Abstract
Phenolic acids are the main polyphenols made by plants. These compounds have diverse functions and are immensely important in plant-microbe interactions/ symbiosis. Phenolic compounds act as signaling molecules in the initiation of legume-rhizobia symbioses, establishment of arbuscular mycorrhizal symbioses and can act as agents in plant defense. Flavonoids are a diverse class of polyphenolic compounds that have received considerable attention as signaling molecules involved in plant-microbe interactions compared to the more widely distributed, simple phenolic acids; hydroxybenzoic and hydroxycinnamic acids, which are both derived from the general phenylpropanoid pathway. This review describes the well-known roles attributed to phenolic compounds as nod gene inducers of legume-rhizobia symbioses, their roles in induction of the GmGin1 gene in fungus for establishment of arbuscular mycorrhizal symbiosis, their roles in inducing vir gene expression in Agrobacterium, and their roles as defense molecules operating against soil borne pathogens that could have great implications for rhizospheric microbial ecology. Amongst plant phenolics we have a lack of knowledge concerning the roles of phenolic acids as signaling molecules beyond the relatively well-defined roles of flavonoids. This may be addressed through the use of plant mutants defective in phenolic acids biosynthesis or knock down target genes in future investigations
Introduction
Phenolic acids and their derivatives are a diverse class of phenolic compounds made by plants. They are known to play multifunctional roles in rhizospheric plant-microbe interactions.Citation1 Phenolic acids are produced in plants via shikimic acid through the phenylpropanoid pathway, as by-products of the monolignol pathway and as breakdown products of lignin and cell wall polymers in vascular plant.Citation2–Citation5 Additionally, some phenolic acids are of microbial origin.Citation6 The phenolic acids found in plant cell walls and lignin have a unique chemical structure of C6-C3 (phenylpropanoid type) whereas those of microbial origin are of the form C6-C1 (Phenylmethyl type).Citation7
In response to microbial attack, plants activate defense responses that lead to induction of a broad spectrum of antimicrobial compounds some of which may be species specific.Citation8 These induced defense mechanisms are expressed at site of attack (hypersensitive response) as well as at a distance (signaled by methyl salisylate) to the site of primary infection and protect the plant from the spread of infection and future attack. Induced resistance is regulated by a network of interconnecting signal transduction pathways in which phenolic acids are key signaling molecules.Citation9,Citation10 Phenolic acids are relatively resistant biochemical species; they undergo transformation in the soil because some microorganisms have the capacity to utilize them as carbon sources.Citation11 It is well established that several soil bacteria have the ability to oxidize aromatic compounds.Citation12,Citation13 Simple phenolic compounds, such as methoxy and hydroxy benzoic acid and cinnamic acids, are commonly formed in decaying plant residues.Citation14 Phenolic acids may provide alternative carbon sources for some diazotrophs in limited environmentsCitation15 and may also serve as precursors for the synthesis of phenolic lipids.Citation16
Mycorrhizae are formed in symbiotic interactions between plants and fungi. Growth promotion in plants can be attributed to arbuscular mycorrhizal fungal (AMF) associations in some situations such as macronutrient-limited soils, because mycorrhizal plants grow better than non-mycorrhizal plants.Citation17 Considerable increases in phenolic compounds in host plants as a result of arbuscular mycorrhizal (AM) fungus inoculation have been reported.Citation18 Generally, inoculation of plants with AMF results in an overall increase in the production of some new phenolic compounds during the progression of the infection.Citation19
In legumes, phenolic acids are released rapidly from emerging roots during seed germination and seedling growth.Citation20,Citation21 The Rhizobium community in the rhizosphere undergoes changes in response to phenolic acids when they accumulate in the soil which provides a competitive advantage for nodulation by selective rhizobial strains.Citation22 It is well documented that Rhizobia species have the ability to utilize phenolic acids as a carbon source.Citation23,Citation24 A range of soluble and conjugated phenolic acids involved in rhizobial defense and nodule morphogenesis have been detected in roots and root nodules of Arachis hypogaea L.Citation25 Recently, it has been shown that some endogenous phenolic acids present in root nodules of Vigna mungo stimulate the efficiency of IAA production by its symbionts (Rhizobium sp.) and regulate nodule morphogenesis.Citation26 Over the last few decades, various functions for these phenolic compounds in root nodules have been investigated (). Microbes and plants have evolved complex signal exchange mechanisms that allow a specific bacterial species to induce its host plant to form invasion structures through which the bacteria can enter the plant root or shoot. This review aims to summarize: how rhizosphere microbial communities impact phenolic signaling and how metabolic plasticity in microsymbionts alters the symbiotic morphogenesis that follows phenolic signaling?
Biosynthesis of Phenolic Acids
Plants produce an extremely diverse array of low molecular mass compounds, often called secondary metabolites or ‘natural products’ that bestow metabolic plasticity essential for anticipating and responding to biotic and abiotic stress(es). Such metabolites are generally derived from isopropanoid, phenylpropanoid, alkaloid or fatty acid pathways.Citation27 All terpenoids are synthesized from the five carbon precursor, isopentanyl diphosphate. Alkaloids are synthesized principally from amino acids and phenolics are derived from either the shikimic acid pathway or the malonate acetate pathway or both. Phenolic acids, characterised by hydroxylated aromatic ring(s) are ubiquitous secondary metabolites in plants and provide one of the most studied and widely exploited metabolic pathways in plant research.Citation28,Citation29
The shikimic acid pathway is involved in the synthesis of a majority of the phenolic compounds in plants fungi and bacteria converting simple carbohydrate precursors derived from glycolysis and the pentose phosphate pathway derived to the aromatic amino acids, phenylalanine and tryptophan (). The malonic acid pathway is of less significance in the formation of phenolic acids in higher plants compared to fungi and bacteria. Flavonoids, biosynthetically derived from malonyl CoA and p-coumaroyl CoA, derived from acetate and shikimate respectively, are the largest single group of phenolic C15 compounds composed of two phenolic rings connected by a three carbon unit.Citation30 They may be considered as phenolic acid derivatives.
First described by Koukol and Conn,Citation31 phenylalanine ammonia lyase (PAL) mediates the formation of cinnamic acid from phenylalanine; is a pivotal branch point of primary and secondary metabolism and is the first and most important regulatory step in the formation of many phenolic acids. The production of phenolics is modulated in response to various stimuli by PAL gene expression and protein activityCitation32 by mechanisms involving feedback regulation,Citation33 post transcriptional changesCitation34 and metabolite channeling.Citation35,Citation36 It has been shown in transgenic tobacco that the subcellular localization of two PAL isoforms (PAL 1 and PAL 2) is different and depends on the complex between cinnamate 4-hydroxylase (C4H) and PAL 1 which is believed to help partitioning phenylpropanoid biosynthesis into different branch pathways by differential subcellular distribution of cinnamic acid.Citation37 However in yeast, no interaction was detected in effective channeling of carbon through PAL to p-coumaric acid.Citation38 The PAL-mediated phenolic pathways of general phenylpropanoid metabolism have been studied extensively and do not fall within the preview of this review. However the recent reports of the presence of PAL homologs in prokaryotes do deserve special mention.Citation39,Citation40 The general absence of phenylpropanoids and flavonoids in bacteria was attributed to the absence of the enzymes, PAL and chalone synthase. PAL has been encountered in a few prokaryotes such as the marine bacterium Streptomyces maritimes,Citation39 Sorangium cellulosumCitation41 and Streptomyces verticillatus.Citation42 PAL products are a key component in the benzoyl-coenzyme A pathway in S. maritimus for the production of benzoate—primed polyketides enterocin and wailupemycin G.Citation43 Disruption of the encP gene in S. maritimus inhibited the production of cinnamate and enterocin.Citation40 Enterocins are charecterized by their broad range of activity against gram positive bacteria and play an important role in maintaining bacterial community structures.Citation44–Citation46 PAL proteins have also been reported recently from cyanobacteria that are similar in tertiary and quarternary structure to plant and yeast PALs and associate with secondary metabolite biosynthetic gene clusters as observed for other eubacterial PAL genes.Citation47 Prokaryotic PALs have been suggested as a viable alternative to plant and yeast PAL in enzyme substitution therapy in patients suffering from phenylketonuria.Citation47 Evidence for the existence of phenylpropanoid pathway and flavonoid biosynthesis in the industrially important fungus Aspergillus oryzaeCitation48 raises new questions about uv light treatment in decontamination of food products as well as opening new avenues for improved production of important and novel metabolites.Citation49 The superfamily of ammonia lyases also includes histidine ammonia lyase (HAL), common in both prokaryotes and eukaryotes catalyzing deamination of L-His in the histidine degradation pathway and tyrosine ammonia lyase (TAL) that specifically deaminates L-Tyr to p-coumarate.Citation50 Recent research has extended the understanding of the enzymes of phenylpropanoid metabolism including the upstream enzymes instrumental in recruiting PAL and TAL, a number of reductases, acyltransferases and associated enzymes tailoring phenylpropanoid derived metabolites providing experimental templates for enzyme and metabolite engineering.Citation51
Plant cell walls are composed of polysaccharides contained in the microfibrillar and amorphous phases: cellulose, hemicellulose and pectic substances, as well as the other components: lignin, proteins and enzymes.Citation52 Lignin is crucial for structural integrity of the cell wall and the most abundant biopolymer after cellulose.Citation53 The biosynthesis of lignin involves hydrogenative polymerization of monolignols.Citation54 Monolignol biosynthesis starts with the deamination of phenylalanine and although the pathway is still debated, it involves the phenolic acids p-coumaric, ferulic and sinapic acid and a number of enzymes including caffeoyl-CoA O-methyltransferase.Citation55 Lignin plays an important role in protecting plants against pathogens. Lignin transformation and decomposition products are generally considered a major source of stable soil organic matter but this process remains poorly understood.Citation56,Citation57 Lignin is considerably resistant to microbial degradation. Some white rot basidomycetes like Coriolus versicolor, Pleurotus eryngii, Phlebia radiata are typically lignin degrading.Citation58 Complete degradation of lignin model compounds have also been reported by bacteria like Pseudomonas sp.Citation59 Lignin degradation by the white rot fungi is a complex process mediated by the action of several extra cellular enzymes, of which lignin peroxidases are the most important.Citation60 Ligninolytic micro-organisms are important in agriculture and industry with a wide tolerance of temperature, pH, oxygen limitation and identification of such organisms from soils by techniques such as DNA fingerprinting is increasingly being applied.Citation61
Phenolic acids are incorporated to cell wall of plants in response to biotic stress with increased flux through the phenylpropanoid pathway resulting in synthesis of cinnamic acid and benzoic acid derivatives that are esterified and incorporated in the cell wall fraction.Citation62 A significant portion of the phenolic acids present in the plants are in conjugated form, principally with a sugar residue linked through one or more of the phenolic hydroxyl groups, or as conjugated esters. Acid or alkali hydrolysis releases a number of soluble phenolic acids that are either associated with lignin or as simple glycosides.Citation63 Phenolics from root and seed exudates, leaf leachetes, decaying plant matter play multiple roles in soil formation and pedogenesis influencing mineral elements and organic matter dynamics.Citation64,Citation65 Soil characteristics like pH inturn are important factor in the release of phenolic acid in root exudates. It is reported that at acidic pH, nitrate reduces phenolic exudation and at pH 7.5, it becomes restricted to some root zones.Citation66 Green leaves and decomposing litter can influence rhizosphere nitrogen through phenolics such as chlorogenic acid.Citation67 There is increasing evidence that some plants may directly modify rhizosphere to gain access to unavailable soil nitrogen and phosphorus reserves.Citation68,Citation69 Phenolic exudates are also reported to increase the availability of micro and macronutrients by formation of organic metal complexes.Citation70
Rhizobium-Legume Symbiosis
The root nodule is a unique and highly organized structure developed as a result of the symbiotic relationship between leguminous plants and bacteria of the genus Rhizobium. Within the root nodule, the invading bacteria (Rhizobium sp.) differentiate into nitrogen-fixing bacteroids that provide reduced nitrogen to the plant in exchange for carbohydrates and shelter.Citation71,Citation72 Different aspects of the physiology of legume-Rhizobium symbiosis and mechanism of nitrogen fixation have been extensively investigated by various workers.Citation73,Citation74 The establishment of the symbiosis requires signaling and recognition by both the partners; various signaling molecules are exchanged between the plant and the infecting bacteria to regulate nodule initiation, differentiation and functioning. Thus, many facets of plant-bacterium recognition, nodule formation and nitrogen fixation have been well studied,Citation75,Citation76 but the mechanisms by which root nodules phenylpropanoids regulate the infection efficiency of rhizobia for nodule formation has not been studied in detail. Recent studies in A. hypogaea indicate a temporal and spatial differentiation in the accumulation and expression of phenolic acids during nodulation.Citation25
The process of nodule formation in legumes involves the production by the plant of flavonoids, betaines and aldonic acids in its seed and root exudates as signals to the microbial symbiont.Citation77 Although phenolic acids are the main polyphenols involved, their regulation has not received the same attention as flavonoids. Some phenolic compounds from symbiotic legumes are known to promote growth of rhizobial bacteria in the rhizosphere78 and also to serve as chemoattractants that guide rhizobial cells to legume root hairs.Citation79 Fox et al.Citation80 demonstrated that planar phenolic compounds with free hydroxyl groups can interfere with legume-rhizobium flavonoid signalling, and alter the nitrogen fixing symbiosis. Flavonoid production is also induced by rhizobia in roots and nodulesCitation81 which regulate Nod (nodulation) factor production prior to and during infection.Citation82 It is possible that flavonoids like phenolic acids protect dividing cells from oxidative damage because of their activity as antioxidants.Citation83 Leguminous plants secrete a variety of phenolic compounds from roots such as flavones and flavonols from Vicia faba,Citation20 isoflavonoids from soybean,Citation84 and vanillin from peanutCitation85 to mention a few. These phenolic compounds regulate nod gene expression by the symbiont (Rhizobium) and so modify the legume-rhizobial symbiosis. The host root secretes phenolic compounds that act as signaling molecule during expression of various symbiotic plasmidencoded nod (nodulation) genes. Some of these nod genes encode enzymes to synthesize a special class of glycolipids (chitolipooligosaccharides). These signal molecules vary somewhat in structure, but their non-reducing end, which contains an N-acyl long-chain fatty acid, is bioactive in the plant host, triggering root hair deformations and cortical cell divisions within the root leading to nodule formation.
It has been reported that in the co-culture of legumes and Rhizobium collectively play a role in the symbiosis.Citation85,Citation86 Some phenolic acids stimulate while others repress nod gene expression in Rhizobium trifolii.Citation87 Seneviratne and JayasinghearachchiCitation64 have suggested a possible mechanistic explanation for the effects of phenolic acids on the protein profiles of Rhizobium. They showed that the strains are initially induced to produce specific enzymes that are capable of degrading the phenolic acid and interaction of phenolic acids with the rhizobial nod gene expression contributes to changes in the protein patterns. The effect of phenolic acids is concentration, and structure dependant, and strain-specific. These potential interactions have implications in modulating nodulation and in establishing the symbiosis ().
The Arbuscular Mycorrhiza Symbiosis
Arbuscular mycorrhizas (AM) are characterized by the formation of unique arbuscules and vesicles by fungi of the phylum Glomeromycota. Growth promotion is attributed to arbuscular mycorrhizal (AM) fungal association in most situations, where mycorrhizal plants grow better than non-mycorrhizal plants, a feature attributed to increased nutrient acquisition (particularly phosphorus) from soilCitation88,Citation89 and better water uptake.Citation90 The successful development of the symbiotic association between AM fungi and their host plants depends on three main steps (a) spore germination, (b) hyphal growth and (c) host recognition and formation of appressoria.Citation91 There is increasing evidence showing that fungus and plant start the recognition process long before the first colonization structures appear on the root epidermis. It has been known for a long time that only compatible host roots are able to elicit hyphal branching.Citation92,Citation93 Earlier evidence showed that exudates from host roots elicit growth stimulation in AM in contrast to non-host exudates.Citation93–Citation95
Plant phenolic compounds are potential candidates as signals during mycorrhizal formation. Some reports show that exogenous application of flavonoids exerts a positive effect on hyphal growth during symbiosis.Citation94,Citation96,Citation97 These effects range from increased spore germination to enhanced hyphal growth, hyphal branching and formation of secondary spores. It has been shown that the AM mycorrhizal formation alters the flavonoid profile of root extracts through modifications to the expression of genes involved in phenylpropanoid, flavonoid and isoflavonoid metabolism.Citation98,Citation99 Flavonoids are specific to mycorrhizal symbiosis formation, a single flavonoid might exert a positive, negative or neutral effect on different fungi.Citation95,Citation97 This may be explained by specific effects of each flavonoid, as recently reported.Citation100,Citation101 Therefore it is likely that the role of flavonoids is limited to a stimulatory effect on AM fungal growthCitation102 thereby facilitating encounters by the fungus with host roots as has been suggested for the role of stimulating microorganisms. A comparative study of flavonoids in Trifolium repens has shown that infection with a mycorrhizal symbiont (Glomus intraradices) can significantly alter the composition of flavonoids accumulated in roots.Citation103
Several workers have observed considerable increases in phenolic compounds in the host as a result of AM inoculation.Citation18,Citation104 Giovannetti et al.Citation93 speculated that only the perception of the right chemical signals, coming from the roots of host plants, promotes differential morphogenesis of AM hyphae in the rhizosphere and that root cell penetration by the fungus depends on the host genome. Therefore, phenolic compounds could enhance the initial stages of AM establishment, but root penetration and AM development are likely regulated by the host plant and subsequent interactions with the fungal partner. Fries et al.Citation105 demonstrated the stimulatory effect of p-coumaric acid, p-hydroxybenzoic acid, or quercetin on growth and colonization of clover (Trifolium repens L. cv. Ladino) and sorghum (Sorghum bicolor L.) roots by the arbuscular mycorrhizal (AM) fungus Glomus. They hypothesized that phenolic compounds, commonly found in many soils, influence the establishment of AM symbioses, stimulated plant growth and AM colonization at low concentration level of phenolic acids. However, the role of flavonoids and other phenolic acids in the vesicular-arbuscular mycorrhizal association is less well understood.
Apart from flavonoids, strigolalactones—a group of sesquiterpene lactones derived from the caroteinoid pathwayCitation106 known to be germination inducers in parasitic plants, have recently been suggested to play a key role in establishment of AM symbiosis.Citation107 They are present in exudates of plants from diverse taxa and are proposed to be essential signals in establishment of AM association, hyphal branching and directional growth of AMF (arbuscular mycorrhizal fungi) towards root.Citation108 Strigolactones may provide new avenues in agriculture for the management of parasitic weeds and beneficial fungal symbionts.Citation109,Citation110
Molecular Response of Microsymbiontsto Phenolic Acids
Phenolic compounds strongly regulate nod gene expression of the symbiont (Rhizobium) and modify the legume-rhizobial symbiosis.Citation75 Rhizobium releases signal molecules by induction of nodulation (nod) genes that control root nodule organogenesis. The expression of these genes is regulated by the nodD activator proteins.Citation111 Rhizobia in turn, respond to the phenolic signal by releasing lipo-chitooligosaccharide Nod factors that cause morphological changes in legume root hairs, leading to infection thread formation, nodule development and symbiotic N2 fixation. Flavonoids are secreted by different root zones and produce divergent effects on nod-gene expressionCitation112 and alter nodule organogeneis. The flavonoids released from the plant roots bind to the transcriptional activation sites of Rhizobial nod genes (NodD1/NodD2/NodD3), and the products of these genes, in turn, activate transcription of other nod genes. The NodD proteins are localized in the cytoplasmic membrane and interact with flavonoids in the inner membrane.Citation113 Exchange of a nodD gene between Rhizobium strains that differ in their sensitivity to different flavonoids changes the sensitivity of the recipient strain to the flavonoid.Citation111,Citation114 Nod gene protein catalyzes synthesis of different nodulating factors which act to induce the first steps in root nodule initiation.
There is also evidence indicating that flavonoids affect cell division either by regulating auxin transport or its turnover, (thereby regulating auxin accumulation) or by direct involvement in cell cycle regulation.Citation115 Prinsen et al.Citation116 have shown that some flavonoids increase IAA production by the symbiont (Rhizobium sp.) in vitro and suggested that these flavonoids may exert this effect through the synthesis of nod signal molecules. Theunis et al.Citation117 have demonstrated that flavonoids secreted by host plants, activate expression of the nod gene locus, y4wEFG. This gene is located downstream of a nod-box, NB15, and encodes proteins involved in IAA synthesis by Rhizobia, which trigger nodule organogenesis. Flavonoids have also been suggested as potential candidates for endogenous inhibitors of auxin transport in the early stage of nodule initiation.Citation118 Studies of auxin import carrier (AUX1) genes in Medicago truncatula indicate that nodule formation requires high auxin levels to initiate the process of cell division and the establishment of the nodule primordium.Citation119
Recently, we observed that some phenolic acids found in root nodules increase the efficiency of IAA production by Rhizobia.Citation26 Further studies should therefore be conducted to evaluate the effects of these compounds on symbiosis, nod-gene expression and nodulation efficiency. The bacterium Agrobacterium tumefaciens has been grouped with Rhizobium into the family Rhizobiaceae,Citation120 and causes crown gall tumors after infecting the wound sites of mostly dicotyledonous and a few of monocotyledonous plants. A specific segment of the Ti (tumor-inducing) plasmid, the T-DNA, is transferred to host plant cells and is then incorporated into the plant nuclear genome. The virulence (vir) genes located on the Ti plasmid, are involved specifically in the processing and transfer of the T-DNA. The vir genes are transcriptionally regulated by two members, virA and virG.Citation121 The VirA protein responds to plant signal molecules and then transduces the signal to the VirG protein, a response regulator. This protein then binds to upstream regions of each of the vir genes and activates transcriptionally vir gene expression. Vir genes are induced at an acidic pH by phenolic compounds that function in concert with monosaccharides synthesized and exuded from wounded plant cells. Lee et al.Citation122,Citation123 clearly stated that the virA locus determines which phenolic compounds can function as vir gene inducers and the VirA protein directly senses the phenolic compounds for vir gene activation.
Only a few studies have identified the fungal transcriptional changes induced by plant signals.Citation124,Citation125 One important limitation for those studies is low amount of fungal biomass within the root, especially during the colonization phase. Requena et al.Citation126 identified few fungal genes regulated during development, used suppressive subtractive hybridization (SSH) to create a subtractive cDNA library from G. mosseae. They identified a novel gene (GmGin1) encoding a two-domain protein that is downregulated in expression upon entry into symbiosis. These authors suggested that GmGin1 could be a sensor for plant signals. This protein is located at the cell membrane, by means of its carboxy terminus, which undergoes splicing in response to signals from the plant. After splicing, the amino terminus remains covalently attached to the plant signal acting as nucleophile. Requena et al.Citation127 suggested that that a modified Gin1 is able to exert a signaling function through its ATPase activity and modulate other downstream signaling proteins. They also showed that chemical communication with the plant symbiont modified fungal gene expression as well as induce post-transcriptional modification of fungal proteins to facilitate the development of successful symbioses.
Phenolic Acid as Defense Molecules
ChesterCitation128 observed that plants possess defense mechanisms that they use in response to the attack of pathogens. Secondary metabolites play important roles either as local or systemic resistance factors in protecting the plants against various pathogens.Citation129–Citation131 Phenolic compounds play a major role in the induction of resistance in plants. Generally, phenolic compounds released from seeds, roots or residue decomposition can act against soil borne pathogens and root-feeding insects.Citation132 Roots are a rich source of specific natural products that contribute to the competitiveness of invasive plant species and have a marked effect plant and soilborne organisms.Citation133,Citation134 Several studies have shown that plant defense against soil born pathogens, nematodes, phytophagous insects is based on the release and accumulation of phenolic compounds in soil.Citation135,Citation136 The activity of particular phenolic compounds depends on their structural diversity. For example, simple and complex phenolic compounds such as cajanin, medicarpin, glyceolin, rotenone, coumestrol, phaseolin, phaseolinin, isoflavonoid, flavonoids act as phytoalexins, phytoanticipins and nematicides against soilborne pathogens and phytophagous insects.Citation132,Citation136 Several phenolic acids possess high antifungal activity.Citation137,Citation138 Phenolic compounds can offer an alternative to the chemical control of pathogens on agricultural crops.Citation136 Accumulation of phenolic compounds at the challenge site also reinforces cell wall which is accompanied by localized production of reactive oxygen species driving cell wall cross linking, antimicrobial activity and defense signaling.Citation139 The presence of microorganisms undoubtedly influences the quality and quantity of flavonoids present in the rhizosphere, both through modification of root exudation patterns and microbial catabolism of exudates. Microbial alteration and attenuation of the signals of phenolic compounds may have ecological consequences for plant-microbe interaction.Citation140
Plant phenolic compounds produced during host-pathogen interactions work by several mechanisms in plant defense.Citation27,Citation141 It has been suggested that the alteration of flavonoid profiles in response to AMF colonization may be a result of initiation of a general plant defense response which is later suppressed.Citation142 Rao and CooperCitation143 have demonstrated that the potential consequences of microbial transformations of pre-existing phenolic compound pools, namely the production of de novo flavonoids which are either nod gene inducers or repressors and also induce rhizobial resistance toward phytoalexins, or the formation of mono-cyclic hydroxy aromatic metabolites which could have implications for competition for nodule occupancy, and chemotactic responses. Such natural defense mechanisms provided by these biomolecules in the rhizosphere deserves more scientific attention because of its dual ecological potential as a sustainable means of reducing soil borne infections and for increasing the soil fertility in the ecosystems. Interestingly, rhizobia induce a number of defense mechanisms in planta thereby conferring increased disease resistance.Citation144 Plant growth promoting rhizobacteria (PGPR) are increasingly being used in agriculture with potential beneficial effects on plant growth while limiting deleterious effects of phytopathogens by the production of antimicrobials.Citation145 Recent advances in genomic research provide vital clues to the enigma of legume—rhizobia recognition by antimicrolbial molecules.Citation146
Future Perspectives and Conclusions
Within the last few decades, strong evidence supporting the roles of phenolic compounds in the establishment of plant-microbe symbioses has been published. Scientists have been focused mostly on the regulation by flavonoid signaling rather than by phenolic acids, although phenolic acids are the major polyphenols in most plants, especially in roots. Obviously there is a complex interrelationship involving phenolic acids and their derivatives such as flavonoids, and the ecology of the plant-microbe symbiosis system. So, microbial attenuation or alteration of phenylpropanoid signals may be an important aspect of rhizosphere ecology and in the establishment of symbiosis. Molecular biological research on plant-microbe symbiosis has addressed several questions about the interaction: (a) how do phenolic acids alter the Rhizobium-legume symbiosis—does this involve binding to transcriptional activation sites (NodD1/NodD2/NodD3) of nod genes, in the same way as is thought to occur for flavonoids? () (b) What effects does phenolic acid induction have on the expression (downregulation or upregulation) of nod cluster? (c) Does the catabolism of phenolic acids or their derivatives in the rhizosphere catabolism have effects on the Rhizobium-legume symbiosis? (d) Is there any effect of phenolic acids on nutritional exchange during AM-plant symbiosis? (e) How is the GmGin1 gene in fungus regulated by altering the functional groups of phenolic acids? Regarding the impact of phenolic acids at the plant-microbe symbiosis level, the simplest way to address these questions would be through collective action among researchers to define in detail the genetic, biochemical and physiological parameters of the interactions that involve phenolic acids. Through the use of plant mutants defective in phenolic acid biosynthesis or knock-out mutants of target genes we can also start to address the question of the significance of phenolic acids in plant-microbe symbioses. Major goals in future research will be to identify networks of signals and receptors that provide the gateway to the establishment of plant-microbe symbioses.
Figures and Tables
Figure 1 Summarized phenolic acids and flavonoids branch of the phenylpropanoid biosynthetic pathway.
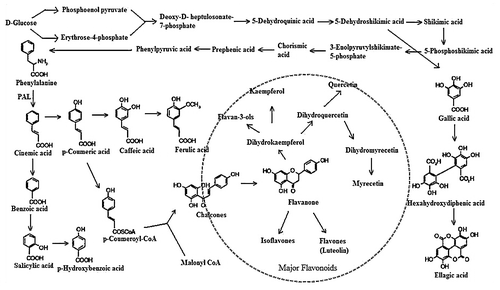
Figure 2 Diagrammatic representation of phenolic compounds signal exchange between the legumes and rhizobia during nodulation. Plant signal activates the nodD product that controls the transcription of other nod genes—in the same way as is it occurs for phenolic acids? The nod boxes are highlighted with black boxes in the organization of nod gene cluster of Rhizobia and the arrows with different nod gene indicate the direction of transcription.
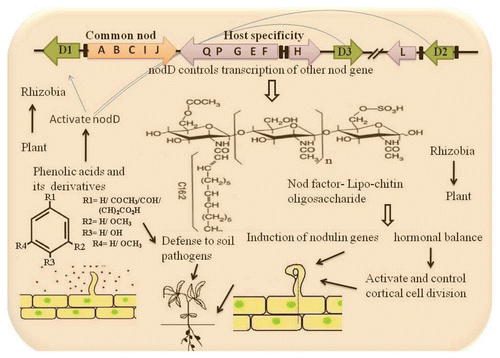
Table 1 Phenolic acids as inducer of plant-microbe symbioses in some selected species
Acknowledgements
We greatly acknowledge Prof. Cathe Martin, Department of Metabolic Biology, John Innes Centre, Colney, Norwich, NR4 7UH, United Kingdom, for her valuable insight, necessary correction and advice on this review. We would like to thank Sudipta Mandal Ghosh for references and figures editing in the manuscript.
References
- Martens DA. Relationship between plant phenolic acids released during soil mineralization and aggregate stabilization. Soil Sci Soc Am J 2002; 66:1857 - 1867
- Harkin JM. Butler GW, Bailey RW. Lignin (forage plants). Chemistry and biochemistry of herbage 1973; Academic Press New York 323 - 373
- Harborne JB. Bell EA, Charlwood BV. Plant phenolics. Encyclopedia of Plant Physiology, Secondary Plant Products 1980; 8:Berlin Heidelberg New York Springer-Verlag 329 - 395
- Carpita N, McCann M. Buchanan, et al. The cell wall. Biochemistry and molecular biology of plants 2000; Rockville MD American Society of Plant Physiologists 52 - 108
- Croteau R, Kutchan TM, Lewis NG. Buchanan, et al. Natural products (secondary metabolites). Biochemistry andmolecular biology of plants 2000; Rockville MD American Society of Plant Physiologists 1250 - 1318
- Moorman TB, Becerril JM, Lydon J, Duke SO. Production of hydroxybenzoic acids by Bradyrhizobiumjaponicum strains after treatment with glyphosphate. J Agric Food Chem 1992; 40:289 - 293
- Sarakanen KV, Ludwig CH. Lignins: occurance, formation, structure and reactions 1971; New York Wiley-Interscience
- van Loon LC. Slusarenko AJ, Fraser RSS, Van Loon LC. Systemic induced resistance. Mechanisms of resistance to plant diseases 2000; Dordrecht, The Netherlands Kluwer Academic Publishers 521 - 574
- Pieterse C, Van Loon LC. Salicylic acid-independent plant defence pathways. Trends Plant Sci 1999; 4:52 - 58
- Feys B, Parker JE. Interplay of signaling pathways in plant disease resistance. Trends Genet 2000; 16:449 - 455
- Kefeli VI, Kalevitch MV, Borsari B. Phenolic cycle in plants and environment. J Cell Mol Biol 2003; 2:13 - 18
- Chen YP, Dilworth MJ, Glenn AR. Uptake and oxidation of aromatic substances by Rhizobium leguminosarum MNF 8841 and Rhizobium trifolii TA1. FEMS Microbiol Lett 1984; 21:201 - 205
- Carmona M, Zamarro MT, Blázquez B, Durante-Rodríguez G, Juárez JF, Valderrama AJ, et al. Anaerobic catabolism of aromatic compounds: a genetic and genomic View. Microbiol Mol Biol R 2009; 73:71 - 133
- Whitehead DC, Dibb H, Hartley RD. Bound phenolic compounds in water extract of soils, plant roots and leaf litter. Soil Biol Biochem 1983; 15:133 - 136
- Chan YK. Utilization of simple phenolics for dinitrogen fixation by soil diazotrophic bacteria. Plant Soil 2006; 90:141 - 150
- Funa N, Ozawa H, Hirata A, Horinouchi S. Phenolic lipid synthesis by type III polyketide synthases is essential for cyst formation in Azotobacter vinelandii. Proc Natl Acad Sci USA 2006; 103:6356 - 6361
- Harley JL, Smith SE. Symbiosis 1983; London UK Academic Press
- Ling-Lee M, Chilvers GA, Ashford AE. A histochemical study of phenolic materials in mycorrhizal and uninfected roots of Eucalyptus fastigata Dean & Maid. New Phytol 1977; 78:313 - 328
- Devi MC, Reddy MN. Phenolic acid metabolism of groundnut (Arachis hypogaea L.) plants inoculated with VAM fungus and Rhizobium. Plant Growth Regul 2002; 37:151 - 156
- Bekkara F, Jay M, Viricel MR, Rome S. Distribution of phenolic compounds within seed and seedlings of two Vicia faba cvs differing in their seed tannin content, and study of their seed and root phenolic exudations. Plant Soil 1998; 203:27 - 36
- Staman K, Blum U, Louws F, Robertson D. Can simultaneous inhibition of seedling growth and stimulation of rhizosphere bacterial populations provide evidence for phytotoxin transfer from plant residues in the bulk soil to the rhizosphere of sensitive species?. J Chem Ecol 2001; 27:807 - 829
- Blum U, Stamen KL, Flint LJ, Shafer SR. Induction and/or selection of phenolic acid-utilizing bulk-soil and rhizosphere bacteria and their influence on phenolic acid phytotoxicity. J Chem Ecol 2000; 26:2059 - 2078
- van Rossum D, Schuuramns FB, Gillis M, Muyotcha A, van Versveld HW, Stouthamer AH, Boogerd FC. Genetic and phenetic analyses of Bradyrhizobium strains nodulating peanut (Arachis hypogaea L.) roots. Appl Environ Microbiol 1995; 61:1599 - 1609
- Irisarri P, Milnitsky, Monza J, Bedmar EJ. Characterization of rhizobia nodulating Lotus subbliforus from Uruguayan soil. Plant Soil 1996; 180:39 - 47
- Chakraborty D, Mandal SM. Fractional changes in phenolic acids composition in root nodules of Arachis hypogaea L. Plant Growth Regul 2008; 55:159 - 163
- Mandal SM, Mandal M, Das AK, Pati BR, Ghosh AK. Stimulation of indoleacetic acid production in a Rhizobium isolate of Vigna mungo by root nodule phenolic acids. Arch Microbiol 2009; 191:389 - 393
- Dixon RA. Natural products and plant disease resistance. Nature 2001; 411:843 - 847
- Boudet AM. Evolution and current status of research in phenolic compounds. Phytochemistry 2007; 68:2722 - 2735
- Whiting DA. Natural phenolic compounds 1900–2000: a bird's eye view of a centuries chemistry. Nat Prod Rep 2001; 18:583 - 606
- Mann J. Secondary Metabolism 1978; Oxford Clarendon Press 316
- Koukol J, Conn EE. The metabolism of aromatic compounds in higher plants. IV Purification and properties of the phenylalaninedea minase of Hordeum vulgare. J Biol Chem 1961; 236:2692 - 2698
- Hahlbrock K, Scheel D. Physiology and molecular biology of phenylpropanoid metabolism. Plant Mol Biol 1989; 40:347 - 369
- Blount JW, Korth KL, Masoud SA, Rasmussen S, Lamb C, Dixon RA. Altering expression of cinnamic acid 4-hydroxylase in transgenic plants provides evidence for a feedback loop at the entry point into the phenylpropanoid pathway. Plant Physiol 2000; 122:107 - 116
- Cheng SH, Sheen J, Gerrish C, Bolwell GP. Molecular identification of phenylalanineammonia-lyase as a substrate of aspecific constitutively active Arabidopsis CDPK expressed in maize protoplasts. FEBS Lett 2001; 503:185 - 188
- Stafford HA. Possible multienzyme complexes regulating the formation of C6-C3 phenolic compounds and lignins in higher plants. Recent Adv Phytochem 1974; 8:53 - 79
- Rasmussen S, Dixon RA. Transgene-mediated and elicitor-induced perturbation of metabolic channeling at the entry point into the phenylpropanoid pathway. Plant Cell 1999; 11:1537 - 1552
- Achnine L, Blancaflor EB, Rasmussen S, Dixon RA. Colocalization of L-phenylalanineammonia-lyase and cinnamate4-hydroxylase for metabolic channeling in phenylpropanoid biosynthesis. Plant Cell 2004; 16:3098 - 3109
- Ro DK, Douglas CJ. Reconstitution of the entry point of plant phenylpropanoid metabolism in yeast (Saccharomyces cerevisiae): implications for control of metabolic flux into the phenylpropanoid pathway. J Biol Chem 2004; 279:2600 - 2607
- Moore BS, Hertweck C, Hopke JN, Izumikawa M, Kalaitzis JS, Nilsen G, et al. Plant-like biosynthetic pathways in bacteria: from benzoic acid to chalone. J Nat Prod 2002; 65:1956 - 1962
- Xiang L, Moore BS. Biochemical characterization of a prokaryotic phenylalanine ammonia lyase. J Bacteriol 2005; 187:4286 - 4289
- Hill AM, Thompson BL, Harris JP, Segret R. Investigation of the early stages in soraphen A biosynthesis. Chem Commun 2003; 21:1358 - 1359
- Bezanson GS, Desaty D, Emes AV, Vining LC. Biosynthesis on cinnamamide and detection of phenylalanine ammonia-lyase in Streptomyces verticillatus. Can J Microbiol 1970; 16:147 - 151
- Kalaitzis JA, Izumikawa M, Xiang L, Hertweck C, Moore BS. Mutasynthesis of enterocin and wailupemycin analogues. J Am Chem Soc 2003; 125:9290 - 9291
- Jack RW, Tagg JR, Ray B. Bacteriocins of grampositive bacteria. Microbiol Rev 1995; 59:171 - 200
- Brook I. Bacterial interference. Crit Rev Microbiol 1999; 25:155 - 172
- van Belkum MJ, Stiles ME. Nonlantibiotic antibacterial peptides from lactic acid bacteria. Nat Prod Rep 2000; 17:323 - 325
- Moffitt MC, Louie GV, Bowman ME, Pence J, Noel JP, Moore BS. Discovery of two cyanobacterial phenylalanine ammonia lyases: kinetic and structural characterization. Biochemistry 2007; 46:1004 - 1012
- Juvvadi PR, Seshime Y, Kitamoto K. Genomicsreveals traces of fungal phenylpropanoid—flavonoid metabolic pathway in the filamentous fungus Aspergillus oryzae. J Microbiol 2005; 43:475 - 486
- Seshime Y, Rao PR, Fujii I, Kitamoto K. Genomic evidences for the existence of a phenylpropanoid metabolic pathway in Aspergillus oryzae. Biochem Biophy Res Comm 2005; 337:747 - 751
- Poppe L, Retey J. Friedel-Crafts-type mechanism for the enzymatic elimination of ammonia from histidine and phenylalanine. Angew Chem Int Ed Engl 2005; 44:3668 - 3688
- Ferrer JL, Austin MB, Stewart C Jr, Noel JP. Structure and function of enzymes involved in the biosynthesis of phenylpropanoids. Plant Physiol Biochem 2008; 46:356 - 370
- Heredia A, Jimenez A, Guillen R. Composition of plant cell walls. Z Lebensm Unters Forsch 1995; 200:24 - 31
- Boerjan W, Ralph J, Baucher M. Lignin Biosynthesis. Annu Rev Plant Biol 2003; 54:519 - 546
- Chen C, Meyermans H, Burggraeve B, De Rycke RM, Inoue K, et al. Cell-specific and conditional expression of caffeoyl-CoA O-methyltransferase in poplar. Plant Physiol 2000; 123:853 - 867
- Ishii T. Structure and functions of feruloylated polysaccharides. Plant Sci 1997; 127:111 - 127
- Lewis NG, Yamamoto E. Lignin occurance, biogenesis and biodegradation. Ann Rev Plant Physiol Plant Mol Biol 1990; 41:455 - 496
- Bahri H, Dignac M-F, Rumpel C, Rasse DP, Chenu C, Mariotti A. Lignin turnover kinetics in an agricultural soil is monomer specific. Soil Biol Biochem 2006; 38:1977 - 1988
- Higuchi T. Biodegradation mechanism of lignin by white-rot basidiomycetes. J Biotechnol 1993; 30:1 - 8
- Odier E, Janin G, Monties B. Poplar lignin decomposition by gram negative aerobic bacteria. Appl Environ Microbiol 1981; 41:337 - 341
- Higuchi T. Microbial degradation of lignin: role of lignin peroxidase, manganese peroxidase and laccase. Proc Jpn Acad Ser (B) 2004; 80:204 - 214
- El Hanafy AA, Abd-Elsalam HE, Hafez EE. Fingerprinting for the lignin degrading bacteria from soil. J Appl Sci Res 2007; 3:470 - 475
- de Ascensao AR, Dubery IA. Soluble and wall bound phenolic polymers in Musa acuminata roots exposed to elicitors from Fusarium oxysporum f.sp. cubense. Phytochem 2003; 63:679 - 686
- Harborne JB. Phytochemical methods 1998; UK Chapman & Hall
- Seneviratne G, Jayasinghearachchi HS. Phenolic acids: possible agents of modifying N2-fixing symbiosis through rhizobial alteration?. Plant Soil 2003; 252:385 - 395
- Makoi JHJR, Ndakidemi PA. Biological, ecological and agronomic significance of plant phenolic compounds in rhizosphere of the symbiotic legumes. Afr J Biotechnol 2007; 6:1358 - 1368
- Wojtaszek P, Peretiatkowicz M. A simple method for visualization of phenolics exudation by roots of intact lupin plants; the effect of nitrate and pH. Acta Biochimca Polonica 1992; 39:307 - 316
- Eva C, Josep P. Is there a feedback between N availability in siliceous and calcareous soils and Cistus albidus leaf chemical composition. Oecoi 2003; 136:183 - 192
- Blum U. Effects of microbial utilization of phenolic acid breakdown products on allelopathic interactions. J Chem Ecol 1998; 24:685 - 708
- Morel C, Hinsinger P. Root induced modifications of the exchange of phosphate ion between soil solution and solid phase. Plant Soil 1999; 211:103 - 110
- Micales JA. Localization and induction of oxalate decarboxylase in the brown rot wood decay fungus Postia placenta. Int Biodereioration Biodegrad 1997; 39:125 - 132
- Becker BU, Kosch K, Parniske M, Müller P. Exopolysaccharide (EPS) synthesis in Bradyrhizobium japonicum: sequence, operon structure and mutational analysis of an exo gene cluster. Mol Gen Genet 1998; 259:161 - 171
- Schauser L, Roussis A, Stiller J, Stougaard J. A plant regulator controlling development of symbiotic root nodules. Nature 1999; 402:191 - 195
- Broughton WJ, Jabbouri S, Perret X. Keys to symbiotic harmony. J Bacteriol 2000; 182:5641 - 5652
- Jones KM, Kobayashi H, Davies BW, Taga ME, Walker GC. How rhizobial symbionts invade plants: the Sinorhizobium-Medicago model. Nature Rev Microbiol 2007; 5:619 - 633
- Kondorsi A, Schultze M. Regulation of symbiotic root nodule development. Annu Rev Genet 1998; 32:33 - 57
- Geurts R, Fedorova E, Bisseling T. Nod factor signaling genes and their function in the early stages of Rhizobium infection. Curr Opin Plant Biol 2005; 8:346 - 352
- Phillips DA, Torrey JG. Studies on cytokinin production by Rhizobium. Plant Physiol 1972; 49:11 - 15
- Hartwig U, Joseph C, Phillips DA. Flavonoids released naturally from alfalfa seeds enhance growth rate of Rhizobium meliloti. Plant Physiol 1991; 95:797 - 803
- Caetano-Anolles G, Wall LG, De Micheli AT, Macchi EM, Bauer WD, Favelukes G. Role of motility and chemotaxis in efficiency of nodulation by Rhizobium meliloti. Plant Physiol 1988; 86:1228 - 1235
- Fox JE, Starcevic M, Kow KY, Burow ME, McLachlan JA. Endocrine disrupters and flavonoid signaling. Nature 2001; 413:128 - 129
- Cooper JE, Rao R. Localised changes in flavonoid biosynthesis in roots of Lotus pendiculatus after infection by Rhizobium loti. Plant Physiol 1992; 100:444 - 450
- Zuanazzi JAS, Clergeot PH, Quirion J-C, Husson HP, Kondorosi P, Ratet P. Production of Sinorhizobium meliloti nod gene activation and repressor flavonoids from Medicago sativa roots. Mol Plant Microbe Interact 1998; 11:784 - 794
- Rice-Evans C. Flavonoid antioxidants. Curr Med Chem 2001; 8:797 - 807
- D'Arcy-Lameta A, Jay M. Study of soybean and lentil root exudates III. Influence of soybean isoflavonoids on the growth of rhizobia and some rhizospheric microorganisms. Plant Soil 1987; 101:267 - 272
- Zawoznik MS, Garrido LM, del Pero Martinez MA, Tomaro ML. Effect of vanillin on growth and symbiotic ability of Bradyrhizobium sp. (Arachis) strain. Proc Int Plant Growth-promoting Rhizobacteria 2000; http:/www.ag.auburn.edu/argentina
- Novikova TL. Influence of natural phenols on Trifolium pratense-Rhizobium trifoli symbiosis. Acta Hort 1994; 381:421 - 424
- Djordjevic M, Rolfe B. Bothe H, de Bruijn F, Newton F. Recognition process in the Rhizobium trifolii-white clover symbiosis. nitrogen fixation: hundred years after 1988; Stuttgart, Germany Gustav Fischer 431 - 436
- Gerdemann JW. Vesicular-arbuscular mycorrhiza and plant growth. Ann Rev Phytopathol 1968; 6:397 - 418
- Bago B, Pfeffer PE, Abubaker J, Jun J, Allen JW, Brouillette J, et al. Carbon export from arbuscular mycorrhizal roots involves the translocation of carbohydrate as well as lipid. Plant Physiol 2003; 131:1496 - 1507
- Mosse B. Some studies relating to “independent” growth of vesicular-arbuscular endophytes. Can J Bot 1988; 66:2533 - 2540
- Nagahashi G, Douds DD Jr. Partial separation of root exudates components and their effects upon the growth of germinated spores of AM fungi. Mycol Res 2000; 104:1453 - 1464
- Mosse B, Hepper CE. Vesicular-arbuscular mycorrhizal infections in root organ cultures. Physiol Plant Pathol 1975; 5:2015 - 2223
- Giovannetti M, Sbrana C, Citernesi AS, Avio L. Analysis of factors involved in fungal recognition responses to host derived signals by arbuscular mycorrhizal fungi. New Phytol 1996; 133:65 - 71
- Gianinazzi-Pearson V, Branzanti B, Gianinazzi S. In vitro enhancement of spore germination and early hyphal growth of a vesicular-arbuscular mycorrhizal fungus by host root exudates and plant flavonoids. Symbiosis 1989; 7:243 - 255
- Siqueira J, Safir G, Nair M. Stimulation of vesicular-arbuscular mycorrhiza formation and growth of white clover by flavonoid compounds. New Phytol 1991; 118:87 - 93
- Tsai SM, Phillips DA. Flavonoids released naturally from alfalfa promote development of symbiotic Glomus spores in vitro. Appl Environ Microbiol 1991; 57:85 - 88
- Poulin MJ, Simard J, Catford JG, Labrie F, Piche Y. Response of symbiotic endomycorrhizal fungi to estrogen and antiestrogens. Mol Plant Microbe Interact 1997; 10:481 - 487
- Harrison MJ, Dixon RA. Isoflavonoid accumulation and expression of defense gene transcripts during the establishment of vesicular arbuscular mycorrhizal associations in roots of Medicago truncatula. Mol Plant-Microbe Interact 1993; 6:643 - 654
- Harrison MJ, Dixon RA. Spatial patterns of expression of flavonoid/isoflavonoid pathway genes during interactions between roots of Medicago truncatula and the mycorrhizal fungus Glomus versiforme. Plant J 1994; 6:9 - 20
- Scervino JM, Ponce MA, Erra-Bassells R, Vierheilig H, Ocampo JA, Godeas A. Flavonoids exhibit fungal species and genus specific effects on the presymbiotic growth of Gigaspora and Glomus. Mycol Res 2005; 109:789 - 794
- Scervino JM, Ponce MA, Erra-Bassells R, Vierheilig H, Ocampo JA, Godeas A. Arbuscular mycorrhizal colonization of tomato by Gigaspora and Glomus species in the presence of root flavonoids. J Plant Physiol 2005; 162:625 - 633
- Silva-Junior P, Siqueira JO. Colonização micorrízica e crescimento da soja com diferentes fungos e aplicação do isoflavonóide formononetina. Pesquisa Agropecuária Brasileira 1998; 33:953 - 959
- Ponce M, Scervino J, Erra-Balsells R, Ocampo J, Godeas A. Flavonoids from shoots and roots of Trifolium repens (white clover) grown in presence or absence of the arbuscular mycorrhizal fungus Glomus intraradices. Phytochemistry 2004; 65:1925 - 1930
- Selvaraj JT, Subramanian G. Phenols and lipids in mycorrhizal and non-mycorrhizal roots of Sesamum indicum. Cur Sci 1990; 59:471 - 473
- Fries LLM, Pacovsky RS, Safir GR, Siqueira JO. Plant growth and arbuscular mycorrhizal fungal colonization affected by exogenously applied phenolic compounds. J Chem Ecol 1997; 23:755 - 1767
- Matusova R, Rani K, Verstappen FWA, Franssen MCR, Beale MH, Bouwmeester HJ. The strigolactone germination stimulants of the plant-parasitic Striga and Orobanche spp. are derived from the carotenoid pathway. Plant Physiol 2005; 139:920 - 934
- Steinkellner S, Lendzemo V, Langer I, Schweiger P, Khaosaad T, Toussaint JP, Vierhailig H. Flavonoids and strigolactones in root exudates as signals in symbiotic and pathogenic plant-fungus interactions. Molecules 2007; 12:1290 - 1306
- Gomez-Roldan V, Roux C, Girard D, Bécard G, Puech V. Strigolactones: Promising plant signals. Plant Signal Behav 2007; 2:163 - 164
- Akiyama K, Hayashi H. Strigolactones: chemical signals for fungal symbionts and parasitic weeds in plant roots. Ann Bot 2006; 97:925 - 931
- Bouwmeester HJ, Matusova R, Zhongkui S, Beale MH. Secondary metabolites signaling in host-parasitic plant interactions. Curr Opin Plant Biol 2003; 6:358 - 364
- Horvath B, Bachem CWB, Schell J, Kondorsi A. Host specific regulation of nodulation genes in Rhizobium is mediated by a plant signal, interacting with the nodD gene product. The EMBO J 1987; 6:841 - 848
- Redmond JW, Batley M, Djordjevic MA, Innes RW, Kuempel PL, Rolfe BG. Flavones induce expression of nodulation genes in Rhizobium. Nature 1986; 323:632 - 635
- van Rhijn P, Vanderleyden J. The Rhizobium-plant symbiosis. Microbiol Rev 1995; 59:124 - 142
- Spaink HP. Root nodulation and infection factors produced by rhizobial bacteria. Annu Rev Microbiol 2000; 54:257 - 288
- Ferguson BJ, Mathesius U. Signaling interactions during nodule development. Plant Growth Regul 2003; 22:47 - 72
- Prinsen E, Chauvaux N, Schmidt J, John M, Wieneke U, Greef JD, et al. Stimulation of indole-3-acetic acid production in Rhizobium by flavonoids. FEBS Lett 1991; 282:53 - 55
- Theunis M, Kobayashi H, Broughton WJ, Prinsen E. Flavonoids, NodD1, NodD2 and nod-box NB15 modulate expression of the y4wEFG locus that is required for indole-3-acetic acid synthesis in Rhizobium sp. strain NGR234. Mol Plant-Microbe Interact 2004; 10:1153 - 1161
- Hirsch A. Developmental biology of legume nodulation. New Phytol 1992; 122:211 - 237
- deBilly F, Grosjean C, May S, Bennett M, Cullimore JV. Expression studies on AUX1-like genes in Medicago truncatula suggest that auxin is required at two steps in early nodule development. Mol Plant Microbe Interact 2001; 14:267 - 277
- Lippincott JA, Lippincott BB, Starr MP. Starr MP, Stoip H, Trupper HG, Balows A, Schiegel HG. The Prokaryotes 1981; 1:New York Springer 842 - 855
- Heath JD, Charles TC, Nester EW. Hoch JA, Silhavy TJ. Two-component signal transduction 1995; Washington, DC Am Soc Microbiol 367 - 385
- Lee K, Dudley MW, Hess KM, Lynn DG, Joerger RD, Binns AN. Mechanism of activation of Agrobacterium virulence genes: identification of phenol-binding proteins. Proc Natl Acad Sci USA 1992; 89:8666 - 8670
- Lee YW, Jin S, Sims WS, Nester EW. Genetic evidence for direct sensing of phenolic compounds by the VirA protein of Agrobacterium tumefaciens. Proc Natl Acad Sci USA 1995; 92:12245 - 12249
- Harrier LA, Wright F, Hooker JE. Isolation of the 3-phosphoglycerate kinase gene of the arbuscular mycorrhizal fungus Glomus mosseae (Nicol. & Gerd.) Gerdemann & Trappe. Curr Genet 1998; 34:386 - 392
- Martin-Laurent F, van Tuinen D, Dumas-Gaudot E, Gianinazzi-Pearson V, Gianinazzi S, Franken P. Differential display analysis of RNA accumulation in arbuscular mycorrhiza of pea and isolation of a novel symbiosis-regulated plant gene. Mol Genet Genomics 1997; 256:37 - 44
- Requena N, Mann P, Hampp R, Franken P. Early developmentally regulated genes in the arbuscular mycorrhizal fungus Glomus mosseae: identification of GmGin1 a novel gene with homology to the C-terminus of metazoan hedgehog proteins. Plant Soil 2002; 244:129 - 139
- Requena N, Serrano E, Ocon A, Breuninger M. Plant signals and fungal perception during arbuscular mycorrhiza establishment. Phytochemistry 2007; 68:33 - 40
- Chester KS. The problem of acquired physiological immunity in plants. Quart Rev Biol 1993; 8:275 - 324
- Aist JR, Gold RE. Prevention of fungal ingress: The role of papillae and calcium 1987; Tokyo/Springer-Verlag, Berlin Japan Sci Soc Press 47 - 58
- Redman RS, Freeman S, Clifton DR, Morrel J, Brown G, Rodriguez RJ. Biochemical analysis of plant protection afforded by a nonpathogenic endophytic mutant of colletotrichum magna. Plant Physiol 2003; 119:795 - 804
- Metraux JP. Systemic acquired resistance and salicylic acid: current state of knowledge. Eur J Plant Pathol 2001; 107:13 - 18
- Ndakidemi PA, Dakora FD. Legume seed flavonoids and nitrogenous metabolites as signals and protectants in early seedling development. Rev Funct Plant Biol 2003; 30:729 - 745
- Inderjit, Duke SO. Ecophysiological aspects of allelopathy. Planta 2003; 217:529 - 539
- Bais HP, Weir TL, Perry LG, Gilroy S, Vivanco JM. The role of root exudates in rhizosphere interactions with plants and other organisms. Annu Rev Plant Biol 2006; 57:233 - 266
- Dakora FD. Plant flavonoids: biological molecules for useful exploitation. Aust J Plant Physiol 1995; 22:87 - 99
- Dakora FD, Phillips DA. Diverse functions of isoflavonoids in legumes transcend ant-microbial definitions of phytoalexins. Physiol Mol Plant Pathol 1996; 49:1 - 20
- Nicholson RL, Hammerschmidt R. Phenolic compounds and their role in disease resistance. Annu Rev Phytopathol 1992; 30:369 - 389
- Sarma BK, Singh UP. Ferulic acid prevents infection by Sclerotium rolfsii in Cicer arietinum. World J Microbiol Biotechnol 2003; 19:123 - 127
- Field B, Jordon F, Osbourn. First encounters-deployment of defence related natural products by plants. New Phytol 2006; 172:193 - 207
- Shaw L, Phil Morris J, Hooker JE. Perception and modification of plant flavonoid signals by rhizosphere microorganisms. Environ Microbiol 2006; 8:1867 - 1880
- Walker T, Bias H, Grotewold E, Vivanco J. Root exudation and rhizosphere biology. Plant Physiol 2003; 132:44 - 51
- Volpin H, Elland Y, Okon Y, Kapulnik Y. A vesicular arbuscular mycorrhizal fungus (Glomus intraradix) induces a defence response in alfalfa roots. Plant Physiol 1994; 104:683 - 689
- Rao JR, Cooper JE. Soybean nodulating rhizobia modify nod gene inducers daidzein and genistein to yield aromatic products that can influence geneinducing activity. Mol Plant Microbe Interact 1995; 8:855 - 862
- Pieterse CMJ, Van Pelt JA, Van Wees SCM, Ton J, L'eon-Kloosterziel KM, Keurentjes JJB, et al. Rhizobacteria-mediated induced systemic resistance: triggering, signalling and expression. Eur J Plant Pathol 2001; 107:51 - 61
- Castro-Sowinski S, Herschkovitz Y, Okon Y, Jurkevitch E. Effects of inoculation with plant growth-promoting rhizobacteria on resident rhizosphere microorganisms. FEMS Microbiol Lett 2007; 276:1 - 11
- Samac DA, Graham MA. Recent advances in legume-microbe interactions: recognition, defense response and symbiosis from a genomic perspective. Plant Physiol 2007; 144:582 - 587
- Vaughan D, Ord B. Influence of phenolic acids on morphological changes in roots of Pisum sativum. J Sci Food Agric 1990; 52:289 - 299
- Mishra RPN, Singh RK, Jaiswal HK, Kumar V, Maurya S. Rhizobium-Mediated Induction of Phenolics and Plant Growth Promotion in Rice (Oryza sativa L.). Curr Microbiol 2006; 52:383 - 389
- Aguilar JMM, Ashby AM, Richards AJM, Loake GJ, Watson MD, Shaw CH. Chemotaxis of Rhizobium leguminosarum biovar phaseoli towards flavonoid inducers of the symbiotic nodulation genes. J Gen Microbiol 1988; 134:2741 - 2746
- Moran JF, Klucas RV, Grayer RJ, Abian J, Becana M. Complexes of Iron with Phenolic Compounds from Soybean Nodules and Other Legume Tissues: Prooxidant and Antioxidant Properties. Free Radic Biol Med 1997; 22:861 - 870
- Sobolev VS, Horn BW, Potter TL, Deyrup ST, Gloer JB. Production of Stilbenoids and Phenolic Acids by the Peanut Plant at Early Stages of Growth. J Agric Food Chem 2006; 54:3505 - 3511
- van Spronsen PC, Tak T, Rood AMM, van Brussel AAN, Kijne JW, Boot KJM. Salicylic acid inhibits indeterminate-type nodulation but not determinate-type nodulation. Mol Plant-Microb Interact 2003; 16:83 - 91
- Stacey G, Mc Alvin CB, Kim SY, Olivares J, Soto MJ. Effects of Endogenous Salicylic Acid on Nodulation in the Model Legumes Lotus japonicus and Medicago truncatula. Plant Physiol 2006; 141:1473 - 1481
- Sato T, Fujikake H, Ohtake N, Sueyoshi K, Takahashi T, Sato A, Ohyama T. Effect of exogenous salicylic acid supply on nodulation formation of hypernodulating mutant and wild type of soybean. Soil Sci Plant Nutr 2002; 48:413 - 420