Abstract
Homeostasis in the central nervous system (CNS) is maintained by active interfaces between the bloodstream and the brain parenchyma. The blood-brain barrier (BBB) constitutes a selective filter for exchange of water, solutes, nutrients, and controls toxic compounds or pathogens entry. Some parasites, bacteria, and viruses have however developed various CNS invasion strategies, and can bypass the brain barriers. Concerning viruses, these strategies include transport along neural pathways, transcytosis, infection of the brain endothelial cells, breaching of the BBB, and passage of infected-leukocytes. Moreover, neurotropic viruses can alter BBB functions, thus compromising CNS homeostasis. Retroviruses have been associated to human neurological diseases: HIV (human immunodeficiency virus 1) can induce HIV-associated dementia, and HTLV-1 (human T lymphotropic virus 1) is the etiological factor of tropical spastic paraparesis/HTLV-1 associated myelopathy (TSP/HAM). The present review focuses on how the different retroviruses interact with this structure, bypass it and alter its functions.
Introduction
The Central Nervous System (CNS), which is composed of the brain and the spinal cord, is mainly isolated from the rest of the organism by blood/brain interfaces, including the Blood-Brain Barrier (BBB) and the Blood-Cerebrospinal Fluid (CSF) barrier. These barriers maintain the CNS homeostasis, by regulating ions, water and solutes exchanges, and protect it from toxic compounds or pathogens. However, several pathogens have developed invasion strategies of the CNS, bypassing these barriers.
Neurotropic viruses, i.e., viruses that are able to infect neural cells can be retrieved in several viral families, such as Herpesviridae, Paramyxoviridae, Rhabdoviridae, Picornaviridae, Retroviridae,Citation1 etc. They can gain access to the CNS by different routes: either they use the neural network and the axonal transport (rabies virus or herpes simplex virus type I, for exampleCitation2), or they invade the CNS from the bloodstream (West Nile virus, for exampleCitation3). The hematogenous route is the one usually used by the retroviruses.
Retroviruses are enveloped RNA viruses that have the unique property of transcribing RNA into DNA, and integrating their retroviral DNA into the chromosomal DNA of the host cell. Two human retroviruses have been recognized to be responsible for major CNS diseases: human immunodeficiency virus type 1 (HIV-1), which can be responsible of damages to the brain and the spinal cord during acquired immune deficiency syndrome (AIDS) and human T-cell lymphotropic virus type 1 (HTLV-1), which is responsible for a progressive neurodegenerative disease, the tropical spastic paraparesis/HTLV-1-associated myelopathy (TSP/HAM) in some HTLV-1 infected persons. Other retroviruses, from the Lentiviral subgroup, are also able to induce neurological syndromes in animals. The present review summarizes the main characteristics of the BBB, and the different interactions between retroviruses and this structure.
Overview of the Blood-Brain Barrier
The brain is a highly perfused organ: it represents only 2% of body weight, but receives 15–20% of total cardiac output. The blood/CNS interfaces are diverse: they include the BBB, as well as the blood cerebrospinal fluid barrier, the blood retinal barrier, the blood nerve barrier and the blood labyrinth barrier.Citation4 The barrier properties are usually determined by the endothelium of brain microvessels, but both in choroid plexus and arachnoid the barrier is determined by an epithelium. The cerebral endothelium is the primary site of oxygen and nutrient exchanges.Citation5 In the brain, it is estimated that nearly every neuron, the core components of the nervous system, has its own microvessel,Citation6 underlining the critical relationship between the neuronal and vascular compartments. The combined surface area of these microvessels, depending on the anatomical region, ranges between 150 and 200 cm2 g−1 tissue, with a total area for exchange in the brain about 12 m2 for the average human adult.Citation7,Citation8
The neurological activity requires a strict cerebral homeostasis. With the exception of the circumventricular organs, which are regions of important exchanges between specialized neurons and the blood stream, the homeostasis of the CNS is achieved by exquisite regulation of nutrients, solutes and water exchanges at blood/CNS interfaces.Citation9
As BBB specificities and characteristics are mainly induced, maintained and/or interfered by cerebral vicinity, it is worth to consider the whole neurovascular unit rather than the brain endothelium as an isolated structure.
The neurovascular unit
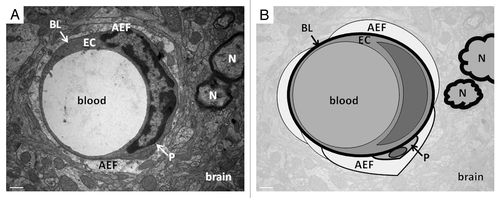
The cerebral endothelium is a very dense network of intercommunicating capillaries and microvessels, composed of specialized endothelial cells (). BBB endothelium is characterized by the presence of tight junctions (TJs). In addition, cerebral microvascular endothelial cells are surrounded by pericytes, which are important for the maintenance of vascular homeostasis and are source of adult pluripotent stem cells.Citation10 Finally, basal lamina surrounds the microvessels and astrocytic end-feet sheath the vascular structure. The importance of astrocyte was for long undermined: their role was purportedly limited to providing trophic, metabolic, and structural support for neural networks. Since, it has been demonstrated that astrocytes also play a signaling role: they communicate with neurons via Ca2+ signaling, and can release signaling transmitters, termed gliotransmitters.Citation11
Figure 1. Transmission electron microscopy (A) and schematic view (B) of rat brain illustrating the neurovascular unit. This complex includes microvessel endothelial cells (EC), based on basal lamina (BL), astrocytes end-feet (AEF) and some neurons (N) in the vicinity. Scale bar: 0.5 μm.
Functions and characteristics of the BBB
The neurovascular unit acts, besides other roles, as a physical barrier for the CNS. Cerebral endothelium is continuous without any fenestration and intercellular junction complexes are formed. Junction complexes comprise adherens and tight junctions; these are accumulated close to the apical side of endothelial cells.Citation12 Adherens junctions are composed of cadherin-catenin complexes, and are important in the initiation, maturation and maintenance of endothelial intercellular contacts. TJs are mainly composed of three transmembrane proteins (claudins, occludin, and Junction Adhesion Molecules) associated with cytoplasmic accessory proteins (Zonula Occludens-1, -2, -3, cingulin). These latter link membrane proteins to the actin cytskeleton, which is involved in the structural and functional integrity of cerebral endothelium.Citation12,Citation13
This typical angioarchitecture, with junction complexes that fasten together adjacent endothelial cells,Citation14 together with minimal vesicular transport activity in the endothelial cells,Citation9,Citation14 explain mostly the restrictiveness of the BBB. As passive diffusion between the bloodstream and the brain is abolished, BBB strictly controls nutrient transport to the brain as well as efflux of metabolites.Citation15 Moreover, BBB confers a large protection against toxicity of many xenobiotics, as it regulates efflux of drugs from the CNS to the blood.Citation16 Finally it is a good protection against pathogens: it restricts the entry of circulating immune cellsCitation17 and pathogens.Citation18 Retroviruses, as other viruses, have developed different ways to overcome this protective barrier, as described as follows.
Blood-Brain Barrier, HIV and Other Lentiviral Infections
HIV-infected patients commonly develop neurological symptoms, such as HIV-associated dementia (HAD), and its pathological correlate, HIV-encephalitis (HIVE). These can occur even in the absence of opportunistic infections and are characterized by motor and cognitive disorders, such as limb muscle weakness, loss of memory, depression and dementia.Citation19,Citation20 The clinical manifestations are accompanied by histological hallmarks, such as neocortical and subcortical damage within the white and gray matter,Citation21-Citation23 presence of multinucleated giant cells, neuronal loss and astrocytosis. Although the introduction of highly active antiretroviral therapy (HAART) has been able to reverse some of the clinical manifestations, pathological alterations persist within the CNS of infected patients as the drugs hardly penetrate the CNS turning difficult the control of HIV replication within the brain.Citation24
Blood-brain barrier alterations during HIV infection
Blood-brain barrier functional perturbations have been early identified during HIV infection, as shown by the presence of cellular infiltrates and diffusion of seric markers within the brain parenchyma,Citation24-Citation27 as well as by dynamic magnetic resonance imaging.Citation28 Additional evidence of BBB alteration in the CNS of infected patients has been brought by observation of TJ disorganization, especially in the expression pattern of occludin and the protein Zonula Occludens 1 (ZO-1).Citation29 Interestingly, whereas brain sections from HIV infected patients without HIVE showed normal amounts of occludin- or ZO-1-reactive blood vessels, brain sections from patients with HIVE showed marked alterations in both the intensity and staining pattern of occludin and ZO-1.Citation29 Moreover, a correlation could be established between the level of ZO-1 disorganization in brain endothelia and the extent of dementia in HAD patients.Citation30
Three main mechanisms were proposed to explain BBB alteration during HIV infection: (1) BBB is altered in response to proinflammatory cytokines secretion by HIV-infected cells or activated endothelial cells, (2) BBB is altered in response to secreted viral proteins (Tat or gp120) and (3) BBB is altered upon infection of endothelial cells by HIV.
Effect of proinflammatory cytokines secretion
During the asymptomatic phase, HIV-infected patients exhibit a chronic activation of immune system, accompanied by dysregulation of cytokine secretion.Citation31,Citation32 At this stage, HIV can be detected in the CSF,Citation33 as the consequence of trafficking of either activated and infected CD4+ T cells, monocytes or dendritic cells from the periphery. After the virus has reached the CNS, productive virus replication occurs in the CNS, which is accompanied by massive cytokine and chemokine secretion in the CNS.Citation34,Citation35 Cerebral endothelial cell functions are deeply impaired during this chronic activation; they overexpress cell adhesion markers such as intercellular adhesion molecule-1 (ICAM-1) and secrete metalloproteases, which induce basal lamina thinning.Citation36-Citation38 This facilitates mononucleated cells extravasation into the CNS.
Effect of secreted viral proteins
Since clinical manifestations of HIV infection often do not correlate directly with viral titers, it has been thought that the effects of infection could be mediated by viral soluble factors secreted from HIV-infected cells.
Among them, the viral Tat protein, which is secreted by infected cellsCitation39 and is able to cross cell membranes, can be detected in both sera and CSFs from HIV-infected patients.Citation40 Evidence of a deleterious effect of Tat on BBB has been brought both on cellular models in vitro and in vivo.Citation41-Citation45 Tax-induced BBB alteration occurs through the disorganization of TJs in vitro: decrease in claudin-1, claudin-5 and ZO-2 expression were observed in cellular models treated with recombinant Tat.Citation44 Moreover, in vivo, exposure to recombinant Tat were sufficient to induce redistribution of claudin-5 immunoreactivity in a murine model.Citation44 Similarly, Tat administration into the brain hippocampi of C57BL/6 mice resulted in decreased mRNA levels of ZO-1 and drastic reduction of ZO-1 continuity in brain microvessels.Citation43 Such changes are mediated by the activation of extracellular signal-regulated kinase 1/2 (ERK1/2), suggesting that Tat-induced oxidative stress may play an important role in affecting BBB integrity via the ERK1/2 pathway. In fact, a dose-dependence relationship was established between the oxidative stress degree and Tat concentration in brain endothelial cell cultures.Citation45 Via the increase in oxidative stress, HIV infection could increase cytoplasmic calcium concentration, thus altering mitochondrial functions and inducing endothelial cell apoptosis. Endothelial cell apoptosis was indeed observed in the brains of some AIDS patientsCitation46; however the importance of such a mechanism has not yet been established in vivo.Citation47
The HIV envelope glycoprotein gp120 could also alter BBB integrity. Alterations in the BBB have been detected in transgenic mice expressing solely the viral gp120. Gp120 expression induces matrix metalloproteinase-2 secretion in vivo that might alter the basal lamina,Citation37 oxydative stressCitation48 and ZO-1 and occludin degradation accompanied with claudins expression alterations.Citation49 TJ proteins degradation might be mediated by proteasome in cultured human brain microvascular endothelial cells.Citation50 Eventually, BBB permeability is impaired and infected monocytes migration increased.Citation51
Direct infection of endothelial cells by HIV
Brain endothelial cells can potentially be infected. Microvascular cerebral endothelial cells express HIV receptor and co-receptors.Citation52,Citation53 Moreover, primary endothelial cells can be productively infected by HIV in vitro,Citation54,Citation55 although such an infection could only been demonstrated for dual-tropic X4R5 HIV strains. Infection of brain endothelial cells could have many deleterious effects on BBB integrity: (1) HIV-infected endothelial cells could secrete cytokines and the viral protein Tat, with deleterious effect on BBB; (2) infection of brain endothelial cells enter apoptosis; (3) infected endothelial cells could secrete metalloproteases able to alter BBB.Citation56
Infection of brain endothelial cells in vivo is still a matter of debate. Some reports suggested the presence of infected endothelial cells through in situ hybridization experiments.Citation57-Citation59 However, these studies could not exclude that such positive signal could correspond to infected perivascular macrophages, in fact they concluded solely based on morphological criteria, in the absence of endothelial cell typing by histochemistry. In addition, using PCR/in situ hybridization technique, no HIV-infected endothelial cells could be detected in the brains of adult patients with AIDS, including patients with HAD.Citation60
Cell trafficking through BBB during HIV infection
In addition to altering BBB functions either by viral-induced TJ disorganization or endothelial cell cytopathic effect, HIV can access the CNS by an increased trafficking of HIV-infected CD4+ T cells or circulating monocytes; this is named the “Trojan horse” mechanism.Citation61
Whereas in healthy individuals leukocyte trafficking toward the CNS is very low, in inflammatory conditions lymphocytes and monocytes/macrophages gain access to the CNS by increased migration through the BBB.Citation17,Citation62 In the case of HIV infection, inflammation in the CNS, and subsequent increased transmigration of lymphocytes and monocytes through the BBB, have been shown (for a recent review, see ref. Citation63). For example, Tat induces adhesion molecules in endothelial cells and chemokines secretion by astrocytes and microglial cells, thus possibly enhancing leukocyte trafficking toward CNS.Citation64 In addition, the viral protein gp120, which can be detected in the brain of HIV-1 infected patients,Citation65 triggers the release of MCP-1, as a potent chemoattractant for monocytes.Citation66 Interestingly, an increased risk of HAD has been shown to be linked to a mutant MCP-1 allele.Citation67 Finally, proinflammatory cytokines levels are elevated in CSF and brain parechyma of HAD patients. These can alter BBB integrity, increase the expression of the adhesion molecules ICAM-1, Vascular Cell Adhesion Molecule-1 (VCAM-1) and E-selectins on endothelial cells, thus facilitating leukocyte adhesion, rolling and estravasation into the brain.Citation68
Interestingly, some hypothesized that the increased trafficking of leukocytes toward the CNS during HAD would be a consequence of early alteration of the gastrointestinal mucosa observed during HIV infection.Citation69 In fact, gastrointestinal rupture could lead to translocation of bacterial endotoxin LPS, which could induce a generalized systemic activation, LPS-induced BBB damageCitation70 and an increase in monocyte transmigration.Citation71,Citation72
Blood-brain barrier and other lentiviruses
Most of the observations reported for HIV seem to stand true for other lentivirus-associated CNS disorders.
As for its human counterpart, the simian immunodeficiency virus can induce encephalitis in macaques.Citation73-Citation76 Brains of SIV-infected macaques exhibit fragmented and reduced immunoreactivity for occludin and ZO-1, in association with accumulation of perivascular macrophages.Citation77
Upon Feline Immunodeficiency Virus (FIV) infection, BBB and choroid plexus functions are impaired.Citation78 In vitro, cat brain microvascular endothelial cells can be infected with FIV, which could lead to important alterations of BBB functions.Citation79 However, as for HIV-1, FIV-infection of brain endothelial cells in vivo remains controversial; the current model favors a CNS invasion by FIV through trafficking of infected lymphocytes.Citation80
Interestingly, both Visna virus and Caprine Arthritis Encephalitis virus can cross the BBB, but since no free virus could be detected in the blood, it has been suggested once again that the major entry mechanism for lentiviruses is the “Trojan horse” mechanism.Citation81,Citation82
Blood-Brain Barrier and HTLV-1
Human T-cell leukemia virus type 1 (HTLV-1), a retrovirus that infects 15 to 20 millions people worldwide, is known to cause a variety of diseases, including a chronic neurological syndrome called either tropical spastic paraparesis or HTLV-1-associated myelopathy (TSP/HAM).Citation83-Citation85 TSP/HAM is a slowly progressive neurological disease, which occurs in less than 3% of HTLV-1 infected people, and is characterized by BBB alterations such as immunoglobulin and fibrinogen deposit in the brain parenchyma,Citation86,Citation87 and crossing of HTLV-1 infected lymphocytes through the BBB.Citation88,Citation89
Three putative mechanisms have been proposed for TSP/HAM pathogenesis. First, the “bystander” model is based on the observation that CSF from HAM/TSP patients is enriched in pro-inflammatory cytokines such as tumor necrosis factor-α (ΤΝF-α), interleukin-1 (IL-1), IL-6 and interferon-γ (IFN-γ).Citation90-Citation92 These pro-inflammatory cytokines, as well as HTLV-1 proteins (such as the viral transactivator Tax protein) are secreted by infiltrating infected lymphocytes that have crossed the BBB and could induce glial cell stress and effect on myelin.Citation93 In the second model, called the “cytotoxic model,” glial cells get infected by HTLV-1.Citation94 Infected glial cells would then be recognized as targets by anti-HTLV-1 cytotoxic lymphocytes (CTL) and subsequently lysed. The third model puts at play an auto-immune mechanism. The viral Tax protein dominant epitope (346–353) cross-reacts with a neuron-specific ribonucleoprotein, hnRNP A1.Citation95 Therefore, the humoral immunity against HTLV-1 can give rise to antibodies directed against neuronal antigens,Citation96 and such antibodies have been detected in sera from TSP/HAM patients.
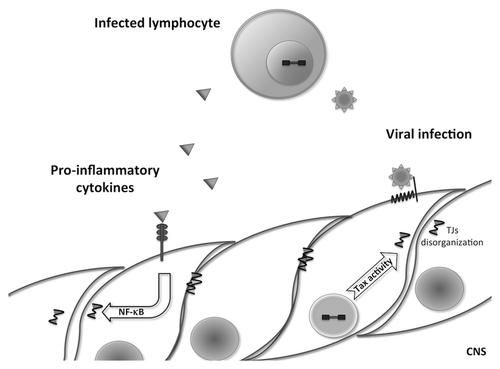
All hypotheses include an initial step of BBB breakdown and/or BBB crossing by HTLV-1 infected lymphocytes. In this context, in vitro and ex vivo studies have been performed to decipher the cellular and molecular mechanisms of BBB alteration and HTLV-1 infected lymphocytes crossing through this latter. In an in vitro rat endothelial model, HTLV-1 infected lymphocytes could alter the endothelial monolayer through TNF-α secretion, and HTLV-1 virions could be transferred through endothelial cells, either by transcytosis or transient infection.Citation97 More recently, in an in vitro model of human brain endothelial cells, HTLV-1 infected lymphocytes were able to alter TJ structures, increase paracellular permeability and transcellular migration, via secretion of both IL-1α and TNF-α.Citation98 These two cytokines have also been shown in an in vitro model of epithelial choroid cells to be able to mediate the alteration of epithelial transport processes induced by HTLV-1 infected lymphocytes.Citation99 In addition, cerebral endothelial cells can be infected by HTLV-1, as shown in vitro, in human brain endothelial cells, and in necropsy spinal cord sections from TSP/HAM patients.Citation100 In vitro, such an infection is productive and alters BBB functions, thus providing additional mechanisms for BBB alteration and viral access to the CNS during TSP/HAM ().
Figure 2. Possible mechanisms of blood-brain barrier disruption by HTLV-1-infected lymphocytes. During TSP/HAM, HTLV-1 infected lymphocytes may disrupt the blood-brain barrier either by proinflammatory cytokine secretion (TNF-α, IL-1α) that activate NFκB pathway in endothelial cells, which induce tight junction disruption, or by infecting endothelial cells; the viral protein Tax could then induce tight junction disorganization.
Conclusion and Perspectives
In conclusion, although the pathological consequences are rather different, the two human retroviruses linked to neurological symptoms, i.e., HIV and HTLV-1, bypass and alter the BBB using very similar mechanisms. Besides the “Trojan horse” strategy to invade the CNS via infected infiltrating cells, the role of the infection of endothelial cells remains to be further investigated. BBB alterations that occur during retroviral infection are often related with a combination of viral-induced proinflammatory cytokines secretion and direct effect of viral proteins (Tat or Tax).
However, the interactions between retroviruses and BBB might have to be revisited in the context of both anti-retroviral therapy and social behaviors, such as drug abuse. Concerning HTLV-1, the use of valproate, a drug often use for epilepsy treatment, has been shown to increase the proviral load and alter motor functions at the beginning of the treatment.Citation101 Concerning HIV, the introduction of highly anti-retroviral therapy (HAART) has drastically limited BBB alterations, which were previously frequently detected.Citation102 Indeed, HAART has been shown to limit or prevent lymphocytic infiltration toward the CNS. However, during the occasional immune reconstitution inflammatory syndrome (IRIS) associated with HAART, a massive lymphocyte extravasation through brain parenchyma has been reported.Citation103 The mechanisms of BBB alteration in this context might be different. In the context of drug abuse, the viral capacity to penetrate the CNS might be modified. As an example, methamphetamines alter BBB permeability through modulation of TJ expression,Citation104 facilitating the entry of the virus or infected cells. In the same manner, use of cocaine can increase HIV-1 neuroinvasion by upregulating the expression of adhesion molecules and matrix metalloproteinases in cultured brain microvascular cells.Citation105,Citation106 The viral protein Tat effects on BBB integrity are also directly exacerbated by cocaine, with a differential effect between the Tat proteins from HIV-1 clades B and C.Citation107 By contrast, cannabinoids can inhibit HIV-1 gp120-induced alterations in cultured microvascular endothelial cells.Citation108 Finally, since combination of HAART (and especially the HIV protease inhibitor saquinavir) with chronic exposure to nicotine has been recently shown to induce BBB integrity alterationCitation109,Citation110it makes clear that retrovirus interaction with the BBB remains a topic of major interest in the next years.
Acknowledgments
The authors are grateful to P.O. Couraud, N. Weiss, B. Saubamea, S. Youssif and A. Schmidt, J.M. Massé (Institut Cochin; Paris) for their contribution to Figure 1. They acknowledge C. Cecilio (Médiathèque Pasteur Institute) for precious help in preparing the manuscript.
References
- Romero JR, Newland JG. Viral meningitis and encephalitis: traditional and emerging viral agents. Semin Pediatr Infect Dis 2003; 14:72 - 82; http://dx.doi.org/10.1053/spid.2003.127223; PMID: 12881794
- Ceccaldi PE, Gillet JP, Tsiang H. Inhibition of the transport of rabies virus in the central nervous system. J Neuropathol Exp Neurol 1989; 48:620 - 30; http://dx.doi.org/10.1097/00005072-198911000-00004; PMID: 2477505
- Ceccaldi PE, Lucas M, Despres P. New insights on the neuropathology of West Nile virus. FEMS Microbiol Lett 2004; 233:1 - 6; http://dx.doi.org/10.1016/j.femsle.2004.01.035; PMID: 15098543
- Neuwelt EA, Bauer B, Fahlke C, Fricker G, Iadecola C, Janigro D, et al. Engaging neuroscience to advance translational research in brain barrier biology. Nat Rev Neurosci 2011; 12:169 - 82; http://dx.doi.org/10.1038/nrn2995; PMID: 21331083
- Cipolla MJ. The Cerebral Circulation. San Rafael (CA), 2009.
- Zlokovic BV. Neurovascular mechanisms of Alzheimer’s neurodegeneration. Trends Neurosci 2005; 28:202 - 8; http://dx.doi.org/10.1016/j.tins.2005.02.001; PMID: 15808355
- Pan W, Kastin AJ. Why study transport of peptides and proteins at the neurovascular interface. Brain Res Brain Res Rev 2004; 46:32 - 43; http://dx.doi.org/10.1016/j.brainresrev.2004.04.006; PMID: 15297153
- Begley DJ, Brightman MW. Structural and functional aspects of the blood-brain barrier. Prog Drug Res 2003; 61:39 - 78; PMID: 14674608
- Reese TS, Karnovsky MJ. Fine structural localization of a blood-brain barrier to exogenous peroxidase. J Cell Biol 1967; 34:207 - 17; http://dx.doi.org/10.1083/jcb.34.1.207; PMID: 6033532
- Bonkowski D, Katyshev V, Balabanov RD, Borisov A, Dore-Duffy P. The CNS microvascular pericyte: pericyte-astrocyte crosstalk in the regulation of tissue survival. Fluids Barriers CNS 2011; 8:8; http://dx.doi.org/10.1186/2045-8118-8-8; PMID: 21349156
- Nag S. Morphology and properties of astrocytes. Methods Mol Biol 2011; 686:69 - 100; http://dx.doi.org/10.1007/978-1-60761-938-3_3; PMID: 21082367
- Weiss N, Miller F, Cazaubon S, Couraud PO. The blood-brain barrier in brain homeostasis and neurological diseases. Biochim Biophys Acta 2009; 1788:842 - 57; http://dx.doi.org/10.1016/j.bbamem.2008.10.022; PMID: 19061857
- Correale J, Villa A. Cellular elements of the blood-brain barrier. Neurochem Res 2009; 34:2067 - 77; http://dx.doi.org/10.1007/s11064-009-0081-y; PMID: 19856206
- Brightman MW, Reese TS. Junctions between intimately apposed cell membranes in the vertebrate brain. J Cell Biol 1969; 40:648 - 77; http://dx.doi.org/10.1083/jcb.40.3.648; PMID: 5765759
- Bickel U, Yoshikawa T, Pardridge WM. Delivery of peptides and proteins through the blood-brain barrier. Adv Drug Deliv Rev 2001; 46:247 - 79; http://dx.doi.org/10.1016/S0169-409X(00)00139-3; PMID: 11259843
- Löscher W, Potschka H. Drug resistance in brain diseases and the role of drug efflux transporters. Nat Rev Neurosci 2005; 6:591 - 602; http://dx.doi.org/10.1038/nrn1728; PMID: 16025095
- Engelhardt B, Wolburg H. Mini-review: Transendothelial migration of leukocytes: through the front door or around the side of the house?. Eur J Immunol 2004; 34:2955 - 63; http://dx.doi.org/10.1002/eji.200425327; PMID: 15376193
- Nassif X, Bourdoulous S, Eugène E, Couraud PO. How do extracellular pathogens cross the blood-brain barrier?. Trends Microbiol 2002; 10:227 - 32; http://dx.doi.org/10.1016/S0966-842X(02)02349-1; PMID: 11973156
- Janssen RS, Nwanyanwu OC, Selik RM, Stehr-Green JK. Epidemiology of human immunodeficiency virus encephalopathy in the United States. Neurology 1992; 42:1472 - 6; PMID: 1641138
- Reger M, Welsh R, Razani J, Martin DJ, Boone KB. A meta-analysis of the neuropsychological sequelae of HIV infection. J Int Neuropsychol Soc 2002; 8:410 - 24; http://dx.doi.org/10.1017/S1355617702813212; PMID: 11939699
- Navia BA, Cho ES, Petito CK, Price RW. The AIDS dementia complex: II. Neuropathology. Ann Neurol 1986; 19:525 - 35; http://dx.doi.org/10.1002/ana.410190603; PMID: 3014994
- Navia BA, Jordan BD, Price RW. The AIDS dementia complex: I. Clinical features. Ann Neurol 1986; 19:517 - 24; http://dx.doi.org/10.1002/ana.410190602; PMID: 3729308
- Wiley CA, Schrier RD, Morey M, Achim C, Venable JC, Nelson JA. Pathogenesis of HIV encephalitis. Acta Pathol Jpn 1991; 41:192 - 6; PMID: 2068943
- Toborek M, Lee YW, Flora G, Pu H, András IE, Wylegala E, et al. Mechanisms of the blood-brain barrier disruption in HIV-1 infection. Cell Mol Neurobiol 2005; 25:181 - 99; http://dx.doi.org/10.1007/s10571-004-1383-x; PMID: 15962513
- Budka H. The definition of HIV-specific neuropathology. Acta Pathol Jpn 1991; 41:182 - 91; PMID: 2068942
- Tardieu M. HIV-1-related central nervous system diseases. Curr Opin Neurol 1999; 12:377 - 81; http://dx.doi.org/10.1097/00019052-199908000-00002; PMID: 10555824
- Singer EJ, Syndulko K, Fahy-Chandon B, Schmid P, Conrad A, Tourtellotte WW. Intrathecal IgG synthesis and albumin leakage are increased in subjects with HIV-1 neurologic disease. J Acquir Immune Defic Syndr 1994; 7:265 - 71; PMID: 7906304
- Avison MJ, Nath A, Berger JR. Understanding pathogenesis and treatment of HIV dementia: a role for magnetic resonance?. Trends Neurosci 2002; 25:468 - 73; http://dx.doi.org/10.1016/S0166-2236(02)02234-8; PMID: 12183208
- Dallasta LM, Pisarov LA, Esplen JE, Werley JV, Moses AV, Nelson JA, et al. Blood-brain barrier tight junction disruption in human immunodeficiency virus-1 encephalitis. Am J Pathol 1999; 155:1915 - 27; http://dx.doi.org/10.1016/S0002-9440(10)65511-3; PMID: 10595922
- Boven LA, Middel J, Verhoef J, De Groot CJ, Nottet HS. Monocyte infiltration is highly associated with loss of the tight junction protein zonula occludens in HIV-1-associated dementia. Neuropathol Appl Neurobiol 2000; 26:356 - 60; http://dx.doi.org/10.1046/j.1365-2990.2000.00255.x; PMID: 10931369
- Mogensen TH, Paludan SR. Virus-cell interactions: impact on cytokine production, immune evasion and tumor growth. Eur Cytokine Netw 2001; 12:382 - 90; PMID: 11566618
- Mogensen TH, Paludan SR. Molecular pathways in virus-induced cytokine production. Microbiol Mol Biol Rev 2001; 65:131 - 50; http://dx.doi.org/10.1128/MMBR.65.1.131-150.2001; PMID: 11238989
- Gabuzda DH, Sobel RA. HIV antigen in brains of patients with AIDS. Ann Neurol 1987; 22:668; http://dx.doi.org/10.1002/ana.410220526; PMID: 3480698
- Cinque P, Vago L, Mengozzi M, Torri V, Ceresa D, Vicenzi E, et al. Elevated cerebrospinal fluid levels of monocyte chemotactic protein-1 correlate with HIV-1 encephalitis and local viral replication. AIDS 1998; 12:1327 - 32; http://dx.doi.org/10.1097/00002030-199811000-00014; PMID: 9708412
- Bernasconi S, Cinque P, Peri G, Sozzani S, Crociati A, Torri W, et al. Selective elevation of monocyte chemotactic protein-1 in the cerebrospinal fluid of AIDS patients with cytomegalovirus encephalitis. J Infect Dis 1996; 174:1098 - 101; http://dx.doi.org/10.1093/infdis/174.5.1098; PMID: 8896515
- Conant K, McArthur JC, Griffin DE, Sjulson L, Wahl LM, Irani DN. Cerebrospinal fluid levels of MMP-2, 7, and 9 are elevated in association with human immunodeficiency virus dementia. Ann Neurol 1999; 46:391 - 8; http://dx.doi.org/10.1002/1531-8249(199909)46:3<391::AID-ANA15>3.0.CO;2-0; PMID: 10482270
- Marshall DC, Wyss-Coray T, Abraham CR. Induction of matrix metalloproteinase-2 in human immunodeficiency virus-1 glycoprotein 120 transgenic mouse brains. Neurosci Lett 1998; 254:97 - 100; http://dx.doi.org/10.1016/S0304-3940(98)00674-0; PMID: 9779929
- Sporer B, Paul R, Koedel U, Grimm R, Wick M, Goebel FD, et al. Presence of matrix metalloproteinase-9 activity in the cerebrospinal fluid of human immunodeficiency virus-infected patients. J Infect Dis 1998; 178:854 - 7; http://dx.doi.org/10.1086/515342; PMID: 9728558
- Weeks BS, Lieberman DM, Johnson B, Roque E, Green M, Loewenstein P, et al. Neurotoxicity of the human immunodeficiency virus type 1 tat transactivator to PC12 cells requires the Tat amino acid 49-58 basic domain. J Neurosci Res 1995; 42:34 - 40; http://dx.doi.org/10.1002/jnr.490420105; PMID: 8531224
- Hudson L, Liu J, Nath A, Jones M, Raghavan R, Narayan O, et al. Detection of the human immunodeficiency virus regulatory protein tat in CNS tissues. J Neurovirol 2000; 6:145 - 55; http://dx.doi.org/10.3109/13550280009013158; PMID: 10822328
- Arese M, Ferrandi C, Primo L, Camussi G, Bussolino F. HIV-1 Tat protein stimulates in vivo vascular permeability and lymphomononuclear cell recruitment. J Immunol 2001; 166:1380 - 8; PMID: 11145723
- Oshima T, Flores SC, Vaitaitis G, Coe LL, Joh T, Park JH, et al. HIV-1 Tat increases endothelial solute permeability through tyrosine kinase and mitogen-activated protein kinase-dependent pathways. AIDS 2000; 14:475 - 82; http://dx.doi.org/10.1097/00002030-200003310-00002; PMID: 10780709
- Pu H, Tian J, Flora G, Lee YW, Nath A, Hennig B, et al. HIV-1 Tat protein upregulates inflammatory mediators and induces monocyte invasion into the brain. Mol Cell Neurosci 2003; 24:224 - 37; http://dx.doi.org/10.1016/S1044-7431(03)00171-4; PMID: 14550782
- András IE, Pu H, Deli MA, Nath A, Hennig B, Toborek M. HIV-1 Tat protein alters tight junction protein expression and distribution in cultured brain endothelial cells. J Neurosci Res 2003; 74:255 - 65; http://dx.doi.org/10.1002/jnr.10762; PMID: 14515355
- Toborek M, Lee YW, Pu H, Malecki A, Flora G, Garrido R, et al. HIV-Tat protein induces oxidative and inflammatory pathways in brain endothelium. J Neurochem 2003; 84:169 - 79; http://dx.doi.org/10.1046/j.1471-4159.2003.01543.x; PMID: 12485413
- Shi B, De Girolami U, He J, Wang S, Lorenzo A, Busciglio J, et al. Apoptosis induced by HIV-1 infection of the central nervous system. J Clin Invest 1996; 98:1979 - 90; http://dx.doi.org/10.1172/JCI119002; PMID: 8903316
- Acheampong EA, Parveen Z, Muthoga LW, Kalayeh M, Mukhtar M, Pomerantz RJ. Human Immunodeficiency virus type 1 Nef potently induces apoptosis in primary human brain microvascular endothelial cells via the activation of caspases. J Virol 2005; 79:4257 - 69; http://dx.doi.org/10.1128/JVI.79.7.4257-4269.2005; PMID: 15767427
- Price TO, Ercal N, Nakaoke R, Banks WA. HIV-1 viral proteins gp120 and Tat induce oxidative stress in brain endothelial cells. Brain Res 2005; 1045:57 - 63; http://dx.doi.org/10.1016/j.brainres.2005.03.031; PMID: 15910762
- Kanmogne GD, Primeaux C, Grammas P. HIV-1 gp120 proteins alter tight junction protein expression and brain endothelial cell permeability: implications for the pathogenesis of HIV-associated dementia. J Neuropathol Exp Neurol 2005; 64:498 - 505; PMID: 15977641
- Nakamuta S, Endo H, Higashi Y, Kousaka A, Yamada H, Yano M, et al. Human immunodeficiency virus type 1 gp120-mediated disruption of tight junction proteins by induction of proteasome-mediated degradation of zonula occludens-1 and -2 in human brain microvascular endothelial cells. J Neurovirol 2008; 14:186 - 95; http://dx.doi.org/10.1080/13550280801993630; PMID: 18569453
- Kanmogne GD, Schall K, Leibhart J, Knipe B, Gendelman HE, Persidsky Y. HIV-1 gp120 compromises blood-brain barrier integrity and enhances monocyte migration across blood-brain barrier: implication for viral neuropathogenesis. J Cereb Blood Flow Metab 2007; 27:123 - 34; http://dx.doi.org/10.1038/sj.jcbfm.9600330; PMID: 16685256
- Rottman JB, Ganley KP, Williams K, Wu L, Mackay CR, Ringler DJ. Cellular localization of the chemokine receptor CCR5. Correlation to cellular targets of HIV-1 infection. Am J Pathol 1997; 151:1341 - 51; PMID: 9358760
- Sanders VJ, Pittman CA, White MG, Wang G, Wiley CA, Achim CL. Chemokines and receptors in HIV encephalitis. AIDS 1998; 12:1021 - 6; http://dx.doi.org/10.1097/00002030-199809000-00009; PMID: 9662198
- Moses AV, Bloom FE, Pauza CD, Nelson JA. Human immunodeficiency virus infection of human brain capillary endothelial cells occurs via a CD4/galactosylceramide-independent mechanism. Proc Natl Acad Sci U S A 1993; 90:10474 - 8; http://dx.doi.org/10.1073/pnas.90.22.10474; PMID: 7504264
- Moses AV, Nelson JA. HIV infection of human brain capillary endothelial cells--implications for AIDS dementia. Adv Neuroimmunol 1994; 4:239 - 47; http://dx.doi.org/10.1016/S0960-5428(06)80262-7; PMID: 7533040
- Sporer B, Koedel U, Paul R, Kohleisen B, Erfle V, Fontana A, et al. Human immunodeficiency virus type-1 Nef protein induces blood-brain barrier disruption in the rat: role of matrix metalloproteinase-9. J Neuroimmunol 2000; 102:125 - 30; http://dx.doi.org/10.1016/S0165-5728(99)00170-8; PMID: 10636480
- Rostad SW, Sumi SM, Shaw CM, Olson K, McDougall JK. Human immunodeficiency virus (HIV) infection in brains with AIDS-related leukoencephalopathy. AIDS Res Hum Retroviruses 1987; 3:363 - 73; http://dx.doi.org/10.1089/aid.1987.3.363; PMID: 3447627
- Ward JM, O’Leary TJ, Baskin GB, Benveniste R, Harris CA, Nara PL, et al. Immunohistochemical localization of human and simian immunodeficiency viral antigens in fixed tissue sections. Am J Pathol 1987; 127:199 - 205; PMID: 3472469
- Wiley CA, Schrier RD, Nelson JA, Lampert PW, Oldstone MB. Cellular localization of human immunodeficiency virus infection within the brains of acquired immune deficiency syndrome patients. Proc Natl Acad Sci U S A 1986; 83:7089 - 93; http://dx.doi.org/10.1073/pnas.83.18.7089; PMID: 3018755
- Takahashi K, Wesselingh SL, Griffin DE, McArthur JC, Johnson RT, Glass JD. Localization of HIV-1 in human brain using polymerase chain reaction/in situ hybridization and immunocytochemistry. Ann Neurol 1996; 39:705 - 11; http://dx.doi.org/10.1002/ana.410390606; PMID: 8651642
- Liu Y, Tang XP, McArthur JC, Scott J, Gartner S. Analysis of human immunodeficiency virus type 1 gp160 sequences from a patient with HIV dementia: evidence for monocyte trafficking into brain. J Neurovirol 2000; 6:Suppl 1 S70 - 81; PMID: 10871768
- Engelhardt B, Ransohoff RM. The ins and outs of T-lymphocyte trafficking to the CNS: anatomical sites and molecular mechanisms. Trends Immunol 2005; 26:485 - 95; http://dx.doi.org/10.1016/j.it.2005.07.004; PMID: 16039904
- Gras G, Kaul M. Molecular mechanisms of neuroinvasion by monocytes-macrophages in HIV-1 infection. Retrovirology 2010; 7:30; http://dx.doi.org/10.1186/1742-4690-7-30; PMID: 20374632
- Wu DT, Woodman SE, Weiss JM, McManus CM, D’Aversa TG, Hesselgesser J, et al. Mechanisms of leukocyte trafficking into the CNS. J Neurovirol 2000; 6:Suppl 1 S82 - 5; PMID: 10871769
- Jones MV, Bell JE, Nath A. Immunolocalization of HIV envelope gp120 in HIV encephalitis with dementia. AIDS 2000; 14:2709 - 13; http://dx.doi.org/10.1097/00002030-200012010-00010; PMID: 11125889
- Fantuzzi L, Spadaro F, Vallanti G, Canini I, Ramoni C, Vicenzi E, et al. Endogenous CCL2 (monocyte chemotactic protein-1) modulates human immunodeficiency virus type-1 replication and affects cytoskeleton organization in human monocyte-derived macrophages. Blood 2003; 102:2334 - 7; http://dx.doi.org/10.1182/blood-2002-10-3275; PMID: 12805068
- Gonzalez E, Rovin BH, Sen L, Cooke G, Dhanda R, Mummidi S, et al. HIV-1 infection and AIDS dementia are influenced by a mutant MCP-1 allele linked to increased monocyte infiltration of tissues and MCP-1 levels. Proc Natl Acad Sci U S A 2002; 99:13795 - 800; http://dx.doi.org/10.1073/pnas.202357499; PMID: 12374865
- Yadav A, Collman RG. CNS inflammation and macrophage/microglial biology associated with HIV-1 infection. J Neuroimmune Pharmacol 2009; 4:430 - 47; http://dx.doi.org/10.1007/s11481-009-9174-2; PMID: 19768553
- Banks WA, Kastin AJ, Brennan JM, Vallance KL. Adsorptive endocytosis of HIV-1gp120 by blood-brain barrier is enhanced by lipopolysaccharide. Exp Neurol 1999; 156:165 - 71; http://dx.doi.org/10.1006/exnr.1998.7011; PMID: 10192787
- Stolp HB, Dziegielewska KM, Ek CJ, Potter AM, Saunders NR. Long-term changes in blood-brain barrier permeability and white matter following prolonged systemic inflammation in early development in the rat. Eur J Neurosci 2005; 22:2805 - 16; http://dx.doi.org/10.1111/j.1460-9568.2005.04483.x; PMID: 16324115
- Persidsky Y, Stins M, Way D, Witte MH, Weinand M, Kim KS, et al. A model for monocyte migration through the blood-brain barrier during HIV-1 encephalitis. J Immunol 1997; 158:3499 - 510; PMID: 9120312
- Wang H, Sun J, Goldstein H. Human immunodeficiency virus type 1 infection increases the in vivo capacity of peripheral monocytes to cross the blood-brain barrier into the brain and the in vivo sensitivity of the blood-brain barrier to disruption by lipopolysaccharide. J Virol 2008; 82:7591 - 600; http://dx.doi.org/10.1128/JVI.00768-08; PMID: 18508884
- Baskin GB, Murphey-Corb M, Watson EA, Martin LN. Necropsy findings in rhesus monkeys experimentally infected with cultured simian immunodeficiency virus (SIV)/delta. Vet Pathol 1988; 25:456 - 67; http://dx.doi.org/10.1177/030098588802500609; PMID: 2850650
- Sharer LR, Baskin GB, Cho ES, Murphey-Corb M, Blumberg BM, Epstein LG. Comparison of simian immunodeficiency virus and human immunodeficiency virus encephalitides in the immature host. Ann Neurol 1988; 23:Suppl S108 - 12; http://dx.doi.org/10.1002/ana.410230727; PMID: 2831797
- Chakrabarti L, Hurtrel M, Maire MA, Vazeux R, Dormont D, Montagnier L, et al. Early viral replication in the brain of SIV-infected rhesus monkeys. Am J Pathol 1991; 139:1273 - 80; PMID: 1750503
- Hurtrel B, Chakrabarti L, Hurtrel M, Maire MA, Dormont D, Montagnier L. Early SIV encephalopathy. J Med Primatol 1991; 20:159 - 66; PMID: 1942006
- Luabeya MK, Dallasta LM, Achim CL, Pauza CD, Hamilton RL. Blood-brain barrier disruption in simian immunodeficiency virus encephalitis. Neuropathol Appl Neurobiol 2000; 26:454 - 62; http://dx.doi.org/10.1046/j.1365-2990.2000.00275.x; PMID: 11054186
- Ryan G, Grimes T, Brankin B, Mabruk MJ, Hosie MJ, Jarrett O, et al. Neuropathology associated with feline immunodeficiency virus infection highlights prominent lymphocyte trafficking through both the blood-brain and blood-choroid plexus barriers. J Neurovirol 2005; 11:337 - 45; http://dx.doi.org/10.1080/13550280500186445; PMID: 16162477
- Steffan AM, Lafon ME, Gendrault JL, Koehren F, De Monte M, Royer C, et al. Feline immunodeficiency virus can productively infect cultured endothelial cells from cat brain microvessels. J Gen Virol 1994; 75:3647 - 53; http://dx.doi.org/10.1099/0022-1317-75-12-3647; PMID: 7996160
- Fletcher NF, Meeker RB, Hudson LC, Callanan JJ. The neuropathogenesis of feline immunodeficiency virus infection: barriers to overcome. Vet J 2011; 188:260 - 9; http://dx.doi.org/10.1016/j.tvjl.2010.03.022; PMID: 20418131
- Georgsson G. Neuropathologic aspects of lentiviral infections. Ann N Y Acad Sci 1994; 724:50 - 67; http://dx.doi.org/10.1111/j.1749-6632.1994.tb38895.x; PMID: 8030977
- Zink MC, Yager JA, Myers JD. Pathogenesis of caprine arthritis encephalitis virus. Cellular localization of viral transcripts in tissues of infected goats. Am J Pathol 1990; 136:843 - 54; PMID: 2327471
- Osame M, Izumo S, Igata A, Matsumoto M, Matsumoto T, Sonoda S, et al. Blood transfusion and HTLV-I associated myelopathy. Lancet 1986; 2:104 - 5; http://dx.doi.org/10.1016/S0140-6736(86)91636-3; PMID: 2873363
- Osame M, Usuku K, Izumo S, Ijichi N, Amitani H, Igata A, et al. HTLV-I associated myelopathy, a new clinical entity. Lancet 1986; 1:1031 - 2; http://dx.doi.org/10.1016/S0140-6736(86)91298-5; PMID: 2871307
- Gessain A, Barin F, Vernant JC, Gout O, Maurs L, Calender A, et al. Antibodies to human T-lymphotropic virus type-I in patients with tropical spastic paraparesis. Lancet 1985; 2:407 - 10; http://dx.doi.org/10.1016/S0140-6736(85)92734-5; PMID: 2863442
- Jernigan M, Morcos Y, Lee SM, Dohan FC Jr., Raine C, Levin MC. IgG in brain correlates with clinicopathological damage in HTLV-1 associated neurologic disease. Neurology 2003; 60:1320 - 7; PMID: 12707436
- Ozden S, Seilhean D, Gessain A, Hauw JJ, Gout O. Severe demyelinating myelopathy with low human T cell lymphotropic virus type 1 expression after transfusion in an immunosuppressed patient. Clin Infect Dis 2002; 34:855 - 60; http://dx.doi.org/10.1086/338868; PMID: 11830796
- Cartier L, Ramirez E. Presence of HTLV-I Tax protein in cerebrospinal fluid from HAM/TSP patients. Arch Virol 2005; 150:743 - 53; http://dx.doi.org/10.1007/s00705-004-0443-3; PMID: 15592888
- Cavrois M, Gessain A, Gout O, Wain-Hobson S, Wattel E. Common human T cell leukemia virus type 1 (HTLV-1) integration sites in cerebrospinal fluid and blood lymphocytes of patients with HTLV-1-associated myelopathy/tropical spastic paraparesis indicate that HTLV-1 crosses the blood-brain barrier via clonal HTLV-1-infected cells. J Infect Dis 2000; 182:1044 - 50; http://dx.doi.org/10.1086/315844; PMID: 10979898
- Kuroda Y, Matsui M, Takashima H, Kurohara K. Granulocyte-macrophage colony-stimulating factor and interleukin-1 increase in cerebrospinal fluid, but not in serum, of HTLV-I-associated myelopathy. J Neuroimmunol 1993; 45:133 - 6; http://dx.doi.org/10.1016/0165-5728(93)90173-V; PMID: 8331158
- Kuroda Y, Matsui M. Cerebrospinal fluid interferon-gamma is increased in HTLV-I-associated myelopathy. J Neuroimmunol 1993; 42:223 - 6; http://dx.doi.org/10.1016/0165-5728(93)90014-P; PMID: 8429106
- Nakamura S, Nagano I, Yoshioka M, Shimazaki S, Onodera J, Kogure K. Detection of tumor necrosis factor-alpha-positive cells in cerebrospinal fluid of patients with HTLV-I-associated myelopathy. J Neuroimmunol 1993; 42:127 - 30; http://dx.doi.org/10.1016/0165-5728(93)90001-F; PMID: 8429097
- Brosnan CF, Selmaj K, Raine CS. Hypothesis: a role for tumor necrosis factor in immune-mediated demyelination and its relevance to multiple sclerosis. J Neuroimmunol 1988; 18:87 - 94; http://dx.doi.org/10.1016/0165-5728(88)90137-3; PMID: 3257971
- Lehky TJ, Fox CH, Koenig S, Levin MC, Flerlage N, Izumo S, et al. Detection of human T-lymphotropic virus type I (HTLV-I) tax RNA in the central nervous system of HTLV-I-associated myelopathy/tropical spastic paraparesis patients by in situ hybridization. Ann Neurol 1995; 37:167 - 75; http://dx.doi.org/10.1002/ana.410370206; PMID: 7847858
- Jacobson S. Immunopathogenesis of human T cell lymphotropic virus type I-associated neurologic disease. J Infect Dis 2002; 186:Suppl 2 S187 - 92; http://dx.doi.org/10.1086/344269; PMID: 12424696
- Muller S, Boire G, Ossondo M, Ricchiuti V, Smadja D, Vernant JC, et al. IgG autoantibody response in HTLV-I-infected patients. Clin Immunol Immunopathol 1995; 77:282 - 90; http://dx.doi.org/10.1006/clin.1995.1154; PMID: 7586738
- Romero IA, Prevost MC, Perret E, Adamson P, Greenwood J, Couraud PO, et al. Interactions between brain endothelial cells and human T-cell leukemia virus type 1-infected lymphocytes: mechanisms of viral entry into the central nervous system. J Virol 2000; 74:6021 - 30; http://dx.doi.org/10.1128/JVI.74.13.6021-6030.2000; PMID: 10846084
- Afonso PV, Ozden S, Prevost MC, Schmitt C, Seilhean D, Weksler B, et al. Human blood-brain barrier disruption by retroviral-infected lymphocytes: role of myosin light chain kinase in endothelial tight-junction disorganization. J Immunol 2007; 179:2576 - 83; PMID: 17675520
- Khuth ST, Strazielle N, Giraudon P, Belin MF, Ghersi-Egea JF. Impairment of blood-cerebrospinal fluid barrier properties by retrovirus-activated T lymphocytes: reduction in cerebrospinal fluid-to-blood efflux of prostaglandin E2. J Neurochem 2005; 94:1580 - 93; http://dx.doi.org/10.1111/j.1471-4159.2005.03309.x; PMID: 16026393
- Afonso PV, Ozden S, Cumont MC, Seilhean D, Cartier L, Rezaie P, et al. Alteration of blood-brain barrier integrity by retroviral infection. PLoS Pathog 2008; 4:e1000205; http://dx.doi.org/10.1371/journal.ppat.1000205; PMID: 19008946
- Lezin A, Gillet N, Olindo S, Signaté A, Grandvaux N, Verlaeten O, et al. Histone deacetylase mediated transcriptional activation reduces proviral loads in HTLV-1 associated myelopathy/tropical spastic paraparesis patients. Blood 2007; 110:3722 - 8; http://dx.doi.org/10.1182/blood-2007-04-085076; PMID: 17717136
- Boissé L, Gill MJ, Power C. HIV infection of the central nervous system: clinical features and neuropathogenesis. Neurol Clin 2008; 26:799 - 819, x; http://dx.doi.org/10.1016/j.ncl.2008.04.002; PMID: 18657727
- Anthony IC, Bell JE. The Neuropathology of HIV/AIDS. Int Rev Psychiatry 2008; 20:15 - 24; http://dx.doi.org/10.1080/09540260701862037; PMID: 18240059
- Mahajan SD, Aalinkeel R, Sykes DE, Reynolds JL, Bindukumar B, Fernandez SF, et al. Tight junction regulation by morphine and HIV-1 tat modulates blood-brain barrier permeability. J Clin Immunol 2008; 28:528 - 41; http://dx.doi.org/10.1007/s10875-008-9208-1; PMID: 18574677
- Zhang L, Looney D, Taub D, Chang SL, Way D, Witte MH, et al. Cocaine opens the blood-brain barrier to HIV-1 invasion. J Neurovirol 1998; 4:619 - 26; http://dx.doi.org/10.3109/13550289809114228; PMID: 10065903
- Fiala M, Gan XH, Zhang L, House SD, Newton T, Graves MC, et al. Cocaine enhances monocyte migration across the blood-brain barrier. Cocaine’s connection to AIDS dementia and vasculitis?. Adv Exp Med Biol 1998; 437:199 - 205; http://dx.doi.org/10.1007/978-1-4615-5347-2_22; PMID: 9666272
- Gandhi N, Saiyed ZM, Napuri J, Samikkannu T, Reddy PV, Agudelo M, et al. Interactive role of human immunodeficiency virus type 1 (HIV-1) clade-specific Tat protein and cocaine in blood-brain barrier dysfunction: implications for HIV-1-associated neurocognitive disorder. J Neurovirol 2010; 16:294 - 305; http://dx.doi.org/10.3109/13550284.2010.499891; PMID: 20624003
- Lu TS, Avraham HK, Seng S, Tachado SD, Koziel H, Makriyannis A, et al. Cannabinoids inhibit HIV-1 Gp120-mediated insults in brain microvascular endothelial cells. J Immunol 2008; 181:6406 - 16; PMID: 18941231
- Manda VK, Mittapalli RK, Bohn KA, Adkins CE, Lockman PR. Nicotine and cotinine increases the brain penetration of saquinavir in rat. J Neurochem 2010; 115:1495 - 507; http://dx.doi.org/10.1111/j.1471-4159.2010.07054.x; PMID: 20950334
- Manda VK, Mittapalli RK, Geldenhuys WJ, Lockman PR. Chronic exposure to nicotine and saquinavir decreases endothelial Notch-4 expression and disrupts blood-brain barrier integrity. J Neurochem 2010; 115:515 - 25; http://dx.doi.org/10.1111/j.1471-4159.2010.06948.x; PMID: 20722969