
Abstract
Radiotherapy is one of the most effective therapeutic strategies for breast cancer patients, although its efficacy may be reduced by intrinsic radiation resistance of cancer cells. Recent investigations demonstrate a link between cancer cell radio-resistance and activation of sphingosine kinase (SphK1), which plays a key role in the balance of lipid signaling molecules. Sphingosine kinase (SphK1) activity can alter the sphingosine-1-phosphate (S1P)/ceramide ratio leading to an imbalance in the sphingolipid rheostat. Fingolimod (FTY720) is a novel sphingosine analog and a potent immunosuppressive drug that acts as a SphK1 antagonist, inhibits the growth, and induces apoptosis in different human cancer cell lines. We sought to investigate the in vitro radiosensitizing effects of FTY720 on the MDA-MB-361 breast cancer cell line and to assess the effects elicited by radiation and FTY720 combined treatments. We found that FTY720 significantly increased anti-proliferative and pro-apoptotic effects induced by a single dose of ionizing radiation while causing autophagosome accumulation. At the molecular level, FTY720 significantly potentiated radiation effects on perturbation of signaling pathways involved in regulation of cell cycle and apoptosis, such as PI3K/AKT and MAPK. In conclusion, our data highlight a potent radiosensitizing effect of FTY720 on breast cancer cells and provide the basis of novel therapeutic strategies for breast cancer treatment.
Introduction
Breast cancer is the most common female cancer and the leading cause of cancer deaths among women in the economically developed countries.Citation1-Citation3 Treatment of breast cancer requires the use of multiple therapeutic modalities including radiation therapy. Resistance to radiotherapy often results in treatment failure, particularly for loco-regional recurrence.
Hence, new therapeutic strategies are required for overcoming radioresistance and improving prognosis of breast cancer patients.Citation4
So far, multiple alterations of signal transduction pathways have been correlated with resistance of cancer cells against conventional anticancer therapies.Citation5 Among them, the aberrant sphingolipid metabolism is an important mediator of both chemo- and radio-therapy resistance.Citation6 The enzyme sphingosine kinase (SphK1) converts the pro-apoptotic, anti-proliferative ceramide into its pro-survival and mitogenic metabolite S1P (sphingosine 1-phosphatase);Citation7,Citation8 high cellular levels of SphK1 can protect against ionizing radiation. Preclinical evidence suggests that pharmacological inhibition of SphK1 results in sensitization of human prostate cancer cells to radiotherapy.Citation9
FTY720 is a sphingosine analog and a potent immunosuppressive drug currently approved for treating multiple sclerosis.Citation10,Citation11 Moreover, FTY720 has also been shown to induce apoptosis in multiple myeloma,Citation12 prostate,Citation13 breast,Citation14 kidney,Citation15 and bladder cancer cells.Citation16
Based on this experimental evidence, we sought to analyze the radiosensitizing effects of FTY720 in human breast cancer cells, in order to clearly assess its effect on growth, proliferation, and apoptosis.
Results
Effects of FTY720 and ionizing radiations on MDA-MB-361 breast cancer cells
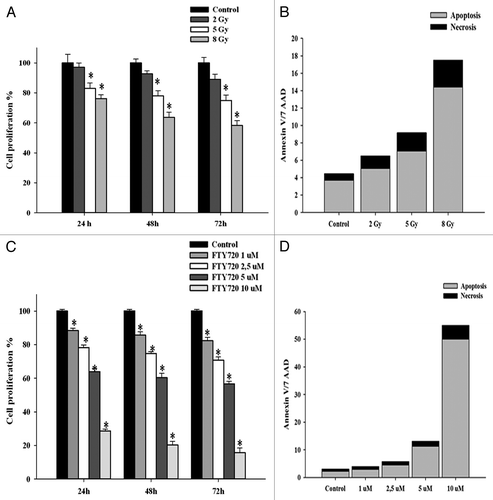
Trypan blue exclusion assay was used to determine the effects of FTY720, either alone or in combination with radiation, on cell proliferation in vitro. Ionizing radiation caused a dose-dependent inhibition of cell proliferation: the greatest effects were obtained by a single 8 Gy administration that induced a decrease in cancer cell proliferation of 24% at 24 h, 37% at 48 h, and 42% at 72 h (). These effects were paralleled by a dose-dependent induction of apoptotic cell death after treatment with ionizing radiations (). Moreover, we evaluated the effects of FTY720 on proliferation of MDA-MB361 cancer cells: a dose-dependent inhibition of cell proliferation was indeed observed after treatment with the drug (), which occurred together to apoptosis induction ().
Figure 1. Antitumor effect of FTY720 and radiation in MDA-MB 361 cells. (A) Cells were plated overnight and then exposed to radiation up to 8 Gy as indicated. Viability of cells was assessed by trypan blue assay. (B) Number of apoptotic cells was quantified by flow cytometry staining with Annexin V/7AAD after 24 h of incubation. (C) Cell viability of MDA-MB-361 treated with FTY720. Plated cells were treated with FTY720 at varying doses for the indicated times. (D) Number of apoptotic cells after drug administration at 24 h. Data are the average of three independent experiments performed in triplicate. P values were obtained using two-tailed t test. *P < 0.01.
FTY720 enhances radiation-induced growth inhibition and apoptosis
Next we assessed whether FTY720 treatment could enhance radiation effects on MDA-MB 361 cells. The suboptimal dose of 5 μM FTY720 was chosen for combinatory studies since lower concentrations resulted ineffective. Notably, FTY720 strongly increased the anti-proliferative effects of 8 Gy radiation (the greatest effects were obtained at 72 h: % of growth inhibition was of 42% for radiation, 42% for FTY720, and 90% for combination treatment []). Simultaneous exposure to FTY720 synergistically interacted with ionizing radiation. The combination of FTY720 and ionizing radiation displayed a strong synergistic effect against mammalian cancer cells (CI = 1.1).
Figure 2. FTY720 sensitizes MDA-MB 361 cells to X-irradiation. Plated cells were pretreated for 1 h with FTY720 or vehicle at indicated concentration (5 uM), then exposed to 8 Gy X-irradiation. (A) Cell viability of MDA-MB 361 cells was measured by trypan blue at indicated time. (B) Apoptotic MDA-MB 361 cells after 24 h of incubation were assessed by flow cytometry after Annexin V/PI staining. Columns and points, mean of three independent experiment performed in triplicate; bars, *P < 0.01; (C–F) Observed nuclei morphological changes in MDA MB 361 cells after staining with DAPI using fluorescent microscopy with 40× magnification. (C) Untreated cells control; (D) treated cells with 5 uM FTY720; (E) treated cells with 8 Gy radiation; (F) treated cells with combination of FTY720 plus radiation.
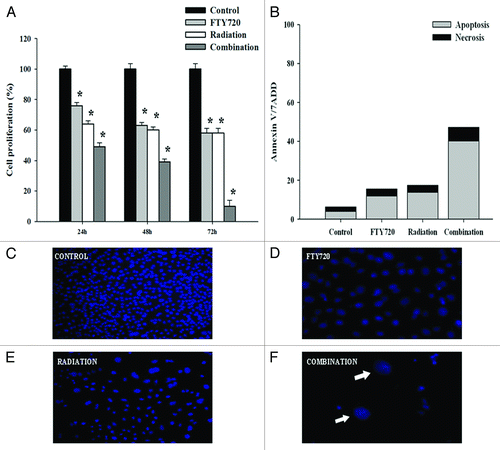
These results suggest that FTY720 potentiates the growth-inhibitory effects of radiation in breast cancer cells. To establish if the growth-inhibitory effects were dependent on apoptosis, we evaluated the proportion of apoptotic cells after treatment with FTY720 and radiation, using Annexin V/7-AAD staining and flow cytometry analysis. Results indicated that FTY720 combined with radiation induced 40% of apoptosis and 7% of necrosis at 24 h in MDA-MB 361, which was significantly higher than radiation alone (17% in MDA-MB 361) or FTY720 alone (13% in MDA-361) (CI = 2). Hence these data indicate that apoptotic induction plays a significant role in FTY720-mediated radiosensitization of human breast cancer cells (). Moreover, apoptosis induction in MDA-MB 361 cancer cells was assessed by the DAPI staining assay. Combination treatment led to shrinkage of nuclei, as shown in , further underlying the induction of apoptotic cell death.
Combination of FTY720 with RT induces MDA-MB 361 cell accumulus in G0/1 phase
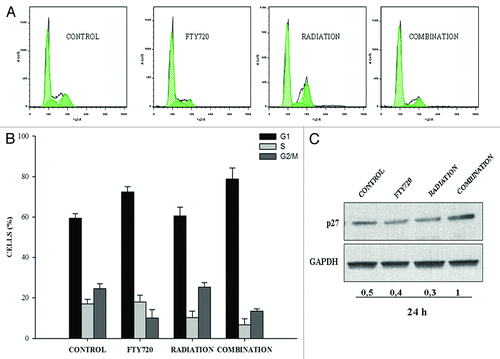
To assess whether FTY720 enhancement of radiation-induced growth inhibition was also dependent on cell cycle alterations, we analyzed the distribution of cell cycle phases by flow cytometry. Interestingly, the two agents mediated different effects on cell cycle, since 24 h exposure to FTY720 caused a significant increase of cells at G0/G1 phase and a decrease of cells at S and G2/M phase, whereas radiation slightly reduced S phase. FTY720 in combination with radiation enhanced blockage at G0/G1 cell cycle phase (). Consistently with this finding, an increase in the expression of the cell cycle inhibitor p27Kip1 was observed after combined treatment but was not affected by the individual treatments ().
Figure 3. Effects of FTY720 or ionizing radiation as individual agents or in combination on cell cycle distribution. (A) Perturbations of the cell cycle, analyzed by flow cytometry, after 24 h exposure to single or combined treatments. The data shown are representative of the combined mean of two independent experiments. (B) p27 immunoblot, at 24 h after treatment. Standardization was performed with GAPDH stained in the same blots with anti-GAPDH antibody.
FTY720 inhibits radiation-dependent activation of prosurvival factors and potentiates RT-dependent caspase 3/7 activation
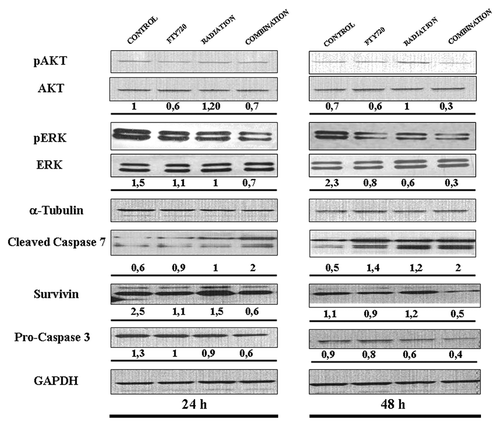
To investigate the molecular mechanisms underlying the radio-sensitizing effects of FTY720, we analyzed the role of FTY720 plus radiation on the activation of signal transduction pathways involved in regulation of survival and apoptosis. Cells were treated with radiation (8 Gy) and FTY720 (5 μM) for 24, 48, and 72 h and then the expression of phospho-ERK1/2, a member of the MAPK family, and phospho-AKT were assessed by western blot. Combination treatments reduced the expression of phospho-ERK1/2 compared with either FTY720 or radiation alone. In our experimental setting, FTY720 in combination with radiation strongly decreased the levels of phospho-AKT especially if compared with radiation alone at 48 h time points. Furthermore, we examined the expression of the anti-apoptotic protein survivin and observed a decline in its expression 48 h following combination treatment.
We evaluated caspase 3 and caspase 7 activity: a marked time-dependent increase in the cleavage of caspase-7 was observed in cells treated with FTY720 (5 μM) in combination with radiation (8 Gy), 24 and 48 h after treatment. Moreover, pro-caspase 3 expression was decreased by the combinatory treatment with a stronger effect after 48 h of treatment ().
Figure 4. Combination treatment regulates the apoptotic and survival pathways. Immunoblot of pAKT/AKT and pERK/ERK, 24 and 48 h after individual or combination treatments. Standardization was performed with α-tubuline measured in the same blots with anti-α-tubuline antibody. Moreover, immunoblot of cleaved caspase 7, pro-caspase 3, and survivin at time and conditions indicated are described. The protein loading control was performed using GAPDH.
Role of autophagy in FTY720/radiation-induced cell death
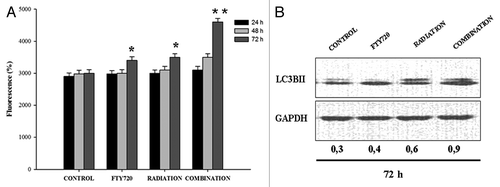
To assess if FTY720 could also modulate radiation effects on autophagy, we analyzed the regulation of autophagy after staining with mono-dansyl-cadaverine (MDC), a probe for detection of late autophagic vacuoles that is incorporated into multilamellar bodies by both an ion trapping mechanism and interaction with membrane lipids.Citation17 FTY720 (5 μM) plus ionizing radiation (8 Gy) treatment increased MDC fluorescence intensity, indicating that combination caused an accumulation of autophagosome likely due to the block of the autophagic flux determined by the combined treatment in MDA-MB361 cells (). Moreover, we analyzed conversion of microtubule associated protein 1 light chain 3 (LC3)-I to LC3-II, that is a marker of autophagosome degradation, by western blotting with anti-LC3 antibody. FTY720 alone could induce this effect, but increased conversion from LC3-I to LC3-II was demonstrated after combined treatment with radiation (). This finding suggests that the combined treatment with FTY720 and radiation likely blocks autophagic processes switching the programmed cell death toward apoptosis.
Figure 5. Combination treatment induces autophagosome accumulation in MDA-MB-361 cells. (A) To quantify fluorescent compound MDC incorporated in MDA-MB-361, cells autophagy test was used as described in Methods. *P > 0.05; **P = 0.02. (B) Western blot analysis was performed for LC3B. The equal loading of protein was confirmed by probing with GAPDH.
Discussion
Radiation therapy is now of routine value after conservative surgery to reduce loco-regional tumor recurrence in breast cancer.Citation18 Unfortunately, the efficacy of radiotherapy is often limited by the intrinsic cell resistance to ionizing radiation of tumor cells.Citation19
In the last decade, the development of radiosensitizers have led to increased efficacy and reduced toxicity of radiation therapy. S1P is a potent sphingolipid mediator of diverse processes important for tumor cells, including cell growth, survival, migration, invasion, and angiogenesis. SphK1 is upregulated in various human tumor tissue.Citation20 The SphK1/S1P pathway contributes to cancer progression and leads to increased cell proliferation, impairment of apoptosis, and oncogenic transformation.Citation21 Furthermore, high levels of SphK1 expression and activity are associated with a poor prognosis in breast cancer.Citation22 This information indicates that high SphK1 activity can protect against ionizing radiation and could induce radioresistance. Many therapeutic agents act against cancer cells by inducing apoptosis and resistance to cancer therapies, which usually involves intrinsic or extrinsic abrogation of the apoptotic machinery. Accumulating evidence has indicated that SphK1 could protect cancer cells from apoptosis;Citation23 in addition, breast cancer cell radioresistance has been linked with sustained activation of SphK1.Citation10 Therefore, a combination therapy based on a SphK1 inhibitor and radiation could provide a useful strategy for the treatment of therapeutic-resistant cancers.Citation24
Here we report that targeting SphK1 with FTY720 potentiated radiation-induced suppression of cell proliferation and induction of apoptosis. Our findings are in agreement with a recent work by Pchekectski et al.Citation9 showing that FTY720 acts as a radiosensitizer on prostate cancer cell lines. It is also well accepted that S1P produced by activation of SphK1 is released from cells and stimulates its receptors that are linked to activation of AKT.Citation25 The PI3/AKT pathway is implicated in radioresistance of human tumor cells. Inhibition of the AKT pathway can modulate sensitivity of tumor cells to apoptosis to increase radiation-induced cell death.Citation26,Citation27 The reduction of S1P levels by FTY720 contributes to decreased phosphorylation of AKT in combined treatment with radiation, as we have shown by western blot analysis. Moreover, the levels of the anti-apoptotic protein survivin, which is increased in most malignancies and confers radiation resistance in cancer,Citation28,Citation29 was strongly decreased in cells treated with FTY720 plus radiation. It is therefore tempting to speculate that FTY720/radiation induced-apoptosis might be dependent on downregulation of survivin. Moreover, in the present study we report, together with the induction of apoptosis, the increased accumulation of autophagosome vesicles in cells treated with FTY720/radiation. It is well established that apoptosis is important in determining the outcome of chemo- and radiation therapy and can be triggered by anticancer drugs and radiation. However, apoptosis is not the unique form of cell death. Autophagy is an evolutionarily conserved catabolic process for the degradation and recycling of cytosolic, long lived, or aggregated proteins, and excess or defective organelles, and is primarily a response to the stress by radiation and chemotherapy agents. Recently, different studies have suggested a dual role for autophagy in regulating cell death and it has recently emerged as a survival response of cancer cells to chemotherapy agents.Citation30 In this light, it has been reported that the autophagy induced by FTY720 worked as a protective function in ovarian cancerCitation31 and acute lymphoblastic leukemia cells;Citation32 on the other hand, a recent study on multiple myeloma cells has demonstrated the ability of FTY720 to induce autophagic cell death and apoptosis.Citation33,Citation34 This discrepancy may be attributable to the complex and diverse interplays between autophagy and other mechanisms of cell death.Citation35 In our experimental setting, we found that the decrease in cell viability and the induction of apoptosis occurred together to the increase of autophagosome vesicles, as assessed by upregulation of apoptotic, i.e., cleaved caspase 7, and autophagic markers, i.e., cleaved LC3B. The regulatory pathways of autophagy and apoptosis share several molecules, among these PI3K/AKT/mTOR plays an important role in radiation-induced autophagy.Citation36,Citation37 SIP is a potent inhibitor of ceramide-induced apoptosis but also induces autophagy, which indicates how the intracellular milieu can contribute to “deciding” between the two processes.
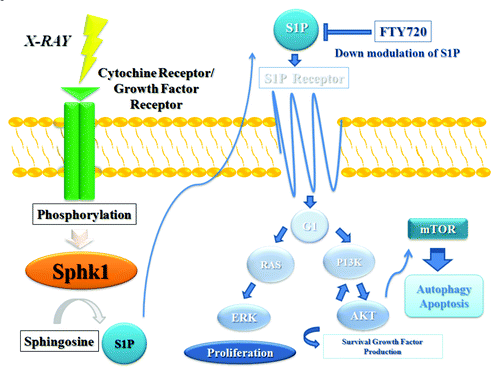
An explanatory cartoon of the effects of fingolimod on signal transduction pathways in our experimental model is reported in .
Figure 6. Explanatory cartoon of fingolimod effects on signal transduction pathways. Sphingosine kinase (SphK1) is a key regulator of the dynamic ceramide/sphingosine-1-phosphate (SP1) rheostat balance which is important in the pathological cancerogenesis, progression, metastasis process and resistance to treatments. FTY720 acts as a SphK1 antagonist reducing SP1 levels with perturbation of PI3K/AKT/mTor pathway involved in apoptosis and autophagy of the cancer cells and modulating sensitivity of tumor cells to radiation-induced cell death.
On the basis of our results, it is possible to hypothesize that the combined treatment potentiates apoptosis occurrence and blocks the autophagy flux causing a paradoxical increase of autophagosomes in breast cancer cells that decide to die through apoptosis. On the other hand, autophagy can facilitate apoptosis by maintaining ATP levels during starvation to promote ATP-dependent apoptotic processes in embryonic cells.Citation38
In conclusion, our findings indicate that FTY720 induces radiosensitization of breast cancer cells, providing the first preclinical framework for a potential clinical use of this agent, in combination with radiotherapy, to overcome radioresistance and potentially improve the outcome of breast cancer patients.
Materials and Methods
Cell lines and drug
Human breast cancer cells (MDA-MB-361) were obtained from the American Type Culture Collection and maintained as adherent monolayer cultures previously reported.Citation39
FTY720(Amino-2-[2-(4-octylphenyl)ethyl]propane-1,3-diol hydrochloride) was purchased from Selleck Chemicals, and stored as 100 mM stock solution in DMSO, protected from light at −20 °C. The drug was diluted in fresh media prior to each experiment.
Radiation treatment
Exponentially growing tumor cells were plated in 100 mm tissue culture dishes or in 6-well plates, allowed to attach for 24 h, and treated with 3 different doses of radiation (2, 5, and 8 Gy) at room temperature, at the dose of 1, 8 Gy/min and Source Surface Distance (SSD) of 80 cm2, by using a 6 MV photon linear accelerator (CLINAC 600 Varian) as previously reported.Citation40
Trypan blue viability assay
Cell lines were plated on 6-well plates at the density of 5.0 × 104/well. Twenty-four hours later, cells were left untreated, irradiated with different doses of radiation or treated with various concentrations of FTY720; for combination treatments, FTY720 was added 1 h before irradiation. After 24, 48, and 72 h incubation, cell survival was determined with 0.2% trypan blue solution.
Analysis of apoptosis
Cells were treated as previously described with ionizing radiation, FTY720, or combinations; after 24 h, cells were harvested, washed twice with cold 1× PBS and stained with Annexin V-PE/7-aminoactinomycin D (7AAD) solution (BD Bioscience) as indicated by the manufacturerCitation41 and then analyzed by flow cytometery (FACS-Calibur, BD Bioscience).
Apoptosis induction was also evaluated by fluorescence dye staining using DAPI to identify the condensation and fragmentation of nucleic DNA of apoptotic cells.
Cancer cells were treated with the combination of FTY720 and ionizing radiation (5 μM and 8 Gy respectively) and incubated for 72 h. The culture medium was removed and cells were rinsed by 1× PBS and finally fixed using 50 μL methanol. Subsequently 50 μL of DAPI (Invitrogen, Life Technologies) was added to cells at the final concentration of 1 μg/mL. The stained nucleic DNA was captured under fluorescence microscopy at a magnitude of 40× (LEICA DM4000 B LED, Leica Microsystems).
Combination index
Interaction between FTY720 and ionizing radiation was assessed using the combination index (CI), where CI > 1, CI = 1, and CI < 1 indicated synergistic, additive, and antagonistic effects, respectively. Data analysis was done by the following formula: CI = (inib A + B) / (inib A) + (inib B) as previously reported.Citation42
Cell cycle analysis
Cancer cells (1 × 106 mL) were seeded in 100-mm tissue culture dishes and allowed to attach for 24 h. After treatments, cells were trypsinized, washed, fixed with 70% ethanol and stored overnight at –20 °C, followed by washing and staining with a solution containing propidium iodide, RNase A, and NP40, according to previous published work;Citation43 cell cycle distribution was determined by flow cytometry.
Antibodies and western blot
Total protein extracts from untreated cells or from cells treated with FTY720 alone were lysed with NP40 buffer supplemented with protease inhibitor cocktails (Roche) and phosphatase inhibitor cocktails (Sigma Aldrich), and separated by Nu-PAGE gel electrophoresis as previously reported.Citation44 Antibodies specific for total Akt, phospho-Akt (Ser473) and for caspase3/7, ERK1/2, phospho-ERK1/2 (Thr202/Tyr204), LC3BII were obtained from Cell Signaling Technologies.
Antibodies specific for survivin, α-Tubulin and GAPDH were from Santa Cruz Biotechnology.
Immunofluorescence and light microscopy
Cells (2.0 × 105) were seeded into glass coverslips and left attach for 24 h, then were treated as above described. Seventy two hours after incubation, cells were washed with 1× PBS and fixed with 3% paraformaldehyde, 0.2% Triton X-100 in PBS for 20 min. Cells were stained with DAPI and examined by light or fluorescence microscopy (Leica Microsystems).
Autophagy assessment
MDA-MB-361 cells were seeded in 96 well plates at a density of 5.0 × 104 cells/well and incubated over-night at 37 °C. Twenty-four hours later, cells were treated as described earlier and incubated over-night. On the third day, cells were stained with the vital dye mono-dansyl-cadaverine (Autophagy/Cytotoxicity dual staining kit, Caymann) and fluorescence was quantified using a plate reader.
Statistical analysis
All the tests were done in triplicate and experiments performed at least three times. The results were expressed as a mean ± standard deviation. Differences between groups were analyzed by Student's two-tailed t test used to calculate all reported P values using GraphPad software (www.graphpad.com).
Asterisks denote statistical significance as indicated in the legends.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
References
- Ferlay J, Shin HR, Bray F, Forman D, Mathers C, Parkin DM. Estimates of worldwide burden of cancer in 2008: GLOBOCAN 2008. Int J Cancer 2010; 127:2893 - 917; http://dx.doi.org/10.1002/ijc.25516; PMID: 21351269
- Jemal A, Siegel R, Xu J, Ward E. Cancer statistics, 2010. CA Cancer J Clin 2010; 60:277 - 300; http://dx.doi.org/10.3322/caac.20073; PMID: 20610543
- Kols A. Breast cancer: increasing incidence. Limited Options. Outlook 2002; 19:1 - 8
- Jung S, Lee S, Lee J, Li C, Ohk JY, Jeong HK, Lee S, Kim S, Choi Y, Kim S, et al. Protein expression pattern in response to ionizing radiation in MCF-7 human breast cancer cells. Oncol Lett 2012; 3:147 - 54; PMID: 22740871
- Tassone P, Tagliaferri P, Perricelli A, Blotta S, Quaresima B, Martelli ML, Goel A, Barbieri V, Costanzo F, Boland CR, et al. BRCA1 expression modulates chemosensitivity of BRCA1-defective HCC1937 human breast cancer cells. Br J Cancer 2003; 88:1285 - 91; http://dx.doi.org/10.1038/sj.bjc.6600859; PMID: 12698198
- Baran Y, Salas A, Senkal CE, Gunduz U, Bielawski J, Obeid LM, Ogretmen B. Alterations of ceramide/sphingosine 1-phosphate rheostat involved in the regulation of resistance to imatinib-induced apoptosis in K562 human chronic myeloid leukemia cells. J Biol Chem 2007; 282:10922 - 34; http://dx.doi.org/10.1074/jbc.M610157200; PMID: 17303574
- Maceyka M, Payne SG, Milstien S, Spiegel S. Sphingosine kinase, sphingosine-1-phosphate, and apoptosis. Biochim Biophys Acta 2002; 1585:193 - 201; http://dx.doi.org/10.1016/S1388-1981(02)00341-4; PMID: 12531554
- Taha TA, Kitatani K, El-Alwani M, Bielawski J, Hannun YA, Obeid LM. Loss of sphingosine kinase-1 activates the intrinsic pathway of programmed cell death: modulation of sphingolipid levels and the induction of apoptosis. FASEB J 2006; 20:482 - 4; PMID: 16507765
- Pchejetski D, Golzio M, Bonhoure E, Calvet C, Doumerc N, Garcia V, Mazerolles C, Rischmann P, Teissié J, Malavaud B, et al. Sphingosine kinase-1 as a chemotherapy sensor in prostate adenocarcinoma cell and mouse models. Cancer Res 2005; 65:11667 - 75; http://dx.doi.org/10.1158/0008-5472.CAN-05-2702; PMID: 16357178
- Chiba K. FTY720, a new class of immunomodulator, inhibits lymphocyte egress from secondary lymphoid tissues and thymus by agonistic activity at sphingosine 1-phosphate receptors. Pharmacol Ther 2005; 108:308 - 19; http://dx.doi.org/10.1016/j.pharmthera.2005.05.002; PMID: 15951022
- Kappos L, Antel J, Comi G, Montalban X, O’Connor P, Polman CH, Haas T, Korn AA, Karlsson G, Radue EW, FTY720 D2201 Study Group. Oral fingolimod (FTY720) for relapsing multiple sclerosis. N Engl J Med 2006; 355:1124 - 40; http://dx.doi.org/10.1056/NEJMoa052643; PMID: 16971719
- Yasui H, Hideshima T, Raje N, Roccaro AM, Shiraishi N, Kumar S, Hamasaki M, Ishitsuka K, Tai YT, Podar K, et al. FTY720 induces apoptosis in multiple myeloma cells and overcomes drug resistance. Cancer Res 2005; 65:7478 - 84; http://dx.doi.org/10.1158/0008-5472.CAN-05-0850; PMID: 16103102
- Wang JD, Takahara S, Nonomura N, Ichimaru N, Toki K, Azuma H, Matsumiya K, Okuyama A, Suzuki S. Early induction of apoptosis in androgen-independent prostate cancer cell line by FTY720 requires caspase-3 activation. Prostate 1999; 40:50 - 5; http://dx.doi.org/10.1002/(SICI)1097-0045(19990615)40:1<50::AID-PROS6>3.0.CO;2-N; PMID: 10344723
- Azuma H, Horie S, Muto S, Otsuki Y, Matsumoto K, Morimoto J, Gotoh R, Okuyama A, Suzuki S, Katsuoka Y, et al. Selective cancer cell apoptosis induced by FTY720; evidence for a Bcl-dependent pathway and impairment in ERK activity. Anticancer Res 2003; 23:3183 - 93; PMID: 12926052
- Ubai T, Azuma H, Kotake Y, Inamoto T, Takahara K, Ito Y, Kiyama S, Sakamoto T, Horie S, Muto S, et al. FTY720 induced Bcl-associated and Fas-independent apoptosis in human renal cancer cells in vitro and significantly reduced in vivo tumor growth in mouse xenograft. Anticancer Res 2007; 27:1A 75 - 88; PMID: 17352219
- Tanaka T, Takahara S, Hatori M, Suzuki K, Wang J, Ichimaru N, Suzuki S, Morozumi K, Okuyama A, Yamanaka H. A novel immunosuppressive drug, FTY720, prevents the cancer progression induced by cyclosporine. Cancer Lett 2002; 181:165 - 71; http://dx.doi.org/10.1016/S0304-3835(01)00799-6; PMID: 12175531
- Jin S, White E. Tumor suppression by autophagy through the management of metabolic stress. Autophagy 2008; 4:563 - 6; PMID: 18326941
- Cutuli B. [Radiotherapy for breast cancer: which strategy in 2012?]. Cancer Radiother 2012; 16:493 - 502; http://dx.doi.org/10.1016/j.canrad.2012.07.185; PMID: 22925489
- Begg AC, Stewart FA, Vens C. Strategies to improve radiotherapy with targeted drugs. Nat Rev Cancer 2011; 11:239 - 53; http://dx.doi.org/10.1038/nrc3007; PMID: 21430696
- Takabe K, Paugh SW, Milstien S, Spiegel S. “Inside-out” signaling of sphingosine-1-phosphate: therapeutic targets. Pharmacol Rev 2008; 60:181 - 95; http://dx.doi.org/10.1124/pr.107.07113; PMID: 18552276
- Pyne S, Bittman R, Pyne NJ. Sphingosine kinase inhibitors and cancer: seeking the golden sword of Hercules. Cancer Res 2011; 71:6576 - 82; http://dx.doi.org/10.1158/0008-5472.CAN-11-2364; PMID: 21940750
- Ruckhäberle E, Rody A, Engels K, Gaetje R, von Minckwitz G, Schiffmann S, Grösch S, Geisslinger G, Holtrich U, Karn T, et al. Microarray analysis of altered sphingolipid metabolism reveals prognostic significance of sphingosine kinase 1 in breast cancer. Breast Cancer Res Treat 2008; 112:41 - 52; http://dx.doi.org/10.1007/s10549-007-9836-9; PMID: 18058224
- Guan H, Song L, Cai J, Huang Y, Wu J, Yuan J, Li J, Li M. Sphingosine kinase 1 regulates the Akt/FOXO3a/Bim pathway and contributes to apoptosis resistance in glioma cells. PLoS One 2011; 6:e19946; http://dx.doi.org/10.1371/journal.pone.0019946; PMID: 21625639
- Shida D, Takabe K, Kapitonov D, Milstien S, Spiegel S. Targeting SphK1 as a new strategy against cancer. Curr Drug Targets 2008; 9:662 - 73; http://dx.doi.org/10.2174/138945008785132402; PMID: 18691013
- Stevenson CE, Takabe K, Nagahashi M, Milstien S, Spiegel S. Targeting sphingosine-1-phosphate in hematologic malignancies. [Review] Anticancer Agents Med Chem 2011; 11:794 - 8; http://dx.doi.org/10.2174/187152011797655122; PMID: 21707492
- Chautard E, Loubeau G, Tchirkov A, Chassagne J, Vermot-Desroches C, Morel L, Verrelle P. Akt signaling pathway: a target for radiosensitizing human malignant glioma. Neuro Oncol 2010; 12:434 - 43; PMID: 20406894
- Toker A, Yoeli-Lerner M. Akt signaling and cancer: surviving but not moving on. [Review] Cancer Res 2006; 66:3963 - 6; http://dx.doi.org/10.1158/0008-5472.CAN-06-0743; PMID: 16618711
- Rödel F, Hoffmann J, Distel L, Herrmann M, Noisternig T, Papadopoulos T, Sauer R, Rödel C. Survivin as a radioresistance factor, and prognostic and therapeutic target for radiotherapy in rectal cancer. Cancer Res 2005; 65:4881 - 7; http://dx.doi.org/10.1158/0008-5472.CAN-04-3028; PMID: 15930309
- Asanuma K, Kobayashi D, Furuya D, Tsuji N, Yagihashi A, Watanabe N. A role for survivin in radioresistance of pancreatic cancer cells. Jpn J Cancer Res 2002; 93:1057 - 62; http://dx.doi.org/10.1111/j.1349-7006.2002.tb02483.x; PMID: 12359060
- Shen S, Kepp O, Kroemer G. The end of autophagic cell death?. Autophagy 2012; 8:1 - 3; http://dx.doi.org/10.4161/auto.8.1.16618; PMID: 22082964
- Zhang N, Qi Y, Wadham C, Wang L, Warren A, Di W, Xia P. FTY720 induces necrotic cell death and autophagy in ovarian cancer cells: a protective role of autophagy. Autophagy 2010; 6:1157 - 67; http://dx.doi.org/10.4161/auto.6.8.13614; PMID: 20935520
- Wallington-Beddoe CT, Hewson J, Bradstock KF, Bendall LJ. FTY720 produces caspase-independent cell death of acute lymphoblastic leukemia cells. Autophagy 2011; 7:707 - 15; http://dx.doi.org/10.4161/auto.7.7.15154; PMID: 21460633
- Liao AJ, Hu R, Li YC, Yao K, Wang HH, Zhang R, Yang W, Liu ZG. [The study of FTY720 on inducing apoptosis and autophagy in multiple myeloma cell line U266]. Zhonghua Xue Ye Xue Za Zhi 2011; 32:664 - 7; PMID: 22339823
- Liao A, Hu R, Zhao Q, Li J, Li Y, Yao K, Zhang R, Wang H, Yang W, Liu Z. Autophagy induced by FTY720 promotes apoptosis in U266 cells. Eur J Pharm Sci 2012; 45:600 - 5; http://dx.doi.org/10.1016/j.ejps.2011.12.014; PMID: 22281442
- Young MM, Kester M, Wang HG. Sphingolipids: regulators of crosstalk between apoptosis and autophagy. J Lipid Res 2013; 54:5 - 19; http://dx.doi.org/10.1194/jlr.R031278; PMID: 23152582
- Klionsky DJ, Emr SD. Autophagy as a regulated pathway of cellular degradation. Science 2000; 290:1717 - 21; http://dx.doi.org/10.1126/science.290.5497.1717; PMID: 11099404
- Meijer AJ, Codogno P. Regulation and role of autophagy in mammalian cells. Int J Biochem Cell Biol 2004; 36:2445 - 62; http://dx.doi.org/10.1016/j.biocel.2004.02.002; PMID: 15325584
- Qu X, Zou Z, Sun Q, Luby-Phelps K, Cheng P, Hogan RN, Gilpin C, Levine B. Autophagy gene-dependent clearance of apoptotic cells during embryonic development. Cell 2007; 128:931 - 46; http://dx.doi.org/10.1016/j.cell.2006.12.044; PMID: 17350577
- Tassone P, Blotta S, Palmieri C, Masciari S, Quaresima B, Montagna M, D’Andrea E, Eramo OP, Migale L, Costanzo F, et al. Differential sensitivity of BRCA1-mutated HCC1937 human breast cancer cells to microtubule-interfering agents. Int J Oncol 2005; 26:1257 - 63; PMID: 15809716
- Marvaso G, Barone A, Vaccaro C, Bruzzaniti V, Grespi S, Scotti V, Bianco C. Repeat stereotactic radiosurgery in the management of brain metastases from NSCLC: A case report and review of the literature. Oncol Lett 2013; 6:897 - 900; PMID: 24137433
- Amodio N, Di Martino MT, Foresta U, Leone E, Lionetti M, Leotta M, Gullà AM, Pitari MR, Conforti F, Rossi M, et al. miR-29b sensitizes multiple myeloma cells to bortezomib-induced apoptosis through the activation of a feedback loop with the transcription factor Sp1. Cell Death Dis 2012; 3:e436; http://dx.doi.org/10.1038/cddis.2012.175; PMID: 23190608
- Tagliaferri P, Katsaros D, Clair T, Neckers L, Robins RK, Cho-Chung YS. Reverse transformation of Harvey murine sarcoma virus-transformed NIH/3T3 cells by site-selective cyclic AMP analogs. J Biol Chem 1988; 263:409 - 16; PMID: 2826444
- Amodio N, Leotta M, Bellizzi D, Di Martino MT, D’Aquila P, Lionetti M, Fabiani F, Leone E, Gullà AM, Passarino G, et al. DNA-demethylating and anti-tumor activity of synthetic miR-29b mimics in multiple myeloma. Oncotarget 2012; 3:1246 - 58; PMID: 23100393
- Amodio N, Scrima M, Palaia L, Salman AN, Quintiero A, Franco R, Botti G, Pirozzi P, Rocco G, De Rosa N, et al. Oncogenic role of the E3 ubiquitin ligase NEDD4-1, a PTEN negative regulator, in non-small-cell lung carcinomas. Am J Pathol 2010; 177:2622 - 34; http://dx.doi.org/10.2353/ajpath.2010.091075; PMID: 20889565