
Abstract
The nucleus is a cellular compartment that hosts several macro-molecular machines displaying a highly complex spatial organization. This tight architectural orchestration determines not only DNA replication and repair but also regulates gene expression. In budding yeast microtubules play a key role in structuring the nucleus since they condition the Rabl arrangement in G1 and chromosome partitioning during mitosis through their attachment to centromeres via the kinetochore proteins. Recently, we have shown that upon quiescence entry, intranuclear microtubules emanating from the spindle pole body elongate to form a highly stable bundle that spans the entire nucleus. Here, we examine some molecular mechanisms that may underlie the formation of this structure. As the intranuclear microtubule bundle causes a profound re-organization of the yeast nucleus and is required for cell survival during quiescence, we discuss the possibility that the assembly of such a structure participates in quiescence establishment.
Introduction
Cells perpetually face the decision to proliferate or to enter a non-dividing state. Non-dividing states may be irreversible, such as senescence, or only temporary. Quiescence, a state defined as a reversible arrest of proliferation, is probably the most common cellular situation found on earth, as it concerns a wide range of cells, from microbes to human stem cells.Citation1-Citation3 Quiescent cells are confronted with quite a few challenges. On the one hand, quiescent cells need to preserve their ability to proliferate, sometimes over several years.Citation4 As they age, quiescent cells must cope with extrinsic or intrinsic harmful events that cause the accumulation of damaged macromolecules. The inability to handle these stresses can ultimately lead to cell death. Additionally, for the sake of tissues, organisms, and the species itself, quiescent cells must produce offspring born “damage-free.” On the other hand, cells must enter quiescence and return into the proliferation cycle in a tightly regulated manner, uncontrolled transitions being potentially deleterious for the whole organism, as exemplified by stem cell depletion or cancer.Citation1,Citation5-Citation7 Finally, in the case of microorganisms competing for their environmental niche, quiescence exit must be swift to ensure the prevalence of the species. Therefore, quiescence is at the heart of crucial biological issues including development, aging, and evolution.
Quiescence encompasses highly diverse physiological situations that differ not only depending on the cell type but also, in the case of the same type of cells, depending on both the stimuli that triggered quiescence entry (contact inhibition, exhaustion of growth factors or nutrients, hypoxia, or other) and on the time spent in quiescence.Citation2,Citation8,Citation9 In single cell eukaryotes like S. cerevisiae, quiescence is primarily induced by the limitation of essential nutrients.Citation4,Citation10 When the carbon source is exhausted, most cells—but not allCitation11-Citation13—enter quiescence from the G1 phase of the cell cycle.Citation14,Citation15 Quiescent yeast cells are not simply resting G1 cells as they acquire a variety of specific properties, including a gene expression profile that differs significantly from the one displayed by G1 arrested cells.Citation16,Citation17 In addition, the establishment of quiescence in yeast is accompanied by a decrease in protein synthesis rate, the accumulation of storage molecules,Citation4,Citation10 and the reorganization of various cellular machinery. Indeed, upon quiescence entry, the mitochondria tubular network fragments,Citation18 the proteasome exits the nucleus and joins cytoplasmic structures called proteasome storage granules,Citation19 actin filaments collapse into actin bodies,Citation20 and various chaperonesCitation21,Citation22 and metabolic enzymes are re-localized.Citation23,Citation24 Some of these reorganizations have been described in both S. cerevisiae and S. pombe,Citation19 but also in metazoans, suggesting that they may be conserved among eukaryotes.Citation24,Citation25 The assembly of quiescence specific structures implies a cell’s commitment to this specific cellular state.Citation13 As some of these structures have been shown to be required for survival and/or exit from quiescence, one key question is to now decipher whether the observed reorganizations are a passive consequence of the cell’s entry into quiescence or if they actively participate in the process of quiescence establishment.
The Nucleus is Drastically Reorganized upon Quiescence Establishment
In the G1 phase of the cell cycle, the budding yeast nucleus is arranged in a Rabl configuration ().Citation26 Each centromere is attached via its kinetochore complex to a short nuclear microtubule (<300 nm) that emanates from the spindle pole body (SPB, the yeast equivalent of the centrosome).Citation27-Citation29 While centromeres are highly clustered next to the SPB, most of the telomeres are localized into 8–10 foci distributed over the nuclear periphery. The nucleolus, built around the rDNA, is opposed to the SPB on the other half of the nucleus.Citation30,Citation31 This typical organization strictly depends on nuclear microtubules. Indeed, it was shown in vivo that microtubule depolymerization causes centromere declustering,Citation32 and in silico modeling predicts that, in the absence of microtubules, the rDNA would fail to position opposite to the SPB.Citation33 Besides, telomere cluster formation and distribution at the nuclear periphery was shown to be influenced by the amount of the silencing protein Sir3.Citation34 Therefore, the nuclear architecture is highly intricate and its compartmentalization tightly controlled.
Figure 1. Schematic model of the nuclear reorganizations occurring upon transition between proliferation and quiescence. The nuclear membrane is in gray, the SPB in blue, and the nucleolus in purple. Microtubules are represented by green lines, the centromeres by red spheres, and the telomeres by yellow parallelepipeds. For simplicity, the chromosome arms are not shown.
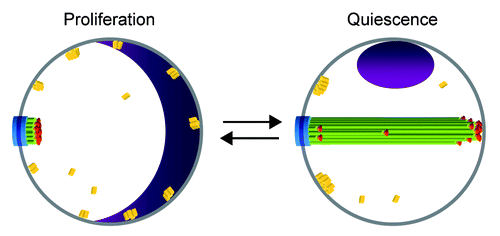
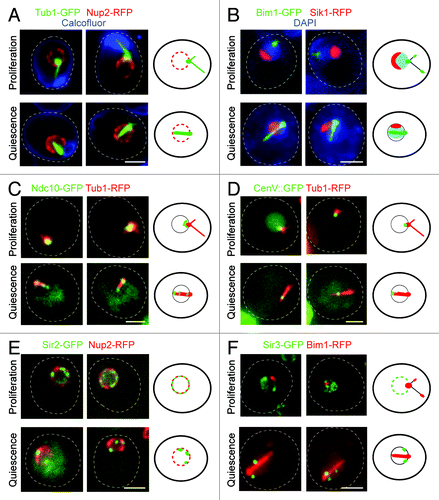
Upon quiescence entry, we have shown that the S. cerevisiae nucleus is drastically reorganized ().Citation35 SPB-associated nuclear microtubules elongate to form an extended and extremely stable bundle that spans the entire nucleus (). Because centromeres remain attached to the microtubule ends, they are mostly found at the tip of the microtubule bundle, opposite to the SPB (). Yet, as microtubules within the bundle are not all of the same length, centromeres can occasionally be found close to the SPB or along the length of the microtubule array (). The nucleolus is displaced to one side of the microtubule array, often close to the SPB (). Of note, the size of the nucleolus is greatly diminished (), in agreement with the drastic decrease in ribosome synthesis and translation rate in quiescent cells.Citation4,Citation10 In mutants that do not assemble a microtubule array in quiescence, the centromeres and the nucleolus are no longer displaced, demonstrating that microtubule elongation and stabilization in quiescence are causing the nuclear rearrangements.Citation35 We have also found that the silencing proteins Sir2 and Sir3 are localized into 2 to 3 foci at the nuclear periphery of quiescent cells, probably reflecting an hyper-clustering of telomeres (D.L. and I.S., unpublished results; ). Interestingly, these foci are often localized next to the SPB (D.L. and I.S., unpublished results; ). The existence of a possible interplay between microtubules and telomere rearrangement remain to be investigated. Yet, it is worth noting that in early prophase of meiosis I, the yeast nucleus adopts an organization called the “bouquet” in which, like in quiescent cells, the telomeres gather together close to the SPB.Citation35,Citation36 Does the telomere congregation next to the SPB observed in quiescence and meiosis involve similar molecular processes? In the bouquet arrangement, centromeres are no longer found in proximity to the SPB like they are in G1, but rather are scattered throughout the nucleoplasm.Citation35 Intriguingly, upon transition from the Rabl configuration to the bouquet arrangement, the assembly of a short-lived intranuclear microtubule bundle was reported.Citation37 Concomitantly, centromeres are found transiently arranged along “a line,” but whether the lining up of the centromeres takes place along the microtubule bundle has not been investigated. Ultimately, the microtubule structure disappears, and, at the pachytene stage, centromeres are unleashed into the nucleoplasm.Citation37 One interesting possibility is that the microtubule bundle observed in meiosis may be comparable to the structure assembled in quiescent cells. Another intriguing analogy between quiescence and meiosis is that both processes are induced by nutrient starvation. Are the signals and the mechanisms responsible for microtubule array formation similar for both cellular situations? The discovery of molecular congruence between these two nuclear reorganizations would be striking.
Figure 2. (A) Wild type cells expressing Tub1-GFP (microtubules, green) and Nup2-RFP (nuclear membrane, red) stained with calcofluor (cell wall, blue). (B) Wild type cells expressing Bim1-GFP (green) and Sik1-RFP (nucleolus, red) stained with DAPI (DNA, blue). (C) Wild type cells expressing Ndc10-GFP (kinetochore, green) and Tub1-RFP (microtubules, red). (D) Wild type cells in which LacO sequences have been integrated within the pericentromeric region of chromosome V and expressing LacI-GFP (centromere, green) and Tub1-RFP (microtubules, red). (E) Wild type cells expressing Sir2-GFP (green) and Nup2-RFP (nuclear membrane, red). (F) Wild type cells expressing Sir3-GFP (green) and Bim1-RFP (red). For each section cells in the upper panel are in G1 phase of the proliferating cycle, and cells in the lower panel are in quiescence (7 d). A schematic representation is shown on the right of each panel; scale bar is 2 microns.
Molecular Mechanisms Underlying Microtubule Array Formation in Quiescent Cells
Microtubules are composed of α- and β-tubulin heterodimers that are assembled after a nucleation step typically requiring the gamma tubulin complex. In proliferating yeast cells, the gamma tubulin complex is embedded into the SPB, and while the microtubule “minus end” stays closely associated with this structure, the opposite end, the “plus end,” is highly dynamic and alternates between periods of growth, pause, and shrinkage. In yeast, like in other eukaryotes, microtubule dynamic is influenced by a plethora of microtubule-associated proteins (MAPs) that can either stabilize or destabilize microtubules.Citation38-Citation41
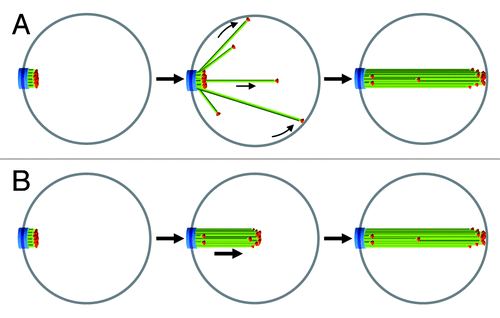
Quiescent cells assemble an array of microtubules that spans the entire nucleus, sometimes even causing the deformation of the nuclear membrane.Citation35 The net elongation of microtubules could result from an increased polymerization rate, a decreased depolymerization rate, a diminished catastrophe frequency, an enhanced rescue frequency, or a combination of all these events. Upon quiescence entry microtubules may grow from the SPB in various directions until they reach the nuclear membrane. Due to the spatial constraint imposed by this physical barrier, growing microtubules would slide along the membrane toward the opposite side of the nucleus, where they would reach their “maximum” length (). Alternatively, packing forces due to microtubule bundling proteins or to interaction between neighboring kinetochores may result in multiple microtubules simultaneously elongating in front of the SPB in a constrained manner ().
Figure 3. Models for the formation of the nuclear microtubule array upon quiescence establishment. In (A) microtubules elongate from the SPB in all directions and reach a maximum size at the nuclear membrane opposite of the SPB. In (B) microtubules are maintained together as they elongate. Legends are the same as in . See text for details.
In quiescence nuclear microtubules are not only long but amazingly stable. This stability could be actively triggered by enhanced microtubule stabilization activity and/or may be the consequence of the depletion of microtubule destabilizing proteins. We have found that a large number of MAPs that influence microtubule dynamics are not required for the formation and the stability of microtubule array, even though some of these MAPs were clearly detected along the microtubule bundle.Citation35 This is the case for Bim1, the EB1 homolog known to decrease the catastrophe frequency.Citation42 Astonishingly, we have found that dynein, a microtubule minus-end-directed motor and its regulator, the dynactin complex, are required for the microtubule array formation. Yet, in quiescent cells, none of these proteins could be detected in the nucleus but were rather localized on the cytoplasmic face of the SPB.Citation35 If these MAPs are indeed absent in the nucleus, how could they affect the formation of a nuclear microtubule structure? In budding yeast, the SPB traverses the nuclear envelope.Citation40 Thus, one possibility could be that deletion of cytoplasmic proteins, such as dynein, alters the composition and/or architecture of the SPB, which causes its incapacity to nucleate (or stabilize) microtubules inside the nucleus. In S. pombe it was suggested that dynein deletion may impede on SPB integrity.Citation43,Citation44 Whether the SPB is affected in dynein and dynactin mutants in quiescent S. cerevisiae is not known.
At the other end of the bundle, the plus ends of microtubules may be “capped” by proteins, thereby preventing their depolymerization. One obvious candidate is the kinetochore complex. In proliferating cells it was shown that microtubules that are attached to kinetochore are more stable than unattached ones.Citation45-Citation47 In quiescence the microtubule/kinetochore attachment could be locked, leading to an enhanced kinetochore microtubule-stabilizing activity and the “freezing” of microtubule plus end. A peculiar observation is the localization of members of the Dam complex, a yeast kinetochore sub-module, all along the microtubule bundle. In vitro, the Dam complex forms a ring around the microtubule.Citation48-Citation51 Could the Dam complex, by forming collars all along the microtubules participate in microtubule stabilization and/or bundling in quiescent cells? Deciphering the roles of each kinetochore sub complex will likely shed light on the molecular mechanisms driving the formation of the atypically stable bundle of parallel microtubules assembled in quiescent yeast cells.
The Nuclear Microtubule Bundle May Influence Gene Expression and Thereby be Required for Quiescence Establishment
Whatever the molecular mechanisms involved in the formation of the microtubule bundle in the nucleus of quiescent cells, one key question is: what is the physiological “raison d’être” of this structure? Does it play a crucial role in quiescence establishment? Is this structure needed for the fitness during quiescence exit? We have found that mutants unable to assemble a microtubule bundle in quiescence are impaired for quiescence survival.Citation35 One intriguing possibility could be that the chromosome rearrangement imposed by the formation of the microtubule array contributes to the specific gene expression profile adopted by the cells upon quiescence entry. Indeed, although chromosomes display confined movement in interphase,Citation52 it was proposed that the localization of a gene within the nucleus correlates with its transcriptional activity. In the current model, genes expressed at a basal level preferentially localize toward the center of the nucleus while actively silenced chromosome regions are associated with the nuclear membrane. Massively transcribed genes would be located rather close to nuclear pores, a proximity that is supposed to improve the export efficiency of the newly synthesized mRNA (for a review see ref. Citation30 and refs. herein). By inducing the displacement of most centromeres and the relocalization of the nucleolus, the microtubule array would cause severe variation in gene position within the nuclear compartment and thereby modify the transcriptional profile of the cell. Therefore, by modifying gene expression, the nuclear microtubule array may be needed for quiescence establishment. Mutants that are not able to assemble this structure would fail in modifying their transcriptome and, consequently, would be incapable of installing quiescence. This would explain why such mutants have a reduced viability in this non-proliferating cellular state. Interestingly, drastic and reversible chromosome displacements have been reported upon proliferation/quiescence transitions in primary human fibroblasts.Citation53 Like in yeast, the nuclear micro-environment influences gene regulation in mammals.Citation54 Therefore, chromosome re-localization may also be involved in quiescence establishment in metazoan cells.
Microtubule and/or Centromere Attachment and the Fitness of Quiescence Exit
When yeast cells proliferate, centromeres stay connected to the microtubule plus end throughout cell cycle progression. In fact, the centromere-microtubule attachment is lost for only a few minutes, which is just the time needed to replicate centromeric DNA.Citation55-Citation57 In mitosis, a single unattached centromere can activate the spindle assembly checkpoint and delay cell division until the defect is solved.Citation58,Citation59 In quiescent cells centromeres are mostly localized at the tip of the microtubule bundle. When quiescence exit is triggered by nutrient addition, the nuclear microtubule array slowly shortens, and about an hour after re-feeding, cells have recovered a typical Rabl configuration. During this entire process, microtubules remain bundled and the centromeres stay apparently attached to the depolymerizing microtubule plus ends. In fact, yeast cells appear to keep their centromeres attached to the microtubule plus end throughout the quiescence cycle, just as they do during proliferation.Citation35 Keeping centromeres attached to microtubules during quiescence could be a mechanism to avoid the recapture step that would delay re-entry into the cell cycle. Indeed, for single cell eukaryotes like yeast that are in direct competition for nutrients with other species, quiescent exit must be fast since it conditions the survival of the species in the environmental niche.
Most cells unable to assemble a microtubule bundle die after a few days in quiescence. Interestingly, the survivors show a delay in resuming the cell cycle upon refeeding and not many are capable of producing healthy progeny. In fact, most of the cells either do not divide or give rise to a micro-colony.Citation35 Mutants unable to assemble a microtubule bundle may experience genome instability in quiescence or chromosome segregation defects upon quiescence exit, processes that would ultimately lead to unfit progeny.
Upon quiescence establishment, the formation of a stable microtubule bundle inside the nucleus may be a S. cerevisiae specificity that is linked to the particular architecture of its spindle pole body. Indeed, the budding yeast SPB is permanently inserted across the nuclear membrane and displays a cytoplamic and a nuclear face that are both capable of nucleating microtubule. Intriguingly, in mammals, quiescence entry has been associated with the formation of a primary cilium, a microtubule containing-structure that is nucleated from the elder centriole of the centrosome.Citation60 It has been proposed that the primary cilium may function as a “physical checkpoint” for cell cycle re-entry since its resorption is apparently required for quiescence exit.Citation61 Might the microtubule intranuclear bundle assembled in S. cerevisiae be the distant cousin of the primary cilium?
Disclosure of Potential Conflicts of Interest
No potential conflict of interest was disclosed.
Acknowledgments
We are very thankful to B Daignan-Fornier, J Dompierre, and M Gupta for their precious comments on this manuscript. We are particularly grateful to JP Javerzat for enlightening discussions about our work and his tremendous help in writing this manuscript. This work was supported by a Region Aquitaine post fellowship (to D.L.), a Young Investigator Grant from the Agence Nationale pour la Recherche (grant number JC08 310804 to I.S.), and an Association pour la Recherche sur le Cancer Grant (grant number SFI20101201558 to I.S.).
References
- Cheung TH, Rando TA. Molecular regulation of stem cell quiescence. Nat Rev Mol Cell Biol 2013; 14:329 - 40; http://dx.doi.org/10.1038/nrm3591; PMID: 23698583
- O’Farrell PH. Quiescence: early evolutionary origins and universality do not imply uniformity. Philos Trans R Soc Lond B Biol Sci 2011; 366:3498 - 507; http://dx.doi.org/10.1098/rstb.2011.0079; PMID: 22084377
- Valcourt JR, Lemons JM, Haley EM, Kojima M, Demuren OO, Coller HA. Staying alive: metabolic adaptations to quiescence. Cell Cycle 2012; 11:1680 - 96; http://dx.doi.org/10.4161/cc.19879; PMID: 22510571
- De Virgilio C. The essence of yeast quiescence. FEMS Microbiol Rev 2012; 36:306 - 39; http://dx.doi.org/10.1111/j.1574-6976.2011.00287.x; PMID: 21658086
- Wells A, Griffith L, Wells JZ, Taylor DP. The dormancy dilemma: quiescence versus balanced proliferation. Cancer Res 2013; 73:3811 - 6; http://dx.doi.org/10.1158/0008-5472.CAN-13-0356; PMID: 23794703
- Ro SH, Liu D, Yeo H, Paik JH. FoxOs in neural stem cell fate decision. Arch Biochem Biophys 2013; 534:55 - 63; http://dx.doi.org/10.1016/j.abb.2012.07.017; PMID: 22902436
- Malumbres M, Barbacid M. To cycle or not to cycle: a critical decision in cancer. Nat Rev Cancer 2001; 1:222 - 31; http://dx.doi.org/10.1038/35106065; PMID: 11902577
- Coller HA, Sang L, Roberts JM. A new description of cellular quiescence. PLoS Biol 2006; 4:e83; http://dx.doi.org/10.1371/journal.pbio.0040083; PMID: 16509772
- Klosinska MM, Crutchfield CA, Bradley PH, Rabinowitz JD, Broach JR. Yeast cells can access distinct quiescent states. Genes Dev 2011; 25:336 - 49; http://dx.doi.org/10.1101/gad.2011311; PMID: 21289062
- Gray JV, Petsko GA, Johnston GC, Ringe D, Singer RA, Werner-Washburne M. “Sleeping beauty”: quiescence in Saccharomyces cerevisiae.. Microbiol Mol Biol Rev 2004; 68:187 - 206; http://dx.doi.org/10.1128/MMBR.68.2.187-206.2004; PMID: 15187181
- Wei W, Nurse P, Broek D. Yeast cells can enter a quiescent state through G1, S, G2, or M phase of the cell cycle. Cancer Res 1993; 53:1867 - 70; PMID: 8467507
- Laporte D, Lebaudy A, Sahin A, Pinson B, Ceschin J, Daignan-Fornier B, Sagot I. Metabolic status rather than cell cycle signals control quiescence entry and exit. J Cell Biol 2011; 192:949 - 57; http://dx.doi.org/10.1083/jcb.201009028; PMID: 21402786
- Daignan-Fornier B, Sagot I. Proliferation/Quiescence: When to start? Where to stop? What to stock?. Cell Div 2011; 6:20; http://dx.doi.org/10.1186/1747-1028-6-20; PMID: 22152110
- Pardee AB. A restriction point for control of normal animal cell proliferation. Proc Natl Acad Sci U S A 1974; 71:1286 - 90; http://dx.doi.org/10.1073/pnas.71.4.1286; PMID: 4524638
- Cooper S. Reappraisal of serum starvation, the restriction point, G0, and G1 phase arrest points. FASEB J 2003; 17:333 - 40; http://dx.doi.org/10.1096/fj.02-0352rev; PMID: 12631573
- Martinez MJ, Roy S, Archuletta AB, Wentzell PD, Anna-Arriola SS, Rodriguez AL, Aragon AD, Quiñones GA, Allen C, Werner-Washburne M. Genomic analysis of stationary-phase and exit in Saccharomyces cerevisiae: gene expression and identification of novel essential genes. Mol Biol Cell 2004; 15:5295 - 305; http://dx.doi.org/10.1091/mbc.E03-11-0856; PMID: 15456898
- Radonjic M, Andrau JC, Lijnzaad P, Kemmeren P, Kockelkorn TT, van Leenen D, van Berkum NL, Holstege FC. Genome-wide analyses reveal RNA polymerase II located upstream of genes poised for rapid response upon S. cerevisiae stationary phase exit. Mol Cell 2005; 18:171 - 83; http://dx.doi.org/10.1016/j.molcel.2005.03.010; PMID: 15837421
- Yotsuyanagi Y. [Study of yeast mitochondria. I. Variations in mitochondrial ultrastructure during the aerobic growth cycle]. J Ultrastruct Res 1962; 7:121 - 40; http://dx.doi.org/10.1016/S0022-5320(62)80031-8; PMID: 14002756
- Laporte D, Salin B, Daignan-Fornier B, Sagot I. Reversible cytoplasmic localization of the proteasome in quiescent yeast cells. J Cell Biol 2008; 181:737 - 45; http://dx.doi.org/10.1083/jcb.200711154; PMID: 18504300
- Sagot I, Pinson B, Salin B, Daignan-Fornier B. Actin bodies in yeast quiescent cells: an immediately available actin reserve?. Mol Biol Cell 2006; 17:4645 - 55; http://dx.doi.org/10.1091/mbc.E06-04-0282; PMID: 16914523
- Liu IC, Chiu SW, Lee HY, Leu JY. The histone deacetylase Hos2 forms an Hsp42-dependent cytoplasmic granule in quiescent yeast cells. Mol Biol Cell 2012; 23:1231 - 42; http://dx.doi.org/10.1091/mbc.E11-09-0752; PMID: 22337769
- Tapia H, Morano KA. Hsp90 nuclear accumulation in quiescence is linked to chaperone function and spore development in yeast. Mol Biol Cell 2010; 21:63 - 72; http://dx.doi.org/10.1091/mbc.E09-05-0376; PMID: 19889838
- Narayanaswamy R, Levy M, Tsechansky M, Stovall GM, O’Connell JD, Mirrielees J, Ellington AD, Marcotte EM. Widespread reorganization of metabolic enzymes into reversible assemblies upon nutrient starvation. Proc Natl Acad Sci U S A 2009; 106:10147 - 52; http://dx.doi.org/10.1073/pnas.0812771106; PMID: 19502427
- Noree C, Sato BK, Broyer RM, Wilhelm JE. Identification of novel filament-forming proteins in Saccharomyces cerevisiae and Drosophila melanogaster.. J Cell Biol 2010; 190:541 - 51; http://dx.doi.org/10.1083/jcb.201003001; PMID: 20713603
- Jensen PV, Larsson LI. Actin microdomains on endothelial cells: association with CD44, ERM proteins, and signaling molecules during quiescence and wound healing. Histochem Cell Biol 2004; 121:361 - 9; http://dx.doi.org/10.1007/s00418-004-0648-2; PMID: 15103468
- Rabl C. Uber zellteilung. Morphol Jahrbuch. 1885; 10:214 - 330
- Winey M, Mamay CL, O’Toole ET, Mastronarde DN, Giddings TH Jr., McDonald KL, McIntosh JR. Three-dimensional ultrastructural analysis of the Saccharomyces cerevisiae mitotic spindle. J Cell Biol 1995; 129:1601 - 15; http://dx.doi.org/10.1083/jcb.129.6.1601; PMID: 7790357
- Jin Q, Trelles-Sticken E, Scherthan H, Loidl J. Yeast nuclei display prominent centromere clustering that is reduced in nondividing cells and in meiotic prophase. J Cell Biol 1998; 141:21 - 9; http://dx.doi.org/10.1083/jcb.141.1.21; PMID: 9531545
- O’Toole ET, Winey M, McIntosh JR. High-voltage electron tomography of spindle pole bodies and early mitotic spindles in the yeast Saccharomyces cerevisiae.. Mol Biol Cell 1999; 10:2017 - 31; http://dx.doi.org/10.1091/mbc.10.6.2017; PMID: 10359612
- Taddei A, Gasser SM. Structure and function in the budding yeast nucleus. Genetics 2012; 192:107 - 29; http://dx.doi.org/10.1534/genetics.112.140608; PMID: 22964839
- Zimmer C, Fabre E. Principles of chromosomal organization: lessons from yeast. J Cell Biol 2011; 192:723 - 33; http://dx.doi.org/10.1083/jcb.201010058; PMID: 21383075
- Jin QW, Fuchs J, Loidl J. Centromere clustering is a major determinant of yeast interphase nuclear organization. J Cell Sci 2000; 113:1903 - 12; PMID: 10806101
- Wong H, Marie-Nelly H, Herbert S, Carrivain P, Blanc H, Koszul R, Fabre E, Zimmer C. A predictive computational model of the dynamic 3D interphase yeast nucleus. Curr Biol 2012; 22:1881 - 90; http://dx.doi.org/10.1016/j.cub.2012.07.069; PMID: 22940469
- Laporte D, Courtout F, Salin B, Ceschin J, Sagot I. An array of nuclear microtubules reorganizes the budding yeast nucleus during quiescence. J Cell Biol 2013; 203:585 - 94; http://dx.doi.org/10.1083/jcb.201306075; PMID: 24247429
- Trelles-Sticken E, Loidl J, Scherthan H. Bouquet formation in budding yeast: initiation of recombination is not required for meiotic telomere clustering. J Cell Sci 1999; 112:651 - 8; PMID: 9973600
- Dresser ME, Giroux CN. Meiotic chromosome behavior in spread preparations of yeast. J Cell Biol 1988; 106:567 - 73; http://dx.doi.org/10.1083/jcb.106.3.567; PMID: 2450094
- Hayashi A, Ogawa H, Kohno K, Gasser SM, Hiraoka Y. Meiotic behaviours of chromosomes and microtubules in budding yeast: relocalization of centromeres and telomeres during meiotic prophase. Genes Cells 1998; 3:587 - 601; http://dx.doi.org/10.1046/j.1365-2443.1998.00215.x; PMID: 9813109
- Howard J, Hyman AA. Microtubule polymerases and depolymerases. Curr Opin Cell Biol 2007; 19:31 - 5; http://dx.doi.org/10.1016/j.ceb.2006.12.009; PMID: 17184986
- Mitchison T, Kirschner M. Dynamic instability of microtubule growth. Nature 1984; 312:237 - 42; http://dx.doi.org/10.1038/312237a0; PMID: 6504138
- Winey M, Bloom K. Mitotic spindle form and function. Genetics 2012; 190:1197 - 224; http://dx.doi.org/10.1534/genetics.111.128710; PMID: 22491889
- van der Vaart B, Akhmanova A, Straube A. Regulation of microtubule dynamic instability. Biochem Soc Trans 2009; 37:1007 - 13; http://dx.doi.org/10.1042/BST0371007; PMID: 19754441
- Blake-Hodek KA, Cassimeris L, Huffaker TC. Regulation of microtubule dynamics by Bim1 and Bik1, the budding yeast members of the EB1 and CLIP-170 families of plus-end tracking proteins. Mol Biol Cell 2010; 21:2013 - 23; http://dx.doi.org/10.1091/mbc.E10-02-0083; PMID: 20392838
- Miki F, Okazaki K, Shimanuki M, Yamamoto A, Hiraoka Y, Niwa O. The 14-kDa dynein light chain-family protein Dlc1 is required for regular oscillatory nuclear movement and efficient recombination during meiotic prophase in fission yeast. Mol Biol Cell 2002; 13:930 - 46; http://dx.doi.org/10.1091/mbc.01-11-0543; PMID: 11907273
- Courtheoux T, Gay G, Reyes C, Goldstone S, Gachet Y, Tournier S. Dynein participates in chromosome segregation in fission yeast. Biol Cell 2007; 99:627 - 37; http://dx.doi.org/10.1042/BC20070047; PMID: 17561805
- Hunt AJ, McIntosh JR. The dynamic behavior of individual microtubules associated with chromosomes in vitro. Mol Biol Cell 1998; 9:2857 - 71; http://dx.doi.org/10.1091/mbc.9.10.2857; PMID: 9763448
- Zhai Y, Kronebusch PJ, Borisy GG. Kinetochore microtubule dynamics and the metaphase-anaphase transition. J Cell Biol 1995; 131:721 - 34; http://dx.doi.org/10.1083/jcb.131.3.721; PMID: 7593192
- Mitchison TJ. The role of microtubule polarity in the movement of kinesin and kinetochores. J Cell Sci Suppl 1986; 5:Suppl 5 121 - 8; http://dx.doi.org/10.1242/jcs.1986.Supplement_5.7; PMID: 3115997
- Miranda JJ, De Wulf P, Sorger PK, Harrison SC. The yeast DASH complex forms closed rings on microtubules. Nat Struct Mol Biol 2005; 12:138 - 43; http://dx.doi.org/10.1038/nsmb896; PMID: 15640796
- Joglekar AP, Bloom KS, Salmon ED. Mechanisms of force generation by end-on kinetochore-microtubule attachments. Curr Opin Cell Biol 2010; 22:57 - 67; http://dx.doi.org/10.1016/j.ceb.2009.12.010; PMID: 20061128
- Westermann S, Wang HW, Avila-Sakar A, Drubin DG, Nogales E, Barnes G. The Dam1 kinetochore ring complex moves processively on depolymerizing microtubule ends. Nature 2006; 440:565 - 9; http://dx.doi.org/10.1038/nature04409; PMID: 16415853
- Ramey VH, Wang HW, Nakajima Y, Wong A, Liu J, Drubin D, Barnes G, Nogales E. The Dam1 ring binds to the E-hook of tubulin and diffuses along the microtubule. Mol Biol Cell 2011; 22:457 - 66; http://dx.doi.org/10.1091/mbc.E10-10-0841; PMID: 21169562
- Chuang CH, Belmont AS. Moving chromatin within the interphase nucleus-controlled transitions?. Semin Cell Dev Biol 2007; 18:698 - 706; http://dx.doi.org/10.1016/j.semcdb.2007.08.012; PMID: 17905613
- Mehta IS, Amira M, Harvey AJ, Bridger JM. Rapid chromosome territory relocation by nuclear motor activity in response to serum removal in primary human fibroblasts. Genome Biol 2010; 11:R5; http://dx.doi.org/10.1186/gb-2010-11-1-r5; PMID: 20070886
- Hübner MR, Eckersley-Maslin MA, Spector DL. Chromatin organization and transcriptional regulation. Curr Opin Genet Dev 2013; 23:89 - 95; http://dx.doi.org/10.1016/j.gde.2012.11.006; PMID: 23270812
- Tanaka K, Kitamura E, Tanaka TU. Live-cell analysis of kinetochore-microtubule interaction in budding yeast. Methods 2010; 51:206 - 13; http://dx.doi.org/10.1016/j.ymeth.2010.01.017; PMID: 20117214
- Winey M, O’Toole ET. The spindle cycle in budding yeast. Nat Cell Biol 2001; 3:E23 - 7; http://dx.doi.org/10.1038/35050663; PMID: 11146646
- Kitamura E, Tanaka K, Kitamura Y, Tanaka TU. Kinetochore microtubule interaction during S phase in Saccharomyces cerevisiae.. Genes Dev 2007; 21:3319 - 30; http://dx.doi.org/10.1101/gad.449407; PMID: 18079178
- Rieder CL, Cole RW, Khodjakov A, Sluder G. The checkpoint delaying anaphase in response to chromosome monoorientation is mediated by an inhibitory signal produced by unattached kinetochores. J Cell Biol 1995; 130:941 - 8; http://dx.doi.org/10.1083/jcb.130.4.941; PMID: 7642709
- Foley EA, Kapoor TM. Microtubule attachment and spindle assembly checkpoint signalling at the kinetochore. Nat Rev Mol Cell Biol 2013; 14:25 - 37; http://dx.doi.org/10.1038/nrm3494; PMID: 23258294
- Tucker RW, Pardee AB, Fujiwara K. Centriole ciliation is related to quiescence and DNA synthesis in 3T3 cells. Cell 1979; 17:527 - 35; http://dx.doi.org/10.1016/0092-8674(79)90261-7; PMID: 476831
- Kim S, Tsiokas L. Cilia and cell cycle re-entry: more than a coincidence. Cell Cycle 2011; 10:2683 - 90; http://dx.doi.org/10.4161/cc.10.16.17009; PMID: 21814045