Abstract
The pathogenesis of acne, a disease of the pilosebaceous follicle and one of the most common chronic skin disorders, is attributed to multiple factors such as increased sebum production, alteration of the quality of sebum lipids, inflammatory processes, dysregulation of the hormone microenvironment, interaction with neuropeptides, follicular hyperkeratinisation and the proliferation of Propionibacterium acnes within the follicle. In particular, the sebaceous gland plays an exquisite role in the initiation of the disease as it possesses all the enzyme machinery for the production of hormones and cytokines. In addition, in response to the altered tissue environment in the pilosebaceous follicle as well as in answer to emotional fret, stress response system mechanisms with induction of central and local expression of neuropeptides, are also initiated. This review summarises the latest advances in understanding the role of sebaceous gland cells in the pathomechanism of acne.
Introduction
Sebaceous glands are holocrine glands found over the entire surface of the body except the palms, soles and dorsum of the feet. They are largest and most concentrated in the face and scalp where they are the sites of origin of acne (). The normal function of sebaceous glands is to produce and secrete sebum, a group of complex oils including triglycerides and fatty acid breakdown products, wax esters, squalene, cholesterol esters and cholesterol.Citation1–Citation4 Sebum lubricates the skin to protect against friction and makes it more impervious to moisture. Furthermore, the sebaceous gland transports antioxidants in and on the skin and exhibits a natural light protective activity. It possesses an innate antibacterial activity and has a pro- and anti-inflammatory function. It can regulate the activity of xenobiotics and is actively involved in the wound healing process.Citation5
In the last years, acne research has made a remarkable progress in understanding the mechanisms involved in the pathogenesis of the disease by using cell culture models and new molecular techniques. Mammal sebocytes and sebocyte-like cells (human, mouse, hamster and rat) and human sebaceous gland cell lines (SZ95, SEB-1, Seb-E6E7)Citation6–Citation8 have been used in monolayer cultures as models to study specific functions involved in development, growth and differentiation of sebaceous gland cells. More complex culture systems, including three-dimensional models, are under development.
Sebum and Acne
Increased sebum excretion, alteration of lipid composition and the oxidant/antioxidant ratio characteristic of the skin surface lipids are major concurrent events associated with the development of acne.Citation5 If sebum interferes with the process of follicular keratinisation in the pilosebaceous unit, pore blockage may occur, contributing to lesion formation and acne. However, seborrhoea per se is not considered to be the only responsible factor for the development of acne, as demonstrated by the success of treatment with agents with no effect on sebum secretion rate that can inhibit the inflammatory process, such as antibiotics, topical retinoids, azelaic acid and benzoyl peroxide.Citation9 The composition of the produced lipids is also of great importance. Lower essential fatty acid levels were found in wax esters in twins with acne rather than in twins with no acne.Citation10 Moreover, low levels of linoleic acid have been observed in skin surface lipids of acne patients.Citation11 Evidence suggests that diet may be an important source of substrate for the synthesis of sebaceous lipids.Citation12 This notion is supported also by the observation that sebum contains linoleic acid, an essential fatty acid that cannot be synthesised in vivo and therefore must be obtained from the diet. It has recently been hypothesised that low glycemic load diet may influence sebum production based on the beneficial endocrine effects of its components.Citation13
On the other hand, extreme caloric restriction dramatically decreases the sebum excretion rate and these changes can be reversed when a normal diet is resumed.Citation14,Citation15 Other studies have demonstrated that increased consumption of dietary fat or carbohydrate increases sebum production and modifications to the type of carbohydrate can also alter sebum composition.Citation16 Typical western diet, comprised of milk and hyperglycaemic foods, may have potentiating effects on serum insulin and insulin-like growth factor-I (IGF-I) levels, thereby promoting the development of acne.Citation17
Another hallmark of sebum in acne patients is the presence of lipoperoxides, mainly due to the peroxidation of squalene and a decrease in the level of vitamin E, the major sebum antioxidant.Citation18 Both lipoperoxides and monounsaturated fatty acid (MUFA) are capable of inducing alteration in keratinocyte proliferation and differentiation, whereas peroxides are capable of inducing production of pro-inflammatory cytokines and activation of peroxisome proliferator-activated receptors (PPAR).Citation13,Citation18
The biological function of sebocytes is further regulated by several factors including ligands of receptors expressed in sebocytes, such as androgens and estrogens, PPAR ligands and neuropeptides (NP), liver-X receptor ligands (LXR), histamines, retinoids and vitamin D. The ligand-receptor complexes activate pathways involving cell proliferation, differentiation, lipogenesis, hormone metabolism and cytokine and chemokine releaseCitation19 ().
LXR, which are members of the nuclear receptor superfamily, and play a critical role in cholesterol homeostasis and lipid metabolism have been documented to regulate lipid synthesis in the immortalised human sebaceous gland cell line SZ95. Treatment of SZ95 sebocytes with LXR ligands such as TO901317 or 22(R)-hydroxycholesterol, enhanced accumulation of lipid droplets in the cells which could be explained through induction of the expression of the LXRalpha receptor and known LXR targets, such as fatty acid synthase and sterol regulatory-binding protein-1.Citation20,Citation21
On the other hand, sebaceous function can be also significantly modified by histamine and conversely, antihistamines. Diphenhydramine (DPH), a histamine 1 receptor antagonist, significantly decreases squalene levels in human sebaceous gland cells as determined by means of high-performance liquid chromatography. These data were further verified by the identification of histamine 1 receptor in human sebaceous glands.Citation22
Retinoids are also suggested to influence the biological function of sebocytes. Retinoic acid receptors (RAR; isotypes α and γ) and retinoid X receptors (RXR; isotypes α, β, γ) are expressed in human sebocytes.Citation23 The natural ligands for RAR and RXR are all-trans (atRA) and 9-cis retinoic acid (9cRA). In SZ95 sebocytes 13-cis retinoic acid (13cRA) may unfold its action through a marked isomerisation to atRA. All three isoforms atRA, 13cRA and 9cRA exhibit anti-proliferative effectsCitation24 and inhibit sebocyte differentiation and lipid synthesis.Citation25 RXR agonists stimulate sebocyte differentiation and proliferation. The RXR agonist rexinoid in combination with the specific PPAR agonists, WY 14643, troglitazone and cabaprostacyclin affect differentiation and growth in cultured primary sebocyte-like rat preputial cells.Citation26
The enzymatic machinery for the local synthesis and metabolism of 1, 25-dihydroxyvitamin D (3) [1,25(OH)(2) D(3), calcitriol] has been also investigated in human sebocytes. Vitamin D receptor, vitamin D-25-hydroxylase, 25-hydroxyvitamin D-1alpha-hydroxylase and 1, 25-dihydroxyvitamin D-24-hydroxylase are expressed in SZ95 sebocytes in vitro. Furthermore, incubation of SZ95 sebocytes with 1,25(OH)(2)D(3) leads to a dose-dependent modulation of cell proliferation, cell cycle regulation, lipid content and interleukin (IL)-6/IL-8 secretion in vitro.Citation27 In hamster auricular sebocytes while epidermal growth factor and atRA can decrease the intracellular accumulation of triglycerides and free fatty acids in the cells, 1α, 25-dihydroxyvitamin D3 decreases the triglyceride level but augments the accumulation of wax esters. No difference has been detected in the level of cholesterol after the above treatments.Citation28
Effects of Hormones on Sebaceous Gland Cells
Sex steroids.
Several studies have demonstrated that there is an association between local overproduction of active androgens and acne. Acne patients produced higher rates of testosterone and 5α-dihydrotestosterone (5α-DHT) in their skin than healthy individuals.Citation29 High testosterone levels have been implicated with enhanced sebaceous gland activity in humansCitation30,Citation31 and consequently with diseases marked by hyperseborrhea, such as Acne vulgaris. However, only a few patients with androgenic disorders exhibit hyperandrogenemia, an observation which indicates the predominance of peripheral tissue events for the occurrence of clinical signs.Citation32
Enhanced sebaceous gland activity is attributed to the potent androgen 5α-DHTCitation5 as sebaceous gland cells possess all necessary enzymes for conversion of testosterone to 5α-DHT.Citation33 The isozyme 5α-reductase type I, which catalyses the conversion from testosterone to 5α-DHT in peripheral tissues by a NADPH-dependent reaction is expressed predominantly in skin. It is present in the cytoplasm and cell membrane compartment in skin cellsCitation34 and particularly in facial sebocytes,Citation33 illustrating the key role of sebaceous gland cells in androgen metabolism.
The effects of testosterone and 5α-DHT are mediated by binding to the nuclear androgen receptor (AR), also expressed in human sebaceous gland cells.Citation35 AR is a member of the steroid superfamily of ligand-dependent transcription factors. 5α-DHT binds to the AR with greater affinity than testosterone and the 5α-DHT/AR complex appears to be more stableCitation36 and therefore, more effective.
In contrast to the in vivo observations, in vitro experiments with human sebocytes have shown that testosterone affects proliferation in a dose-dependent mannerCitation6,Citation37 but does not affect lipid synthesis.Citation38,Citation39 This contradiction has led to the assumption that co-factors may be required for the induction of the entire so-called androgenic influence of the sebaceous gland.Citation40 Current research has indicated that PPAR and their ligands, may be the primary candidates.Citation38,Citation39 PPAR regulate multiple lipid metabolism genes in mitochondria, peroxisomes and microsomes, all prominent in sebocyte cytoplasm.Citation38,Citation39
Indeed, Rosenfield et al.Citation38 have previously demonstrated the interaction of 5α-DHT with PPAR ligands in inducing differentiation of sebocyte-like rat preputial cells and lipid synthesis. PPARα is the most important PPAR that regulates lipid synthesis and inflammation.Citation40,Citation41 In addition, PPAR-α, -δ, -γ1 and -γ2 have been shown to be expressed at mRNA and protein levels in SZ95 sebocytes.Citation39
Dehydroepiandrosterone (DHEA) has been also shown to regulate sebum production especially in postmenopausal women.Citation22 Consequently, several researchers have suggested the use of DHEA as an anti-aging agent.Citation42,Citation43 However, in in vitro experiments DHEA has been shown to have no direct effect on the biological activity of human sebocytes. Substitution with DHEA in elderly persons is accompanied by a small increase of testosterone and estradiol, which may indeed yield an explanation of the clinical change demonstratedCitation22 suggesting that the action of DHEA may be implemented through indirect pathways.
Growth factors.
Growth hormone (GH) activity is considered to be mainly attributed to IGF but GH has also been shown to exhibit direct effects on human skin cells.Citation44 The increased serum GH levels in acromegaly are associated with enhanced sebum secretion,Citation45 an observation that could be confirmed by GH treatment of human SZ95 sebocytes in vitro.Citation46 In acne vulgaris, increased sebum production peaks in mid-adolescence at a time that GH and IGF-I reach their highest serum levels.Citation47 In mini rats, suppression of GH gene expression by an antisense transgene, leads among to thinner skin with less collagen and increase of subcutaneous adipose tissue also to small-sized sebaceous glands.Citation48
Increased serum levels of IGF-I have been observed in adult women and men with acne and the number of total acne lesions, inflammatory lesions, serum levels of 5α-DHT and DHEA sulphate, each correlated with serum IGF-I levels in women with acne.Citation49,Citation50 A correlation between the mean facial sebum excretion rate and serum IGF-I levels has been demonstrated in postadolescent acne patients.Citation51 IGF-I has been localised to the peripheral cells of sebaceous glands in the rat,Citation52 while in human skin the strongest expression of IGF-I protein has been found in maturing sebocytes and suprabasal cells of sebaceous ducts.Citation53 The expression of IGF-I receptor mRNA is the strongest in basal cells of sebaceous glands and immature sebocytes, whereas IGF-I receptor protein expression was uniform and intense in all regions of the gland.Citation53 In animal studies, IGF-I has been shown to stimulate sebocyte differentiation in vitro especially in combination with GH,Citation47 while in human keratinocytes it acts as a mitogen.Citation54 On the other hand, in humans, IGF-I plays a key role in the induction of lipid synthesis in human sebocytes.Citation46,Citation55 In SEB-1 sebocytes, IGF-I increases lipogenesis by the induction of sterol response element-binding protein-1 (SREBP-1)Citation55 through activation of PI3K/Akt and MAPK/ERK-signal transduction pathway.Citation56 SREBP-1 preferentially regulates genes of fatty acid synthesis.Citation56 In the hamster ear sebaceous model, androgens rapidly induce the expression of SREBP-1.Citation57 In addition, an interaction between the IGF-I and estradiol signalling pathway has been described in human SZ95 sebocytes, implicating that estrogens may have an indirect effect on the pathogenesis of acne.Citation46
Recent data suggest that incubation of human sebaceous gland cells with a hormone mixture consisting of growth factors and sex steroids at age-specific levels may alter the biological activity of the cells by regulating their transcriptome and thus illustrate the importance of the hormone environment for cell function.Citation58 Human SZ95 sebocytes treated with hormone levels that can be found in 60 year-old women produce less lipids than sebocytes treated with a hormone mixture representing that found in serum of 20 year-old women.Citation58 Gene expression profiling via cDNA microarray between SZ95 sebocytes under the 20 and 60 year-old hormone mixture detected differentially expressed genes, which are involved in biological processes such as DNA repair and stability, mitochondrial function, oxidative stress, cell cycle and apoptosis, ubiquitin-induced proteolysis and transcriptional regulation. The most significantly altered signalling pathway was that of transforming growth factor-β (TGFβ). A disturbed function of this cascade has been also associated with tumourigenesis, i.e., in pancreatic, prostate, intestine, breast and uterine cancer. Interestingly, genes expressed in signalling pathways operative in age-associated diseases such as Huntington's disease, dentatorubral-pallidoluysian atrophy and amyotrophic lateral sclerosis were also identified. These data demonstrate that hormones interact in a complex fashion, and sebocytes may be affected to a large extent by the changes in their circulating blood levels with age.Citation58
Effects of NPs on Sebaceous Gland Cells
NP are a heterogeneous group of biologically active peptides that are present in neurons of both the central and peripheral nervous systems. However, human skin and in particular the human sebaceous gland has been shown to express functional receptors for NP, such as corticotropin-releasing hormone (CRH), melanocortins, β-endorphin, vasoactive intestinal polypeptide, neuropeptide Y and calcitonin gene-related peptide. These receptors modulate the production of inflammatory cytokines, proliferation, differentiation, lipogenesis and androgen metabolism in human sebocytes.Citation5
CRH the most proximal element of the hypophysis-pituitary-adrenal (HPA) axis acts as central coordinator for neuroendocrine and behavioural responses to stress. It has been shown that CRH, CRH-binding protein, CRH-receptor®1 and CRH-R2 are expressed in SZ95 sebocytes at mRNA and protein level, while CRH-R1 is the predominant type (CRH-R1/CRH-R2 = 2). In addition, CRH significantly induces sebaceous lipids production, IL-6 and -8 synthesis and may upregulate mRNA levels of 3β-hydroxysteroid dehydrogenase/Δ5-4 isomerase.Citation59,Citation60 In acne-involved skin the complete CRH system is abundant especially in the sebaceous glands (), possibly activating pathways which affect immune and inflammatory processes leading to the development and stress-induced exacerbation of acne.Citation61
Melanocortin (MC) peptides can also directly affect the function of human sebocytes via MC receptors. α-Melanocyte-stimulating hormone (α-MSH) has been demonstrated to act as a modulator of the rat preputial gland, a specialised sebaceous gland-like structure of rodents.Citation62 The presence of both MC-1R and MC-5R which bind α-MSH were detected in primary cell cultures of facial human sebocytes. The expression of MC-5R is weaker than that of MC1-R but it has been shown to be a marker of human sebocyte differentiation, since it is expressed in differentiated, lipid-containing sebocytes only.Citation63,Citation64 In acne-involved skin, sebocytes and keratinocytes of the ductus seboglandularis showed very intense MC-1R expression in contrast to less intense scattered immunoreactivity in normal skin samples suggesting that this receptor is involved in the initiation of acne.Citation65 MC-1R expression has been shown to be upregulated by proinflammatory signals.Citation66,Citation67 As proinflammatory cytokines are upregulated in acne lesions,Citation68 sebocytes may respond to these signals with increased MC-1R expression, thereby generating a negative feedback mechanism for α-MSH which exerts direct anti-inflammatory actions, i.e., inhibition of IL-1-mediated IL-8 secretion.Citation64,Citation65
Cannabinoid receptors (CR), which mediate the psychopharmacological action of marijuana have been not only localised in the central and peripheral nervous system but also in human skin. CR1 and 2 are expressed in human sebaceous glands,Citation69 whereas the CB2 and other prototypic endocannabinoids are present in SZ95 sebocytes and may induce in a dose-dependent manner lipid production and cell death. These actions are selectively mediated by CB2-coupled signalling involving the MAPK pathway.Citation70
Other NP such as substance P or vasointestinal peptide may also be involved in the pathogenesis of acne vulgaris Substance P, which can be elicited by stress may promote the development of cytoplasmic organelles in sebaceous cells, stimulate sebaceous germinative cells, and induce significant increases in the area of sebaceous glands. It also increases the size of individual sebaceous cells and the number of sebum vacuoles for each differentiated sebaceous cell, all of which suggests that substance P promotes both the proliferation and the differentiation of sebaceous glands. Substance P induces the expression of neutral endopeptidase, a potent NP-degrading enzyme, in sebaceous germinative cells and of E-selectin by perisebaceous venules. Facial skin from acne patients is characterised by rich innervation, by increased numbers of substance P-containing nerves and mast cells, and by strong expression of neutral endopeptidase in sebaceous glands and E-selectin in venules around sebaceous glands, compared with normal skin.Citation71 The ectopeptidases dipeptidyl peptidase IV (DP IV or CD26) and aminopeptidase N (APN or CD13), which have been shown to be involved in the degradation of several NP, especially of substance P, have been found to be highly expressed in human sebocytes in vivo and in vitro. Further studies have shown unexpectedly that inhibitors of DP IV and APN can suppress proliferation and slightly decrease neutral lipids, but can also enhance terminal differentiation in SZ95 sebocytes. This suggests that ectopeptidases may be new targets to modulate certain sebocyte functions, and that ectopeptidase inhibitors may have potential therapeutic roles in acne pathogenesis.Citation72
A central integrator of nociception, the transient receptor potential vanilloid-1 (TRPV1) is expressed in human skin, sebaceous glands in situ and in SZ95 sebocytes in vitro. It has been documented that the prototypic TRPV1 agonist, capsaicin, selectively inhibits basal and arachidonic acid-induced lipid synthesis in a dose-, time- and extracellular calcium-dependent and TRPV1-specific manner. Low-dose capsaicin stimulates cellular proliferation via TRPV1, whereas higher concentrations inhibit sebocyte growth and induce cell death independent of TRPV1.Citation73 These findings suggest the strong involvement of neurogenic factors and sebocytes in the disease process of acne.
Inflammation, Sebaceous Gland Cells and Acne
Inflammation is being regarded as a key component of the pathogenesis of acne.Citation74 In the last few years, there has been a debate as to whether hyperkeratinisation of the follicular duct precedes the influx of inflammatory cells or vice versa. Recent studies support the latter hypothesis by demonstrating that an increase in IL-1 activity occurs before the hyperproliferation around uninvolved follicles and this triggers the activation of the keratinocytes.Citation68,Citation75 Expression profiling of acne-involved and uninvolved skin from acne patients and from subjects without acne via cDNA microarrays have given us a better insight into the etiological factors giving rise to acne.Citation76 In inflammatory acne lesions, the majority of the regulated genes, which showed to be upregulated are involved in inflammatory processes. These include matrix metalloproteinases, β-defensin 4, IL-8 and granulysin. No differences were noted between normal skin from acne patients and that from patients without acne in the array analysis. NFκB, a transcription factor critical for upregulation of many proinflammatory cytokine genes has been shown to be activated in acne lesions.Citation77 NFκB-regulated cytokine mRNA genes levels of TNFα, IL-1β, IL-8 and IL-10 are significantly upregulated in acne-involved skin compared to uninvolved normal adjacent skin. Elevated expression of the chemokine IL-8 is able to attract circulating inflammatory cells into the tissue. Indeed, in lesional skin of acne, there is a marked increase in the presence of neutrophils, as compared to the uninvolved skin whereas lymphocytes are prominently visible in inflammatory acne lesions as compared to normal controls.Citation77 Another transcription factor involved in inflammation, AP-1 has been shown to be activated in inflammatory acne lesions in vivo as well. Levels of the proinflammatory cytokine IL-1 were also upregulated perifollicularly in uninvolved skin from acne patients. This cytokine may be responsible for the cutaneous inflammation and the resulting keratinocyte proliferation and may play a profound role in the transformation of a normal follicle into an acne lesion.Citation68
Inflammation is further characterised by action of active lipid mediators, such as leucotrienes (LT), prostaglandins (PG) and 15-hydroxyeicosatetraenoic acids (15-HETE). These molecules are synthesised from arachidonic acid (AA) or linolenic acid by the enzymes lipoxygenase (LOX) and cyclooxygenase (COX), respectively. Both COX isozymes, COX-1 and COX-2, are expressed in human sebocytes in vitro, in particular COX-2 expression is selectively upregulated in acne involved sebaceous glands in vivoCitation41 (). In hamster sebocytes the expression of COX-2 has been also documented,Citation78 while the 15-desoxy-Δ12,14-PGJ2 has been shown to induce the lipid synthesis in the cells.Citation79 Activation of the platelet-activating factor signalling pathway (PAF, 1-O-alkyl-2-acetyl-sn-glycero-3-phosphocholine) which consists of a group of phosphocholines with various biological effects, including modulation of keratinocyte function and skin inflammation, can regulate the expression of inflammatory mediators, e.g., COX-2 and PGE2, as well as IL-8 in SZ95 sebocytes.Citation80 Transgenic keratin 5 promoter driven overexpression of COX-2 in the basal compartment of the epidermis of the mouse and increased PGE2 levels have been documented to cause sebaceous gland hyperplasia and overshooting sebum production pointing to a role of COX-2-mediated PGE2 synthesis in this process.Citation81 Activation of PPARγ by UVB irradiation and the potent lipid soluble oxidant tert-butylhydroperoxide (TBH) induces COX-2 expression in SZ95 sebocytes and this finding indicates a PPARγ COX-2-mediated pathway regulating sebocyte proliferation and/or lipogenesis.Citation82
LT are potent proinflammatory mediators and neutrophil attractants produced from arachidonic acid by the enzyme 5-LOX. Human sebocytes express all necessary enzymes for a functional LT pathway. The enzymes 5-LOX and LTA4 hydrolase are expressed in SZ95 sebocytes at protein and mRNA level. These enzymes are essential for the formation of LTB4. On the other hand, 15-LOX expression shows a weak expression in SZ95 sebocytes, indicating that sebocytes do not play a significant role in the biosynthesis of the anti-inflammatory 15-HETE. Treatment of SZ95 sebocytes with AA stimulates 5-LOX expression and induces LTB4 synthesis.Citation41 In addition, AA induces the expression of the IL-6 and IL-8 cytokines. 5-LOX and LTA4 hydrolase show a stronger expression in acne lesions than in normal skin and in uninvolved skin of acne patients.Citation41 The involvement of 5-LOX in the pathogenesis of acne has led to new therapeutic strategies to deal with the disease.Citation83
Cytokines are present in normal sebaceous glands, and they are affected by many factors. IL-1α, TNFα, IL-6 and IL-8 are released into supernatant in unstressed sebocyte culture.Citation41 In a stressed environment, the amounts of released cytokines increase significantly. AA and calcium ionophore enhance the level of IL-6 and IL-8, but that of IL-1β and TNFα is not affected.Citation9,Citation41
Psoriasin, a member of the S100 gene family, was shown to be highly expressed in the epidermis and the ductus seboglandularis of acne-involved skin in contrast to uninvolved control.Citation84 Psoriasin has been suggested to be involved in the pathogenesis of several inflammatory skin diseases, and its levels increase in response to inflammatory stress. RA and inflammatory agents have been also implicated in the upregulation of psoriasin.Citation85,Citation86
Propionibacterium acnes (p. acnes)
P. acnes is a gram-positive anaerobic bacterium which with other non-pathogenic microorganisms, such as coagulase negative staphylococci and diphtheroid rods, resides in pilosebaceous follicles as a member of the resident bacterial flora. The mechanism by which P. acnes contributes to the pathogenesis of acne is debated. While in several studies it could be shown that P. acnes numbers are higher in acne patients than in healthy individuals, other studies found no difference between the numbers of P. acnes in affected and non-affected follicles. Nevertheless, an abnormal colonisation by P. acnes has been implicated in the occurrence of acne via the induction of inflammatory mediators. The bacteria stimulate the production of proinflammatory cytokines, including interleukins-1β, -8 and -12, and TNFα. It is known that P. acnes-induced cytokine production is mediated by Toll-like receptor (TLR) 2.Citation87–Citation90 The pilosebaceous unit is an immunocompetent organ. Keratinocytes and sebocytes may act as immune cells capable of pathogen recognition and abnormal lipid presentation. Both cell types can be activated by P. acnes via toll-like receptors (TLR), CD14 and CD1 molecules.Citation88 The expression of TLR2, TLR4, TLR6 and CD14 has been already documented in SZ95 sebocytes.Citation91,Citation92 Recent evidence has indicated that human sebaceous glands may contribute to the skin immune defense by releasing antimicrobial peptides (AMP). For example, human β-defensins (hBD) are expressed in human pilosebaceous units and their expression is upregulated in acne lesions.Citation93 Cathelicidin and hBD-2 are detected in cultured human sebocytes, the predominant cells residing in the sebaceous gland, and their expression levels are upregulated in the presence of P. acnes.Citation89,Citation94 Each P. acnes strain has been shown to influence sebocyte viability and differentiation differently, a fact that raises the possibility that certain P. acnes strains to be responsible for opportunistic infections worsening acne lesions.Citation89,Citation94,Citation95 A description of phylogenetically distinct P. acnes clusters has been already undertaken.Citation96
MUFA, mainly palmitic acid (C16:1) and oleic acid (C18:1), both of which are bactericidal against gram-positive organisms,Citation91 are produced by the sebaceous gland, as is sapienic acid, an important antimicrobial lipid. Stearoyl coenzyme A desaturase (SCD) 1, an enzyme responsible for the biosynthesis of MUFA, is also expressed by the sebaceous gland.Citation97 The TLR-2 ligand macrophage-activating lipopeptide-2 stimulates both SCD and fatty acid desaturase-2 mRNA expression in SZ95 sebocytes.Citation91 Lauric acid (C12:0), one of the sebum free fatty acids (FFA) has strong antimicrobial activity in vitro against skin bacteria, including P. acnes. Topical application or intradermal injection of lauric acid in vivo shows remarkable therapeutic effectiveness against P. acnes-induced inflammation and significant reduction in the number of bacteria.Citation98 Furthermore, lauric acid, palmitic acid (16:0) and oleic acid (C18:1, cis-9), which are the typical FFA found in human sebum, enhanced the hBD-2 expression and antimicrobial activity of human sebocytes against P. acnesCitation99 indicating that sebum FFA are involved in the disinfecting activity of the human skin both through their direct antimicrobial characteristics and by inducing AMP in human sebocytes to enhance their innate immune defense ability.
The treatment of cultured sebocytes with P. acnes and lipopolysaccharides (LPS) significantly upregulates the expression of proinflammatory cytokines.Citation89 There is a difference in the cytokine production curve over time after treatment between P. acnes and LPS. While LPS stimulates CXCL8, TNFα and IL-1α, P. acnes stimulates CXCL8 and TNFα only. P. acnes has no effect on IL-1α regulation. Furthermore, viable P. acnes and not heat-killed organisms can stimulate the release of cytokines such as IL-1β, GM-CSF and IL-8.Citation100,Citation101
Conclusion
Current research results have allowed us to elucidate a part of the mechanisms involved in the pathogenesis of one of the most common skin disorders, acne and critically revisit conventional concepts of its pathogenesis. In addition, it has helped us to determine new targets for future drug development. The sebaceous gland cell is a key player in the initiation of the disease and sebocyte culture models have become so far very useful tools to provide new chances for further research.
Abbreviations
| AA | = | arachidonic acid |
| ACTH | = | adrenocorticotropine hormone |
| AMPs | = | antimicrobial peptides |
| AR | = | androgen receptor |
| COX | = | cycloxygenase |
| CR | = | cannabinoid receptor |
| CRH | = | corticotropin-releasing hormone |
| DHEAS | = | dehydroepiandrosterone sulphate |
| 5α-DHT | = | 5α-dihydrostestosterone |
| DPH | = | diphenhydramine |
| GH | = | growth hormone |
| H-1 receptor | = | histamine-1 receptor |
| hBDs | = | human β-defensins |
| IGF-I | = | insulin growth factor-I |
| LXR | = | liver-X receptor |
| LOX | = | lipoxygenase |
| LT | = | leucotrienes |
| α-MSH | = | α-melanocyte-stimulating hormone |
| MMPs | = | matrix metalloproteinases |
| MC | = | melanocortin |
| MUFAs | = | monounsaturated fatty acid |
| NP | = | neuropeptide |
| PG | = | prostaglandins |
| P. acnes | = | Propionibacterium acnes |
| POMC | = | propiomelanocortin |
| PPAR | = | peroxisome proliferator-activated receptor |
| RA | = | retinoic acid |
| RAR | = | retinoid acid receptor |
| RXR | = | retinoid X receptor |
| SCD | = | stearoyl coenzyme A |
| SREBP-1 | = | sterol response element-binding protein-1 |
| TLRs | = | toll-like receptors |
| TNFα | = | tumor necrosis factorα |
| TRPV1 | = | transient receptor potential vanilloid-1 |
| VDR | = | vitamin D receptor |
Figures and Tables
Figure 1 Pilosebaceous unit in facial skin of acne patients. Faintly hypertrophic sebaceous gland are observed. Dilated capillaries and perivascular lymphocytes (A and C) are early signs of inflammatory process in acne-involved skin. Dilated plugged orifice of hair follicle—sign of acne comedo (B).
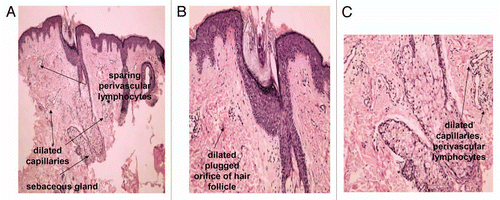
Figure 2 Regulation of the biological function of human sebaceous gland cells. Schematic overview. [LXR: liver X receptors, PPAR: peroxisome-proliferator activated receptors].
![Figure 2 Regulation of the biological function of human sebaceous gland cells. Schematic overview. [LXR: liver X receptors, PPAR: peroxisome-proliferator activated receptors].](/cms/asset/8f9286f3-bd7d-4a9c-8b03-ad783d46aa4d/kder_a_10913900_f0002.gif)
Figure 3 Localisation of CRH immunostaining in the sebaceous gland of acne patients (A) and in the normal skin of healthy controls (B). Very intensive expression of the gene in all types of sebocytes—basal, differentiating and mature cells—and keratinocytes of ductus seboglandularis in acne skin is shown (A) (x400); significant weaker and dependent upon sebocytes differentiation stage immunoreaction of sebaceous gland in normal skin (B) (x400).
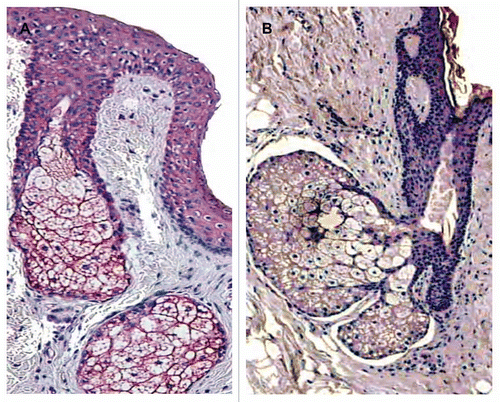
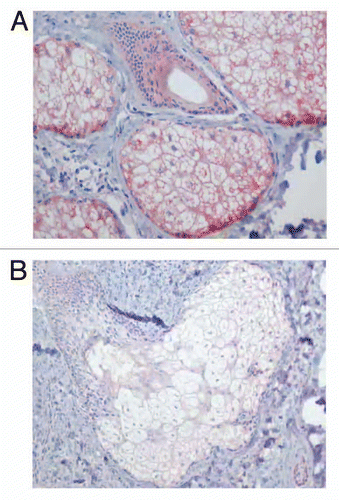
Figure 4 Localisation of COX-2 immunostaining in the sebaceous gland of acne patients (A) and in the normal skin of healthy controls (B). Very strong immunoreaction of the gene within the sebaceous gland of acne skin, especially, in undifferentiated and early differentiated sebocytes is seen (A) (x400). Weak immunoexpression of COX-2 in the sebaceous gland and ductal cells of healthy skin (B) (x400).
Acknowledgements
This work was supported by a research grant by the German Federal Ministry of Education and Research to C.C.Z. E.M. is grateful for having received the R.O.S.A. Prize (Research on Skin Dryness Award) 2008 and the Beauty Care Prize 2009 of the German Society of Dermatology.
References
- Downing DT, Stewart ME, Wertz PW, et al. Skin lipids: an update. J Invest Dermatol 1987; 88:2 - 6
- Thody AJ, Shuster S. Control and function of sebaceous glands. Physiol Rev 1989; 69:1 - 4
- Nikkari T, Schreibman PH, Ahrens EH Jr. In vivo studies of sterol and squalene secretion by human skin. J Lipid Res 1974; 15:563 - 573
- Ramasastry P, Downing DT, Pochi PE, et al. Chemical composition of human skin surface lipids from birth to puberty. J Invest Dermatol 1970; 54:139 - 144
- Zouboulis CC. Acne and sebaceous gland function. Clin Dermatol 2004; 22:360 - 366
- Zouboulis CC, Seltmann H, Neitzel H, et al. Establishment and characterization of an immortalized human sebaceous gland cell line (SZ95). J Invest Dermatol 1999; 113:1011 - 1020
- Thiboutot D, Jabara S, McAllister JM, et al. Human skin is a steroidogenic tissue: steroidogenic enzymes and cofactors are expressed in epidermis, normal sebocytes and an immortalized sebocyte cell line (SEB-1). J Invest Dermatol 2003; 120:905 - 914
- Lo Celso C, Berta MA, Braun KM, et al. Characterization of bipotential epidermal progenitors derived from human sebaceous gland: contrasting roles of c-Myc and beta-catenin. Stem Cells 2008; 26:1241 - 1252
- Kurokawa I, Danby FW, Ju Q, et al. New developments in our understanding of acne pathogenesis and treatment. Exp Dermatol 2009; 18:821 - 832
- Stewart ME. Sebaceous gland lipids. Semin Dermatol 1992; 11:100 - 105
- Downing DT, Stewart ME, Wertz PW, et al. Essential fatty acids and acne. J Am Acad Dermatol 1986; 14:221 - 225
- Rasmussen JE. Diet and acne. Int J Dermatol 1977; 16:488 - 492
- Smith RN, Braue A, Varigos GA, et al. The effect of a low glycemic load diet on acne vulgaris and the fatty acid composition of skin surface triglycerides. J Dermatol Sci 2008; 50:41 - 52
- Downing DT, Strauss JS, Pochi PE. Changes in skin surface lipid composition induced by severe caloric restriction in man. Am J Clin Nutr 1972; 25:365 - 367
- Pochi PE, Downing DT, Strauss JS. Sebaceous gland response in man to prolonged total caloric deprivation. J Invest Dermatol 1970; 55:303 - 309
- Macdonald I. Changes in the Fatty Acid Composition of Sebum Associated with High Carbohydrate Diets. Nature 1964; 203:1067 - 1068
- Melnik BC, Schmitz G. Role of insulin, insulin-like growth factor-1, hyperglycaemic food and milk consumption in the pathogenesis of acne vulgaris. Exp Dermatol 2009; 18:833 - 841
- Ottaviani M, Alestas T, Flori E, et al. Peroxidated squalene induces the production of inflammatory mediators in HaCaT keratinocytes: a possible role in acne vulgaris. J Invest Dermatol 2006; 126:2430 - 2437
- Zouboulis CC, Schagen S, Alestas T. The sebocyte culture: a model to study the pathophysiology of the sebaceous gland in sebostasis, seborrhoea and acne. Arch Dermatol Res 2008; 300:397 - 413
- Russell LE, Harrison WJ, Bahta AW, et al. Characterization of liver X receptor expression and function in human skin and the pilosebaceous unit. Exp Dermatol 2007; 16:844 - 852
- Hong I, Lee MH, Na TY, et al. LXRalpha enhances lipid synthesis in SZ95 sebocytes. J Invest Dermatol 2008; 128:1266 - 1272
- Baulieu EE, Thomas G, Legrain S, et al. Dehydroepiandrosterone (DHEA), DHEA sulfate and aging: contribution of the DHEAge Study to a sociobiomedical issue. Proc Natl Acad Sci USA 2000; 97:4279 - 4284
- Reichrath J, Mittmann M, Kamradt J, et al. Expression of retinoid-X receptors (-alpha, -beta, -gamma) and retinoic acid receptors (-alpha, -beta, -gamma) in normal human skin: an immunohistological evaluation. Histochem J 1997; 29:127 - 133
- Tsukada M, Schroder M, Roos TC, et al. 13-cis retinoic acid exerts its specific activity on human sebocytes through selective intracellular isomerization to all-trans retinoic acid and binding to retinoid acid receptors. J Invest Dermatol 2000; 115:321 - 327
- Zouboulis CC, Korge B, Akamatsu H, et al. Effects of 13-cis-retinoic acid, all-trans-retinoic acid and acitretin on the proliferation, lipid synthesis and keratin expression of cultured human sebocytes in vitro. J Invest Dermatol 1991; 96:792 - 797
- Kim MJ, Deplewski D, Ciletti N, et al. Limited cooperation between peroxisome proliferator-activated receptors and retinoid X receptor agonists in sebocyte growth and development. Mol Genet Metab 2001; 74:362 - 369
- Kramer C, Seltmann H, Seifert M, et al. Characterization of the vitamin D endocrine system in human sebocytes in vitro. J Steroid Biochem Mol Biol 2009; 113:9 - 16
- Sato T, Imai N, Akimoto N, et al. Epidermal growth factor and 1alpha,25-dihydroxyvitamin D3 suppress lipogenesis in hamster sebaceous gland cells in vitro. J Invest Dermatol 2001; 117:965 - 970
- Sansone G, Reisner RM. Differential rates of conversion of testosterone to dihydrotestosterone in acne and in normal human skin—a possible pathogenic factor in acne. J Invest Dermatol 1971; 56:366 - 372
- Pochi PE, Strauss JS. Sebaceous gland response in man to the administration of testosterone, delta-4-androstenedione and dehydroisoandrosterone. J Invest Dermatol 1969; 52:32 - 36
- Giltay EJ, Gooren LJ. Effects of sex steroid deprivation/administration on hair growth and skin sebum production in transsexual males and females. J Clin Endocrinol Metab 2000; 85:2913 - 2921
- Orfanos CE, Adler YD, Zouboulis CC. The SAHA syndrome. Horm Res 2000; 54:251 - 258
- Fritsch M, Orfanos CE, Zouboulis CC. Sebocytes are the key regulators of androgen homeostasis in human skin. J Invest Dermatol 2001; 116:793 - 800
- Chen W, Zouboulis CC, Fritsch M, et al. Heterogeneity and quantitative differences of type 1, 5-alpha-reductase expression in cultured skin epithelial cells. Dermatology 1998; 196:51 - 52
- Fimmel S, Saborowski A, Orfanos CE, et al. Development of efficient transient transfection systems for introducing antisense oligonucleotides into human epithelial skin cells. Horm Res 2000; 54:306 - 311
- Anderson KM, Liao S. Selective retention of dihydrotestosterone by prostatic nuclei. Nature 1968; 219:277 - 279
- Akamatsu H, Zouboulis CC, Orfanos CE. Control of human sebocyte proliferation in vitro by testosterone and 5-alpha-dihydrotestosterone is dependent on the localization of the sebaceous glands. J Invest Dermatol 1992; 99:509 - 511
- Rosenfield RL, Deplewski D, Kentsis A, et al. Mechanisms of androgen induction of sebocyte differentiation. Dermatology 1998; 196:43 - 46
- Chen W, Yang CC, Sheu HM, et al. Expression of peroxisome proliferator-activated receptor and CCAAT/enhancer binding protein transcription factors in cultured human sebocytes. J Invest Dermatol 2003; 121:441 - 447
- Zouboulis CC, Eady A, Philpott M, et al. What is the pathogenesis of acne?. Exp Dermatol 2005; 14:143 - 152
- Alestas T, Ganceviciene R, Fimmel S, et al. Enzymes involved in the biosynthesis of leukotriene B(4) and prostaglandin E(2) are active in sebaceous glands. J Mol Med 2006; 84:75 - 87
- Minghetti P, Cilurzo F, Casiraghi A, et al. Development of patches for the controlled release of dehydroepiandrosterone. Drug Dev Ind Pharm 2001; 27:711 - 717
- Shin MH, Rhie GE, Park CH, et al. Modulation of collagen metabolism by the topical application of dehydroepiandrosterone to human skin. J Invest Dermatol 2005; 124:315 - 323
- Deplewski D, Rosenfield RL. Role of hormones in pilosebaceous unit development. Endocr Rev 2000; 21:363 - 392
- Burton JL, Libman LJ, Cunliffe WJ, et al. Sebum excretion in acromegaly. Br Med J 1972; 1:406 - 408
- Makrantonaki E, Vogel K, Fimmel S, et al. Interplay of IGF-I and 17beta-estradiol at age-specific levels in human sebocytes and fibroblasts in vitro. Exp Gerontol 2008; 43:939 - 946
- Deplewski D, Rosenfield RL. Growth hormone and insulin-like growth factors have different effects on sebaceous cell growth and differentiation. Endocrinology 1999; 140:4089 - 4094
- Ikawa A, Ishii Y, Suzuki K, et al. Age-related changes in the dorsal skin histology in Mini and Wistar rats. Histol Histopathol 2002; 17:419 - 426
- Aizawa H, Niimura M. Elevated serum insulin-like growth factor-1 (IGF-1) levels in women with postadolescent acne. J Dermatol 1995; 22:249 - 252
- Cappel M, Mauger D, Thiboutot D. Correlation between serum levels of insulin-like growth factor 1, dehydroepiandrosterone sulfate and dihydrotestosterone and acne lesion counts in adult women. Arch Dermatol 2005; 141:333 - 338
- Vora S, Ovhal A, Jerajani H, et al. Correlation of facial sebum to serum insulin-like growth factor-1 in patients with acne. Br J Dermatol 2008; 159:990 - 991
- Hansson HA, Nilsson A, Isgaard J, et al. Immunohistochemical localization of insulin-like growth factor I in the adult rat. Histochemistry 1988; 89:403 - 410
- Rudman SM, Philpott MP, Thomas GA, et al. The role of IGF-I in human skin and its appendages: morphogen as well as mitogen?. J Invest Dermatol 1997; 109:770 - 777
- Tavakkol A, Varani J, Elder JT, et al. Maintenance of human skin in organ culture: role for insulin-like growth factor-1 receptor and epidermal growth factor receptor. Arch Dermatol Res 1999; 291:643 - 651
- Smith TM, Cong Z, Gilliland KL, et al. Insulin-like growth factor-1 induces lipid production in human SEB-1 sebocytes via sterol response element-binding protein-1. J Invest Dermatol 2006; 126:1226 - 1232
- Smith TM, Gilliland K, Clawson GA, et al. IGF-1 induces SREBP-1 expression and lipogenesis in SEB-1 sebocytes via activation of the phosphoinositide 3-kinase/Akt pathway. J Invest Dermatol 2008; 128:1286 - 1293
- Rosignoli C, Nicolas JC, Jomard A, et al. Involvement of the SREBP pathway in the mode of action of androgens in sebaceous glands in vivo. Exp Dermatol 2003; 12:480 - 489
- Makrantonaki E, Adjaye J, Herwig R, et al. Age-specific hormonal decline is accompanied by transcriptional changes in human sebocytes in vitro. Aging Cell 2006; 5:331 - 344
- Krause K, Schnitger A, Fimmel S, et al. Corticotropin-releasing hormone skin signaling is receptor-mediated and is predominant in the sebaceous glands. Horm Metab Res 2007; 39:166 - 170
- Zouboulis CC, Seltmann H, Hiroi N, et al. Corticotropin-releasing hormone: an autocrine hormone that promotes lipogenesis in human sebocytes. Proc Natl Acad Sci USA 2002; 99:7148 - 7153
- Ganceviciene R, Graziene V, Fimmel S, et al. Involvement of the corticotropin-releasing hormone system in the pathogenesis of acne vulgaris. Br J Dermatol 2009; 160:345 - 352
- Thody AJ, Cooper MF, Bowden PE, et al. Effect of alpha-melanocyte-stimulating hormone and testosterone on cutaneous and modified sebaceous glands in the rat. J Endocrinol 1976; 71:279 - 288
- Zhang L, Li WH, Anthonavage M, et al. Melanocortin-5 receptor: a marker of human sebocyte differentiation. Peptides 2006; 27:413 - 420
- Bohm M, Schiller M, Stander S, et al. Evidence for expression of melanocortin-1 receptor in human sebocytes in vitro and in situ. J Invest Dermatol 2002; 118:533 - 539
- Ganceviciene R, Graziene V, Bohm M, et al. Increased in situ expression of melanocortin-1 receptor in sebaceous glands of lesional skin of patients with acne vulgaris. Exp Dermatol 2007; 16:547 - 552
- Bhardwaj R, Becher E, Mahnke K, et al. Evidence for the differential expression of the functional alpha-melanocyte-stimulating hormone receptor MC-1 on human monocytes. J Immunol 1997; 158:3378 - 3384
- Hartmeyer M, Scholzen T, Becher E, et al. Human dermal microvascular endothelial cells express the melanocortin receptor type 1 and produce increased levels of IL-8 upon stimulation with alpha-melanocytestimulating hormone. J Immunol 1997; 159:1930 - 1937
- Jeremy AH, Holland DB, Roberts SG, et al. Inflammatory events are involved in acne lesion initiation. J Invest Dermatol 2003; 121:20 - 27
- Stander S, Schmelz M, Metze D, et al. Distribution of cannabinoid receptor 1 (CB1) and 2 (CB2) on sensory nerve fibers and adnexal structures in human skin. J Dermatol Sci 2005; 38:177 - 188
- Dobrosi N, Toth BI, Nagy G, et al. Endocannabinoids enhance lipid synthesis and apoptosis of human sebocytes via cannabinoid receptor-2-mediated signaling. Faseb J 2008; 22:3685 - 3695
- Toyoda M, Nakamura M, Morohashi M. Neuropeptides and sebaceous glands. Eur J Dermatol 2002; 12:422 - 427
- Thielitz A, Reinhold D, Vetter R, et al. Inhibitors of dipeptidyl peptidase IV and aminopeptidase N target major pathogenetic steps in acne initiation. J Invest Dermatol 2007; 127:1042 - 1051
- Toth BI, Geczy T, Griger Z, et al. Transient receptor potential vanilloid-1 signaling as a regulator of human sebocyte biology. J Invest Dermatol 2009; 129:329 - 339
- Zouboulis CC. Is acne vulgaris a genuine inflammatory disease?. Dermatology 2001; 203:277 - 279
- Freedberg IM, Tomic-Canic M, Komine M, et al. Keratins and the keratinocyte activation cycle. J Invest Dermatol 2001; 116:633 - 640
- Trivedi NR, Gilliland KL, Zhao W, et al. Gene array expression profiling in acne lesions reveals marked upregulation of genes involved in inflammation and matrix remodeling. J Invest Dermatol 2006; 126:1071 - 1079
- Kang S, Cho S, Chung JH, et al. Inflammation and extracellular matrix degradation mediated by activated transcription factors nuclear factorkappaB and activator protein-1 in inflammatory acne lesions in vivo. Am J Pathol 2005; 166:1691 - 1699
- Ito A, Sakiguchi T, Kitamura K, et al. Establishment of a tissue culture system for hamster sebaceous gland cells. Dermatology 1998; 197:238 - 244
- Iwata C, Akimoto N, Sato T, et al. Augmentation of lipogenesis by 15-deoxy-Delta12,14-prostaglandin J2 in hamster sebaceous glands: identification of cytochrome P-450-mediated 15-deoxy-Delta12,14-prostaglandin J2 production. J Invest Dermatol 2005; 125:865 - 872
- Zhang Q, Seltmann H, Zouboulis CC, et al. Activation of platelet-activating factor receptor in SZ95 sebocytes results in inflammatory cytokine and prostaglandin E2 production. Exp Dermatol 2006; 15:769 - 774
- Neufang G, Furstenberger G, Heidt M, et al. Abnormal differentiation of epidermis in transgenic mice constitutively expressing cyclooxygenase-2 in skin. Proc Natl Acad Sci USA 2001; 98:7629 - 7634
- Zhang Q, Seltmann H, Zouboulis CC, et al. Involvement of PPARgamma in oxidative stress-mediated prostaglandin E(2) production in SZ95 human sebaceous gland cells. J Invest Dermatol 2006; 126:42 - 48
- Zouboulis CC, Seltmann H, Alestas T. Zileuton prevents the activation of the leukotriene pathway and reduces sebaceous lipogenesis. Exp Dermatol 2009;
- Ganceviciene R, Fimmel S, Glass E, et al. Psoriasin and follicular hyperkeratinization in acne comedones. Dermatology 2006; 213:270 - 272
- Zouboulis CC, Voorhees JJ, Orfanos CE, et al. Topical all-trans retinoic acid (RA) induces an early, coordinated increase in RA-inducible skin-specific gene/psoriasin and cellular RA-binding protein II mRNA levels which precedes skin erythema. Arch Dermatol Res 1996; 288:664 - 669
- Tavakkol A, Zouboulis CC, Duell EA, et al. A retinoic acid-inducible skin-specific gene (RIS-1/psoriasin): molecular cloning and analysis of gene expression in human skin in vivo and cultured skin cells in vitro. Mol Biol Rep 1994; 20:75 - 83
- Kim J, Ochoa MT, Krutzik SR, et al. Activation of toll-like receptor 2 in acne triggers inflammatory cytokine responses. J Immunol 2002; 169:1535 - 1541
- Kim J. Review of the innate immune response in acne vulgaris: activation of Toll-like receptor 2 in acne triggers inflammatory cytokine responses. Dermatology 2005; 211:193 - 198
- Nagy I, Pivarcsi A, Kis K, et al. Propionibacterium acnes and lipopolysaccharide induce the expression of antimicrobial peptides and proinflammatory cytokines/chemokines in human sebocytes. Microbes Infect 2006; 8:2195 - 2205
- Koreck A, Pivarcsi A, Dobozy A, et al. The role of innate immunity in the pathogenesis of acne. Dermatology 2003; 206:96 - 105
- Georgel P, Crozat K, Lauth X, et al. A toll-like receptor 2-responsive lipid effector pathway protects mammals against skin infections with gram-positive bacteria. Infect Immun 2005; 73:4512 - 4521
- Oeff MK, Seltmann H, Hiroi N, et al. Differential regulation of Toll-like receptor and CD14 pathways by retinoids and corticosteroids in human sebocytes. Dermatology 2006; 213:266
- Chronnell CM, Ghali LR, Ali RS, et al. Human beta defensin-1 and -2 expression in human pilosebaceous units: upregulation in acne vulgaris lesions. J Invest Dermatol 2001; 117:1120 - 1125
- Lee DY, Yamasaki K, Rudsil J, et al. Sebocytes express functional cathelicidin antimicrobial peptides and can act to kill Propionibacterium acnes. J Invest Dermatol 2008; 128:1863 - 1866
- Graham GM, Farrar MD, Cruse-Sawyer JE, et al. Proinflammatory cytokine production by human keratinocytes stimulated with Propionibacterium acnes and P. acnes GroEL. Br J Dermatol 2004; 150:421 - 428
- McDowell A, Valanne S, Ramage G, et al. Propionibacterium acnes types I and II represent phylogenetically distinct groups. J Clin Microbiol 2005; 43:326 - 334
- Harrison WJ, Bull JJ, Seltmann H, et al. Expression of lipogenic factors galectin-12, resistin, SREBP-1 and SCD in human sebaceous glands and cultured sebocytes. J Invest Dermatol 2007; 127:1309 - 1317
- Nakatsuji T, Kao MC, Fang JY, et al. Antimicrobial property of lauric acid against Propionibacterium acnes: its therapeutic potential for inflammatory acne vulgaris. J Invest Dermatol 2009; 129:2480 - 2488
- Nakatsuji T, Kao MC, Zhang L, et al. Sebum Free Fatty Acids Enhance the Innate Immune Defense of Human Sebocytes by Upregulating beta-Defensin-2 Expression. J Invest Dermatol 2009;
- Nagy I, Pivarcsi A, Koreck A, et al. Distinct strains of Propionibacterium acnes induce selective human betadefensin-2 and interleukin-8 expression in human keratinocytes through toll-like receptors. J Invest Dermatol 2005; 124:931 - 938
- Schaller M, Loewenstein M, Borelli C, et al. Induction of a chemoattractive proinflammatory cytokine response after stimulation of keratinocytes with Propionibacterium acnes and coproporphyrin III. Br J Dermatol 2005; 153:66 - 71