiang Y. Tumor necrosis factor modulates fibroblast apoptosis, PMN recruitment, and osteoclast formation in response to P. gingivalis infection. J Dent Res 2001; 80:1875-9; PMID:11706944; http://dx.doi.org/10.1177/00220345010800100301
- Garlet GP, Cardoso CR, Campanelli AP, Ferreira BR, Avila-Campos MJ, Cunha FQ, Silva JS. The dual role of p55 tumour necrosis factor-alpha receptor in Actinobacillus actinomycetemcomitans-induced experimental periodontitis: host protection and tissue destruction. Clin Exp Immunol 2007; 147:128-38; PMID:17177972
- Seok J, Warren HS, Cuenca AG, Mindrinos MN, Baker HV, Xu W, Richards DR, McDonald-Smith GP, Gao H, Hennessy L, et al. Genomic responses in mouse models poorly mimic human inflammatory diseases. Proc Natl Acad Sci U S A 2013; 110:3507-12; PMID:23401516; http://dx.doi.org/10.1073/pnas.1222878110
- Ebersole JL, Kirakodu S, Novak MJ, Stromberg AJ, Shen S, Orraca L, Gonzalez-Martinez J, Burgos A, Gonzalez OA. Cytokine Gene Expression Profiles during Initiation, Progression and Resolution of Periodontitis. J Clin Periodontol 2014; 41:853-61; PMID:24975876; http://dx.doi.org/10.1111/jcpe.12286
- Graves D. Cytokines that promote periodontal tissue destruction. J Periodontol 2008; 79:1585-91; PMID:18673014
- Glowacki AJ, Yoshizawa S, Jhunjhunwala S, Vieira AE, Garlet GP, Sfeir C, Little SR. Prevention of inflammation-mediated bone loss in murine and canine periodontal disease via recruitment of regulatory lymphocytes. Proc Natl Acad Sci U S A 2013; 110:18525-30; PMID:24167272; http://dx.doi.org/; http://dx.doi.org/10.1073/pnas.1302829110
- Noguchi K, Ishikawa I. The roles of cyclooxygenase-2 and prostaglandin E2 in periodontal disease. Periodontol 2000 2007; 43:85-101; PMID:17214837; http://dx.doi.org/10.1111/j.1600-0757.2006.00170.x
- Darveau RP, Belton CM, Reife RA, Lamont RJ. Local chemokine paralysis, a novel pathogenic mechanism for Porphyromonas gingivalis. Infect Immun 1998; 66:1660-5; PMID:9529095
- Takeuchi H, Hirano T, Whitmore SE, Morisaki I, Amano A, Lamont RJ. The serine phosphatase SerB of Porphyromonas gingivalis suppresses IL-8 production by dephosphorylation of NF-κB RelA/p65. PLOS Pathog 2013; 9:e1003326; PMID:23637609
- Abe T, Hosur KB, Hajishengallis E, Reis ES, Ricklin D, Lambris JD, Hajishengallis G. Local complement-targeted intervention in periodontitis: proof-of-concept using a C5a receptor (CD88) antagonist. J Immunol 2012; 189:5442-8; PMID:23089394; http://dx.doi.org/10.4049/jimmunol.1202339
- Wang M, Krauss JL, Domon H, Hosur KB, Liang S, Magotti P, Triantafilou M, Triantafilou K, Lambris JD, Hajishengallis G. Microbial hijacking of complement-toll-like receptor crosstalk. Sci Signal 2010; 3:ra11; PMID:20159852
- Maekawa T, Krauss JL, Abe T, Jotwani R, Triantafilou M, Triantafilou K, Hashim A, Hoch S, Curtis MA, Nussbaum G, et al. Porphyromonas gingivalis manipulates complement and TLR signaling to uncouple bacterial clearance from inflammation and promote dysbiosis. Cell Host Microbe 2014; 15:768-78; PMID:24922578; http://dx.doi.org/10.1016/j.chom.2014.05.012
- Coats SR, Jones JW, Do CT, Braham PH, Bainbridge BW, To TT, Goodlett DR, Ernst RK, Darveau RP. Human Toll-like receptor 4 responses to P. gingivalis are regulated by lipid A 1- and 4'- phosphatase activities. Cell Microbiol 2009; 11:1587-99; PMID:19552698; http://dx.doi.org/10.1111/j.1462-5822.2009.01349.x
- Zenobia C, Hasturk H, Nguyen D, Van Dyke TE, Kantarci A, Darveau RP. Porphyromonas gingivalis lipid A phosphatase activity is critical for colonization and increasing the commensal load in the rabbit ligature model. Infect Immun 2014; 82:650-9; PMID:24478080; http://dx.doi.org/10.1128/IAI.01136-13
- Stecher B, Maier L, Hardt WD. 'Blooming' in the gut: how dysbiosis might contribute to pathogen evolution. Nat Rev Microbiol 2013; 11:277-84; PMID:23474681; http://dx.doi.org/10.1038/nrmicro2989
- Hajishengallis G, Darveau RP, Curtis MA. The keystone-pathogen hypothesis. Nat Rev Microbiol 2012; 10:717-25; PMID:22941505; http://dx.doi.org/10.1038/nrmicro2873
- Ohmori Y, Fukumoto S, Hamilton TA. Two structurally distinct kappa B sequence motifs cooperatively control LPS-induced KC gene transcription in mouse macrophages. J Immunol 1995; 155:3593-600; PMID:7561058
- Tateno N, Matsumoto N, Motowaki T, Suzuki K, Aratani Y. Myeloperoxidase deficiency induces MIP-2 production via ERK activation in zymosan-stimulated mouse neutrophils. Free Radic Res 2013; 47:376-85; PMID:23438680; http://dx.doi.org/10.3109/10715762.2013.778990
- Chandrasekar B, Smith JB, Freeman GL. Ischemia-reperfusion of rat myocardium activates nuclear factor-KappaB and induces neutrophil infiltration via lipopolysaccharide-induced CXC chemokine. Circulation 2001; 103:2296-302; PMID:11342480; http://dx.doi.org/10.1161/01.CIR.103.18.2296
- Madianos PN, Papapanou PN, Sandros J. Porphyromonas gingivalis infection of oral epithelium inhibits neutrophil transepithelial migration. Infect Immun 1997; 65:3983-90; PMID:9316996
- Bainbridge B, Verma RK, Eastman C, Yehia B, Rivera M, Moffatt C, Bhattacharyya I, Lamont RJ, Kesavalu L. Role of Porphyromonas gingivalis phosphoserine phosphatase enzyme SerB in inflammation, immune response, and induction of alveolar bone resorption in rats. Infect Immun 2010; 78:4560-9; PMID:20805334; http://dx.doi.org/10.1128/IAI.00703-10
- Hajishengallis G. Aging and its impact on innate immunity and inflammation: implications for periodontitis. J Oral Biosci 2014; 56:30-7; PMID:24707191; http://dx.doi.org/10.1016/j.job.2013.09.001
- Barnett M, Fraser A. Animal models of colitis: lessons learned, and their relevance to the clinic. In: O'Connor M, ed. Ulcerative colitis—treatments, special populations and the future. Rijeka, Croatia; Shangai, China: InTech, 2011:161-78.
- Gudjonsson JE, Johnston A, Dyson M, Valdimarsson H, Elder JT. Mouse models of psoriasis. J Invest Dermatol 2007; 127:1292-308; PMID:17429444; http://dx.doi.org/10.1038/sj.jid.5700807
- t Hart BA, Gran B, Weissert R. EAE: imperfect but useful models of multiple sclerosis. Trends Mol Med 2011; 17:119-25; PMID:21251877; http://dx.doi.org/10.1016/j.molmed.2010.11.006
- Brand DD, Latham KA, Rosloniec EF. Collagen-induced arthritis. Nat Protoc 2007; 2:1269-75; PMID:17546023; http://dx.doi.org/10.1038/nprot.2007.173
- Chapple IL, Genco R, working group 2 of the joint EFPAAPw. Diabetes and periodontal diseases: consensus report of the Joint EFP/AAP workshop on periodontitis and systemic diseases. J Periodontol 2013; 84:S106-12; PMID:23631572; http://dx.doi.org/10.1902/jop.2013.1340011
- Naguib G, Al-Mashat H, Desta T, Graves D. Diabetes prolongs the inflammatory response to a bacterial stimulus through cytokine dysregulation. J Invest Dermatol 2004; 123:87-92; PMID:15191547; http://dx.doi.org/10.1111/j.0022-202X.2004.22711.x
- Graves DT, Naguib G, Lu H, Leone C, Hsue H, Krall E. Inflammation is more persistent in Type 1 diabetic mice. J Dent Res 2005; 84:324-8; PMID:15790737; http://dx.doi.org/10.1177/154405910508400406
- Al-Mashat HA, Kandru S, Liu R, Behl Y, Desta T, Graves DT. Diabetes enhances mRNA levels of proapoptotic genes and caspase activity, which contribute to impaired healing. Diabetes 2006; 55:487-95; PMID:16443785; http://dx.doi.org/10.2337/diabetes.55.02.06.db05-1201
- Liu R, Bal HS, Desta T, Behl Y, Graves DT. Tumor necrosis factor-alpha mediates diabetes-enhanced apoptosis of matrix-producing cells and impairs diabetic healing. Am J Pathol 2006; 168:757-64; PMID:16507891; http://dx.doi.org/10.2353/ajpath.2006.050907
- Pacios S, Kang J, Galicia J, Gluck K, Patel H, Ovaydi-Mandel A, Petrov S, Alawi F, Graves DT. Diabetes aggravates periodontitis by limiting repair through enhanced inflammation. FASEB J 2012; 26:1423-30; PMID:22179526; http://dx.doi.org/10.1096/fj.11-196279
- Pacios S, Andriankaja O, Kang J, Alnammary M, Bae J, de Brito Bezerra B, Schreiner H, Fine DH, Graves DT. Bacterial infection increases periodontal bone loss in diabetic rats through enhanced apoptosis. Am J Pathol 2013; 183:1928-35; PMID:24113454; http://dx.doi.org/10.1016/j.ajpath.2013.08.017
- Kang J, de Brito Bezerra B, Pacios S, Andriankaja O, Li Y, Tsiagbe V, Schreiner H, Fine DH, Graves DT. Aggregatibacter actinomycetemcomitans infection enhances apoptosis in vivo through a caspase-3-dependent mechanism in experimental periodontitis. Infection and immunity 2012; 80:2247-56; PMID:22451521; http://dx.doi.org/10.1128/IAI.06371-11
- Lalla E, Lamster IB, Feit M, Huang L, Spessot A, Qu W, Kislinger T, Lu Y, Stern DM, Schmidt AM. Blockade of RAGE suppresses periodontitis-associated bone loss in diabetic mice. J Clin Invest 2000; 105:1117-24; PMID:10772656; http://dx.doi.org/10.1172/JCI8942
- Slocum C, Coats SR, Hua N, Kramer C, Papadopoulos G, Weinberg EO, Gudino CV, Hamilton JA, Darveau RP, Genco CA. Distinct lipid A moieties contribute to pathogen-induced site-specific vascular inflammation. PLOS Pathog 2014; 10:e1004215; PMID:25010102
- Maresz KJ, Hellvard A, Sroka A, Adamowicz K, Bielecka E, Koziel J, Gawron K, Mizgalska D, Marcinska KA, Benedyk M, et al. Porphyromonas gingivalis facilitates the development and progression of destructive arthritis through its unique bacterial peptidylarginine deiminase (PAD). PLOS Pathog 2013; 9:e1003627; PMID:24068934
- Slots J, Rosling BG. Suppression of the periodontopathic microflora in localized juvenile periodontitis by systemic tetracycline. J Clin Periodontol 1983; 10:465-86; PMID:6579058; http://dx.doi.org/10.1111/j.1600-051X.1983.tb02179.x
- Golub LM, Lee HM, Lehrer G, Nemiroff A, McNamara TF, Kaplan R, Ramamurthy NS. Minocycline reduces gingival collagenolytic activity during diabetes. Preliminary observations and a proposed new mechanism of action. J Periodontal Res 1983; 18:516-26; PMID:6315909; http://dx.doi.org/10.1111/j.1600-0765.1983.tb00388.x
- Caton J, Ryan ME. Clinical studies on the management of periodontal diseases utilizing subantimicrobial dose doxycycline (SDD). Pharmacol Res 2011; 63:114-20; PMID:21182947; http://dx.doi.org/10.1016/j.phrs.2010.12.003
- Iyer RP, Patterson NL, Fields GB, Lindsey ML. The history of matrix metalloproteinases: milestones, myths, and misperceptions. Am J Physiol Heart Circ Physiol 2012; 303:H919-30; PMID:22904159; http://dx.doi.org/10.1152/ajpheart.00577.2012