Abstract
Because of rapidly increasing market demand and rising cost pressure, the innovator of etanercept (Enbrel®) will inevitably face competition from biosimilar versions of the product. In this study, to elucidate the differences between the reference etanercept and its biosimilars, we characterized and compared the quality attributes of two commercially available, biosimilar TNF receptor 2-Fc fusion protein products. Biosimilar 1 showed high similarity to Enbrel® in critical quality attributes including peptide mapping, intact mass, charge variant, purity, glycosylation and bioactivity. In contrast, the intact mass and MS/MS analysis of biosimilar 2 revealed a mass difference indicative of a two amino acid residue variance in the heavy chain (Fc) sequences. Comprehensive glycosylation profiling confirmed that biosimilar 2 has significantly low sialylated N-oligosaccharides. Biosimilar 2 also displayed significant differences in charge attributes compared with the reference product. Interestingly, biosimilar 2 exhibited similar affinity and bioactivity levels compared with the reference product despite the obvious difference in primary structure and partial physiochemical properties. For a biosimilar development program, comparative analytical data can influence decisions about the type and amount of animal and clinical data needed to demonstrate biosimilarity. Because of the limited clinical experience with biosimilars at the time of their approval, a thorough knowledge surrounding biosimilars and a case-by-case approach are needed to ensure the appropriate use of these products.
Introduction
Muromonab-CD3 (Orthoclone OKT3) was the first monoclonal antibody approved by the US. Food and Drug Administration (FDA) in 1986 for clinical use. Since then, more than 30 therapeutic antibodies and antibody-derivatives have received regulatory approval for the treatment of various diseases.Citation1-Citation5 Like small molecule drugs, biological products also inevitably confront patent expirations and potential threats from competitors. Patents for many biologics have either expired or are going to expire. Thus, the biopharmaceutical market has opened to “generic-like” versions of these products, called biosimilars. Since 2004, the European Medicines Agency’s Committee for Human Medicinal Products has developed three categories of guidelines for biosimilars, including (1) an overarching guideline to define the principle of biosimilar products, (2) general guidelines addressing quality (e.g., manufacturing processes and quality control), non-clinical and clinical issues, and (3) annex guidelines.Citation6-Citation8
Biosimilars are defined as biological medicinal products comparable (but not identical) in quality, safety and efficacy to reference products.Citation6 Compared with generic chemical drugs, biosimilars require a much more extensive assessment for comparability, in which the boundaries of criteria (usually on ad hoc basis) are not usually well-defined due to the complex nature of biologics and their manufacturing process.Citation9,Citation10 Because of the complex post-translational modifications (PTMs) of the glycosylated biomolecules, even a well-controlled product may consist of several hundred or more isoforms with the same amino acid sequence, but different modifications and different batches may exhibit different heterogeneity.Citation11 Thus, to what extent a biosimilar should demonstrate similarity compared with its reference product is currently the most controversial regulatory question. Reports exist that claim there are significant differences in the efficacy and safety profiles of the authorized biosimilar products vs. the reference products, but the direct relationships between those differences and the biophysical characteristics of each drug are not defined.Citation12-Citation27 Whenever possible, various aspects of similarity, including biophysical and clinical results, of biosimilars need to be extensively evaluated and claims of similarity justified on a case-by-case basis.
Etanercept, treatment for moderate to severe plaque psoriasis, psoriatic arthritis, ankylosing spondylitis and moderate to severe rheumatoid arthritis (RA), is a recombinant protein of human soluble tumor necrosis factor receptor 2 (TNFR2) coupled to the Fc portion of human IgG1.Citation28 Enbrel® (etanercept manufactured by Amgen Inc.) is one of the top-selling prescription drugs overall and one of the top-selling biological products on a global basis ($7.4 billion in 2010). Therefore, Enbrel® is an obvious target for biosimilar developers. Although Amgen announced the issuance of a composition of matter patent (US patent number 8,063,182) protecting the branded Enbrel® from etanercept biosimilar competition for another 17 y (through 2028) in the United States, many pharmaceutical companies in other countries have not been discouraged from developing etanercept biosimilars. For example, LG Life Sciences is currently evaluating TNFcept in preclinical studies in South Korea, and Mycenax has completed its Phase 1/2 clinical trial of TuNEX® in Taiwan and is initializing a Phase 3 trial. China’s SFDA has approved two etanercept biosimilar products for the treatment of RA and ankylosing spondylitis; biosimilar 1 has been successfully used to improve the health of RA patients for six years since it was approved for the Chinese market in 2005.
In this study, to elucidate the differences between the products, we characterized and compared the quality attributes of Enbrel®, referred to herein as the reference product, to two commercially available biosimilars from multiple aspects, including primary sequence, peptide mapping, intact mass, charge variants, glycosylations, bioactivity, and affinity.
Results
Amino acid sequences of the reference and biosimilar products
Assessing the molecular similarity of the etanercept biosimilars to the reference product is a critical task during biosimilar development because of its complex molecular structure (934 amino acid) and PTMs. In this study, HPLC-peptide mapping was first used to evaluate identity. As shown in , the peptide mapping of four batches of biosimilar 1 was highly similar to that of the reference product, including the peak number and retention time. The peptide mapping of biosimilar 2 was also similar to that of the reference product, albeit a new peak (between 30–32 min) was found in biosimilar 2 (). A MS/MS analysis was selected to reveal if the difference in peptide mapping resulted from the primary sequence differences.
Figure 1. Comparison of HPLC peptide mapping between the etanercept reference product (Enbrel®) and its biosimilars. (B) was the partial zoom of (A) (26–35min). Line 1 to 7 was blank control, biosimilar 1 (Lot 1), biosimilar 1 (Lot 2), biosimilar 1 (Lot 3), biosimilar 1 (Lot 4), reference product (Wyeth, Lot F12508), and biosimilar 2 (Lot 1), respectively.
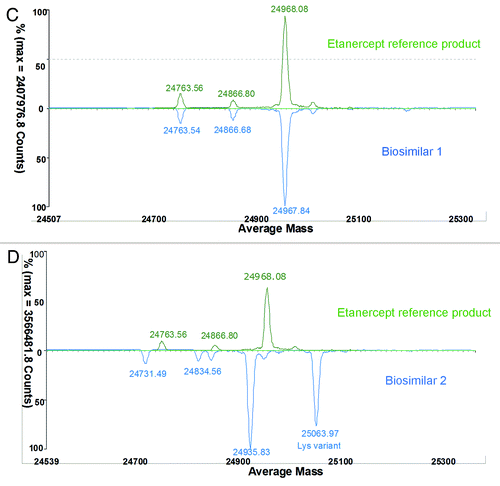
Etanercept is a 150 kDa recombinant dimer protein consisting of two soluble p75 TNFR (sTNFR2) molecules fused to the Fc fragment of human IgG1. It consists of 934 amino acids and each monomer of the dimer contains 467 amino acids. Of the 467 amino acids, 235 amino acids form the extracellular portion of sTNFR2. The remaining 232 amino acids form the Fc domain of human immunoglobulin G1, including CH3, CH2, and the hinge region. In addition to N-glycosylation modification in the Fc’s CH2 region, etanercept has complicated N-linked and O-linked glycosylation in sTNFR2 region (glycosylation covers 1/3 of the whole molecular weight).Citation29-Citation32 To reduce the complexity caused by PTMs, the etanercept protein was digested into two parts (Fc and TNFR2 fragment) using papain treatment. Then, PNGF deglycosylated Fc was analyzed by observing intact mass measurements (Fig. 2). Compared with an Fc sample of the reference product, biosimilar 1 had the same intact mass (49930.1Da/49930.05Da, ), but biosimilar 2 had a -64 Da mass difference for the main peak (49866.47 Da) (). Biosimilar 2 also contained high proportional Lys-variants (1 K and 2 K charge variants). To make further analysis more straightforward, the same Fc samples were then reduced with DTT to remove complexity caused by the disulfide pairs. The accurately measured mass of the reference product (exp. 23968.08 Da) was in accordance with the theoretical value of 23970.34 Da calculated according to the amino acid sequence included for etanercept in the World Health Organization’s international non-proprietary name (INN) list (). Compared with a reduced Fc sample of the reference product, biosimilar 1 had the same intact mass profile (), suggesting the same amino acid sequence information. However, biosimilar 2 had a −32 Da mass difference (half of −64 Da mass difference for intact Fc) for each peak, and also a high proportion of Lys-variant (). These results revealed that two single Fc chains of biosimilar 2 had the same sequence variance or modifications. MS/MS analysis was used to investigate the differences in protein sequence and PTMs between the biosimilar 2 and the reference product.
Figure 2A and B. Comparison of deconvoluted masses of Fc (A and B).
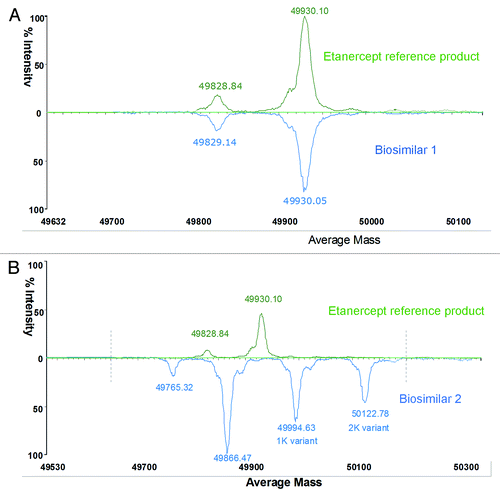
Figure 2C and D. Comparison of reduced Fc (C and D) between the reference product and two etanercept biosimilar products.
An automated data processing and annotation procedure was applied with BiopharmaLynx 1.2 to the entire set of MS/MS data collected. Both the accurate mass of peptide precursors and singly-charged fragment ion spectra were provided for sequence confirmation. Identified peptides were automatically annotated by the application manager with annotation scheme depicting whether they were tryptic peptides from the theoretical sequence (Txx). The total ion chromatogram (TIC) of biosimilar 1 was almost identical with that of the reference product, varying only in peak shapes (). Compared with the TIC chromatogram of the reference product, biosimilar 2 had differences between RT 30–33 min (), and the charge-reduced, isotope-deconvoluted LC-MS chromatogram verified that the two different peptides resulted from the modification or amino acid variations of the peptide T15–16, showing a 32 Da mass difference ().
Figure 3. Mirror plots of LC-MS peptide maps from 4 h tryptic digests of the Fc of the reference product and two etanercept biosimilar products. (A) Comparison of LC-MS (TIC) chromatogram between the reference and biosimilar 1 products; (B) Comparison of LC-MS (TIC) chromatogram between the reference and biosimilar 2 products. (C) A zoom charge-reduced, isotope-deconvoluted LC-MS chromatograms from 22–36 min.
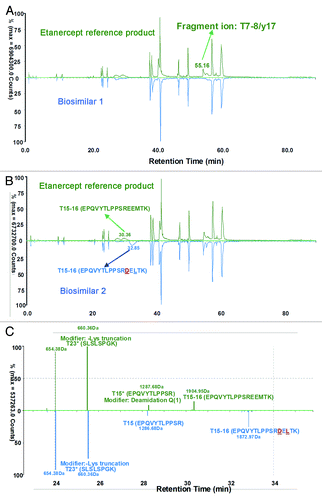
It is interesting that the LC-MS of peptide T15 of biosimilar 2 was in accordance with the reference and biosimilar 1 products (), so the difference must exist in the peptide T16. Analysis revealed that peptide T16 of biosimilar 2 was not identical to that of the reference and biosimilar 1 products (). Conversely, a new mass at RT 5.75 min (), was observed in the biosimilar 2 map; this did not have corresponding matches in the reference product map and the biosimilar 1 map.
Figure 4. Mirror plots of zoom charge-reduced, isotope-deconvoluted LC-MS chromatograms (1–6min) of the FC of reference and two etanercept biosimilar products. (A) Comparison of biosimilar 1 with the reference product; (B) Comparison of biosimilar 2 with the reference product.
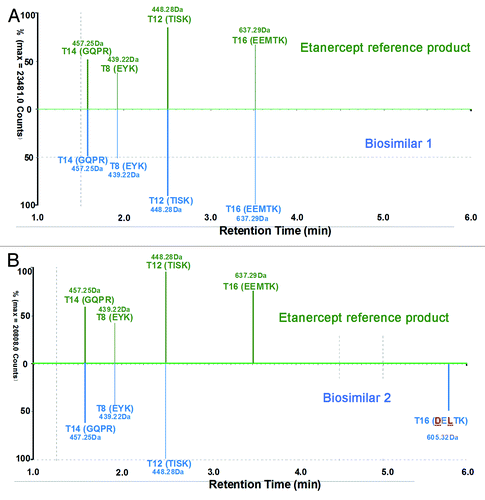
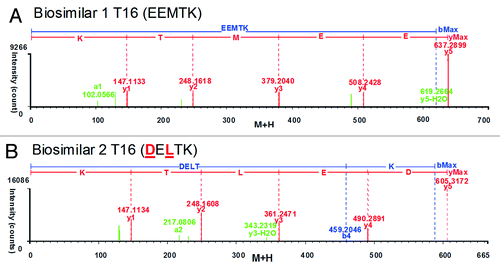
To identify the new mass in the peptide map of biosimilar 2 and uncover its potential relationship with the unmatched T16 peptide of the reference and biosimilar 1 products, we examined three MSE spectra of T16 peptide. The automated assignment of the T16 (EEMTK, RT = 3.51 min, MH+ = 637.29 Da) for the reference and biosimilar 1 products was correct according to the continuously matched fragment ion spectra ( and B); however, there was no matching fragment ion data for this sequence in biosimilar 2. The new unmatched mass T16 in biosimilar 2 had MH+ 605.32 Da (at RT 5.75min), showing a 31.97 Da mass difference from T16 of the reference and biosimilar 1 products (). The mass difference was consistent with the observation of the intact mass measurements. The “EEMTK” sequence of the reference and biosimilar 1 products at peptide T16 was found to be consistent with the sequence data from DrugBank and INN drug information for etanercept,Citation33,Citation34 but the peptide T16 sequence of “DELTK” is included in a patent.Citation35 We found that the sequence “DELTK” exactly has a 32 Da mass difference with “EEMTK,” and we used this sequence variation to find the matched precursor and fragment ion spectra in biosimilar 2. It is interesting that T16 in biosimilar 2 could be confirmed as “DELTK” from the accurate mass of precursor and continuously matched fragment ion spectra (; ).
Figure 5. Charge-reduced, isotope-deconvoluted MSE spectra (fragment ion matched profiles) of tryptic peptide T16. (A) T16 (EEMTK) of biosimilar 1; (B) T16 (DELTK) of biosimilar 2.
Impurity and charge variant analysis
SDS-PAGE purity is often referred to as one of the specifications for etanercept product release. The SDS-PAGE purity specification for the reference product was set to where the content of monomer, high molecular weight components (HMWC), and low molecular weight components (LMWC) must be ≥ 88%, ≤ 6% and ≤ 8%, respectively. As shown in , no obvious difference was found from the SDS-PAGE profiles of the reference and biosimilar products. Compared with the monomers position of the biosimilar 1 and reference products, the monomer of biosimilar 2 was shown to have a relatively low molecular weight (). We thought the molecular weight difference resulted from the sialylation difference because all samples displayed the same electrophoresis positions after the desialylation treatment (). From the SDS-PAGE scanning densitometry results shown in , we found that the reference product had a relatively lower content of HMWC and LMWC impurities.
Figure 6. Non-reduced SDS-PAGE analysis of the reference etanercept product and its biosimilar products before (A) and after (B) desialylation. E1 is the reference product (Enbrel® Lot F12508); Y1, Y2, Y3 and Y4 are 4 lots of biosimilar 1. Q1 is one commercial lot of biosimilar 2.
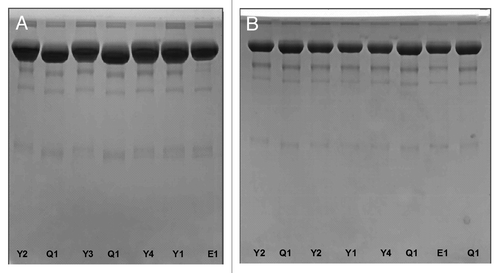
Table 1. TNFα receptor II–Fc fusion proteins*
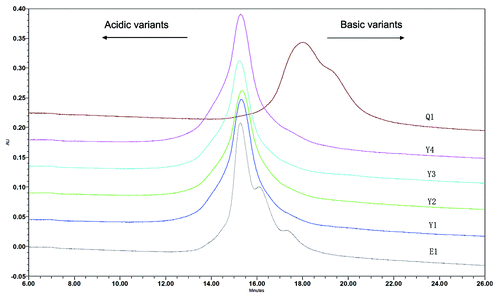
Examples of heterogeneities in mAb or Fc fusion protein products include deamidation, glycosylation, oxidation, aggregation, proteolytic cleavage, and disulfide bond isoforms. These modifications, especially the oxidation, deamidation, and proteolytic cleavage, often result in charge heterogeneities. Ion exchange chromatography (IEX) is frequently used as a tool for analyzing the charge heterogeneity of therapeutic mAbs. To reduce the complexity of the products, sialic acid was removed before IEX separation. Cation exchange chromatography (CEX) analysis showed that the main peak of biosimilar 1 was in accordance with the reference product, but differed in the amount of the basic variants, which corresponds primarily to C-terminal lysine variants from 15–30% in the biosimilar 1 to 40–60% in the reference product (). In fact, the basic variants of the reference product have been shown to vary between pre-manufacturing change and post-manufacturing change batches.Citation11 The main peak of the biosimilar 2, however, eluted much slower than that of either biosimilar 1 or the reference product, showing its relatively basic charge attributes ().
Figure 7. Comparison of IEX charge variant between the reference etanercept product and its biosimilar products (after desialylation). E1 is the reference product (Enbrel® Lot F12508); Y1, Y2, Y3 and Y4 are 4 lots of biosimilar 1. Q1 is one commercial lot of biosimilar 2.
N-glycosylation difference
Etanercept is a dimeric fusion protein that binds TNF. It comprises two extracellular ligand-binding domains of human TNFR2 and the Fc portion of a human IgG1 antibody. The protein contains the IgG1-specific N-glycosylation sites in the Fc portion, and two N-glycosylation sites and multiple O-glycan sites in the receptor portion.Citation32 Particular glycoforms are involved in important immune cytotoxic effector functions, such as cell dependent cytotoxicity and antibody dependent cellular cytotoxicity.Citation36,Citation37 Glycosylation also plays a role in the plasmatic half-life because of its affect on binding of glycoproteins to neonatal Fc gamma receptor (FcγRn).Citation38 Glycans present considerable heterogeneity, with potentially more than 400 glycoforms due to the random pairing of heavy chain glycans of different structures.Citation39 Even minor changes in glycan distribution, such as increased afucosylation, can have effects on the activity of a biopharmaceutical product.Citation40,Citation41
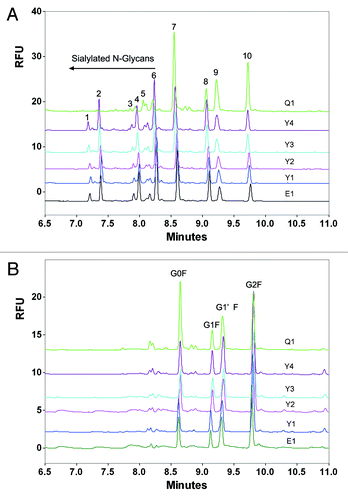
In this study, capillary electrophoresis with laser-induced fluorescence detection (CE-LIF) was used to evaluate the differences in N-glycan profiles between the reference product and its biosimilars. First, N-linked oligosaccharides were removed from the fusion protein samples using the N-glycosidase PNGase F, followed by derivatization of the fluorophore aminopyrene tri-sulfonic acid (APTS) via reductive amination at the reducing end of the oligosaccharide. Electrophoretic separation was conducted utilizing a carbohydrate separation gel buffer. As shown in , the N-glycan profile of the biosimilar 1 was highly similar with the reference product, but that of the bioimilar 2 exhibited major differences in glycoforms and glycan proportion. Both the glycoforms and glycan proportions of the biosimilar 1 and reference products were similar (). After desialylation, the glycoforms of the biosimilar 2 was in accordance with that of the biosimilar 1 and reference products (), although the major difference was found in the glycan proportion (), suggesting that the major difference is the sialylation of the N-oligosaccharides.
Figure 8. Typical CE-LIF electropherogram of released N-oligosaccharides from etanercept reference and biosimilar products before (A) and after desialylation (B). E1 is the reference product (Enbrel® Lot F12508); Y1, Y2, Y3 and Y4 are 4 lots of biosimilar 1. Q1 is one commercial lot of biosimilar 2.
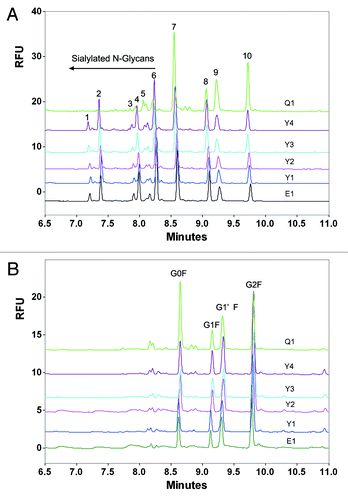
Table 2. Composition percentage of N- Glycans from the innovator Enbrel® and two commercially available biosimilar products
Table 3. Composition percentage of N-oligosaccharides from desialylated TNFα receptor II–Fc fusion proteins*
Affinity and bioactivity analysis
Etanercept has been shown to be effective in the treatment of several autoimmune diseases by reducing TNF bioavailability.Citation42 Affinity analysis based on interactions between TNF and TNF receptor is often used to evaluate the binding properties of the etanercept products with TNF. Biacore systems exploit the phenomenon of surface plasmon resonance to monitor the interaction between molecules in real time. The bivalent etanercept molecule forms a 1:1 complex with the TNF trimer in which two of the three receptor binding sites on TNF are occupied by etanercept, and the third receptor binding site is open.Citation43
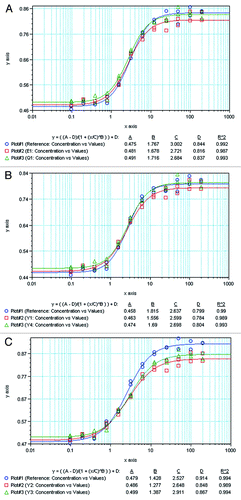
In this study, intrinsic binding affinities of the etanercept products were determined on a surface plasmon resonance instrument. TNF was first immobilized on the surface of a CM-5 sensor chip. Then, analytes (Enbrel® and its biosimilars), in a series of concentrations, were injected sequentially to perform surface-bound analysis to obtain multi-cycle association and dissociation sensorgrams. Then, the multi-cycle curves were fitted with the 1:1 binding model, and the association and dissociation rates (Kon and Koff) were calculated automatically using evaluation software (). As shown in , the determined equilibrium dissociation constant (KD) of the reference product with TNF is 2.083 × 10−11. Biosimilar 1(KD = 2.284 × 10−11, average of 4 lots data) showed about 91.2% of relative affinity compared with the reference product. Biosimilar 2 (KD = 1.316 × 10−11) exhibited an affinity that was ~58.3% higher than the reference product because of its low dissociation rate.
Table 4. Comparison of kinetics fitting results between the etanercept reference product and two commercially available biosimilars*
A TNF-sensitive L929 cell cytotoxicity neutralization assay was used to assess the in vitro bioactivity of the reference product and its biosimilars when bound to TNF. As shown in , the cytotoxicity neutralization experiments of different samples were compared with that of internal reference at each plate. The relative bioactivity could be calculated according to the following formula: Relative bioactivity = CF*ED50(Standard)/ED50(Sample). The relative bioactivity of Y1, Y2, Y3, Y4, Q1 and E1 was 113%, 111%, 96%, 104%, 124%, and 122%, respectively. Interestingly the bioactivity of biosimilar 2 remained comparable with the reference product despite major differences in the primary structure and PTMs.
Figure 9. Four-parameters fitting curve for the potency determination of the etanercept reference and biosimilar products. E1 is the reference product (Enbrel® Lot F12508); Y1, Y2, Y3 and Y4 are 4 lots of biosimilar 1. Q1 is one commercial lot of biosimilar 2.
Discussion
Our study of the structure, physicochemical characteristics, and potency of three marketed etanercept products showed that biosimilar 1 was highly similar to the reference product, but that biosimilar 2 differed in primary structure and glycosylation. The identified sequence variants demonstrate that biosimilar 2 is a different allotype compared with the reference product. A two-amino acid variation occurred in the sequence of the Fc portion in biosimilar 2 (“EEMTK” changed to “DELTK”). By definition, biosimilar products must have the identical amino acid sequence as the reference product to be considered ‘biosimilar’ in Europe.Citation44 In this situation, can we term etanercept biosimilar 2 as a biosimilar of the reference product? Clearly not in Europe, but the definition of biosimilars currently differs between regulatory agencies around the world. If the definition of biosimilars in China or other countries is different from that of Europe, should more comparability studies be conducted to verify the safety and efficacy of products such as biosimilar 2? This remains a challenging question for the regulatory assessment and licensing of such a product.
Another major difference between biosimilar 2 and the reference product was in the sialylation profile. Sialic acid analysis showed that the content of biosimilar 2 was obviously lower than that of the biosimilar 1 and reference products. Sialic acids occupy terminal positions on many mammalian glycoprotein and glycolipid oligosaccharides. Sialylation can significantly influence the safety and efficacy profiles of glycoprotein drugs.Citation45 Therefore, it is important to know the sialic acid content of a glycoprotein when assaying its function or its efficacy as a pharmaceutical therapeutic. When a glycoprotein loses its sialic acid residues, the glycoprotein has reduced serum half-life, and in some cases reduced activity.Citation42 A limited amount of data on half-life is available from the package inserts of the three etanercept products: (1) the half-life of Enbrel® is 102 ± 30 h after administration of 25 mg of Enbrel® by a single subcutaneous (SC) injection to 25 patients; (2) the half-life of biosimilar 1 is 74 ± 4 h after administration of 25 mg of biosimilar 1 by twice weekly SC injection for a period of 6 weeks to 11 RA patients; and (3) the half-life of biosimilar 2 is 60–90 h after a single SC injection of 12.5–25 mg in healthy subjects. From this data, it appears that biosimilar 2 may have a shorter half-life, although most of its range is within that of biosimilar 1 and the reference product.
Despite differences in primary structure and sialylation, in our study biosimilar 2 showed comparable bioactivity with the reference and biosimilar 1 products. This suggests that it may be difficult to define how many comparability studies will be required to demonstrate that similar biological and reference medicinal products have similar profiles in terms of quality, safety and efficacy. The good news is that modern analytical technology, such as peptide mapping with LC-MSE, MALDI-imaging MS, and ESI-Q-TOF-MS, is enabling increasingly comprehensive comparisons between candidate biosimilar and reference products.Citation46,Citation47 The application of analytical technologies will play an important role in enhancing our understanding and reducing some of the risks associated with biopharmaceuticals.Citation48 In this situation, regulators need to be forthcoming with information about their requirements for demonstration of comparability of biosimilars so that more biosimilars can be developed for clinical use.
Materials and Methods
Samples and materials
The commercial etanercept reference product (Enbrel®, Wyeth, Lot No F12508), Biosimilar 1 and Biosimilar 2 were acquired from the Chinese market. NaBHCN (1 M/THF), β-Mercaptoethanol (β-ME), dithiothereitol (DTT), iodoacetic acid (IAA), iodoacetamide (IAM), guanidine hydrochloride (GuHCl), ammonium bicarbonate (NH4HCO3), trypsin, papain, trifluroacetic (TFA), formic acid (FA), peptide N-glycosidase (PNGase F), TNF, and acetonitrile (ACN) were purchased from Sigma Chemical Co. The Carbohydrate Labeling and Analysis Kit were obtained from Beckman. The Amine Coupling kit, Regeneration Scouting Kit, and the CM-5 Sensor Chip were purchased from GE Healthcare
Charge variants analysis
After adding neuraminidase (10 mU/μL, Roche) to form a resulting solution with a concentration of approximately 2.5 mg/mL protein (enzyme/protein ratio is 0.2 mU/μg fusion protein), the sample mixtures were incubated overnight at 37°C in a thermomixer. Desialylated proteins were collected through ultrafiltration (10 KD cut-off of membrane). The desialylated proteins were subsequently separated on a cation exchange chromatographic column (100 × 4.6 mm, 3 μm, 1000 Å, Poly LC). Separations were performed at a 20 min gradient (30–65% B) using 50 mM sodium acetate, pH 5.2 as mobile phase A and 50 mM sodium acetate, 250 mM sodium chloride, pH 5.2 as mobile phase B. We performed UV detection at 214 nm.
HPLC Peptide mapping analysis
Tryptic and alkylated samples were injected onto a Symmetry C18 RP column (250 × 4.6 mm, 5μm). We separated the peaks by applying a linear gradient from 0% to 45% solvent B in 115 min (Solvent A: 0.1% TFA in ddH2O; Solvent B: 0.08% TFA in ACN). The column temperature was 40°C and we performed UV detection at 214 nm.
MS/MS analysis
For the preparation of Fc fragment of the fusion protein, 2 mg/ml fusion protein was treated with papain (1:100 W/W) in a reaction buffer (20 mM PB + 150 mM NaCl +2 mM EDTA + 10 mM Cys pH7.0) at 37°C for 4 h. After the column balance, papain-treated fusion proteins were loaded onto the affinity column (GE, rProtein A 4FF) with a 1 ml/min of flow rate while the non-binding proteins were flushed with PBS. Then, the binding Fc fragments were eluted with 20 mM citric acid (pH 3.0). The collected Fc fragments were then buffer interchanged to a 50 mM NH4HCO3 solution.
The intact and reduced Fc fragment samples were analyzed by reverse LC-MS. A Waters UPLC system was used to generate a gradient flow at 200 µL/min, using typical mobile phases of 0.1% FA in MS-grade water (solvent A) and 0.1% FA in ACN (solvent B). The reverse-phase desalting separation of intact and reduced Fc was performed on a Waters BEH300 C4 Column (2.1 × 50 mm, 1.7 µm) using a 4 min gradient (2–6min, 25–34% B). The column temperature was maintained at 60°C. Mass spectrometric analysis was performed on a Waters Xevo G2 Q-TOFMS system, with a positive ion V mode. The desolvation gas and source temperature were set to 300°C and 120°C, respectively. The capillary and cone voltages were set at 3000 and 40 V, respectively. The m/z scan range was set to 500–3000. The deconvolution of ESI mass spectra of intact and reduced Fc was performed by Mass Biopharmalynx software using Maximum Entropy algorithm.
For the preparation of tryptic digests of etanercept Fc, reduced and alkylated Fc was treated with sequence-grade trypsin (1:20 W/W) at 37°C for 4–16 h. Peptide mixture was separated on a BEH300 C18 column (2.1 × 150 mm, 1.7 µm) at 45°C using a 80 min gradient (0–80 min, 1–36% B). The LC mobile phases and flow rate were the same as in the above intact protein mass analysis. LC-eluted peptides were detected by MS with an alternating low collision energy (6 V) and elevated collision (ramping from 15 to 30 V) ESI+ acquisition data (MSE), respectively. Scan time was 0.5 sec. The capillary and cone voltages were set at 3000 and 35 V, respectively. The source temperature was set to 120°C, and the cone gas flow was set to 50 L/h. An auxiliary pump was used to spray a solution of 100 fmol/µL Glul-fibrinopeptide B in 50/50 ACN/water containing 0.1% FA for mass accuracy (lockmass channel), with a flow rate of 5 µL/min and sampling every 1 min.
The collected LC-MS/MS data was processed using Biopharmalynx software. We can set the fixed modification (alkylation) and variable modification (e.g., oxidation, deamidation, glycosylation) while searching the protein digest library. The mass tolerance for both precursors and fragments was set to less than 30 ppm. The identified peptides were confirmed by MSE spectra with at least three continuously matched b/y fragment ions.
N-Glycosylation analysis
Under denaturation conditions, the desialylated or non-desialylated samples were deglycosylated with PNGase F for 20–24 h at 37°C. After cold ethanol precipitation of deglycoslated samples, the mixture was centrifuged at 14,000 × g for 10 min. The supernatant containing the released oligosaccharides was transferred to a new sample tube and evaporated to dryness. Dried oligosaccharide sample were labeled with 2 μL of 1 M NaBHCN/THF and 2 μL of APTS Labeling Reagent for 4 h at 37°C, and then diluted to 50 μl with Milli-Q water. CE-LIF analysis was performed on Beckman PA800 PLUS system using an N-CHO capillary (50 µm I.D., 40 cm effective length, 50.2 cm total length, Beckman Coulter) in a Carbohydrate Separation Buffer. Prior to sample injection, the capillary was rinsed with carbohydrate separation buffer for 3 min. Then samples were injected for three seconds at 0.5 psi. Separation was conducted in the negative polarity mode (−30 kV) at 20°C for 20 min. LIF detection was conducted at 488nm of excitation and 520 nm of emission wavelength.
Potency determination
A TNF-sensitive L929 bioassay was used to assess the bioactivity of the reference product and its biosimilars when bound to TNF. 8 x 104 cells/well of L929 cells were incubated in 100 μL RPMI-1640/DMEM supplemented with 10% NBS in a 96-well tissue culture plate and cultured overnight (18–24 h) at 37°C, 8% CO2. 100 μL of prepared serially diluted test sample was added to each well of a 96-well tissue culture plate. After 18–22 h of incubation at 37°C, 20 μL MTS/PMS (Promega) solution was added before another 2–4 h of incubation. The absorbance was read and recorded at 490 and 630 nm. The ED50 value was obtained by logistics four parameter curve fit.
Affinity analysis
The affinity analysis of the reference product and its biosimilars were performed using a Biacore T200. TNF protein solution at a concentration of 10 µg/mL (in 10 mM Acetate, pH 5.0) was amine coupled on CM5 chip according to the automated immobilization procedure, including the pre-concentration, activation, injection, immobilization, and blocking, and the target response value was set as 800 RU. First, 0.039 to 2.5 µg/mL gradient of the analytes (etanercept proteins) were acquired by serial 2-fold HBS-EP buffer dilution. Next, two vials of HBS-EP buffer and one vial of 4 M MgCl2 were prepared for blank test, startup and regeneration steps separately. Then, other parameters of affinity/kinetics assay were set, including injection time 180 sec, dissociation time 1800 sec, regeneration time 60 sec at flow rate 30 µL/min after each injection and two startups. At last, two parallel runs were performed for all samples.
Using global fit analysis with Bia-evaluation 4.0.1 software for kinetic analyses, rate equations derived from the 1:1 Langmuir binding model were fitted to the experimentally obtained association-phase and dissociation-phase binding curves for all concentration-gradient injections. Kinetic rate constants for association rate constant (Kon) and dissociation rate constant (Koff) were determined, and the equilibrium dissociation rate constant (KD) was calculated with the formula: KD = Koff/Kon.
| Abbreviations: | ||
| Fab | = | fragment antigen binding region |
| Fc | = | fragment crystallizable region |
| MS | = | mass spectrometry |
| MS/MS | = | tandem mass spectrometry |
| MALDI | = | Matrix-assisted laser desorption/ionization |
| ESI-Q-TOF | = | electrospray ionization-time-of-flight |
| ESI | = | electrospray ionization |
| IEX | = | ion exchange chromatography |
| CE-LIF | = | capillary electrophoresis with laser induced fluorescence detector |
| IAA | = | Iodoacetic acid |
| IAM | = | Iodoacetamide |
| DTT | = | dithiothereitol |
| β-ME | = | β-mercaptoethanol |
| TFA | = | trifluroacetic |
| FA | = | formic acid |
| PNGase F | = | peptide N-glycosidase |
| CAN | = | acetonitrile |
| SEC | = | size exclusion chromatography |
| ED50 | = | 50% effective dose |
| PTMs | = | post-translational modifications |
Acknowledgments
This work was supported by grants from National Natural Science Foundation of China, Ministry of Science and Technology of China (973 and 863 program projects), State Key Project for New Drug Development (2012ZX09304008; 2010ZX09401-407), Ministry of Education (Key Laboratory), Shanghai Commission of Science and Technology (Key Laboratory and Projects), and Shanghai Leading Academic Discipline Project (B905).
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
References
- Jenkins N, Parekh RB, James DC. Getting the glycosylation right: implications for the biotechnology industry. Nat Biotechnol 1996; 14:975 - 81; http://dx.doi.org/10.1038/nbt0896-975; PMID: 9631034
- Reichert JM, Rosensweig CJ, Faden LB, Dewitz MC. Monoclonal antibody successes in the clinic. Nat Biotechnol 2005; 23:1073 - 8; http://dx.doi.org/10.1038/nbt0905-1073; PMID: 16151394
- Beck A, Wagner-Rousset E, Bussat MC, Lokteff M, Klinguer-Hamour C, Haeuw JF, et al. Trends in glycosylation, glycoanalysis and glycoengineering of therapeutic antibodies and Fc-fusion proteins. Curr Pharm Biotechnol 2008; 9:482 - 501; http://dx.doi.org/10.2174/138920108786786411; PMID: 19075687
- Reichert JM. Antibodies to watch in 2010. MAbs 2010; 2:84 - 100; http://dx.doi.org/10.4161/mabs.2.1.10677; PMID: 20065640
- Oldham RK, Dillman RO. Monoclonal antibodies in cancer therapy: 25 years of progress. J Clin Oncol 2008; 26:1774 - 7; http://dx.doi.org/10.1200/JCO.2007.15.7438; PMID: 18398141
- European Medicines Agency. Guideline on similar biological medicinal products. CHMP/437/04. 2005. www.ema.europa.eu/pdfs/human/biosimilar/043704en.pdf
- European Medicines Agency. Guideline on similar biological medicinal products containing biotechnology-derived proteins as active substance: quality issues. EMEA/CHMP/BWP/49348/2005. 2006. http://www.emea.europa.eu/docs/en_GB/document_library/Scientific_guideline/2009/09/WC500003953.pdf
- European Medicines Agency. Guideline on similar biological medicinal products containing biotechnology-derived proteins as active substance: non-clinical and clinical issues. EMEA/CHMP/BMWP/42832/2005. 2006. http://www.ema.europa.eu/docs/en_GB/document_library/Scientific_guideline/2009/09/WC500003920.pdf
- Gottlieb S. Biosimilars: policy, clinical, and regulatory considerations. Am J Health Syst Pharm 2008; 65:Suppl 6 S2 - 8; http://dx.doi.org/10.2146/ajhp080210; PMID: 18591712
- Ahmed I, Kaspar B, Sharma U. Biosimilars: impact of biologic product life cycle and European experience on the regulatory trajectory in the United States. Clin Ther 2012; 34:400 - 19; http://dx.doi.org/10.1016/j.clinthera.2011.12.005; PMID: 22244050
- Schiestl M, Stangler T, Torella C, Cepeljnik T, Toll H, Grau R. Acceptable changes in quality attributes of glycosylated biopharmaceuticals. Nat Biotechnol 2011; 29:310 - 2; http://dx.doi.org/10.1038/nbt.1839; PMID: 21478841
- Stanhope R, Sörgel F, Gravel P, Pannatier Schuetz YB, Zabransky M, Muenzberg M. Bioequivalence studies of omnitrope, the first biosimilar/rhGH follow-on protein: two comparative phase 1 randomized studies and population pharmacokinetic analysis. J Clin Pharmacol 2010; 50:1339 - 48; http://dx.doi.org/10.1177/0091270009359005; PMID: 20173088
- Romer T, Saenger P, Peter F, Walczak M, Le Bouc Y, Khan-Boluki J, et al. Seven years of safety and efficacy of the recombinant human growth hormone Omnitrope in the treatment of growth hormone deficient children: results of a phase III study. Horm Res 2009; 72:359 - 69; http://dx.doi.org/10.1159/000249164; PMID: 19844125
- Pavlovic M, Girardin E, Kapetanovic L, Ho K, Trouvin JH. Similar biological medicinal products containing recombinant human growth hormone: European regulation. Horm Res 2008; 69:14 - 21; http://dx.doi.org/10.1159/000111790; PMID: 18059081
- Romer T, Peter F, Saenger P, Starzyk J, Koehler B, Korman E, et al. Efficacy and safety of a new ready-to-use recombinant human growth hormone solution. J Endocrinol Invest 2007; 30:578 - 89; PMID: 17848841
- Peterkova V, Arslanoglu I, Bolshova-Zubkovskaya E, Romer T, Zdravkovic D, Kratzsch J, et al. A randomized, double-blind study to assess the efficacy and safety of valtropin, a biosimilar growth hormone, in children with growth hormone deficiency. Horm Res 2007; 68:288 - 93; http://dx.doi.org/10.1159/000105494; PMID: 17627092
- Skrlin A, Radic I, Vuletic M, Schwinke D, Runac D, Kusalic T, et al. Comparison of the physicochemical properties of a biosimilar filgrastim with those of reference filgrastim. Biologicals 2010; 38:557 - 66; http://dx.doi.org/10.1016/j.biologicals.2010.05.002; PMID: 20637652
- Waller CF, Bronchud M, Mair S, Challand R. Pharmacokinetic profiles of a biosimilar filgrastim and Amgen filgrastim: results from a randomized, phase I trial. Ann Hematol 2010; 89:927 - 33; http://dx.doi.org/10.1007/s00277-010-0961-x; PMID: 20428872
- Waller CF, Bronchud M, Mair S, Challand R. Comparison of the pharmacodynamic profiles of a biosimilar filgrastim and Amgen filgrastim: results from a randomized, phase I trial. Ann Hematol 2010; 89:971 - 8; http://dx.doi.org/10.1007/s00277-010-0973-6; PMID: 20567823
- Waller CF, Semiglazov VF, Tjulandin S, Bentsion D, Chan S, Challand R. A phase III randomized equivalence study of biosimilar filgrastim versus Amgen filgrastim in patients receiving myelosuppressive chemotherapy for breast cancer. Onkologie 2010; 33:504 - 11; http://dx.doi.org/10.1159/000319693; PMID: 20926897
- Sörgel F, Lerch H, Lauber T. Physicochemical and biologic comparability of a biosimilar granulocyte colony-stimulating factor with its reference product. BioDrugs 2010; 24:347 - 57; http://dx.doi.org/10.2165/11585100-000000000-00000; PMID: 20873878
- Gascon P, Fuhr U, Sörgel F, Kinzig-Schippers M, Makhson A, Balser S, et al. Development of a new G-CSF product based on biosimilarity assessment. Ann Oncol 2010; 21:1419 - 29; http://dx.doi.org/10.1093/annonc/mdp574; PMID: 20019087
- Lubenau H, Sveikata A, Gumbrevicius G, Macijauskiene J, Fokas V, Kazlauskas S, et al. Bioequivalence of two recombinant granulocyte colony-stimulating factor products after subcutaneous injection in healthy volunteers. Int J Clin Pharmacol Ther 2009; 47:275 - 82; PMID: 19356394
- Lubenau H, Bias P, Maly AK, Siegler KE, Mehltretter K. Pharmacokinetic and pharmacodynamic profile of new biosimilar filgrastim XM02 equivalent to marketed filgrastim Neupogen: single-blind, randomized, crossover trial. BioDrugs 2009; 23:43 - 51; http://dx.doi.org/10.2165/00063030-200923010-00005; PMID: 19344191
- Haag-Weber M, Vetter A, Thyroff-Friesinger U, INJ-Study Group. Therapeutic equivalence, long-term efficacy and safety of HX575 in the treatment of anemia in chronic renal failure patients receiving hemodialysis. Clin Nephrol 2009; 72:380 - 90; PMID: 19863881
- Krivoshiev S, Todorov VV, Manitius J, Czekalski S, Scigalla P, Koytchev R, Epoetin Zeta Study Group. Comparison of the therapeutic effects of epoetin zeta and epoetin alpha in the correction of renal anaemia. Curr Med Res Opin 2008; 24:1407 - 15; http://dx.doi.org/10.1185/030079908X297402; PMID: 18394266
- Wizemann V, Rutkowski B, Baldamus C, Scigalla P, Koytchev R, Epoetin Zeta Study Group. Comparison of the therapeutic effects of epoetin zeta to epoetin alfa in the maintenance phase of renal anaemia treatment. Curr Med Res Opin 2008; 24:625 - 37; http://dx.doi.org/10.1185/030079908X273264; PMID: 18208642
- Grattendick KJ, Nakashima JM, Feng L, Giri SN, Margolin SB. Effects of three anti-TNF-α drugs: etanercept, infliximab and pirfenidone on release of TNF-α in medium and TNF-α associated with the cell in vitro. Int Immunopharmacol 2008; 8:679 - 87; http://dx.doi.org/10.1016/j.intimp.2008.01.013; PMID: 18387510
- Peppel K, Crawford D, Beutler B. A tumor necrosis factor (TNF) receptor-IgG heavy chain chimeric protein as a bivalent antagonist of TNF activity. J Exp Med 1991; 174:1483 - 9; http://dx.doi.org/10.1084/jem.174.6.1483; PMID: 1660525
- Peppel K, Poltorak A, Melhado I, Jirik F, Beutler B. Expression of a TNF inhibitor in transgenic mice. J Immunol 1993; 151:5699 - 703; PMID: 7693816
- Kolls J, Peppel K, Silva M, Beutler B. Prolonged and effective blockade of tumor necrosis factor activity through adenovirus-mediated gene transfer. Proc Natl Acad Sci U S A 1994; 91:215 - 9; http://dx.doi.org/10.1073/pnas.91.1.215; PMID: 8278368
- Tracey D, Klareskog L, Sasso EH, Salfeld JG, Tak PP. Tumor necrosis factor antagonist mechanisms of action: a comprehensive review. Pharmacol Ther 2008; 117:244 - 79; http://dx.doi.org/10.1016/j.pharmthera.2007.10.001; PMID: 18155297
- http://www.drugbank.ca/drugs/DB00005.
- RECOMMENDED INN. List 43. WHO Drug Information 2000; 14:51
- U.S. Patent number: 5605690.
- Raju TS. Terminal sugars of Fc glycans influence antibody effector functions of IgGs. Curr Opin Immunol 2008; 20:471 - 8; http://dx.doi.org/10.1016/j.coi.2008.06.007; PMID: 18606225
- Nimmerjahn F, Ravetch JV. The antiinflammatory activity of IgG: the intravenous IgG paradox. J Exp Med 2007; 204:11 - 5; http://dx.doi.org/10.1084/jem.20061788; PMID: 17227911
- Magdelaine-Beuzelin C, Kaas Q, Wehbi V, Ohresser M, Jefferis R, Lefranc MP, et al. Structure-function relationships of the variable domains of monoclonal antibodies approved for cancer treatment. Crit Rev Oncol Hematol 2007; 64:210 - 25; http://dx.doi.org/10.1016/j.critrevonc.2007.04.011; PMID: 17624800
- Walsh G, Jefferis R. Post-translational modifications in the context of therapeutic proteins. Nat Biotechnol 2006; 24:1241 - 52; http://dx.doi.org/10.1038/nbt1252; PMID: 17033665
- Hancock WS. Glycosylation, who cares?. J Proteome Res 2002; 1:297; http://dx.doi.org/10.1021/pr0203363; PMID: 12645884
- Jefferis R. Glycosylation of recombinant antibody therapeutics. Biotechnol Prog 2005; 21:11 - 6; http://dx.doi.org/10.1021/bp040016j; PMID: 15903235
- Grattendick KJ, Nakashima JM, Feng L, Giri SN, Margolin SB. Effects of three anti-TNF-alpha drugs: etanercept, infliximab and pirfenidone on release of TNF-alpha in medium and TNF-alpha associated with the cell in vitro. Int Immunopharmacol 2008; 8:679 - 87; http://dx.doi.org/10.1016/j.intimp.2008.01.013; PMID: 18387510
- Scallon B, Cai A, Solowski N, Rosenberg A, Song XY, Shealy D, et al. Binding and functional comparisons of two types of tumor necrosis factor antagonists. J Pharmacol Exp Ther 2002; 301:418 - 26; http://dx.doi.org/10.1124/jpet.301.2.418; PMID: 11961039
- Reichert JM, Beck A, Iyer H. European Medicines Agency workshop on biosimilar monoclonal antibodies: July 2, 2009, London, UK. MAbs 2009; 1:394 - 416; http://dx.doi.org/10.4161/mabs.1.5.9630; PMID: 20065643
- Takeuchi M, Inoue N, Strickland TW, Kubota M, Wada M, Shimizu R, et al. Relationship between sugar chain structure and biological activity of recombinant human erythropoietin produced in Chinese hamster ovary cells. Proc Natl Acad Sci U S A 1989; 86:7819 - 22; http://dx.doi.org/10.1073/pnas.86.20.7819; PMID: 2813359
- Xie H, Chakraborty A, Ahn J, Yu YQ, Dakshinamoorthy DP, Gilar M, et al. Rapid comparison of a candidate biosimilar to an innovator monoclonal antibody with advanced liquid chromatography and mass spectrometry technologies. MAbs 2010; 2:379 - 94; PMID: 20458189
- Beck A, Sanglier-Cianférani S, Van Dorsselaer A. Biosimilar, biobetter, and next generation antibody characterization by mass spectrometry. Anal Chem 2012; 84:4637 - 46; http://dx.doi.org/10.1021/ac3002885; PMID: 22510259
- Berkowitz SA, Engen JR, Mazzeo JR, Jones GB. Analytical tools for characterizing biopharmaceuticals and the implications for biosimilars. Nat Rev Drug Discov 2012; 11:527 - 40; http://dx.doi.org/10.1038/nrd3746; PMID: 22743980