Abstract
Anti-angiogenic vascular endothelial growth factor A (VEGF)165b and pro-angiogenic VEGF165 are generated from the same transcript, and their relative amounts are dependent on alternative splicing. The role of VEGF165b has not been investigated in as much detail as VEGF165, although it appears to be highly expressed in non-angiogenic tissues and, in contrast with VEGF165, is downregulated in tumors and other pathologies associated with abnormal neovascularization such as diabetic retinopathy or Denys Drash syndrome. VEGF165b inhibits VEGFR2 signaling by inducing differential phosphorylation, and it can be used to block angiogenesis in in vivo models of tumorigenesis and angiogenesis-related eye disease. Recent reports have identified three serine/arginine-rich proteins, SRSF1, SRSF2 and SRSF6, and studied their role in regulating terminal splice-site selection. Since the balance of VEGF isoforms is lost in cancer and angiogenesis-related conditions, control of VEGF splicing could also be used as a basis for therapy in these diseases.
Cells of pluricellular organisms require access to the circulation to subsist. Blood vessels carry the nutrients and oxygen necessary for the survival of these cells, and remove metabolic waste and carbon dioxide away from them. In the adult, the formation of new blood vessels is a tightly regulated phenomenon, becoming activated during brief periods of time such as wound healing or the female reproductive cycle, and subsequently inhibited.Citation1 In a physiological context, stimulators and inhibitors apply positive and negative influences on blood vessel formation to ensure that it is limited to the demands of the growing tissue. This balance is lost in many pathological conditions, leading to an angiogenic phenotype. Vascular endothelial growth factor (VEGF) is a key contributor to this process in health and disease. The VEGF family of growth factors includes VEGF-A, placental growth factor (PGF), VEGF-B, VEGF-C and VEGF-D, being VEGF-A the most important member of the family.
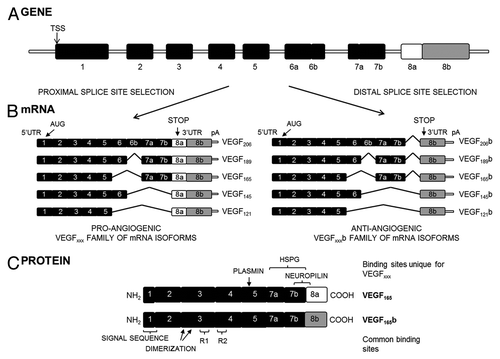
The human vegf-a gene contains eight exons and seven introns.Citation2 Alternative splicing and proteolytic processing of VEGF-A produces various isoforms with distinct biological activities ().Citation3-Citation8 The particular splicing event in the terminal exon creates two whole families of isoforms: the classic pro-angiogenic VEGFxxx isoforms, generated by proximal splice-site (PSS) selection, and the more recently described anti-angiogenic VEGFxxxb isoforms, formed by distal splice-site (DSS) choice, 66 bases downstream ().Citation9 This process gives rise to two families of polypeptides with the same size but different sequence at the C-terminus, maintaining the two of them the dimerization and receptor binding domains unchanged.Citation3 Exon 8a encodes Cys-Asp-Lys-Pro-Arg-Arg instead of the Ser-Leu-Thr-Arg-Lys-Asp sequence encoded by exon 8b. Therefore, the basic properties conferred by the two arginines present in 8a are altered, resulting in the neutral charge conferred by the residues lysine and aspartic acid, and loss of one disulfide bond. These conformational changes result in the inability of VEGFxxxb to bind the VEGFR2 co-receptor neuropilin 1 (NRP1), and have been proposed as the cause for the differences in downstream signaling ().Citation10,Citation11
Figure 1.vegf-a. (A) Gene structure. TSS is the transcriptional start site. (B) mRNA species. Alternative splicing of the vegf-a gene in the terminal exon results in two families of isoforms: the pro-angiogenic VEGFxxx and the anti-angiogenic VEGFxxxb isoforms. AUG, the start site for translation; UTR, the untranslated region; pA, the polyadenylation site. (C) Protein structure of the two major isoforms of each family, VEGF165 and VEGF165b. C-terminal splicing leads to an alternative last six amino acids (CDKPRR or SLTRKD). The isoforms are termed according to the amino acid number of the resulting protein (xxx). HSPG, heparin sulfate proteoglycan; R1, VEGF Receptor 1; R2, VEGF Receptor 2 (from ref. Citation29).
VEGF165b Physiological Expression and Function
VEGF165b, the major anti-angiogenic isoform, was the first member of the VEGFxxxb family to be describedCitation3 and is the most studied member so far. VEGF165b mRNA was first isolated and cloned from human renal cortex tissue, and subsequently identified in other human tissues. In fact, anti-angiogenic VEGF isoforms have been reported to represent a predominant proportion of the total VEGF protein found in normal human tissues such as vitreous fluid, circulating plasma, urine, renal cortex and glomeruli, colonic epithelium, bladder, smooth muscle, lung and pancreatic islets.Citation9,Citation12,Citation13 On the contrary, in placenta, a tissue where angiogenesis takes place under physiological conditions, VEGFxxxb represents only 1.5% of total VEGF.Citation12 In vitro, VEGFxxxb also predominates in primary cultured cells, such as differentiated podocytes or retinal pigmented epithelial cells. VEGFxxxb isoforms have also been reported in other species including pig,Citation14 rabbit, mouseCitation15 and rat,Citation16,Citation17 although its relative expression has been found to be decreased from humans to lower mammals, suggesting that the inclusion level of exon 8b, as well as the complexity of the regulatory mechanism of its splicing, increases through the evolutionary process from mouse to human.Citation15 The splice site is not conserved in lower vertebrates,Citation3 and there is no information on the existence of VEGF165b outside mammals.
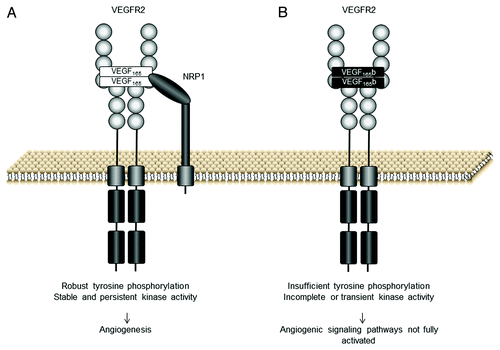
VEGF165 binding to VEGFR2 and NRP1 results in dimerization of the receptor and rotation of its intracellular domain, leading to its autophosphorylation ().Citation18 By contrast, this full rotational change is predicted to not occur when VEGF165b binds VEGFR2, resulting in inefficient autophosphorylation of the receptor (). In fact, the preference of VEGF165b toward the various tyrosine phosphorylation sites of the receptor is different than that observed after VEGF165 activation. Particularly, tyrosine 1054Citation18 in the kinase regulatory site is not phosphorylated following VEGF165b-VEGFR2 binding. Once VEGF165b has bound the receptor, what happens downstream? Little is known about the mechanisms of VEGF165b signaling. VEGF165b stimulates VEGFR2, ERK1/2 and Akt phosphorylation in endothelial cells.Citation8,Citation19 However, VEGF165b-induced phosphorylation of ERK1/2 has been observed to be more transient and weaker than that promoted by VEGF165.Citation4,Citation8,Citation18,Citation19
Figure 2. VEGF165b and VEGF165 interaction with VEGF receptor 2 (VEGFR2). (A) The VEGFR2 binding site of VEGF165 interacts with the VEGFR2 extracellular domain. VEGF165 functions as a dimer and promotes the formation of VEGFR2 dimers resulting in activation of the kinase domains via the phosphorylation of its tyrosine residues. Robust tyrosine phosphorylation results in the activation of angiogenic signaling pathways. (B) VEGF165b binds the VEGFR2 binding site with equal affinity to VEGF165 but does not bind neuropilin 1 (NRP1) co-receptor. The C-terminus of VEGF165b is neutral and there is insufficient torsional rotation for complete tyrosine phosphorylation, although weak phosphorylation can occur (adapted from ref. Citation11)
VEGF165 and VEGF165b appear to have contrasting physiological roles. The functions of VEGF165b are summarized in .
Table 1. VEGF165b physiological functions
Regulation of VEGF165b Expression
VEGF165b expression can be regulated by growth factors, including insulin-like growth factor 1 (IGF-1), tumor necrosis factor α (TNFα) and transforming growth factor β (TGFβ) in retinal pigmented epithelial cells, podocytes and dermal microvascular endothelial cells (MVECs). IGF-1 and TNFα have been shown to modulate VEGF splicing preferentially at the PSS increasing the expression of pro-angiogenic VEGF isoforms. In podocytes, IGF-mediated VEGFxxx upregulation occurs via stimulation of protein kinase C (PKC), which in turn activates the serine/arginine protein kinase 1 (SRPK1). Finally, SRPK1 phosphorylates the serine/arginine-rich splicing factor 1 (SRSF1), also known as ASF/SF2. Once activated, SRSF1, previously identified as an oncogene due to its transforming capacity in vitro and in vivo,Citation20 is able to bind VEGF pre-mRNA in the area adjacent to the PSS promoting VEGFxxx expression ().Citation21,Citation22
Figure 3. Regulation of VEGF alternative splicing in the terminal exon.
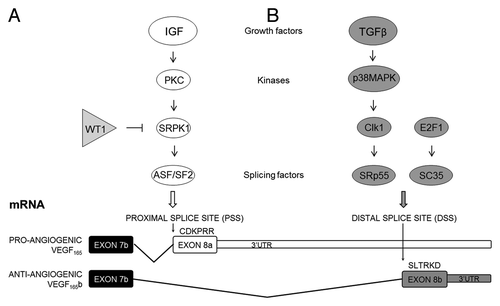
On the other hand, TGFβ upregulates anti-angiogenic VEGF isoforms most likely through stimulation of the p38 MAPK pathway and successive downstream activation of Clk/sty kinases, which results in phosphorylation of the serine/arginine-rich splicing factor 6 (SRSF6), also known as SRp55. SRSF6 binds directly to VEGF pre-mRNA in the exon 8b region favoring DSS selection and therefore, VEGFxxxb expression ().Citation22,Citation23 Finally, transcription factor 1 (E2F1) also favors the expression of the anti-angiogenic isoforms in human lung carcinoma cell lines through upregulation of another serine/arginine-rich splicing factor, SRSFR2, also known as SC35 ().Citation24 SRSF2 overexpression is able to mimic the E2F1-induced VEGF165b expression in vitro, and decreases tumor neovascularization in vivo. In contrast, a direct correlation between VEGFxxx and E2F1 has been observed in colorectal cancer specimensCitation25 suggesting that the regulation of vegf-a splicing by E2F1, and in general by SR splicing factors in human tumor cells, could be more complex and might depend on different factors such as the cell type, the amount of the different SR proteins, or other upstream signals.
VEGF165b in Pathology
The role of VEGF165b in cancer
Vasculature not only provides tumors with an adequate blood supply, but it offers a route for tumor cells to metastasize.Citation26 An upregulation of the pro-angiogenic VEGFxxx variants has been widely reported in human tumors. This upregulation brings about a loss in the balance of isoforms, which causes a drop in the proportion of VEGFxxxb levels. Indeed, VEGFxxxb is downregulated in several adult epithelial cancer types including melanoma,Citation27 renalCitation8 or colorectal carcinoma.Citation25,Citation28 An imbalance of the expression of the two VEGF families of isoforms has also been observed in the pediatric cancer neuroblastoma.Citation29 In contrast, VEGFxxxb is upregulated in intraductal breast carcinoma.Citation19
Overexpression of VEGF165b has been reported to delay the growth of melanoma,Citation8 kidney,Citation30 colon,Citation28 prostate and Ewing sarcoma tumors in mice.Citation31 Recombinant human VEGF165b (rhVEGF165b) treatment in vivo also has a growth-inhibitory effect in nude mice xenograft models of melanoma,Citation8 prostate cancer, renal cell carcinoma, Ewing’s sarcoma,Citation31 colorectal carcinomaCitation28 and neuroblastoma.Citation29 Likewise, rhVEGF165b parenteral administration reduces the growth of melanoma metastases.Citation13 These studies highlight the potential role of VEGF165b as an anti-cancer agent. However, a proper selection of patients that can benefit from this treatment is essential, as rhVEGF165b-based therapies are likely to be effective only in tumors with high VEGF expression vs. those with low VEGF levels, which must rely on other pro-angiogenic factors for their growth.Citation19
Finally, it has been shown that the VEGFxxx/VEGFxxxb ratio has an effect on the sensitivity of tumors to bevacizumab, an anti-VEGF antibody licensed for use in cancer treatment: as both VEGF165 and VEGF165b can bind bevacizumab with a similar affinity, the presence of VEGFxxxb can inhibit the effect of this drug through reducing the amounts of antibody available to bind VEGFxxx.Citation28 Hence, despite having a slower growing rate, tumors with high concentrations of VEGFxxxb might be more resistant to this particular therapy. One alternative treatment option for these tumors would be to use VEGFxxx-specific antibodies, which would inhibit exclusively the pro-angiogenic isoforms. Another possibility would be to develop molecules able to favor the DSS selection, and thus increase the endogenous expression of VEGFxxxb. Indeed, it has been shown that knocking down SRPK1 in a colorectal tumor cell line sensitive to the anti-angiogenic actions of VEGF165b, decreases tumor growth in vivo.Citation32
The role of VEGF165b in eye disease
A high expression of VEGF165b has been reported in human eye tissues, such as the retina or vitreous fluid,Citation9 and in the rodent eye.Citation17,Citation33 However, VEGF165b is relatively downregulated in vitreous fluid of diabetic retinopathy patients, resulting in a switch to an angiogenic phenotype.Citation9 In fact, proliferative diabetic retinopathy is characterized by an abnormal growth of new blood vessels into the vitreous fluid of the eye, stimulated by hypoxia-induced VEGF expression. In a mouse model of oxygen-induced retinopathy (OIR), the development of neovascularization in the retina correlated with an increase of total VEGF, and a specific decrease of VEGF165b, indicating a pro-angiogenic shift.Citation33 In this OIR mouse model, intraocular injection of rhVEGF165b also diminished pre-retinal neovascularization yet permitting the physiological angiogenesis in the inner retina to proceed normally.Citation34 This lack of intra-retinal neovascularization could be a consequence of the cytoprotective effects of VEGF165b on endothelial cells.Citation35 Thus, treatments with VEGF165b would not only inhibit neovascularization but still protect existing endothelial cells in a disease in which angiogenesis is frequently a consequence of endothelial cell loss and ischemia.Citation35 The VEGF balance is also altered in other angiogenic-associated eye conditions, including retinal vein occlusionCitation36 or neovascular age-related macular degeneration (AMD), and rhVEGF165b acted as a powerful anti-angiogenic agent when injected either subcutaneously or intraocularly in a mouse model of AMD, where choroidal neovascularization was laser-induced.Citation37 These results, together with the cytoprotection that VEGF165b confers to cultured retinal epithelial cells,Citation35 point to this anti-angiogenic isoform as a potential therapeutic agent in angiogenesis-associated eye diseases.
In contrast, in avascular eye diseases, including glaucoma,Citation17 rhegmatogenous retinal detachment and proliferative vitreoretinopathy,Citation38 VEGFxxxb has been found to be upregulated suggesting that, besides impeding neovascularization, VEGF165b might play a role in the pathogenesis of these conditions that are not angiogenesis-related.
The role of VEGF165b in renal disease
VEGF165b expression has been reported in the glomeruli of human kidneys.Citation13 However, whereas overexpression of VEGF164 in mouse glomeruli leads to proteinuria, glomerular dysfunction and renal failure,Citation39 a transgenic mouse model where VEGF165b was overexpressed in podocytes, did not have consequences on renal function. Nevertheless, closer observations of these podocytes showed decreased glomerular water permeability, which was dose-dependent.Citation40 In addition, ex vivo incubation of wild-type mouse glomeruli with VEGF165b, decreased permeability to water,Citation41 and constitutive overexpression of VEGF165b could prevent the observed VEGF164-driven increased water permeability and the changes in glomerular ultrastructure in vivo,Citation42 indicating that maintaining an appropriate VEGF isoform balance is also important in the glomerular function of the kidney.
Changes in the balance of VEGF-A variants were also found to be important in Denys Drash syndrome, a urogenital disorder associated with nephropathy and high risk for Wilms tumors. While pro-angiogenic VEGF165 was upregulated, VEGF165b was completely abrogated in glomeruli of patients with Denys Drash syndrome (DDS) suggesting a switch in splicing.Citation43 The authors pointed out that aberrant expression of the WT1 gene may be involved in regulating the vegf-a gene expression, and is the cause of glomerulopathy in DDS. In fact, WT1 in wild-type podocytes could bind the SRPK1 promoter inhibiting its expression (), whereas the mutated form of WT1 in podocytes isolated from a DDS patient did not bind this site, and SRPK1 expression was upregulated. Wild-type WT1 restoration decreased SRPK1 expression in these podocytes.Citation22,Citation32 Nevertheless, WT1 is known for being able to both repress and activate its target genes, hence its transcription function is highly context-specific and can be modulated by several cofactors.Citation44 Thus, modifying vegf-a splicing by preventing SRPK1 activity may be therapeutically effective in DDS.
The role of VEGF165b in fertility control and pregnancy-related conditions
A transgenic mouse model, whereby overexpression of VEGF165b is driven by the mouse mammary tumor virus promoter, shows expression of VEGF165b in tissues such as the ovary or the mammary tissue during mammary development of female mice. These mice had fewer blood vessels in the mammary tissue, an impaired alveolar vascular coverage of the fat pad and a reduced production of milk for nourishment of their pups, which died of starvation shortly after birth.Citation45 Whereas the litter size of wild-type females mated with transgenic males was normal, when transgenic females were mated with wild-type mice, significantly fewer pups were born. However, no developmental defects in the embryos were detected in culture which, together with the normal birth weight of the pups, indicated that the reduction in litter size could depend on ovarian function, as follicular development in the ovary is angiogenesis-dependent. Indeed, VEGF165b overexpression in the mouse ovary caused an increase in the ovulatory cycle length and a delayed follicular development, resulting in fertility defects in these mice.Citation45 Additionally, VEGF165b neutralization with an antibody specific for VEGFxxxb, as well as treatment with the pro-angiogenic isoform VEGF164, showed an enhanced vascular and follicular development in perinatal rat ovaries.Citation16
Total VEGF levels are upregulated and VEGFxxxb isoforms are expressed at even lower levels than usual in the placenta of patients with pre-eclampsia,Citation46 a condition in which the vasculature of pregnant women is vasoconstricted and shows higher permeability.Citation47 Circulating VEGF165b levels are downregulated during the first trimester in pregnancies that will develop pre-eclampsia, making it a useful clinical marker for high risk of pre-eclampsia.Citation48 Although after the first trimester circulating VEGFxxxb levels are normal in pre-eclamptic patients, pre-eclamptic plasma has been shown to increase vascular permeability. This increase could be inhibited by incubating the pre-eclamptic plasma with an antibody specific for VEGFxxxb, suggesting that VEGFxxxb is responsible for the enhanced vascular permeability in pre-eclamptic plasma.Citation47 Therefore, VEGF165b plays a key role not only in reproductive biology, but also in the control of fertility and pre-eclampsia.
The role of VEGF165b in systemic sclerosis
VEGF165b upregulation has been reported in the skin and circulation of patients with systemic sclerosis (SSc), a chronic disease characterized by impaired angiogenesis and vascular repair that affects the skin and internal organs.Citation49,Citation50 Cultured dermal microvascular endothelial cells (MVECs) isolated from SSc patients expressed higher levels of VEGF165b and VEGFR2 than those from healthy individuals. In spite of this, VEGFR2 phosphorylation was reduced and angiogenesis, defective as a consequence. Furthermore, treatment with rhVEGF165b, as well as treatment with conditioned media from SSc MVECs, inhibited VEGF165-induced VEGFR2 phosphorylation and angiogenesis in healthy MVECs, and these anti-angiogenic effects could be neutralized by treatment with a VEGF165b-blocking antibody.Citation23 Thus, neutralization of the elevated levels of VEGF165b might represent a potential therapeutical approach to stimulate a proper angiogenic response in SSc patients.
In addition, the high VEGF165b levels in SSc skin correlated with increased expression of TGFβ and SRSF6, and expression of VEGF165b and SRSF6 was found to be increased and regulated by TGFβ in cultered SSc MVECs, indicating that in SSc, and other non-angiogenic disorders, favoring PSS rather than DSS selection might represent a potential therapeutic strategy to stimulate adequate angiogenesis.
Perspective
Every year there are more and more studies on VEGF165b, and new conditions where VEGF165b is identified as altered or contributory—for instance a polymorphisim in intron 7 that appears to regulate VEGF165b expression predicts susceptibility to asthma.Citation51 Since its discovery, over 50 publications have described VEGF165b as an anti-angiogenic agent that is downregulated in cancer and other angiogenesis-related conditions such as diabetic retinopathy, and plays an important role in permeability regulation and renal disease, pre-eclampsia and fertility control. Nevertheless, there are researchers who are still skeptical about VEGF165b predominance in human or even existence in mouse tissue.Citation52 Indeed, despite being discovered in 2002, few reports on the splicing mechanism that gives rise to VEGFxxxb and its effects on angiogenesis-related diseases have been published during the last decade, and its downstream signaling effects remain practically unreported. One of the reasons that could explain this lack of research is the difficulty of quantifying the absolute amounts of VEGFxxx and VEGFxxxb mRNA and protein.Citation52,Citation53 The 3′UTR secondary structure of VEGF165 and VEGF165b transcripts is likely to be different, and result in differential reverse transcription efficiency. In fact, VEGF165 has been shown to be preferentially amplified by RT-PCR when mixed at equal amounts or even in excess of VEGF165b cDNA.Citation53 When VEGF was first identified, the importance of ruling out misamplification of VEGF165 by VEGF165b primers was highlighted.Citation3 A recent publication reiterated the importance of primer design in avoiding annealing of the reverse primer to VEGFxxx and amplification of a misprimed product,Citation52 although in that report, the authors did not use positive control DNA to ensure that the primers act specifically, and amplify the VEGFxxxb products efficiently. Mispriming is easily prevented by using primers that can simultaneously amplify both VEGFxxx and VEGFxxxb transcripts, and by subsequent sequencing of the obtained amplicons to confirm their nature, while using control DNA to account for differential amplification. Moreover, there is no commercially available VEGFxxx-specific antibody, and the sensitivity of the currently existing VEGFxxxb ELISA for the different isoforms and heterodimers of these isoforms has not been tested. The total VEGF ELISA, for example, underestimates the total VEGF levels as it has less affinity for VEGF165b than for VEGF165.Citation28 Therefore, further efforts need to be made to develop appropriate and reliable methods that allow the accurate quantification of both VEGFxxx and VEGFxxxb isoforms.
Growing evidence shows that vegf-a alternative splicing can be differentially regulated in conditions characterized by an excess of angiogenesis, such as cancer, or a deficit, such as SSc, but the control of this splicing event is still not well understood, and determining how different key splicing factors regulate an angiogenic splicing switch should be studied much more in depth over the next few years. The cellular machinery underlying the control of splice-site selection, and consequently which isoforms are being expressed to the detriment of others, represents a promising field for continuing investigations, taking into consideration that regulation of this splicing is not likely to be restricted to VEGF-A.
| Abbreviations: | ||
| AMD | = | age-related macular degeneration |
| DDS | = | Denys-Drash syndrome |
| DSS | = | distal splice site |
| IGF-1 | = | insulin-like growth factor 1 |
| MVECs | = | microvascular endothelial cells |
| NRP1 | = | neuropilin 1 |
| OIR | = | oxygen-induced retinopathy |
| PSS | = | proximal splice site |
| SRPK1 | = | serine/arginine protein kinase 1 |
| SRSF1 | = | serine/arginine-rich splicing factor 1 |
| SRSF2 | = | serine/arginine-rich splicing factor 2 |
| SRSF6 | = | serine/arginine-rich splicing factor 6 |
| SSc | = | systemic sclerosis |
| TGFβ | = | transforming growth factor beta |
| TNFα | = | tumor necrosis factor alpha |
| UTR | = | untranslated region |
| VEGF | = | vascular endothelial growth factor |
| VEGFR2 | = | vascular endothelial growth factor receptor 2 |
| WT1 | = | Wilms tumor 1 |
Acknowledgments
I apologize to all those colleagues whose work could not be discussed due to space limitations. I would also like to thank Dave Bates for his valuable comments on the manuscript.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Related Research Data
References
- Carmeliet P. Angiogenesis in health and disease. Nat Med 2003; 9:653 - 60; http://dx.doi.org/10.1038/nm0603-653; PMID: 12778163
- Ferrara N, Gerber HP, LeCouter J. The biology of VEGF and its receptors. Nat Med 2003; 9:669 - 76; http://dx.doi.org/10.1038/nm0603-669; PMID: 12778165
- Bates DO, Cui TG, Doughty JM, Winkler M, Sugiono M, Shields JD, et al. VEGF165b, an inhibitory splice variant of vascular endothelial growth factor, is down-regulated in renal cell carcinoma. Cancer Res 2002; 62:4123 - 31; PMID: 12124351
- Cébe Suarez S, Pieren M, Cariolato L, Arn S, Hoffmann U, Bogucki A, et al. A VEGF-A splice variant defective for heparan sulfate and neuropilin-1 binding shows attenuated signaling through VEGFR-2. Cell Mol Life Sci 2006; 63:2067 - 77; http://dx.doi.org/10.1007/s00018-006-6254-9; PMID: 16909199
- Neufeld G, Cohen T, Gengrinovitch S, Poltorak Z. Vascular endothelial growth factor (VEGF) and its receptors. FASEB J 1999; 13:9 - 22; PMID: 9872925
- Robinson CJ, Stringer SE. The splice variants of vascular endothelial growth factor (VEGF) and their receptors. J Cell Sci 2001; 114:853 - 65; PMID: 11181169
- Tischer E, Mitchell R, Hartman T, Silva M, Gospodarowicz D, Fiddes JC, et al. The human gene for vascular endothelial growth factor. Multiple protein forms are encoded through alternative exon splicing. J Biol Chem 1991; 266:11947 - 54; PMID: 1711045
- Woolard J, Wang WY, Bevan HS, Qiu Y, Morbidelli L, Pritchard-Jones RO, et al. VEGF165b, an inhibitory vascular endothelial growth factor splice variant: mechanism of action, in vivo effect on angiogenesis and endogenous protein expression. Cancer Res 2004; 64:7822 - 35; http://dx.doi.org/10.1158/0008-5472.CAN-04-0934; PMID: 15520188
- Perrin RM, Konopatskaya O, Qiu Y, Harper S, Bates DO, Churchill AJ. Diabetic retinopathy is associated with a switch in splicing from anti- to pro-angiogenic isoforms of vascular endothelial growth factor. Diabetologia 2005; 48:2422 - 7; http://dx.doi.org/10.1007/s00125-005-1951-8; PMID: 16193288
- Cébe-Suarez S, Grünewald FS, Jaussi R, Li X, Claesson-Welsh L, Spillmann D, et al. Orf virus VEGF-E NZ2 promotes paracellular NRP-1/VEGFR-2 coreceptor assembly via the peptide RPPR. FASEB J 2008; 22:3078 - 86; http://dx.doi.org/10.1096/fj.08-107219; PMID: 18467594
- Harper SJ, Bates DO. VEGF-A splicing: the key to anti-angiogenic therapeutics?. Nat Rev Cancer 2008; 8:880 - 7; http://dx.doi.org/10.1038/nrc2505; PMID: 18923433
- Bevan HS, van den Akker NM, Qiu Y, Polman JA, Foster RR, Yem J, et al. The alternatively spliced anti-angiogenic family of VEGF isoforms VEGFxxxb in human kidney development. Nephron, Physiol 2008; 110:57 - 67; http://dx.doi.org/10.1159/000177614
- Woolard J, Bevan HS, Harper SJ, Bates DO. Molecular diversity of VEGF-A as a regulator of its biological activity. Microcirculation 2009; 16:572 - 92; http://dx.doi.org/10.1080/10739680902997333; PMID: 19521900
- Ribeiro LA, Bacci ML, Seren E, Tamanini C, Forni M. Characterization and differential expression of vascular endothelial growth factor isoforms and receptors in swine corpus luteum throughout estrous cycle. Mol Reprod Dev 2007; 74:163 - 71; http://dx.doi.org/10.1002/mrd.20589; PMID: 16967516
- Xu J, Dou T, Liu C, Fu M, Huang Y, Gu S, et al. The evolution of alternative splicing exons in vascular endothelial growth factor A. Gene 2011; 487:143 - 50; http://dx.doi.org/10.1016/j.gene.2011.06.024; PMID: 21782909
- Artac RA, McFee RM, Smith RA, Baltes-Breitwisch MM, Clopton DT, Cupp AS. Neutralization of vascular endothelial growth factor antiangiogenic isoforms is more effective than treatment with proangiogenic isoforms in stimulating vascular development and follicle progression in the perinatal rat ovary. Biol Reprod 2009; 81:978 - 88; http://dx.doi.org/10.1095/biolreprod.109.078097; PMID: 19605786
- Ergorul C, Ray A, Huang W, Darland D, Luo ZK, Grosskreutz CL. Levels of vascular endothelial growth factor-A165b (VEGF-A165b) are elevated in experimental glaucoma. Mol Vis 2008; 14:1517 - 24; PMID: 18728749
- Kawamura H, Li X, Harper SJ, Bates DO, Claesson-Welsh L. Vascular endothelial growth factor (VEGF)-A165b is a weak in vitro agonist for VEGF receptor-2 due to lack of coreceptor binding and deficient regulation of kinase activity. Cancer Res 2008; 68:4683 - 92; http://dx.doi.org/10.1158/0008-5472.CAN-07-6577; PMID: 18559514
- Catena R, Larzabal L, Larrayoz M, Molina E, Hermida J, Agorreta J, et al. VEGF₁₂₁b and VEGF₁₆₅b are weakly angiogenic isoforms of VEGF-A. Mol Cancer 2010; 9:320; http://dx.doi.org/10.1186/1476-4598-9-320; PMID: 21194429
- Karni R, de Stanchina E, Lowe SW, Sinha R, Mu D, Krainer AR. The gene encoding the splicing factor SF2/ASF is a proto-oncogene. Nat Struct Mol Biol 2007; 14:185 - 93; http://dx.doi.org/10.1038/nsmb1209; PMID: 17310252
- Nowak DG, Amin EM, Rennel ES, Hoareau-Aveilla C, Gammons M, Damodoran G, et al. Regulation of vascular endothelial growth factor (VEGF) splicing from pro-angiogenic to anti-angiogenic isoforms: a novel therapeutic strategy for angiogenesis. J Biol Chem 2010; 285:5532 - 40; http://dx.doi.org/10.1074/jbc.M109.074930; PMID: 19906640
- Nowak DG, Woolard J, Amin EM, Konopatskaya O, Saleem MA, Churchill AJ, et al. Expression of pro- and anti-angiogenic isoforms of VEGF is differentially regulated by splicing and growth factors. J Cell Sci 2008; 121:3487 - 95; http://dx.doi.org/10.1242/jcs.016410; PMID: 18843117
- Manetti M, Guiducci S, Romano E, Ceccarelli C, Bellando-Randone S, Conforti ML, et al. Overexpression of VEGF165b, an inhibitory splice variant of vascular endothelial growth factor, leads to insufficient angiogenesis in patients with systemic sclerosis. Circ Res 2011; 109:e14 - 26; http://dx.doi.org/10.1161/CIRCRESAHA.111.242057; PMID: 21636803
- Merdzhanova G, Gout S, Keramidas M, Edmond V, Coll JL, Brambilla C, et al. The transcription factor E2F1 and the SR protein SC35 control the ratio of pro-angiogenic versus antiangiogenic isoforms of vascular endothelial growth factor-A to inhibit neovascularization in vivo. Oncogene 2010; 29:5392 - 403; http://dx.doi.org/10.1038/onc.2010.281; PMID: 20639906
- Díaz R, Peña C, Silva J, Lorenzo Y, García V, García JM, et al. p73 Isoforms affect VEGF, VEGF165b and PEDF expression in human colorectal tumors: VEGF165b downregulation as a marker of poor prognosis. Int J Cancer 2008; 123:1060 - 7; http://dx.doi.org/10.1002/ijc.23619; PMID: 18546269
- Bergers G, Benjamin LE. Tumorigenesis and the angiogenic switch. Nat Rev Cancer 2003; 3:401 - 10; http://dx.doi.org/10.1038/nrc1093; PMID: 12778130
- Pritchard-Jones RO, Dunn DB, Qiu Y, Varey AH, Orlando A, Rigby H, et al. Expression of VEGF(xxx)b, the inhibitory isoforms of VEGF, in malignant melanoma. Br J Cancer 2007; 97:223 - 30; http://dx.doi.org/10.1038/sj.bjc.6603839; PMID: 17595666
- Varey AH, Rennel ES, Qiu Y, Bevan HS, Perrin RM, Raffy S, et al. VEGF 165 b, an antiangiogenic VEGF-A isoform, binds and inhibits bevacizumab treatment in experimental colorectal carcinoma: balance of pro- and antiangiogenic VEGF-A isoforms has implications for therapy. Br J Cancer 2008; 98:1366 - 79; http://dx.doi.org/10.1038/sj.bjc.6604308; PMID: 18349829
- Peiris-Pagès M, Harper SJ, Bates DO, Ramani P. Balance of pro- versus anti-angiogenic splice isoforms of vascular endothelial growth factor as a regulator of neuroblastoma growth. J Pathol 2010; 222:138 - 47; http://dx.doi.org/10.1002/path.2746; PMID: 20662003
- Rennel ES, Hamdollah-Zadeh MA, Wheatley ER, Magnussen A, Schüler Y, Kelly SP, et al. Recombinant human VEGF165b protein is an effective anti-cancer agent in mice. Eur J Cancer 2008; 44:1883 - 94; http://dx.doi.org/10.1016/j.ejca.2008.05.027; PMID: 18657413
- Rennel E, Waine E, Guan H, Schüler Y, Leenders W, Woolard J, et al. The endogenous anti-angiogenic VEGF isoform, VEGF165b inhibits human tumour growth in mice. Br J Cancer 2008; 98:1250 - 7; http://dx.doi.org/10.1038/sj.bjc.6604309; PMID: 18349828
- Amin EM, Oltean S, Hua J, Gammons MV, Hamdollah-Zadeh M, Welsh GI, et al. WT1 mutants reveal SRPK1 to be a downstream angiogenesis target by altering VEGF splicing. Cancer Cell 2011; 20:768 - 80; http://dx.doi.org/10.1016/j.ccr.2011.10.016; PMID: 22172722
- Zhao M, Shi X, Liang J, Miao Y, Xie W, Zhang Y, et al. Expression of pro- and anti-angiogenic isoforms of VEGF in the mouse model of oxygen-induced retinopathy. Exp Eye Res 2011; 93:921 - 6; http://dx.doi.org/10.1016/j.exer.2011.10.013; PMID: 22067127
- Konopatskaya O, Churchill AJ, Harper SJ, Bates DO, Gardiner TA. VEGF165b, an endogenous C-terminal splice variant of VEGF, inhibits retinal neovascularization in mice. Mol Vis 2006; 12:626 - 32; PMID: 16735996
- Magnussen AL, Rennel ES, Hua J, Bevan HS, Beazley Long N, Lehrling C, et al. VEGF-A165b is cytoprotective and antiangiogenic in the retina. Invest Ophthalmol Vis Sci 2010; 51:4273 - 81; http://dx.doi.org/10.1167/iovs.09-4296; PMID: 20237249
- Ehlken C, Rennel ES, Michels D, Grundel B, Pielen A, Junker B, et al. Levels of VEGF but not VEGF(165b) are increased in the vitreous of patients with retinal vein occlusion. Am J Ophthalmol 2011; 152:298 - 303, e1; http://dx.doi.org/10.1016/j.ajo.2011.01.040; PMID: 21621189
- Hua J, Spee C, Kase S, Rennel ES, Magnussen AL, Qiu Y, et al. Recombinant human VEGF165b inhibits experimental choroidal neovascularization. Invest Ophthalmol Vis Sci 2010; 51:4282 - 8; http://dx.doi.org/10.1167/iovs.09-4360; PMID: 20237252
- Ricker LJ, Kijlstra A, Kessels AG, de Jager W, Liem AT, Hendrikse F, et al. Interleukin and growth factor levels in subretinal fluid in rhegmatogenous retinal detachment: a case-control study. PLoS One 2011; 6:e19141; http://dx.doi.org/10.1371/journal.pone.0019141; PMID: 21556354
- Eremina V, Sood M, Haigh J, Nagy A, Lajoie G, Ferrara N, et al. Glomerular-specific alterations of VEGF-A expression lead to distinct congenital and acquired renal diseases. J Clin Invest 2003; 111:707 - 16; PMID: 12618525
- Ferguson J, Qiu Y, Sage LM, Neal CR, Bates DO, Harper SJ, et al. Ultrafiltration co-efficient in isolated intact glomeruli from podocyte-specific VEGF165b over-expressing transgenic mice. Microcirculation 14, 643 (Abstract) 2007.
- Qiu Y, Ferguson J, Oltean S, Neal CR, Kaura A, Bevan H, et al. Overexpression of VEGF165b in podocytes reduces glomerular permeability. J Am Soc Nephrol 2010; 21:1498 - 509; http://dx.doi.org/10.1681/ASN.2009060617; PMID: 20688932
- Oltean S, Neal CR, Mavrou A, Patel P, Ahad T, Alsop C, et al. VEGF165b overexpression restores normal glomerular water permeability in VEGF164-overexpressing adult mice. Am J Physiol Renal Physiol 2012; 303:F1026 - 36; http://dx.doi.org/10.1152/ajprenal.00410.2011; PMID: 22811490
- Schumacher VA, Jeruschke S, Eitner F, Becker JU, Pitschke G, Ince Y, et al. Impaired glomerular maturation and lack of VEGF165b in Denys-Drash syndrome. J Am Soc Nephrol 2007; 18:719 - 29; http://dx.doi.org/10.1681/ASN.2006020124; PMID: 17267748
- Reddy JC, Morris JC, Wang J, English MA, Haber DA, Shi Y, et al. WT1-mediated transcriptional activation is inhibited by dominant negative mutant proteins. J Biol Chem 1995; 270:10878 - 84; http://dx.doi.org/10.1074/jbc.270.18.10878; PMID: 7738027
- Qiu Y, Bevan H, Weeraperuma S, Wratting D, Murphy D, Neal CR, et al. Mammary alveolar development during lactation is inhibited by the endogenous antiangiogenic growth factor isoform, VEGF165b. FASEB J 2008; 22:1104 - 12; http://dx.doi.org/10.1096/fj.07-9718com; PMID: 18032632
- Bates DO, MacMillan PP, Manjaly JG, Qiu Y, Hudson SJ, Bevan HS, et al. The endogenous anti-angiogenic family of splice variants of VEGF, VEGFxxxb, are down-regulated in pre-eclamptic placentae at term. Clin Sci (Lond) 2006; 110:575 - 85; http://dx.doi.org/10.1042/CS20050292; PMID: 16451124
- Bills VL, Salmon AH, Harper SJ, Overton TG, Neal CR, Jeffery B, et al. Impaired vascular permeability regulation caused by the VEGF₁₆₅b splice variant in pre-eclampsia. BJOG 2011; 118:1253 - 61; http://dx.doi.org/10.1111/j.1471-0528.2011.02925.x; PMID: 21668772
- Bills VL, Varet J, Millar A, Harper SJ, Soothill PW, Bates DO. Failure to up-regulate VEGF165b in maternal plasma is a first trimester predictive marker for pre-eclampsia. Clin Sci (Lond) 2009; 116:265 - 72; http://dx.doi.org/10.1042/CS20080270; PMID: 18826376
- LeRoy EC. Systemic sclerosis. A vascular perspective. Rheum Dis Clin North Am 1996; 22:675 - 94; http://dx.doi.org/10.1016/S0889-857X(05)70295-7; PMID: 8923590
- Manetti M, Guiducci S, Ibba-Manneschi L, Matucci-Cerinic M. Mechanisms in the loss of capillaries in systemic sclerosis: angiogenesis versus vasculogenesis. J Cell Mol Med 2010; 14:6A 1241 - 54; http://dx.doi.org/10.1111/j.1582-4934.2010.01027.x; PMID: 20132409
- Simpson A, Custovic A, Tepper R, Graves P, Stern DA, Jones M, et al. Genetic variation in vascular endothelial growth factor-a and lung function. Am J Respir Crit Care Med 2012; 185:1197 - 204; http://dx.doi.org/10.1164/rccm.201112-2191OC; PMID: 22461367
- Harris S, Craze M, Newton J, Fisher M, Shima DT, Tozer GM, et al. Do anti-angiogenic VEGF (VEGFxxxb) isoforms exist? A cautionary tale. PLoS One 2012; 7:e35231; http://dx.doi.org/10.1371/journal.pone.0035231; PMID: 22567098
- Qiu Y, Hoareau-Aveilla C, Oltean S, Harper SJ, Bates DO. The anti-angiogenic isoforms of VEGF in health and disease. Biochem Soc Trans 2009; 37:1207 - 13; http://dx.doi.org/10.1042/BST0371207; PMID: 19909248
- Baba T, McLeod DS, Edwards MM, Merges C, Sen T, Sinha D, et al. VEGF 165 b in the developing vasculatures of the fetal human eye. Dev Dyn 2012; 241:595 - 607; http://dx.doi.org/10.1002/dvdy.23743; PMID: 22275161
- Lipinski DM, Singh MS, MacLaren RE. Assessment of cone survival in response to CNTF, GDNF, and VEGF165b in a novel ex vivo model of end-stage retinitis pigmentosa. Invest Ophthalmol Vis Sci 2011; 52:7340 - 6; http://dx.doi.org/10.1167/iovs.11-7996; PMID: 21873685
- Glass CA, Harper SJ, Bates DO. The anti-angiogenic VEGF isoform VEGF165b transiently increases hydraulic conductivity, probably through VEGF receptor 1 in vivo. J Physiol 2006; 572:243 - 57; PMID: 16423853
- Bates DO, Hillman NJ, Williams B, Neal CR, Pocock TM. Regulation of microvascular permeability by vascular endothelial growth factors. J Anat 2002; 200:581 - 97; http://dx.doi.org/10.1046/j.1469-7580.2002.00066.x; PMID: 12162726
- Baltes-Breitwisch MM, Artac RA, Bott RC, McFee RM, Kerl JG, Clopton DT, et al. Neutralization of vascular endothelial growth factor antiangiogenic isoforms or administration of proangiogenic isoforms stimulates vascular development in the rat testis. Reproduction 2010; 140:319 - 29; http://dx.doi.org/10.1530/REP-09-0456; PMID: 20457593