Abstract
Autophagy is a catabolic process that allows cellular macromolecules to be broken down and recycled into metabolic precursors. It is a highly conserved, critical process, allowing cells to gain survival advantages under various stress situations due to growth and environmental changes. In the past few years, mounting evidence indicates that the post-transcriptional and translational controls mediated by non-coding miRNAs contribute significantly to autophagy in cancer. Such acute modulation of protein synthesis mediated by miRNAs provides cells with advantages in response to starvation, genotoxic stress and hypoxia. In this review, we highlight some of the important discoveries and molecular insights of miRNAs in regulating autophagy based on various cancer models.
Introduction
In the past decade, tremendous progress has been made in understanding the molecular and cellular process of autophagy. Macroautophagy (referred to as autophagy) is an essential and highly conserved critical catabolic process that delivers cytoplasmic components to lysosomes for degradation. The process involves enwrapping fractions of the cytoplasm until a double-membrane autophagic vacuole (autophagosome) is formed. The pathways and genes, such as multiple autophagy-related genes (ATGs), involved in autophagy that have been identified in detail in recent years include ATG1, ATG4, LC3/ATG8 and beclin-1.Citation1,Citation2
It is well-established that post-transcriptional and translational controls play important roles during stress situations. Such controls provide cells with acute responses to growth condition changes. Only in the past few years have we begun to appreciate the contribution and involvement of non-coding miRNAs in this process. miRNAs are non-coding RNA molecules, 18–25 nucleotides in length, which regulate the expression of their target genes by translational arrest or mRNA cleavage, most likely, through interaction mainly at the 3′-UTRs of the target mRNAs.Citation3-Citation5 Base pairing between at least six consecutive nucleotides within the 5′-seed of the miRNA with the target site on the mRNA is reported to be a minimum requirement for the miRNA-mRNA interaction.Citation3,Citation4 MiRNAs have been found to regulate many cellular processes, including apoptosis,Citation6-Citation9 differentiationCitation4,Citation10,Citation11 and cell proliferation.Citation6,Citation11-Citation13 Deregulation of miRNAs has been associated with cancer development and progression, and miRNAs have emerged as a new research frontier for understanding cancer development at the post-transcriptional and translational level.Citation14 Most of the past efforts of studying autophagy focused on key proteins with critical roles (first dimension) in the direct autophagy processing and the signaling pathways involved in transcriptional activation (second dimension). The contributions of post-transcriptional and translational controls (third dimension) of autophagy mediated by miRNAs emerged just recently.
MiRNAs Involved in Regulating the Expression of Key Autophagy-Related Proteins
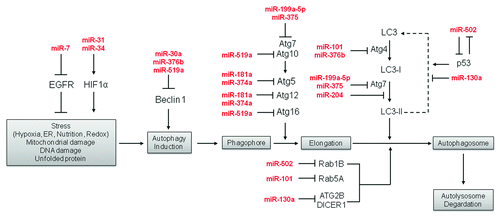
Zhu et al. first reported the involvement of miRNA in autophagy and cancer by providing experimental evidence that miR-30a targets beclin-1, a critical scaffold protein for autophagosome formation.Citation15 They have demonstrated that miR-30a downregulates beclin-1 expression, which mimics blunted activation of autophagy induced by rapamycin. Most recent reports further support the functional significance of miR-30a-mediated autophagy by enhancing Imatinib activity against human chronic myeloid leukemia cells.Citation16,Citation17 miR-30a also sensitizes tumor cells to cisplatin by suppressing beclin-1-mediated autophagy.Citation18 These results support a new treatment development strategy of overcoming chemoresistance by modulating miRNA-mediated autophagy.
Recently, more miRNAs have been reported to be mediators of the autophagic process. Jegga et al. proposed that miR-130, miR-98, miR-124, miR-204 and miR-142 have potential regulatory functions in the autophagic process based on computational analysis.Citation19 Later, miR-130a was shown to inhibit autophagic flux in chronic lymphocytic leukemia (CLL) cells by decreasing the expression level of its targets, ATG2B and DICER1, which are essential for autophagosome formation.Citation20 miR-101 has been recently demonstrated as a potent inhibitor of autophagy. miR-101 suppresses autophagy induced by etoposide or rapamycin in breast cancer cells. A number of key targets, such as STMN1, RAB5A and ATG4D have been identified as direct targets of miR-101.Citation21 miR-375 has been reported to inhibit autophagy through its target, ATG7, in hepatocellular carcinoma (HCC) cells.Citation22 miR-376b expression targets ATG4C and beclin-1, which, in turn, downregulate autophagy induced by nutrition starvation and rapamycin in breast cancer cells.Citation23 On the other hand, ectopic expression of miR-7 in human lung cancer and esophageal cancer cell lines enhances autophagy by suppressing epidermal growth factor receptor (EGFR) expression.Citation24
We have recently identified that miR-502 directly suppresses autophagy by decreasing the expression of RAB1B in colon cancer cell lines.Citation25 RAB1B is a small GTPase from Ras super family that has been demonstrated to modulate autophagic activity in HeLa cells through the regulation of autophagosome formation.Citation26 Rab1B has been shown to regulate vesicle trafficking at multiple stages and directly impact autophagyCitation27,Citation28 and was found to be overexpressed in liver cancer.Citation29 Ectopic expression of miR-502 in HCT-116 cells interrupted autophagic flux under acute and prolonged nutrient starvation.Citation25
Besides the contribution to cell proliferation or cell death, drug-induced autophagy also has been shown to play an important role in cancer chemoresistance, especially for cisplatin.Citation30-Citation32 Cisplatin treatment activates autophagy in multiple cancer cells and decreases the level of several miRNAs at the same time, which target multiple key regulators in autophagy, such as miR-199a-5p against ATG7 in HCC,Citation33 miR-181a and miR-374a against ATG5, miR-630 against ATG12 and miR-519a against beclin-1, ATG10 and ATG16 in squamous cell carcinoma (SCC) cells.Citation34
The miRNA-mediated genes and pathways involved in autophagy are illustrated in .
Figure 1. The miRNA-mediated genes and pathways involved in autophagy.
MiRNAs mediated autophagy in genotoxic stress and hypoxia
Evidence suggests that autophagy plays a prominent role in allowing cells to survive under stressful conditions such as hypoxia. While the exact role autophagy plays in cancer development remains uncertain, it is clear that the induction of autophagy in tumor cells helps them survive the hypoxic conditions associated with the inadequate blood supply to the tumor. It has been shown that miRNAs can mediate this autophagic response to hypoxia. In particular, miR-375, normally downregulated in HCC when exogenously expressed, was shown to inhibit autophagy in response to hypoxia by targeting ATG7, reducing the conversion of LC3I to LC3II. In vivo mouse xenograft studies demonstrated that miR-375 expression reduced tumor growth compared with controls.Citation35 Similar results were found looking at hypoxia in cardiomyocytes. Jian et al. found that miR-204 is downregulated by hypoxia-reoxygenation treatment, but when overexpressed, reduced the conversion of LC3I to LC3II.Citation36 These findings lend support to the idea that the regulation of autophagy by miRNAs may be a viable target for treatment of cancer, making tumor cells less resistant to the inherited hypoxic conditions of the tumor or more susceptible to the effects of other drugs. The story is complex, however, as other miRNAs have been shown to play a role in the activation of autophagy. In Cav(−/−) breast tumor stromal cells, miR-31 and miR-34 were both found to be upregulated. These miRNAs are associated with oxidative stress response and activation of autophagy through HIF1α, promoting cancer cell survival.Citation37,Citation38 miR-34 has also been implicated in the response to DNA damage through both p53-dependent and -independent mechanisms.Citation39 Much like the role of autophagy in cancer development, the role that miRNAs play in cancer development and autophagy regulation is complex. The complexity is most likely influenced by the unique tumor microenvironment. Much work needs to be done to better elucidate the dynamic role of miRNAs in the regulation of autophagy in response to cellular stress, as well as the effects that this response has on the cell.
MiRNA, Cell Cycle Control and Autophagy
Because one particular miRNA can regulate a number of target mRNA transcripts to control their translation, this provides cells with greater flexibility to utilize miRNA in response to growth condition changes and to different environments. This has been recently demonstrated for miR-10b. In breast cancer, miR-10b promotes breast tumor invasion and metastasis by suppressing homeobox D10 (HOXD10).Citation40 In contrast, miR-10b has been demonstrated to promote cell proliferation and prevents death of glioblastoma cells by targeting cell cycle inhibitors and pro-apoptotic genes.Citation41 miR-34c has been shown to regulate the cell cycle in response to DNA damage through its regulation of c-Myc.Citation39 Following DNA damage, miR-34c, activated either by p53 or p32 MAPK/MK2, represses c-Myc, preventing further DNA synthesis, arresting the cells in S-phase.Citation39 We have also demonstrated the flexibility of miRNA to regulate both the cell cycle and autophagy. We showed that miR-502, through its effect on different targets, can both regulate autophagy and cell cycle arrest at both the G1 and G2 phase. Overexpression of miR-502 in HCT-116 cells both inhibited autophagy and increased the number of cells in G1 and G2 phases in comparison to S phase.Citation25
MiRNA: The Third Dimension of p53-Mediated Autophagy
Cisplatin has been shown to induce autophagy in cancer cellsCitation33,Citation34 and activate p53 family members,Citation42,Citation43 which, in turn, function as transcriptional regulators to affect downstream gene expression, including miRNAs.Citation44-Citation46 In SCC cells, cisplatin exposure phosphorylates ΔNp63α, which directly regulates the transcription of many miRNAs.Citation44 Among the direct targets, miR-181a, miR-519a, miR-374a and miR-630 has been shown to regulate autophagy-related proteins,Citation34 which suggests that miRNAs can modulate autophagy pathway at another translational level.
A number of previous studies have reported the direct involvement of p53 in autophagy. The cytoplasmic pool of p53 suppresses autophagy under nutrient deprivation. Moreover, p53 also functions as a key regulator of autophagy.Citation47-Citation50 Under prolonged nutrition starvation, p53 sustained active autophagic flux, which is beneficial for colon cancer cell survival.Citation50
Since translational control provides cells with acute responses to growth stress, we reasoned that some of the p53-mediated miRNAs may play important roles in autophagy. We have provided evidence that certain miRNAs (miR-502) are also involved in the p53-mediated autophagy pathway.Citation25 Ectopic expression of miR-502 in colon cancer cells decreases p53 and upregulation of p53 triggers downregulation of miR-502, suggesting a negative feedback loop between p53 and miR-502. Moreover, overexpression of miR-502 in wild-type p53 cells leads to stalled autophagic flux, and this phenomenon is more prominent in p53-null cells, indicating miR-502 has a broader impact on autophagy that is beyond p53. The impact of miR-502 on autophagy pathway is schematically demonstrated in .
Therapeutic Potential of miRNAs and Autophagic Pathway
There is mounting evidence to link autophagy and cancer,Citation51,Citation52 and the role of autophagy in cancer progression is still unclear. Several reports demonstrated that activation of autophagy may suppress tumor development and lead to cell death.Citation53,Citation54 However, it has been demonstrated extensively that autophagy mediates tumor survival by supplying nutrients to stressed cancer cells.Citation55 Therefore, an anti-autophagy approach may also offer a therapeutic strategy for treating cancer.Citation54,Citation56,Citation57 One of the examples is rapamycin and its analogs (e.g., RAD001), which inhibit a kinase-named mammalian target of rapamycin (mTOR). Moreover, it has been demonstrated that inhibition of autophagy by 3-methyladenine or siRNA against Atg7 enhances the apoptosis induced by 5-FU treatment in colorectal cancer cell lines in vitro and in vivo.Citation58 In Ras-driven tumors, it has shown that Ras mediates cell transformation and activates autophagy, which is essential for tumor cell survival.Citation55 Inhibition of autophagy in Ras-expressing cells leads to decreased cell proliferation, transformation and failure of tumor formation in mice.Citation59-Citation61 Contrary to this, other studies have shown an anticancer role by enhancing autophagy.Citation47,Citation52,Citation62,Citation63 provides a list of miRNAs reported with therapeutic potential for treating cancer. It seems clear that the unique oncogenes/pathways involved in autophagy, the specific tumor type and tumor environment and the disease context are some of the key factors to be considered for developing autophagy-mediated anticancer strategies.
Table 1. miRNAs reported with therapeutic potential for treating cancer
In the case of miRNA, a number of reports have shown that miRNA mimics or anti-miRs can be used as potential therapeutics for anticancer drug development. Breast cancer cells treated with miR-101 showed increased cell death mediated by 4-hydroxytamoxifen (4-OHT) due to the inhibition of autophagy,Citation21 suggesting that microRNAs may modulate chemosensitivity of cancer cells. miR-30a also enhances the tumor cell apoptosis induced by cisplatin through suppressing belcin-1-mediated autophagy.Citation18 miR-221/222 expression on the other hand has been shown to increase tamoxifen resistance in breast cancer by decreasing levels of p27.Citation64 Similarly, miR-21 expression level is found to be higher in radio-resistant glioblastoma cells than in radio-sensitive ones.Citation65 Inhibition of miR-21 can increase the radio-sensitivity of glioblastoma cells and enhance the apoptosis after radiation through upregulate autophagic activity.Citation65
We have demonstrated that miR-502 can significantly decrease colon cancer cell growth in vitro and induce cell cycle arrest. Inhibition of both p53 and RAB1B, the mediators for autophagy, can reproduce this phenotype, suggesting that autophagy plays an important role in its tumor-suppressive function. Profiling of human colon cancer samples reveals that miR-502 is downregulated in tumor tissue as compared with normal tissue. Ectopic expression of miR-502 in human colon cancer xenografts can significantly reduce the tumor sizes, indicating miR-502 as a potential adjuvant treatment for colon cancer patients.Citation25
Conclusion and Future Perspectives
It is clear that miRNAs contribute to autophagy under various stress situations, especially in cancer. As cancer cells grow under stress conditions due to hypoxia, miRNAs will impact cancer cell survival by modulating autophagy. Therefore, it is conceivable that modulating miRNA will open a new direction to change cancer cells in response to stress by altering the autophagy process and, in turn, will provide new therapeutic strategies to overcome chemoresistance.
| Abbreviations: | ||
| ATG | = | autophagy-related gene |
| CLL | = | chronic lymphocytic leukemia |
| EGFR | = | epidermal growth factor receptor |
| HCC | = | heptocellular carcinoma |
| 4-OHT | = | 4-hydroxytamoxifen |
Acknowledgments
We apologize to all scientists whose important work may not be cited in this review due to space and/or time constraints. This study was supported by R01CA155019 (J.J.) and R33CA147966 (J.J.).
References
- Zhou S, Zhao L, Kuang M, Zhang B, Liang Z, Yi T, et al. Autophagy in tumorigenesis and cancer therapy: Dr. Jekyll or Mr. Hyde?. Cancer Lett 2012; 323:115 - 27; http://dx.doi.org/10.1016/j.canlet.2012.02.017; PMID: 22542808
- Fu LL, Wen X, Bao JK, Liu B. MicroRNA-modulated autophagic signaling networks in cancer. Int J Biochem Cell Biol 2012; 44:733 - 6; http://dx.doi.org/10.1016/j.biocel.2012.02.004; PMID: 22342941
- Lee RC, Feinbaum RL, Ambros V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 1993; 75:843 - 54; http://dx.doi.org/10.1016/0092-8674(93)90529-Y; PMID: 8252621
- Wightman B, Ha I, Ruvkun G. Posttranscriptional regulation of the heterochronic gene lin-14 by lin-4 mediates temporal pattern formation in C. elegans. Cell 1993; 75:855 - 62; http://dx.doi.org/10.1016/0092-8674(93)90530-4; PMID: 8252622
- He L, Hannon GJ. MicroRNAs: small RNAs with a big role in gene regulation. Nat Rev Genet 2004; 5:522 - 31; http://dx.doi.org/10.1038/nrg1379; PMID: 15211354
- Brennecke J, Hipfner DR, Stark A, Russell RB, Cohen SM. bantam encodes a developmentally regulated microRNA that controls cell proliferation and regulates the proapoptotic gene hid in Drosophila. Cell 2003; 113:25 - 36; http://dx.doi.org/10.1016/S0092-8674(03)00231-9; PMID: 12679032
- Chan JA, Krichevsky AM, Kosik KS. MicroRNA-21 is an antiapoptotic factor in human glioblastoma cells. Cancer Res 2005; 65:6029 - 33; http://dx.doi.org/10.1158/0008-5472.CAN-05-0137; PMID: 16024602
- Hwang HW, Mendell JT. MicroRNAs in cell proliferation, cell death, and tumorigenesis. Br J Cancer 2006; 94:776 - 80; http://dx.doi.org/10.1038/sj.bjc.6603023; PMID: 16495913
- Ghodgaonkar MM, Shah RG, Kandan-Kulangara F, Affar EB, Qi HH, Wiemer E, et al. Abrogation of DNA vector-based RNAi during apoptosis in mammalian cells due to caspase-mediated cleavage and inactivation of Dicer-1. Cell Death Differ 2009; 16:858 - 68; http://dx.doi.org/10.1038/cdd.2009.15; PMID: 19229243
- Tang F. Small RNAs in mammalian germline: Tiny for immortal. Differentiation 2010; 79:141 - 6; http://dx.doi.org/10.1016/j.diff.2009.11.002; PMID: 20227007
- Navarro F, Lieberman J. Small RNAs guide hematopoietic cell differentiation and function. J Immunol 2010; 184:5939 - 47; http://dx.doi.org/10.4049/jimmunol.0902567; PMID: 20483778
- He L, Thomson JM, Hemann MT, Hernando-Monge E, Mu D, Goodson S, et al. A microRNA polycistron as a potential human oncogene. Nature 2005; 435:828 - 33; http://dx.doi.org/10.1038/nature03552; PMID: 15944707
- Johnson CD, Esquela-Kerscher A, Stefani G, Byrom M, Kelnar K, Ovcharenko D, et al. The let-7 microRNA represses cell proliferation pathways in human cells. Cancer Res 2007; 67:7713 - 22; http://dx.doi.org/10.1158/0008-5472.CAN-07-1083; PMID: 17699775
- Ryan BM, Robles AI, Harris CC. Genetic variation in microRNA networks: the implications for cancer research. Nat Rev Cancer 2010; 10:389 - 402; http://dx.doi.org/10.1038/nrc2867; PMID: 20495573
- Zhu H, Wu H, Liu X, Li B, Chen Y, Ren X, et al. Regulation of autophagy by a beclin 1-targeted microRNA, miR-30a, in cancer cells. Autophagy 2009; 5:816 - 23; PMID: 19535919
- Yu Y, Cao L, Yang L, Kang R, Lotze M, Tang D. microRNA 30A promotes autophagy in response to cancer therapy. Autophagy 2012; 8:853 - 5; http://dx.doi.org/10.4161/auto.20053; PMID: 22617440
- Yu Y, Yang L, Zhao M, Zhu S, Kang R, Vernon P, et al. Targeting microRNA-30a-mediated autophagy enhances imatinib activity against human chronic myeloid leukemia cells. Leukemia 2012;; 26:1752 - 60; http://dx.doi.org/10.1038/leu.2012.65; PMID: 22395361
- Zou Z, Wu L, Ding H, Wang Y, Zhang Y, Chen X, et al. MicroRNA-30a sensitizes tumor cells to cis-platinum via suppressing beclin 1-mediated autophagy. J Biol Chem 2012; 287:4148 - 56; http://dx.doi.org/10.1074/jbc.M111.307405; PMID: 22157765
- Jegga AG, Schneider L, Ouyang X, Zhang J. Systems biology of the autophagy-lysosomal pathway. Autophagy 2011; 7:477 - 89; http://dx.doi.org/10.4161/auto.7.5.14811; PMID: 21293178
- Kovaleva V, Mora R, Park YJ, Plass C, Chiramel AI, Bartenschlager R, et al. miRNA-130a targets ATG2B and DICER1 to inhibit autophagy and trigger killing of chronic lymphocytic leukemia cells. Cancer Res 2012; 72:1763 - 72; http://dx.doi.org/10.1158/0008-5472.CAN-11-3671; PMID: 22350415
- Frankel LB, Wen J, Lees M, Høyer-Hansen M, Farkas T, Krogh A, et al. microRNA-101 is a potent inhibitor of autophagy. EMBO J 2011; 30:4628 - 41; http://dx.doi.org/10.1038/emboj.2011.331; PMID: 21915098
- Chang Y, Yan W, He X, Zhang L, Li C, Huang H, et al. miR-375 inhibits autophagy and reduces viability of hepatocellular carcinoma cells under hypoxic conditions. Gastroenterology 2012; 143:177 - 87, e8; http://dx.doi.org/10.1053/j.gastro.2012.04.009; PMID: 22504094
- Korkmaz G, le Sage C, Tekirdag KA, Agami R, Gozuacik D. miR-376b controls starvation and mTOR inhibition-related autophagy by targeting ATG4C and BECN1. Autophagy 2012; 8:165 - 76; http://dx.doi.org/10.4161/auto.8.2.18351; PMID: 22248718
- Tazawa H, Yano S, Yoshida R, Yamasaki Y, Sasaki T, Hashimoto Y, et al. Genetically engineered oncolytic adenovirus induces autophagic cell death through an E2F1-microRNA-7-epidermal growth factor receptor axis. Int J Cancer 2012; 131:2939 - 50; http://dx.doi.org/10.1002/ijc.27589; PMID: 22492316
- Zhai H, Song B, Xu X, Zhu W, Ju J. Inhibition of autophagy and tumor growth in colon cancer by miR-502. Oncogene 2012; In press http://dx.doi.org/10.1038/onc.2012.167; PMID: 22580605
- Zoppino FC, Militello RD, Slavin I, Alvarez C, Colombo MI. Autophagosome formation depends on the small GTPase Rab1 and functional ER exit sites. Traffic 2010; 11:1246 - 61; http://dx.doi.org/10.1111/j.1600-0854.2010.01086.x; PMID: 20545908
- Plutner H, Cox AD, Pind S, Khosravi-Far R, Bourne JR, Schwaninger R, et al. Rab1b regulates vesicular transport between the endoplasmic reticulum and successive Golgi compartments. J Cell Biol 1991; 115:31 - 43; http://dx.doi.org/10.1083/jcb.115.1.31; PMID: 1918138
- Stenmark H. Rab GTPases as coordinators of vesicle traffic. Nat Rev Mol Cell Biol 2009; 10:513 - 25; http://dx.doi.org/10.1038/nrm2728; PMID: 19603039
- He H, Dai F, Yu L, She X, Zhao Y, Jiang J, et al. Identification and characterization of nine novel human small GTPases showing variable expressions in liver cancer tissues. Gene Expr 2002; 10:231 - 42; PMID: 12450215
- Claerhout S, Verschooten L, Van Kelst S, De Vos R, Proby C, Agostinis P, et al. Concomitant inhibition of AKT and autophagy is required for efficient cisplatin-induced apoptosis of metastatic skin carcinoma. Int J Cancer 2010; 127:2790 - 803; http://dx.doi.org/10.1002/ijc.25300; PMID: 21351258
- Harhaji-Trajkovic L, Vilimanovich U, Kravic-Stevovic T, Bumbasirevic V, Trajkovic V. AMPK-mediated autophagy inhibits apoptosis in cisplatin-treated tumour cells. J Cell Mol Med 2009; 13:9B 3644 - 54; http://dx.doi.org/10.1111/j.1582-4934.2009.00663.x; PMID: 20196784
- Ren JH, He WS, Nong L, Zhu QY, Hu K, Zhang RG, et al. Acquired cisplatin resistance in human lung adenocarcinoma cells is associated with enhanced autophagy. Cancer Biother Radiopharm 2010; 25:75 - 80; http://dx.doi.org/10.1089/cbr.2009.0701; PMID: 20187799
- Xu N, Zhang J, Shen C, Luo Y, Xia L, Xue F, et al. Cisplatin-induced downregulation of miR-199a-5p increases drug resistance by activating autophagy in HCC cell. Biochem Biophys Res Commun 2012; 423:826 - 31; http://dx.doi.org/10.1016/j.bbrc.2012.06.048; PMID: 22713463
- Huang Y, Guerrero-Preston R, Ratovitski EA. Phospho-ΔNp63α-dependent regulation of autophagic signaling through transcription and micro-RNA modulation. Cell Cycle 2012; 11:1247 - 59; http://dx.doi.org/10.4161/cc.11.6.19670; PMID: 22356768
- Chang Y, Yan W, He X, Zhang L, Li C, Huang H, et al. miR-375 inhibits autophagy and reduces viability of hepatocellular carcinoma cells under hypoxic conditions. Gastroenterology 2012; 143:177 - 87, e8; http://dx.doi.org/10.1053/j.gastro.2012.04.009; PMID: 22504094
- Jian X, Xiao-yan Z, Bin H, Yu-feng Z, Bo K, Zhi-nong W, et al. MiR-204 regulate cardiomyocyte autophagy induced by hypoxia-reoxygenation through LC3-II. Int J Cardiol 2011; 148:110 - 2; http://dx.doi.org/10.1016/j.ijcard.2011.01.029; PMID: 21316776
- Pavlides S, Tsirigos A, Migneco G, Whitaker-Menezes D, Chiavarina B, Flomenberg N, et al. The autophagic tumor stroma model of cancer: Role of oxidative stress and ketone production in fueling tumor cell metabolism. Cell Cycle 2010; 9:3485 - 505; http://dx.doi.org/10.4161/cc.9.17.12721; PMID: 20861672
- Martinez-Outschoorn UE, Whitaker-Menezes D, Pavlides S, Chiavarina B, Bonuccelli G, Casey T, et al. The autophagic tumor stroma model of cancer or “battery-operated tumor growth”: A simple solution to the autophagy paradox. Cell Cycle 2010; 9:4297 - 306; http://dx.doi.org/10.4161/cc.9.21.13817; PMID: 21051947
- Cannell IG, Kong YW, Johnston SJ, Chen ML, Collins HM, Dobbyn HC, et al. p38 MAPK/MK2-mediated induction of miR-34c following DNA damage prevents Myc-dependent DNA replication. Proc Natl Acad Sci USA 2010; 107:5375 - 80; http://dx.doi.org/10.1073/pnas.0910015107; PMID: 20212154
- Ma L, Teruya-Feldstein J, Weinberg RA. Tumour invasion and metastasis initiated by microRNA-10b in breast cancer. Nature 2007; 449:682 - 8; http://dx.doi.org/10.1038/nature06174; PMID: 17898713
- Gabriely G, Yi M, Narayan RS, Niers JM, Wurdinger T, Imitola J, et al. Human glioma growth is controlled by microRNA-10b. Cancer Res 2011; 71:3563 - 72; http://dx.doi.org/10.1158/0008-5472.CAN-10-3568; PMID: 21471404
- Murray-Zmijewski F, Lane DP, Bourdon JC. p53/p63/p73 isoforms: an orchestra of isoforms to harmonise cell differentiation and response to stress. Cell Death Differ 2006; 13:962 - 72; http://dx.doi.org/10.1038/sj.cdd.4401914; PMID: 16601753
- Oren M. Decision making by p53: life, death and cancer. Cell Death Differ 2003; 10:431 - 42; http://dx.doi.org/10.1038/sj.cdd.4401183; PMID: 12719720
- Huang Y, Chuang A, Hao H, Talbot C, Sen T, Trink B, et al. Phospho-ΔNp63α is a key regulator of the cisplatin-induced microRNAome in cancer cells. Cell Death Differ 2011; 18:1220 - 30; http://dx.doi.org/10.1038/cdd.2010.188; PMID: 21274007
- Ory B, Ramsey MR, Wilson C, Vadysirisack DD, Forster N, Rocco JW, et al. A microRNA-dependent program controls p53-independent survival and chemosensitivity in human and murine squamous cell carcinoma. J Clin Invest 2011; 121:809 - 20; http://dx.doi.org/10.1172/JCI43897; PMID: 21293058
- Zhang X, Lu X. Posttranscriptional regulation of miRNAs in the DNA damage response. RNA Biol 2011; 8:960 - 3; http://dx.doi.org/10.4161/rna.8.6.17337; PMID: 21941125
- Tasdemir E, Maiuri MC, Galluzzi L, Vitale I, Djavaheri-Mergny M, D’Amelio M, et al. Regulation of autophagy by cytoplasmic p53. Nat Cell Biol 2008; 10:676 - 87; http://dx.doi.org/10.1038/ncb1730; PMID: 18454141
- Tasdemir E, Maiuri MC, Orhon I, Kepp O, Morselli E, Criollo A, et al. p53 represses autophagy in a cell cycle-dependent fashion. Cell Cycle 2008; 7:3006 - 11; http://dx.doi.org/10.4161/cc.7.19.6702; PMID: 18838865
- Tasdemir E, Chiara Maiuri M, Morselli E, Criollo A, D’Amelio M, Djavaheri-Mergny M, et al. A dual role of p53 in the control of autophagy. Autophagy 2008; 4:810 - 4; PMID: 18604159
- Scherz-Shouval R, Weidberg H, Gonen C, Wilder S, Elazar Z, Oren M. p53-dependent regulation of autophagy protein LC3 supports cancer cell survival under prolonged starvation. Proc Natl Acad Sci USA 2010; 107:18511 - 6; http://dx.doi.org/10.1073/pnas.1006124107; PMID: 20937856
- Shintani T, Klionsky DJ. Autophagy in health and disease: a double-edged sword. Science 2004; 306:990 - 5; http://dx.doi.org/10.1126/science.1099993; PMID: 15528435
- White E, DiPaola RS. The double-edged sword of autophagy modulation in cancer. Clin Cancer Res 2009; 15:5308 - 16; http://dx.doi.org/10.1158/1078-0432.CCR-07-5023; PMID: 19706824
- Elgendy M, Sheridan C, Brumatti G, Martin SJ. Oncogenic Ras-induced expression of Noxa and Beclin-1 promotes autophagic cell death and limits clonogenic survival. Mol Cell 2011; 42:23 - 35; http://dx.doi.org/10.1016/j.molcel.2011.02.009; PMID: 21353614
- Liang XH, Jackson S, Seaman M, Brown K, Kempkes B, Hibshoosh H, et al. Induction of autophagy and inhibition of tumorigenesis by beclin 1. Nature 1999; 402:672 - 6; http://dx.doi.org/10.1038/45257; PMID: 10604474
- Mancias JD, Kimmelman AC. Targeting autophagy addiction in cancer. Oncotarget 2011; 2:1302 - 6; PMID: 22185891
- Degenhardt K, Mathew R, Beaudoin B, Bray K, Anderson D, Chen G, et al. Autophagy promotes tumor cell survival and restricts necrosis, inflammation, and tumorigenesis. Cancer Cell 2006; 10:51 - 64; http://dx.doi.org/10.1016/j.ccr.2006.06.001; PMID: 16843265
- Mathew R, Karantza-Wadsworth V, White E. Role of autophagy in cancer. Nat Rev Cancer 2007; 7:961 - 7; http://dx.doi.org/10.1038/nrc2254; PMID: 17972889
- Li J, Hou N, Faried A, Tsutsumi S, Kuwano H. Inhibition of autophagy augments 5-fluorouracil chemotherapy in human colon cancer in vitro and in vivo model. Eur J Cancer 2010; 46:1900 - 9; http://dx.doi.org/10.1016/j.ejca.2010.02.021; PMID: 20231086
- Guo JY, Chen HY, Mathew R, Fan J, Strohecker AM, Karsli-Uzunbas G, et al. Activated Ras requires autophagy to maintain oxidative metabolism and tumorigenesis. Genes Dev 2011; 25:460 - 70; http://dx.doi.org/10.1101/gad.2016311; PMID: 21317241
- Kim MJ, Woo SJ, Yoon CH, Lee JS, An S, Choi YH, et al. Involvement of autophagy in oncogenic K-Ras-induced malignant cell transformation. J Biol Chem 2011; 286:12924 - 32; http://dx.doi.org/10.1074/jbc.M110.138958; PMID: 21300795
- Lock R, Roy S, Kenific CM, Su JS, Salas E, Ronen SM, et al. Autophagy facilitates glycolysis during Ras-mediated oncogenic transformation. Mol Biol Cell 2011; 22:165 - 78; http://dx.doi.org/10.1091/mbc.E10-06-0500; PMID: 21119005
- Boya P, González-Polo RA, Casares N, Perfettini JL, Dessen P, Larochette N, et al. Inhibition of macroautophagy triggers apoptosis. Mol Cell Biol 2005; 25:1025 - 40; http://dx.doi.org/10.1128/MCB.25.3.1025-1040.2005; PMID: 15657430
- Maiuri MC, Zalckvar E, Kimchi A, Kroemer G. Self-eating and self-killing: crosstalk between autophagy and apoptosis. Nat Rev Mol Cell Biol 2007; 8:741 - 52; http://dx.doi.org/10.1038/nrm2239; PMID: 17717517
- Miller TE, Ghoshal K, Ramaswamy B, Roy S, Datta J, Shapiro CL, et al. MicroRNA-221/222 confers tamoxifen resistance in breast cancer by targeting p27Kip1. J Biol Chem 2008; 283:29897 - 903; http://dx.doi.org/10.1074/jbc.M804612200; PMID: 18708351
- Gwak HS, Kim TH, Jo GH, Kim YJ, Kwak HJ, Kim JH, et al. Silencing of microRNA-21 confers radio-sensitivity through inhibition of the PI3K/AKT pathway and enhancing autophagy in malignant glioma cell lines. PLoS One 2012; 7:e47449; http://dx.doi.org/10.1371/journal.pone.0047449; PMID: 23077620