Abstract
Scrapie and CWD are horizontally transmissible, and the environment likely serves as a stable reservoir of infectious prions, facilitating a sustained incidence of CWD in free-ranging cervid populations and complicating efforts to eliminate disease in captive herds. Prions will enter the environment through mortalities and/or shedding from live hosts. Unfortunately, a sensitive detection method to identify prion contamination in environmental samples has not yet been developed. An environmentally-relevant prion model must be used in experimental studies. Changes in PrPSc structure upon environmental exposure may be as significant as changes in PrPSc quantity, since the structure can directly affect infectivity and disease pathology. Prions strongly bind to soil and remain infectious. Conformational changes upon adsorption, competitive sorption, and potential for desorption and transport all warrant further investigation. Mitigation of contaminated carcasses or soil might be accomplished with enzyme treatments or composting in lieu of incineration.
Introduction
Prion diseases, or transmissible spongiform encephalopathies (TSEs), are fatal neurodegenerative diseases impacting a number of mammalian species, including cattle (bovine spongiform encephalopathy, BSE or ‘mad cow’ disease), sheep and goats (scrapie), deer, elk and moose (chronic wasting disease, CWD) and humans (Creutzfeldt-Jakob disease, CJD and others).Citation1 Scrapie and CWD are of particular environmental concern as they are horizontally transmissible and remain infectious after years in the environment.Citation2–Citation5 It is likely that the environment serves as a stable reservoir of infectious CWD and scrapie prions. In addition, the disposal of mortalities during BSE outbreaks, both in the past and potential future disposal events, serves as another environmental source of prions with the potential to infect humans. Therefore, it is clear that prions pose a significant environmental concern.
There has been limited risk assessment of prion exposure through environmental pathways, focusing exclusively on BSE. The risk of BSE from sources including landfills and rendering facilities contaminating drinking water has been evaluated,Citation6,Citation7 as well as the risks to cattle from land-applied fly ash, slag, slaughterhouse sludge and meat and bone meal contaminated with BSE.Citation8–Citation10 Due to the large dilution factors, the calculated risk of transmission was extremely low in all cases. However, CWD risks have not been quantified, and it is uncertain if adequate information is available to make credible assessments of prion transmission via environmental pathways. A recent editorial emphasized the need for more information about prion fate in the environment for the development of quantitative risk assessments.Citation11
Although various aspects of prions in the environment have been reviewed previously, a complete review of literature related to prion occurrence, fate and mitigation in the environment is lacking. The potential role of soil in prion transmission was reviewed in 2006,Citation12 but over 30 pertinent studies have been published during the ensuing two years. Snow and colleagues separately reported the prion environmental literature published in 2006Citation13 and 2007.Citation14 Recently, Wiggens examined prion stability and infectivity in the environment.Citation15 In contrast, this article will comprehensively review the pertinent literature addressing prion occurrence and fate in the environment, with a specific focus on prion interactions with soil. Potential avenues for environmental mitigation of prion infectivity will be considered. Experimental challenges in studying environmental prions will also be discussed, and important unanswered questions in the field will be highlighted.
Prion Diseases and the Prion Protein
Strong evidence indicates that the infectious agent of prion diseases is solely comprised of PrPSc (the prion), an abnormally-folded isoform of a normal cellular protein, PrPc.Citation1,Citation16 The misfolded conformation of PrPSc conveys distinct biological and physicochemical properties, including resistance to proteolysis and inactivation techniques, increased hydrophobicity and a propensity for aggregation.Citation1,Citation17 The CWD and scrapie agent is shed from living hosts and present in mortalities.Citation18,Citation19 CWD and scrapie are orally transmissible,Citation20,Citation21 and the nasal cavity is also an effective route of infectivity.Citation22 Recent bioassay data suggest that prions bound to soil particles remain infectious through oral consumption.Citation23,Citation24
While human prion diseases are very rare, CWD incidence can be over 15% in free-ranging deer and over 90% in captive deer herds.Citation25 CWD has been identified in cervid populations in 16 US states and two Canadian provinces.Citation26 Scrapie occurs in all sheep-farming countries except Australia and New Zealand.Citation27 The economic impacts of CWD on the affected states may be high due to lost revenues in the agriculture, hunting and tourism industries.Citation28,Citation29 Cross-species transmission of prion diseases can occur, as demonstrated by the transmission of BSE to humans, highlighting the public health risk of animal prion diseases.Citation30 A recent report of a single incident where 81 people were exposed to or consumed CWD-infected venison emphasizes the opportunity for CWD transmission to humans.Citation31 There is growing evidence that the potential for CWD transmission to humans is low, but the risk is still not completely known.Citation30,Citation32–Citation35 Natural transmission of CWD to livestock (especially cattle) has not been demonstrated, but remains an unknown risk. Although transmission of CWD to humans and livestock has not been shown, it might occur if a new strain emerges, either through serial passage in cervids or through environmental pressures. Host range is known to vary between prion strains.Citation36 No distinct CWD strains have yet been identified, but evidence suggests they may exist.Citation36–Citation38
Distinct strains have been identified for other prion diseases, including scrapie and BSE. Prion strains are defined by varied clinical symptoms, incubation times and distinct pathological characteristics.Citation39 The conformation of PrPSc (the disease-causing prion protein) is strain dependent.Citation40–Citation44 Because no nucleic acid content has been associated with the prion agent,Citation16,Citation45 it is likely that differences in PrPSc conformation are the sole reason for strain differences.Citation46 Thus, the structure of PrPSc can affect the nature of the disease and, therefore, we propose that changes in PrPSc conformation due to environmental conditions could change prion strain properties.
Routes of Entry, Occurrence and Detection in the Environment
It is now established that scrapie and CWD are horizontally transmissible and can remain infectious after years in the environment.Citation2–Citation4 In one study, the scrapie agent remained infectious after burial in garden soil for three years.Citation47 In another report, a previously scrapie-infected sheephouse and pasture were ‘decontaminated’ and left uninhabited; sheep introduced 16 years later subsequently contracted scrapie suggesting that scrapie remained infectious after 16 years.Citation48 A controlled lab study indicated that the CWD agent remained infectious for at least two years in a pasture.Citation5 Epidemiological modeling suggests that indirect, environmental routes of transmission were responsible for two CWD outbreaks in captive mule deer.Citation49
The existence and nature of environmental reservoirs of prion infectivity have yet to be determined. Recent experiments with prions and soil suggest soil and soil minerals may act as significant reservoirs, but the possibility of environmental transmission sustained by mites or flies cannot be eliminated.Citation50,Citation51 It is also unclear if predators or scavengers (e.g., cougars, vultures) play a significant role in CWD spread in free-ranging cervids.
Prions may enter the environment through a number of routes. First, prions may enter through shedding from live, infected hosts. It has been shown that scrapie and CWD prions can be shed in urine,Citation52,Citation53 feces,Citation54,Citation55 saliva and blood.Citation18 Scrapie can also be shed in birthing matter.Citation56 A second route of entry is through animal mortalities, including farmed sheep, goats and cervids as well as free-ranging cervids. Scrapie, BSE and CWD mortalities contain high levels of infectivity in the central nervous system (CNS), with lower levels of infectivity in extraneural tissues.Citation19 A future outbreak of BSE, scrapie or CWD in captive herds may require the culling of large numbers of animals. While it would be desirable to incinerate these mortalities, biosecurity concerns or other constraints may limit the transport of carcasses over long distances. Thus, other options like on-site burial or composting may be employed. During the early years of the BSE outbreak in the United Kingdom (1988–1991), it is estimated that 6,000 carcasses that were suspected of having BSE were disposed of in 59 landfill sites.Citation7 Another potential route of entry is via solid or liquid waste from rendering plants and slaughterhouses unknowingly processing infected carcasses.
There has been much speculation and interest in environmental locations of concentrated prion infectivity (“hot spots”). Locations of concentrated prion infectivity could be formed at areas of communal activity where shedding of prions in saliva, urine, feces or birthing matter occurs. A recent study of elk wallows suggests that they are used too infrequently to be significant sources of CWD transmission in the wild,Citation57 but mineral licks might be important ‘hot spots’. Animal mortality sites, where highly-infectious CNS matter would enter the environment, could also be hot spots. Hot spots would be important targets for CWD eradiation efforts should a viable mitigation method be developed. These locations would also presumably contain detectable levels of prion contamination, if a sensitive method were successfully developed.
Highly-sensitive and accurate detection of prion infectivity in the environment is not currently possible. Standard methods such as western blotting fail to detect significant levels of infectivity,Citation58–Citation60 and the most sensitive method of prion detection, animal bioassay, would be impractical for use on large numbers of environmental samples. Protein misfolding cyclic amplification (PMCA),Citation61 developed by Soto and colleagues for detecting small amounts of PrPSc, has generated much interest for use as an environmental detection method. PMCA has been used successfully with CWD and with hamster PrPSc extracted from soil with SDS.Citation24,Citation62 The recently developed QUIC (quake-induced conversion) method,Citation63,Citation64 which uses recPrP as a substrate instead of uninfected brain homogenate, might be a viable alternative to PMCA as an environmental diagnostic tool. Quantitative tandem mass spectrometric techniquesCitation65 may also be developed as a sensitive environmental detection and quantification method for PrP.
Infectivity, Fate and Transport of Prions in the Environment
Understanding the significant route(s) of prion entry into the environment will lead to a better understand of prion fate and transport once in the environment. The matrix in which prions enter soil or water environments (e.g., decomposing tissue, feces, urine, saliva) could greatly affect survivability and transmissibility. Once prions enter the environment, they exist in an exceedingly complex system, detailed in , involving a large number of constituents and a wide variety of biological, chemical and physical mechanisms that can influence prion infectivity and fate. Progress towards an understanding of the most significant mechanisms has been made, but more work is needed to determine the behavior of PrPSc upon entering the heterogeneous soil-water environment.
Infectivity of adsorbed prions.
One of the most important tasks in the study of CWD and scrapie transmission is identifying the significant route or routes of natural inoculation so that disease mitigation efforts can be effectively designed. The potential for CWD or scrapie transmission by exposure to contaminated soil is possible since cervids and ruminants are known to ingest and inhale large amounts of soil.Citation66,Citation67 Detection of prion infectivity after exposure to the soil environment was originally demonstrated by Brown and Gajdusek in 1991.Citation47 Scrapie-infected hamster brain homogenate mixed with soil and stored outdoors in buried flowerpots for three years remained infectious. Seidel and colleagues recently conducted a similar experiment, demonstrating high levels of infectivity (by repeated oral inoculation) in prion-contaminated soil after 26 months burial.Citation24
Johnson and colleagues have shown that prions bound to soil minerals remain infectious,Citation23,Citation68 and they report prions bound to soil are more infectious than unbound prions.Citation23 Increased infectivity could be due to preferable conformational changes in PrPSc (including changes in aggregation) upon binding. Uptake of PrPSc upon ingestion or inhalation could also be enhanced by binding to soil, either by increasing PrPSc residence time in the animal or improving PrPSc accessibility to uptake mechanisms. Further studies are needed to determine how prion infectivity is altered upon interaction with soil.
The infectivity of soil-bound prions has also been confirmed by a cell culture model.Citation69 Unfortunately, the cell line is only susceptible to select mouse prion strains, but this technique might be useful in future soil-prion infectivity studies if new cell lines are developed, as it is less expensive and faster than animal bioassay. Nonetheless, animal bioassay will continue to be an important tool in studying not only the infectivity of soil-bound prions but also changes in disease pathology and strain properties due to environmental exposure and binding to soil.
Environmentally-relevant forms of PrP.
In experimental studies, the PrP source and medium are important to accurately simulate the route of entry, subsequent environmental interactions, and the potential transmissibility of prions in the environment. Studies performed to date have used a wide variety of prion materials, including recombinant PrP (recPrP),Citation70–Citation78 purified hamster PrPSc,Citation23,Citation68,Citation79 infectious rodent brain homogenate,Citation23,Citation24,Citation47,Citation68,Citation69,Citation80–Citation84 infectious cervid or ovine brain homogenate,Citation80,Citation82,Citation83 and intact infectious ovine tissueCitation85 (see , discussed in detail below). As outlined in , these prion models vary greatly both in their ease of experimental use and in their applicability to the study of prions in the environment. Recombinant PrP is only useful as a surrogate of PrPc. Purified PrPSc is infectious but artificially aggregated,Citation86 and its production requires a large amount of infectious tissue. Both PrPc and PrPSc exist in vivo as a highly heterogeneous population with both full-length and N-terminal truncated forms as well as glycosylated and unglycosylated forms.Citation87–Citation91 These forms are a small proportion of the overall organic content which will enter the environment simultaneously with prions. This heterogeneity is best modeled using homogenized or intact infectious tissue.
Full-length PrPSc might be rare in the brain matter of animal carcasses greater than 1 week old.Citation83 Thus, a large population of PrP will enter and exist in the environment without the N-terminal domain, and the environmental behavior of PrPSc lacking the N-terminal region could be markedly different from full-length PrPSc. The N-domain has been shown to affect PrP aggregation,Citation92 and evidence suggests the N-domain might play a role in sorption to soils.Citation68,Citation70,Citation73,Citation74,Citation76 Therefore, both full-length and truncated prions should be used to study environmental fate.
Rodent-adapted prions may not simulate prion fate in the environment. In one study, PrP from rodent (HY TME) brain homogenate significantly degraded upon incubation at room temperature while CWD-elk PrP did not.Citation83 A transgenic (Tg) CWD model mimicked the natural elk PrP. Differences between rodent PrP degradation and natural (sheep scrapie, BSE) PrP degradation were also seen in two other degradation studies.Citation80,Citation82 Thus, caution should be taken when extrapolating rodent results to a natural prion system. Recently developed Tg mouse models expressing the elk, ovine, bovine or human prion protein (PrPc) may be valuable research tools in studying the infectivity of environmental samples.Citation37,Citation93,Citation94 In addition, because Tg tissue is in many cases easier to procure than natural CWD or scrapie tissue, transgenic tissue might be used as PrP source material for environmental fate studies.
Because the structure of PrP is likely the sole determinant of infectivity and strain properties,Citation16,Citation41–Citation46,Citation95 changes in PrP structure due to environmental mechanisms could directly impact transmissibility and pathogenesis. This is analogous to other environmental contaminants such as hormones, pesticides or other organic compounds, where metabolites can have differential toxicity from the parent compound. However, the prion-environment system is more complex due to the initial heterogeneity of PrPSc (in vivo) and the three-dimensional and yet undefined structure of PrPSc.
Prion sorption to soil.
Prion sorption (physical or chemical binding) to soil could play an important role in prion transport or immobilization, provide protection from or enhance in situ proteolysis, and induce conformational changes that enhance or decrease infectivity, all of which could affect the transmissibility of CWD, scrapie and BSE in the environment. The fate of proteins in soil environments has relevance not only to the study of prion fate and transmission but also the fate and activity of viruses and bacteria as well as extracellular enzymes such as Bacillus thuringiensis (Bt) toxins released from transgenic crops.Citation96 Answers to basic questions such as the role protein sorption to soil minerals plays in soil ecology are currently lacking. Recent studies, detailed in and summarized below, have started to explore the complex interactions between prions and soil ().
Protein sorption has generally been observed to be strong, fast and irreversible for a wide range of surfaces.Citation97 Prions are no exception and both PrPc and PrPSc appear to have an affinity for quartz sands and soils and a particularly strong affinity for clay minerals.Citation68,Citation73,Citation74,Citation77,Citation79 Prion sorption is strongly irreversible and resistant to detergent and chaotropic treatments.Citation71,Citation81 Studies using recPrP and purified PrPSc hours;Citation68,Citation71,Citation78 have observed maximal sorption in less than two however, PrP sorption kinetics might be significantly longer in the complex, competitive soil environment.
As with other proteins, prion sorption is most likely a function of electrostatic attractions and repulsions and hydrophobic interactions. RecPrP studies have identified electrostatic attraction between positively-charged peptides and negatively-charged mineral surfaces as the most significant adsorption mechanism.Citation70,Citation71 However, since the three-dimensional structure of PrPSc remains unknown, it is a challenge to model the specific mechanisms that are significant in PrPSc adsorption. It is known that PrPSc is highly insoluble and aggregated, and hydrophobic interactions could therefore play a larger role in PrPSc sorption to amorphous clay minerals. PrPSc adsorption to quartz sand was shown to be maximal around the isoelectric point (IEP, at pH ≈ 4 for prions), corresponding to maximal PrPSc aggregation.Citation79 Maximum adsorption near the IEP has been shown for many other proteins and surfaces.Citation98
Because the N-terminal domain is known to be flexibly-disordered and contains a high number of positively-charged amino acids, it may play a significant role in electrostatic attraction to negatively-charged mineral surfaces.Citation70 The N-terminal domain is lost upon desorption of PrPSc from clay,Citation68 and PK-digestion is required to desorb maximal PrPSc from clay and sandy soils.Citation74 More full-length recPrP adsorbs to montmorillonite than N-terminal truncated recPrP.Citation73 However, the N-domain is not needed for prion adsorption.Citation68,Citation74,Citation76
Prions enter the environment concurrently with organic matter from the host. In addition, the soil environment contains native organic matter. Thus, it is important to consider the role organic matter plays in prion sorption. recPrP has been shown to have a high affinity for organic matter, equal to or greater than that calculated for mineral surfaces.Citation76–Citation78 In addition, humic acid was found to increase the recPrP sorption capacity of kaolinite about 10-fold.Citation77 It is also important to note that solution ionic strength and the homoionic salt species (e.g., Na+, Ca2+) can significantly affect the measured sorption capacities for organic matter and clay minerals, including PrP capacity.Citation77,Citation79 While other proteins have been shown to enter the interlayer areas of expandable clays (e.g., smectites),Citation99 potentially decreasing their bioavailability, Johnson et al. found that purified PrPSc did not enter the interlayer area of Na+-montmorillonite.Citation68 However, choice of homoionic species can affect the height of the interlayer area.
Conformational changes in protein structure upon interaction with soil minerals have been well documented and can range from slight unfolding to significant changes in secondary structure and can also include aggregation or disaggregation.Citation97 Proteins can also reorient over time between side-on and end-on orientations. PrPSc is aggregated, and changes in aggregation could occur, potentially affecting infectivity.Citation95 One study did find that recPrP does not form β-sheets or self-aggregate, as seen in vitro, when adsorbed to clay.Citation70 More must be done to determine what conformational changes (if any) occur to PrPSc when it binds to minerals and how these changes affect agent survival and infectivity.
Experimental methods for prion sorption have initially focused on using detergents and boiling to desorb prions into solution where they are detected by immunoblotting techniques ().Citation81 Bound PrP is usually first separated from unsorbed prions by low-speed centrifugation (in some cases by using a sucrose cushion).Citation68,Citation69,Citation81 Unsorbed PrP can be detected by normal immunoblotting, but bound PrP must be desorbed before detection. Unfortunately, detergent and boiling extraction methods typically have very low PrP recoveries, presumably due to the strong and near-irreversible binding of PrP to soil particles. Recoveries reported are 61–67% (wastewater sludge)Citation84 and 5–40% for sandy and clay soils.Citation74,Citation75 An electroelution method developed by Rigou et al. reported similar recoveries (5–40%).Citation73 It is likely that desorption selects for a certain PrP population, such as loosely-bound aggregates.
A direct detection method has been developed by Genovesi and colleagues which bypasses the need to desorb and instead directly detects the PrP bound to soil via immunological methods.Citation69 This method holds promise for use in future studies, including studies of desorption and desiccation behavior. While short-term prion desorption has not been observed except after detergent treatments, longer-term desorption and mobilization could occur. Drying and desiccation, which will undoubtedly take place in many prion-contaminated environments, could significantly alter prion sorption and infectivity. Desiccation can result in protein conformational changes,Citation100 although low-heat treatments of prions do not affect infectivity.Citation101
Abiotic pathways may play a key role in prion transport. Due to their insolubility and high affinity for clays and silts, prions are unlikely to be transported long distances in surface water. Recent studies simulating prion fate in wastewater found that PrP strongly partitioned into the sludge solids.Citation80,Citation84 To date, only one study has directly evaluated the mobility of prions in soil.Citation75 In this study, recPrP was placed in a soil column and unsaturated pore water was sampled for nine months. RecPrP was detected in the original contamination layer throughout the experiment, even in columns with active soil microbial populations. Only slight recPrP migration was observed, indicating that the recPrP was strongly sorbed to soil and underwent limited transport in soil systems. The potential for colloid-facilitated prion transport was not investigated in this study. Colloid-facilitated transport has been shown to be a significant transport process for many strongly-sorbing contaminants.Citation102 In addition, infectious prions can form aggregates of colloidal sizeCitation95 and might be transported unassociated. Macro-pore colloid-facilitated transport could quickly move prions into groundwater or surface waters and therefore warrants further study.
Almost all prion sorption studies have used recPrP or purified PrPSc as the source of infectious material (). These systems do not take into account the competitive matrix (animal tissue, blood, saliva or excreta) in which prions will enter the environment. Competition for sorption sites and interactions on mineral surfaces between prions and other proteins, carbohydrates, lipids and nucleic acids could all be significant factors. Many studies have found significant effects on sorption capacity and on conformational changes due to competition between plasma proteins exposed to surfaces.Citation103 Therefore, it is uncertain how applicable the previously published results are in simulating the complex nature of prion fate in the environment.
Degradation and Mitigation of Prions in the Environment
Prions are subject to degradation in both natural and engineered settings. Bacterial enzymes which effectively degrade prions have been identified, but they are most effective at high pH (10–12) and high temperature (50–60°C).Citation59,Citation104 Microbiological consortia taken from the rumen and colon of cattle could degrade PrPSc to undetectable levels within 20 hours under anaerobic conditions at 37°C, although infectivity remained.Citation60,Citation105 Using an enzyme treatment, it may be possible to lower or eliminate the infectivity at identified or presumed CWD and scrapie ‘hot spots’ in captive and wild settings.
Incineration of prion-contaminated material is considered the most effective method of disposal. Combustion at 1,000°C can destroy prion infectivity, however, low infectivity remains after treatment at 600°C.Citation106 Despite its effectiveness, incineration may not be a practical solution, such as during a large outbreak of BSE, scrapie or CWD requiring a mass culling. Incineration of contaminated soil, vegetation and farm infrastructure (paddocks, fences) to eliminate CWD or scrapie environmental infectivity is also not practical.
Composting mortalities might be an attractive alternative if incineration is not practical. Huang et al. found composting of scrapie-infected tissue reduced or completely eliminated PrPSc as detected by western blot.Citation85 Others have demonstrated PrPSc reductions in CWD-infected elk and deer brain homogenates incubated at 55°C.Citation107 Infectivity was not determined in either study. The risk of landfilling carcasses, residuals from composting or other prioncontaminated materials has not yet been thoroughly investigated. Thus, the ultimate fate of the PrPSc from the 6,000 landfilled carcasses in the UK suspected of being BSE-contaminated remains unclear.
PrPSc degradation might occur in the subsurface environment. One study found recPrP was degraded by soil extracted soluble enzymes after eight days incubation at 28°C,Citation72 but the use of recPrP greatly limits the significance of these results. Previous studies found prion infectivity two or more years after burial of infectious brain homogenates.Citation24,Citation47 Therefore, it is highly likely that infectivity will remain many years after burial. Evaluation of the potential for prion transport in unsaturated and saturated zones will help determine the mobility of prions in landfill environments, better quantifying the risk of landfilling infectious material.
The risk of prions in wastewater and biosolids has recently gained some attention in the wastewater industry.Citation11,Citation108,Citation109 Prions are unlikely to enter wastewater except through effluent from slaughterhouses unknowingly rendering prion mortalities or through contaminated effluent from hospital or research facilities. Hinckley and colleagues recently determined that most PrPSc and prion infectivity would associate with the activated sludge solids, survive mesophilic anaerobic digestion, and be present in the remaining biosolids.Citation84 Likewise, Kirchmayr et al. found no significant decrease in PrPSc after 16 days incubation in mesophilic anaerobic sludge and observed PrPSc solids association.Citation80 PrPSc degradation was observed in thermophilic anaerobic sludge, although maximum degradation occurred in sterilized samples.Citation80 Others found a large decrease in PrPSc within 15 days after incubating BSE brain homogenates in municipal sewage at 20°C.Citation82 Sheep scrapie brain homogenates were somewhat more resistant to degradation. Based on these studies, it can be assumed that most prion infectivity will be conserved during normal wastewater treatment processes, and prions would thus enter the environment, highly diluted, via landfill disposal or landspreading of biosolids.
Conclusion
Determination of the optimum conditions and treatments for prion degradation and immobilization should lead to best management practices for combating CWD and scrapie and for dealing with future outbreaks of prion disease requiring the disposal of a large number of carcasses. To answer practical questions about disease mitigation, it will be necessary to determine (1) the routes of prion entry into the environment, (2) the reservoirs of prions in the environment, and (3) the mechanisms for disease transmission via the environment. The results of fundamental research on prion interactions with soil will undoubtedly begin to answer these questions, and may also lead to a better understanding of basic protein-soil adsorption processes.
The study of prions in the environment is highly challenging. No other environmental ‘contaminant’ has the complex structure of prions. The fact that this structure is not even known and that many standard analytical methods are not well-developed adds to the challenge. Monitoring PrPSc structure during environmental studies is as equally important as quantification measurements, since changes in structure can directly affect infectivity and disease pathology. Although some innovation will be required, applications of analytical techniques such as dynamic light scattering, liquid chromatography-mass spectrometry, and atomic force microscopy hold promise in yielding more quantitative and/or more precise structural measurements of environmental prions compared to current immunological methods, which also require disruptive purification methods. Of critical importance is the ability to study prion quantity and structure in complex systems such as tissue homogenates and intact soils where there are countless interferences and competing factors. In particular, measuring changes in aggregation state as prions are exposed to the environment may be the key to assessing prion mobility, stability, infectivity and ultimately, transmissibility in the environment.
Abbreviations
| CWD | = | chronic wasting disease |
| BSE | = | bovine spongiform encephalopathy |
| PrP | = | prion protein |
| PrPc | = | cellular, noninfectious prion protein |
| PrPSc | = | disease-causing prion protein |
| recPrP | = | recombinant prion protein |
Figures and Tables
Figure 1 The complex nature of prion-soil interactions.
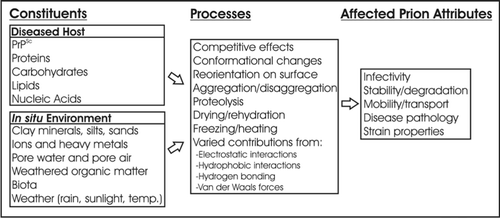
Figure 2 Prion models used to study environmental fate.
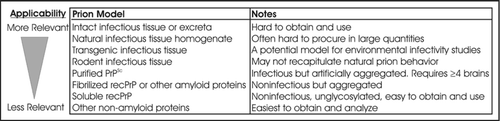
Table 1 Review of prion sorption literature
Acknowledgements
This work was supported by the National Center for Research Resources (C06 RR17417-01).
References
- Prusiner SB. Prusiner SB. An introduction to prion biology and diseases. Prion Biology and Diseases 2004; Cold Spring Harbor, NY Cold Spring Harbor Laboratory Press 1 - 89
- Greig JR. Scrapie: observation on the transmission of the disease by mediate contact. Vet J 1940; 96:203 - 206
- Hadlow WJ, Kennedy RC, Race RE. Natural infection of Suffolk sheep with scrapie virus. J Infect Dis 1982; 146:657 - 664
- Miller MW, Williams ES. Horizontal prion transmission in mule deer. Nature 2003; 425:35 - 36
- Miller MW, Williams ES, Hobbs NT, Wolfe LL. Environmental source of prion transmission in mule deer. Emerging Infect Dis 2004; 10:1003 - 1006
- Gale P, Young C, Stanfield G, Oakes D. Development of a risk assessment for BSE in the aquatic environment. J Appl Microbiol 1998; 84:467 - 477
- The Environmental Agency. Assessment of risk from BSE carcasses in landfills 1997; London, UK Prepared by Det Norske Veritas Limited
- Cummins E, Adkin A. Exposure assessment of TSEs from the landspreading of meat and bone meal. Risk Anal 2007; 27:1179 - 1202
- Paisley LG, Pedersen JH. A quantitative assessment of the BSE risk associated with fly ash slag from the incineration of meat-and-bone meal in a gas-fired power plant in Denmark. Prevent Vet Med 2005; 68:263 - 275
- Yamamoto T, Kobayashi S, Nishiguchi A, Nonaka T, Tstsui T. Evaluation of bovine spongiform encephalopathy (BSE) infection risk of cattle via sewage sludge from wastewater treatment facilities in slaughterhouses in Japan. J Vet Med Sci 2006; 68:137 - 142
- Pedersen JA, McMahon KD, Benson CH. Prions: Novel pathogens of environmental concern?. J Environ Eng 2006; 132:967 - 969
- Schramm PT, Johnson CJ, Mathews NE, McKenzie D, Aiken JM, Pedersen JA. Potential role of soil in the transmission of prion disease. Rev Mineral Geochem 2006; 64:135 - 152
- Snow DD, Bartelt-Hunt SL, Saunders SE, Cassada DA. Detection, occurrence and fate of emerging contaminants in agricultural environments. Water Environ Res 2007; 79:1061 - 1084
- Snow DD, Bartelt-Hunt SL, Saunders SE, Devivo S, Cassada DA. Detection, occurrence and fate of emerging contaminants in agricultural environments. Water Environ Res 2008; 80:868 - 879
- Wiggins RC. Prion stability and infectivity in the environment. Neurochem Res 2008; 34:158 - 168
- Deleault NR, Harris BT, Rees JR, Supattapone S. Formation of native prions from minimal components in vitro. PNAS 2007; 104:9741 - 9746
- Taylor DM. Inactivation of transmissible degenerative encephalopathy agents: a review. Vet J 2000; 159:10 - 17
- Mathiason CK, Powers JG, Dahmes SJ, Osborn DA, Miller KV, Warren RJ, et al. Infectious prions in the saliva and blood of deer with chronic wasting disease. Science 2006; 314:133 - 135
- Angers RC, Browning SR, Seward T, Sigurdarson CJ, Miller MW, Hoover EA, et al. Prions in skeletal muscles of deer with chronic wasting disease. Science 2006; 311:1117
- Sigurdson CJ, Williams ES, Miller MW, Spraker TR, O'Rourke KI, Hoover EA. Oral transmission and early lymphoid tropism of chronic wasting disease PrPres in mule deer fawns (Odocoileus hemionus). J Gen Virol 1999; 80:2757 - 2764
- O'Rourke KI, Holyoak GR, Clark WW, Mickelson JR, Wang S, Melco RP, et al. PrP genotypes and experimental scrapie in orally inoculated Suffolk sheep in the United States. J Gen Virol 1997; 78:975 - 978
- Kincaid AE, Bartz JC. The nasal cavity is a route for prion infection in hamsters. J Virol 2007; 81:4482 - 4491
- Johnson CJ, Pedersen JA, Chappell RJ, McKenzie D, Aiken JM. Oral transmissibility of prion disease Is enhanced by binding of soil particles. PLoS Pathog 2007; 3:93
- Seidel B, Thomzig A, Buschmann A, Groschup MH, Peters R, Beekes M, et al. Scrapie agent (strain 263K) can transmit disease via the oral route after persistence in soil over years. PLoS ONE 2007; 3:435
- Williams ES, Miller MW, Kreeger TJ, Kahn RH, Thorne ET. Chronic wasting disease of deer and elk: a review with recommendation for management. J Wildlife Manage 2002; 66:551 - 563
- USGS National Wildlife Health Center. Map of chronic wasting disease in North America 2008; Accessed at: http://www.nwhc.usgs.gov/disease_information/chronic_wasting_disease/north_america_CWD_map.jsp. 2008
- USDA Animal and Plant Health Inspection Service. Scrapie fact sheet 2004; Accessed at: http://www.aphis.usda.gov/animal_health/animal_diseases/scrapie/downloads/fs_ahscrapie.pdf. 2008
- Bishop RC. Potential economic impacts of chronic wasting disease in Wisconsin. Human Dim Wild 2004; 9:181 - 192
- Seidl AF, Koontz SR. Potential economic impacts of chronic wasting disease in Colorado. Human Dim Wild 2004; 9:241 - 245
- Belay ED, Schonberger LB. The public health impact of prion diseases. Ann Rev Pub Health 2005; 26:191 - 212
- Garruto RM, Reiber C, Alfonso MP, Gastrich H, Needham K, Sunderman S, et al. Risk behaviors in a rural community with a known point-source exposure to chronic wasting disease. Environ Health 2008;
- Belay ED, Maddox RA, Williams ES, Miller MW, Gambetti P, Schonberger LB. Chronic wasting disease and potential transmission to humans. Emerging Infect Dis 2004; 10:977 - 984
- Kong Q, Huang S, Zou WQ, Vanegas D, Wang M, Wu D, et al. Chronic wasting disease of elk: transmissibility to humans examined by transgenic mouse models. J Neurosci 2005; 25:7944 - 7949
- Marsh RF, Kincaid AE, Bessen RA, Bartz JC. Interspecies transmission of chronic wasting disease prions to squirrel monkeys (Saimiri sciureus). J Virol 2005; 79:13794 - 13796
- Raymond GJ, Bossers A, Raymond L, O'Rourke K, McHolland LE, Bryant PK III, et al. Evidence of a molecular barrier limiting susceptibility of humans, cattle and sheep to chronic wasting disease. EMBO J 2000; 19:4425 - 4430
- Raymond GJ, Raymond LD, Meade-White KD, Hughson AG, Favara C, Gardner D, et al. Transmission and adaptation of chronic wasting disease to hamsters and transgenic mice: evidence for strains. J Virol 2007; 81:4305 - 4314
- Browning SR, Mason GL, Seward T, Green M, Eliason GAJ, Mathiason C, et al. Transmission of prions from mule deer and elk with chronic wasting disease to transgenic mice expressing cervid PrP. J Virol 2004; 78:13345 - 13350
- LaFauci G, Carp RI, Meeker HC, Ye X, Kim JI, Natelli M, et al. Passage of chronic wasting disease prion into transgenic mice expressing Rocky Mountain elk (Cervus elaphus nelsoni) PrPc. J Gen Virol 2006; 87:3773 - 3780
- Fraser H, Dickinson AG. Distribution of experimentally induced scrapie lesions in the brain. Nature 1967; 216:1310 - 1311
- Bessen RA, Marsh RF. Biochemical and physical-properties of the prion protein from 2 strains of the transmissible mink encephalopathy agent. J Virol 1992; 66:2096 - 2101
- Bessen RA, Marsh RF. Distinct PrP properties suggest the molecular basis of strain variation in transmissible mink encephalopathy. J Virol 1994; 68:7859 - 7868
- Safar J, Willie H, Itri V, Groth D, Serban H, Torchia M, et al. Eight prion strains have PrPSc molecules with different conformations. Nat Med 1998; 4:1157 - 1165
- Kascsak R, Rubenstein R, Merz PA, Carp RI, Wisniewski HM, Diringer H. Biochemical differences among scrapie-associated fibrils support the biological diversity of scrapie agents. J Gen Virol 1985; 66:1715 - 1722
- Caughey B, Raymond GJ, Bessen RA. Strain-dependent Differences in B-Sheet Conformations of Abnormal Prion Protein. J Biol Chem 1998; 273:32230 - 32235
- Castilla Jn, Saa P, Hetz C, Soto C. In Vitro generation of infectious scrapie prions. Cell 2005; 121:195 - 206
- Castilla Jn, Morales R, Saa P, Barria M, Gambetti P, Soto C. Cell-free propagation of prion strains. EMBO J 2008; 27:2557 - 2566
- Brown P, Gajdusek DC. Survival of scrapie virus after 3 year's interment. Lancet 1991; 337:269 - 270
- Georgsson G, Sigurdarson S, Brown P. Infectious agent of sheep scrapie may persist in the environment for at least 16 years. J Gen Virol 2006; 87:3737 - 3740
- Miller MW, Hobbs NT, Tavener SJ. Dynamics of prion disease transmission in mule deer. Ecol Appl 2006; 16:2208 - 2214
- Carp RI, Meeker HC, Rubenstein R, Sigurdarson S, Papini M, Kascsak RJ, et al. Characteristics of scrapie isolates derived from hay mites. J Neurovirol 2000; 6:137 - 144
- Post K, Riesner D, Walldorf V, Mehlhorn H. Fly larvae and pupae as vectors for scrapie. Lancet 1999; 354:1969 - 1970
- Seeger H, Heikenwalder M, Zeller N, Kranich J, Schwarz P, Gaspert A, et al. Coincident scrapie infection and nephritis lead to urinary prion excretion. Science 2005; 310:324 - 326
- Gonzalez-Romero D, Barria MA, Leon P, Morales R, Soto C. Detection of infectious prions in urine. FEBS letters 2008; 582:3161 - 3166
- Maluquer de Motes C, Grassi J, Simon S, Herva ME, Torres JM, Pumarola M, et al. Excretion of BSE and scrapie prions in stools from murine models. Vet Microbiol 2008; 131:205 - 211
- Safar JG, Lessard P, Tamguney G, Freyman Y, Deering C, Letessier F, et al. Transmission and detection of prions in feces. J Infect Dis 2008; 198:80 - 89
- Race R, Jenny A, Sutton D. Scrapie infectivity and proteinase K-resistant prion protein in sheep placenta, brain, spleen and lymph node: implications for transmission and antemortem diagnosis. J Infect Dis 1998; 178:949 - 953
- VerCauteren KC, Burke PW, Phillips GE, Fischer JW, Seward NW, Wunder BA, et al. Elk use of wallows and potential chronic wasting disease transmission. J Wildlife Dis 2007; 43:784 - 788
- Barron RM, Campbell SL, King D, Bellon A, Chapman KE, Williamson RA, et al. High titers of transmissible spongiform encephalopathy infectivity associated with extremely low levels of PrPSc in vivo. J Bio Chem 2007; 282:35878 - 35886
- McLeod AH, Murdoch H, Dickinson J, Dennis MJ, Hall GA, Buswell CM, et al. Proteolytic inactivation of the bovine spongiform encephalopathy agent. Biochem Biophys Res Com 2004; 317:1165 - 1170
- Scherbel C, Pichner R, Groschup MH, Mueller-Hellwig S, Scherer S, Dietrich R, et al. Infectivity of scrapie prion protein PrPSc following In vitro digestion with bovine gastrointestinal microbiota. Zoo Public Health 2007; 54:185 - 190
- Saa P, Castilla Jn, Soto C. Ultra-efficient replication of infectious prions by automated protein misfolding cyclic amplification. J Bio Chem 2006; 281:35245 - 35252
- Kurt TD, Perrott MR, Wilusz CJ, Wilusz J, Supattapone S, Telling GC, et al. Efficient in vitro amplification of chronic wasting disease PrPres. J Virol 2007; 81:9605 - 9608
- Atarashi R, Wilham JM, Christensen L, Hughson AG, Moore RA, Johnson LM, et al. Simplified ultrasensitive prion detection by recombinant PrP conversion with shaking. Nat Meth 2008; 5:211 - 212
- Atarashi R, Moore RA, Sim VL, Hughson AG, Dorward DW, Onwubiko HA, et al. Ultrasensitive detection of scrapie prion protein using seeded conversion of recombinant prion protein. Nat Meth 2007; 4:645 - 650
- Onisko B, Dynin I, Requena J, Silva C, Erickson M, Carter J. Mass spectrometric detection of attomole amounts of the prion protein by nanoLC/MS/MS. J Am Soc Mass Spec 2007; 18:1070 - 1079
- Arthur WJ, Alldredge AW. Soil ingestion by mule deer in northcentral Colorado. Rang Ecol Manage 1979; 32:67 - 71
- Beyer WN, Connor EE, Gerould S. Estimates of soil ingestion by wildlife. J Wildlife Manage 1994; 58:375 - 382
- Johnson CJ, Phillips KE, Schramm PT, McKenzie D, Aiken JM, Pedersen JA. Prions ddhere to soil minerals and remain infectious. PLoS Pathog 2006; 2:296 - 302
- Genovesi S, Lieta L, Sequi P, Andrighetto I, Sorgato MC, Bertoli A. Direct detection of soil-bound prions. PLoS ONE 2007; 2:1069
- Revault M, Quiquampoix H, Baron MH, Noinville S. Fate of prions in soil: Trapped conformation of full-length ovine prion protein induced by adsorption on clays. Biochim Biophys Acta 2005; 1724:367 - 374
- Vasina EN, Dejardin P, Rezaei H, Grosclaude J, Quiquampoix H. Fate of prions in soil: Adsorption kinetics of recombinant unglycosylated ovine prion protein onto mica in laminar flow conditions and subsequent desorption. Biomacromolecules 2005; 6:3425 - 3432
- Rapp D, Potier P, Jocteur-Monrozier L, Richaume A. Prion degradation in soil: Possible role of microbial enzymes stimulated by the decomposition of buried carcasses. Environ Sci Technol 2006; 40:6324 - 6329
- Rigou P, Rezaei H, Grosclaude J, Staunton S, Quiquampoix H. Fate of prions in soil: Adsorption and extraction by electroelution of recombinant ovine prion protein from montmorillonite and natural soils. Environ Sci Technol 2006; 40:1497 - 1503
- Cooke CM, Rodger J, Smith A, Fernie K, Shaw G, Somerville RA. Fate of prions in soil: detergent extraction of PrP from soils. Environ Sci Technol 2007; 41:811 - 817
- Cooke CM, Shaw G. Fate of prions in soil: longevity and migration of recPrP in soil columns. Soil Biol Biochem 2007; 39:1181 - 1191
- Rao MA, Russo F, Granata V, Berisio R, Zagari A, Gianfreda L. Fate of prions in soil: interaction of a recombinant ovine prion protein with synthetic humic-like mineral complexes. Soil Biol Biochem 2007; 39:493 - 504
- Polano M, Anselmi C, Leita L, Negro A, Nobili MD. Organic polyanions act as complexants of prion protein in soil. Biochem Biophys Res Com 2008; 367:323 - 329
- Pucci A, D'Acqui LP, Calamai L. Fate of prions in soil: Interactions of recPrP with organic matter of soil aggregates as revealed by LTA-PAS. Environ Sci Technol 2008; 42:728 - 733
- Ma X, Benson CH, McKenzie D, Aiken JM, Pedersen JA. Adsorption of pathogenic prion protein to quartz sand. Environ Sci Technol 2007; 41:2324 - 2330
- Kirchmayr R, Reichi HE, Schildorfer H, Braun R, Somerville RA. Prion protein: detection in ‘spiked’ anaerobic sludge and degradation experiments under anaerobic conditions. Water Sci Technol 2006; 53:91 - 98
- Leita L, Fornasier F, Nobili MD, Bertoli A, Genovesi S, Sequi P. Interactions of prion proteins with soil. Soil Biol Biochem 2006; 38:1638 - 1644
- Maluquer de Motes C, Cano MJ, Torres JM, Pumarola M, Girones R. Detection and survival of prion agents in aquatic environments. Water Res 2008;
- Saunders SE, Bartz JC, Telling GC, Bartelt-Hunt SL. Environmentally-relevant forms of the prion protein. Environ Sci Technol 2008; 42:6573 - 6579
- Hinckley GT, Johnson CJ, Jacobson KT, Bartholomay C, McMahon KD, McKenzie D, et al. Persistence of pathogenic prion protein during simulated wastewater treatment processes. Environ Sci Technol 2008; 42:5254 - 5259
- Huang H, Spencer JL, Soutryine A, Guan J, Rendulich J, Balachandran A. Evidence of degradation of abnormal prion protein in tissues from sheep with scrapie during composting. Can J Vet Res 2007; 71:34 - 40
- McKinley MP, Meyer RK, Kenaga L, Rahbar F, Cotter R, Serban A, et al. Scrapie prion rod formation in vitro requires both detergent extraction and limited proteolysis. J Virol 1991; 65:1340 - 1351
- González L, Martin S, Begara-McGorum I, Hunter N, Houston F, Simmons M, et al. Effects of agent strain and host genotype on PrP accumulation in the brain of sheep naturally and experimentally affected with scrapie. J Comp Pathol 2002; 126:17 - 29
- González L, Martin S, Jeffrey M. Distinct profiles of PrPd immunoreactivity in the brain of scrapie- and BSE-infected sheep: implications for differential cell targeting and PrP processing. J Gen Virol 2003; 84:1339 - 1350
- Jeffrey M, Martin S, González L. Cell-associated variants of disease-specific prion protein immunolabelling are found in different sources of sheep transmissible spongiform encephalopathy. J Gen Virol 2003; 84:1033 - 1045
- Kuczius T, Grassi J, Karch H, Groschup MH. Binding of N- and C-terminal anti-prion protein antibodies generates distinct phenotypes of cellular prion proteins (PrPc) obtained from human, sheep, cattle and mouse. FEBS J 2007; 274:1492 - 1502
- Pan T, Wong PK, Chang BG, Li CY, Li RL, Kang SC, et al. Biochemical fingerprints of prion infection: Accumulations of aberrant full-length and N-terminally truncated PrP species are common features in mouse prion disease. J Virol 2005; 79:934 - 943
- Frankenfield KN, Powers ET, Kelly JW. Influence of the N-terminal domain on the aggregation properties of the prion protein. Protein Science 2005; 14:2154 - 2166
- Crozet C, Flamant F, Bencsik A, Aubert D, Samarut J, Baron T. Efficient transmission of two different sheep scrapie Isolates in transgenic mice expressing the ovine PrP gene. J Virol 2001; 75:5328 - 5334
- Scott MR, Safar J, Telling G, Nguyen O, Groth D, Torchia M, et al. Identification of a prion protein epitope modulating transmission of bovine spongiform encephalopathy prions to transgenic mice. PNAS 1997; 94:14279 - 14284
- Silveira JR, Raymond GJ, Hughson AG, Race RE, Sim VL, Hayes SF, et al. The most infectious prion protein particles. Nature 2005; 437:257 - 261
- Quiquampoix H, Burns RG. Interaction between proteins and soil mineral surfaces: environmental and health consequences. Elements 2007; 3:401 - 406
- Brash JL, Horbett TA. Brash JL, Horbett TA. Proteins at interfaces: an overview. Proteins at Interfaces II: Fundamentals and Applications 1995; Washington, DC American Chemical Society 1 - 21
- Quiquampoix H, Abadie J, Baron MH, Leprince F, Matumoto-Pintro PT, Ratcliffe RG, et al. Horbett TA, Brash JL. Mechanisms and consequences of protein adsorption on soil mineral surfaces. Proteins at Interfaces II: Fundamentals and Applications 1995; Washington, DC American Chemical Society 321 - 333
- Harter RD, Stotzky G. X-ray diffraction, electron microscopy, electrophoretic mobility and pH of some stable smectite-protein complexes. Soil Sci Soc Am Proc 1973; 37:116 - 123
- Noinville S, Revault M, Quiquampoix H, Baron MH. Structural effects of drying and rehydration for enzymes in soils: a kinetics-FTIR analysis of alpha-chymotrypsin adsorbed on montmorillonite. J Colloid Interface Sci 2004; 273:414 - 425
- Taylor DM, McConnell I, Fernie K. The effect of dry heat on the ME7 strain of mouse-passaged scrapie agent. J Gen Virol 1996; 77:3161 - 3164
- de Jonge LW, Kjaergaard C, Moldrup P. Colloids and Colloid-Facilitated transport of contaminants in soils: an introduction. Vadose Zone J 2004; 3:321 - 325
- Slack SM, Horbett TA. Brash JL, Horbett TA. The Vroman Effect. Proteins at Interfaces II: Fundamentals and Applications 1995; Washington, DC American Chemical Society 112 - 128
- Yoshioka M, Miwa T, Horii H, Takata M, Yokoyama T, Nishizawa K, et al. Characterization of a proteolytic enzyme derived from a Bacillus strain that effectively degrades prion protein. J Appl Microbiol 2007; 102:509 - 515
- Scherbel C, Richner R, Groschup MH, Mueller-Hellwig S, Scherer S, Dietrich R, et al. Degradation of scrapie associated prion protein (PrPSc) by the gastrointestinal microbiota of cattle. Vet Res 2006; 37:695 - 703
- Brown P, Rau EH, Lemieux P, Johnson BK, Bacote AE, Gajdusek DC. Infectivity studies of both ash and air emissions from simulated incineration of scrapie-contaminated tissues. Environ Sci Technol 2004; 38:6155 - 6160
- Triantis J, Dennis MM, Salman MD, Gould DH. Effect of time and temperature on PrPCWD immunoreactivity as evidenced by western blot. J Vet Diagn Invest 2007; 19:389 - 391
- Epstein E, Beecher N. Mad cow disease, Creuzfeld-Jakob disease, other TSEs and biosolids. J Residual Sci Technol 2005; 2:181 - 187
- Outreach and Education Subcommittee WRBC. Prion potential in biosolids. Water Environ Res 2006; 78:339