
Abstract
The fungal genus Alternaria contains many destructive plant pathogens, including Alternaria brassicicola, which causes black spot disease on a wide range of Brassicaceae plants and which is routinely used as a model necrotrophic pathogen in studies with Arabidopsis thaliana. During host infection, many fungal proteins that are critical for disease progression are processed in the endoplasmic reticulum (ER)/Golgi system and secreted in planta. The unfolded protein response (UPR) is an essential part of ER protein quality control that ensures efficient maturation of secreted and membrane-bound proteins in eukaryotes. This review highlights the importance of the UPR signaling pathway with respect to the ability of A. brassicicola to efficiently accomplish key steps of its pathogenic life cycle. Understanding the pathogenicity mechanisms that fungi uses during infection is crucial for the development of new antifungal therapies. Therefore the UPR pathway has emerged as a promising drug target for plant disease control.
Introduction
The fungal Alternaria genus includes many saprophytic and pathogenic species. It is one of the most common molds associated with allergic diseases and chronic respiratory diseases, such as asthma and chronic rhinosinusitis.Citation1 Moreover, several species cause opportunistic infections in immunocompromised people. This genus is also well-known to contain many destructive plant pathogens. Lawrence et al.Citation2 thus reported that over 4000 Alternaria/host associations have been recorded in the USDA Fungal Host Index, ranking the genus tenth among nearly 2000 fungal genera based on the total number of host records. Alternaria diseases are common on many crops, ornamentals, and a number of weeds. As necrotrophic pathogens, which actively kill host tissue as they colonize and thrive on the contents of dead cells, Alternaria species utilize a variety of secreted pathogenicity factors throughout the infection process.Citation3 For instance, they secrete an arsenal of extracellular compounds and host cell wall degrading enzymes required for plant penetration and nutrient consumption.Citation4,Citation5 In particular, they are known to produce host-specific (HSTs) and/or non-host-specific toxic (NHSTs) toxins.Citation6 Most of these extracellular proteins are folded in the endoplasmic reticulum (ER) prior to secretion, suggesting that the ER folding capacity may be overwhelmed during host infection. We thus hypothesized that the unfolded protein response (UPR) may contribute to necrotrophic virulence by supporting this high secretory load.
The ER orchestrates the processing of nearly all proteins that reside in, or pass through, the endomembrane system of eukaryotic cells. Since a variety of adverse physiological and environmental conditions (such as thermal stress, cell wall stress, hypoxia, nutrient-limiting conditions, or host defense metabolite exposure) can disrupt ER homeostasis, this organelle has evolved a highly specific signaling pathway, termed the UPR, to ensure that its protein-folding capacity is not overwhelmed.Citation7-Citation11 In recent years, considerable efforts have been devoted to identifying components of the fungal UPR pathway, mainly with the aim of overcoming bottlenecks to achieving industrial production of heterologous proteins.Citation12-Citation14 In Saccharomyces cerevisiae, the basic leucine zipper (bZIP)-type transcription factor Hac1 is the major UPR transcriptional regulator. Hac1 synthesis is dependent on splicing of an unconventional intron in the HAC1 mRNA promoted by the ER-located transmembrane protein Ire1.Citation15 This splicing event is activated in response to ER stress and overcomes a translational block, thus allowing translation of Hac1 which regulates the expression of UPR target genes. The predicted proteins encoded by most fungal UPR upregulated genes function as part of the secretory system, including chaperones, foldases, glycosylation enzymes, vesicle transport proteins, and ER-associated degradation proteins. Most of the key elements constituting the signal transduction pathway of the yeast UPR have been identified in filamentous fungi and the central activation mechanism of the pathway, i.e., stress-induced splicing of an unconventional intron, is relatively well conserved among fungi, plants, and even mammalian cells.Citation16 However, the lack of this pathway and the development of alternative mechanisms for the ER stress response were recently reported in Candida glabrata and the fission yeast Schizosaccharomyces pombe.Citation17,Citation18 The UPR also appears to be intimately linked to the ER-associated (protein) degradation pathway, in which misfolded proteins in the ER lumen are retrotranslocated through the translocon to the cytoplasm and are ubiquitinated and degraded by the proteasome.Citation19-Citation21 An additional mechanism, which leads to selective transcriptional downregulation of some genes that encode secreted enzymes, has also been reported in filamentous fungi and termed repression under secretion stress (RESS).Citation22,Citation23 In addition to RESS, Guillemette et al.Citation24 highlighted the possible existence of another feed-back mechanism (that could be termed posttranscriptional RESS, or PT-RESS) occurring at the translational level in A. niger since mRNAs encoding several secreted proteins were found to be redistributed from polysomes to monosomes after ER stress induction. Overall, these ER-stress responses diminish the pool of newly synthesized proteins and provide homeostatic protection for host cells.
In addition to the simple on–off switch model occurring through the canonical Ire1-Hac1 pathway, several other mechanisms have been discovered, which suggest a more complex regulation of the fungal UPR. Leber et al.Citation25 thus detected a process, called S-UPR, which is induced when ER stress is combined with high temperature application or inositol deficiency, two conditions that have an impact on membrane properties. The S-UPR leads to Ire1-independent transcriptional induction of the HAC1 gene and finally results in an increased Hac1 yield in fungal cells. The S-UPR can thus be considered as an adaptation of the classical UPR to more accurately regulate some targets and deal with particular stress conditions. In A. fumigatus, Feng et al.Citation7 also reported differential UPRs according to the ER stress severity and showed that IreA is involved in maintaining continuous ER homeostasis in a HacA-dependent and HacA-independent manner.
Recently, we investigated the importance of the UPR pathway with respect to the pathogenicity of the plant necrotrophic fungus A. brassicicola, the causative agent of black spot disease of Brassicaceae.Citation8 The A. brassicicola–Arabidopsis pathosystem proved to be a suitable model interaction to study molecular aspects of necrotrophic pathogenicity and host plant responses during the plant vegetative and reproductive phases.Citation26
The Alternaria brassicicola–Brassicaceae Pathosystem
Alternaria brassicicola causes dark spot (or black spot) disease, which is one of the most common and destructive fungal diseases of Brassicaceae worldwide. The Brassicaceae family includes many economically important oilseed, vegetable, condiment, and fodder crop species. The pathogen can infect all aerial parts of the plant, including siliques, seeds, and stems. A. brassicicola is notably the dominating Alternaria spp. in Brassica seed crops and may be responsible for high yield losses.Citation27-Citation29 The fungus can overwinter on infected crop residues, seeds, and any related cruciferous weed species (). These sources represent the primary inoculum, which initiate disease in the field. Aerial organs of host plants are affected by typical symptoms corresponding to black necrotic lesions, often surrounded by chlorotic areas (). Then necrotic spots exhibit dense conidia formation on the surface. Conidia may be dispersed relatively long distances by rain and wind and thus constitute the secondary inoculum. At the plant reproductive stage, seed infection through the flowers seems to be less efficient than the fruit route, i.e., direct silique penetration through cellular junctions, replum, and stomata.Citation26 Silique and seed infections are very harmful since they often lead to premature pod shatter and shriveled seed ( and ). No sexual stage occurs during the infection cycle of A. brassicicola, although the genetic structure of populations suggests that sexual recombination occurs.Citation30
Figure 1.A. brassicicola disease cycle. The fungus overwinters on infected crop residues, susceptible weeds, and seed plants (1). It can be seedborne via mycelia within the seed or transitory conidia on the seed surface. Conidia are readily windborne and can be dispersed great distances throughout the growing season (gray arrows represent the aerial spread of conidia). A. brassicicola usually causes damping-off of seedlings (2–3), and dark lesions on aerial parts of adult plants (4). At the plant reproductive stage, seed infection leads to premature pod shatter and shriveled seed (5), causing reduced seed germination and seedling vigor. Black spot disease seriously reduces crop yields, while also reducing the market quality of cauliflower and cabbage heads and oil quality in oilseed species.
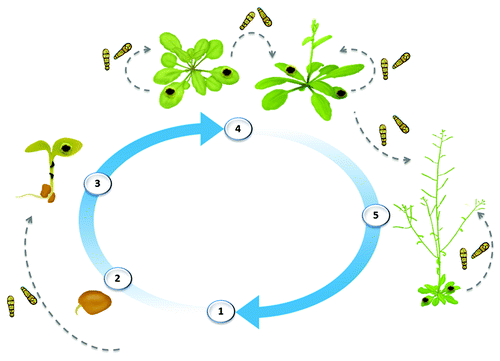
Figure 2. Typical symptoms caused by A. brassicicola on an Arabidopsis silique (A), flower (B), seeds (C) (bars = 1 mm), and cabbage leaf (D) (bars = 1 cm). Arrows show necrotic lesions and/or mycelium development and conidia formation. (E) shows typical A. brassicicola conidia produced from necrotic spots (bars = 10 μm).

As commercial cultivars usually do not express high levels of resistance toward pathogenic Alternaria species, the disease is mainly managed via fungicide treatments by seed dressing and foliar spraying. However, field isolates highly resistant to common broad-spectrum fungicides have been identified, and no fungicides are usually available in organic systems.Citation31 The use of commercial pathogen-free seeds is also essential to limit the spread and incidence of the disease.Citation32 Finally, rotations with noncruciferous crops, crop residue destruction and weed control can limit outbreaks of the disease.
The interaction of Alternaria brassicicola with cruciferous plants, in particular with Arabidopsis, has been extensively used as a model for diseases caused by fungal necrotrophs. A substantial amount of work has been published to characterize plant responses, resistance mechanisms to A. brassicicola and the fungal pathogenicity factor (for a review, see ref. Citation2). Although the breadth of the UPR has been thoroughly analyzed in the plant partner, until very recently no information was available on the UPR impact on the phytopathogenic fungi infectious cycle.Citation33
AbHacA Mediates the UPR in A. brassicicola
The presumed hacA gene (called AbHacA) was identified by a homology search against the A. brassicicola genome assembly (http://genome.jgi-psf.org/Altbr1/Altbr1.home.html) with genes previously described in other filamentous fungi. This gene contains an atypical 20 bp intron that was excised following ER stress. This atypical intron is similar in size to other unconventional hacA introns from fungi (with the exception of 252 nt present in the S. cerevisiae hac1 intron) and has highly conserved border sequences. The unspliced hacA mRNA (hacAu) contains an ORF coding for a 423-amino acid protein. This splicing event alters the C-terminal amino acid sequence of the AbHac1 protein and replaces a portion coding for 187 amino acids, with a different sequence coding for 113 amino acids. By performing complementation tests using a S. cerevisiae hac1 null mutant strain, the authors determined that AbHacA is the ortholog of S. cerevisiae hac1.Citation8
Phenotypic Characteristic of ΔabhacA Mutants
Deletion of AbHacA in A. brassicicola was accomplished by replacing the AbHacA ORF with a hygromycin B resistance cassette.Citation8 Loss of AbHacA is sufficient for disrupting UPR signaling in A. brassicicola. In contrast to the wild-type strain, the mutant strains were indeed unable to increase the level of UPR target genes, such as bipA (coding an ER chaperone) and pdiA (coding a protein disulfide isomerase), when treated with dithiothreitol (DTT) or brefeldin A (BFA), two chemical agents that disrupt ER homeostasis.Citation8 Moreover, AbHacA null strains were growth-impaired in the presence of DTT and BFA concentrations that could be tolerated by the wild-type strain, indicating that AbHacA inactivation increased the sensitivity of A. brassicicola cells to these treatments.Citation8
In addition to the hypersensitivity to UPR-inducing chemical agents, AbHacA null strains share characteristics with other hacA mutants generated in Aspergillus niger, A. fumigatus, Candida albicans, or Cryptococcus neoformans.Citation10,Citation34-Citation36 Most of the mutants are thus hypersensitive to compounds that interfere with cell wall synthesis (Calcofluor white and Congo red), which is indicative of an alteration of the cell wall structure. This feature was confirmed in A. brassicicola conidia by electron microscopy observations and quantification of cell wall compounds. In agreement with the findings of assays performed in A. fumigatus, the amount of glucose was significantly lower in mutant cell walls, suggesting that the glucan fraction is particularly affected by these structural changes.Citation10
Saprophytic growth and hyphal morphology is also strongly affected by the hacA mutation. This phenotype is probably dependent of intracellular vesicular trafficking perturbations and cell wall defects. The germination phase is first affected: wild-type conidia were found to be almost fully germinated after 15 h, while only 20% of Δabhaca conidia were germinated.Citation8 Δabhaca conidia germination rates reached wild-type levels after 24 h. Deletion of AbHacA also dramatically reduced the growth rates and conidiation on PDA (potato dextrose agar) plates at 24 °C. On this medium, the ΔabhacA mutant displayed abnormal hyphae composed of enlarged and swollen cells with many abortive branching structures. Similar observations have been reported in the A. niger hacA mutant.Citation35 However, the mutant growth rates are similar to those of the wild type when the culture medium is synthetic (Vogel’s medium N supplemented with 2% glucose). Conidiation is also partially restored on synthetic medium, which suggests that, even in the absence of ER stress, AbHacA mutants display a reduced secretory capacity and defects in assimilating nutrients from complex substrates. In agreement with this hypothesis, culture supernatants derived from the AbhacA mutant were significantly less efficient at hydrolysing azocoll or p-nitrophenyl butyrate than wild-type cultures, indicating that protease and esterase secretions were impaired in the mutant.Citation8 It was reported that a basal UPR exists in the absence of stress.Citation7,Citation37 This process should allow the cells to make minor adjustments necessary to buffer dynamic fluctuations in ER stress during hyphal growth and maintain continuous ER homeostasis. In a strain lacking the HacA UPR regulator, basal maintenance of ER seems to be difficult or impossible and cause cellular stress quite similar to that generated by chemical agents like DTT and tunicamycin. Loss of AbHacA may thus trigger constitutive activation of secretion stress responses, which are necessarily independent of the canonical HacA pathway. Loss of the secretory capacity of the null mutant under standard conditions could therefore be explained by the constitutive activation of RESS and/or PT-RESS mechanisms that limit the synthesis of secreted proteins.
UPR and A. brassicicola Virulence
The Δabhaca mutants did not cause noticeable disease symptoms on healthy leaves of A. thaliana and B. oleracea and may be considered as avirulent. As few as 50 wild-type conidia in a water drop were sufficient to cause blackspot symptoms on cabbage leaves, while inoculations performed with up to 2500 times more Δabhaca conidia failed to induce typical symptoms.Citation8 The lack of disease symptom development after Δabhaca inoculation on intact leaves was also observed on mechanically wounded leaves or when the conidial drop was supplemented with sorbitol, which stimulated mutant growth. Staining of infection structures with solophenyl flavine or trypan blue and environmental scanning electron microscopic observations showed that both wild-type and mutant conidia germinated with similar efficiency on the epidermal surface and produced similar amounts of appressorial structures with swollen tips.
Using the model pathosystem recently described for investigating seed transmission in Arabidopsis plants, the ability of the AbHacA mutant to transmit to seeds was compared with that of the wild type.Citation26 Silique inoculations with the wild type resulted in mycelium development, conidia formation, and the onset of necrotic lesions typical of black spot on siliques within a few days after inoculation. Some seeds also showed mycelium development and conidia formation. Seeds were then individually harvested 10 d after inoculation and plated on PDA medium, which revealed around 25% global seed transmission rates. In contrast, the ΔabhacA capacity of transmission to A. thaliana seeds was completely abolished. No fungal development was observed from siliques or seeds inoculated with the AbHacA strain.
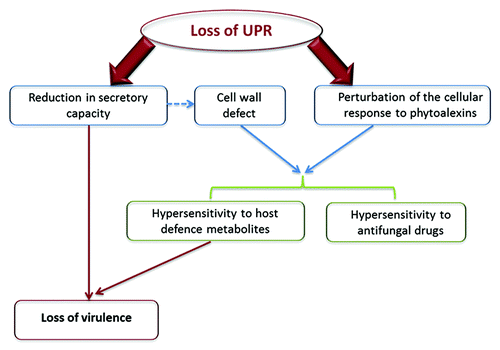
This virulence loss could be explained by at least two features of UPR-deficient mutants (). First, as previously described, they are severely limited in their secretion capacity, which is a major handicap for successful host infection. Like other necrotrophic pathogens, A. brassicicola has a very destructive pathogenesis strategy, resulting in extensive necrosis and tissue maceration. In contrast to biotrophs that feed on living cells, necrotrophs indeed promote the destruction of host cells and derive nutrients from the resulting dead or dying cells. A. brassicicola secretes an arsenal of hydrolytic compounds required for pathogen penetration and nutrient consumption. At early infection stages, the fungus forms penetration pegs, called appressoria, that breach the cuticle and epidermal plant cells. A. brassicicola appressoria are probably not capable of penetrating by physical pressure but presumably secrete a variety of enzymes, such as cell wall-degrading enzymes (CWDEs), proteases, pectinases, and endo-polygalacaturonases, to chemically breakdown the plant surface.Citation4,Citation5 At a later stage, it also produces phytotoxins that likely promote host cell death. UPR may thus contribute to virulence by meeting the increased secretory demand that occurs during host infection.
Figure 3. Different phenotypic characteristics of AbhacA mutants. Loss of virulence may be due to an extreme sensitivity of the UPR-deficient mutant to plant defense metabolites and to a reduced secretory capacity.
Second, in addition to the attenuated secretion, loss of virulence may be due to an extreme sensitivity of the UPR-deficient mutant to various plant defense metabolites. During host infection, A. brassicicola is exposed to high levels of defense compounds, such as phytoalexins and isothiocyanates, the major glucosinolate breakdown products. The in vitro antifungal activities of these compounds against A. brassicicola were assessed by Sellam et al.Citation38 and it was found that the ability to overcome these antimicrobial metabolites is a key factor determining the virulence of fungal necrotrophs.Citation39 AbHaca mutants exhibited increased susceptibility to brassinin and camalexin, two cruciferous indolic phytoalexins, and to allyl-isothiocyanates (AlITC) products. Brassinin and AlITC are produced in a variety of cruciferous vegetables such as cultivated Brassica species, while camalexin is synthesized by A. thaliana and some other brassicaceous weeds.Citation40,Citation41 Δabhaca mutants did not produce any symptoms in the A. thaliana pad3 mutant, which is deficient with respect to camalexin synthesis. However, this Arabidopsis genotype is still able to synthesize other defense compounds at the inoculation site, whose activity can be effective enough to prevent Δabhaca infection.
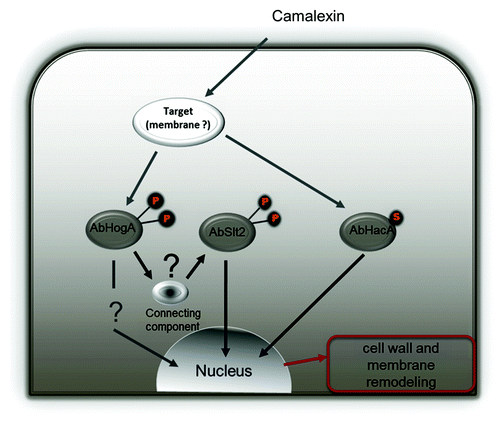
Although the exact mechanism by which cruciferous phytoalexin exerts their toxicity is still unknown, we previously demonstrated that camalexin probably causes membrane damage.Citation42 This toxicity triggers a cellular compensatory response, which particularly involves activating cell wall and membrane repair systems to preserve cell integrity. Recent results showed that two mitogen-activated protein kinase (MAPK) signaling cascades—cell wall integrity (CWI), which plays an essential role in the regulation of the cell wall remodeling process, and high osmolarity glycerol pathways—have a pivotal role in regulating this response during exposure to camalexin.Citation43 Camalexin was found to activate both AbHog1 and AbSlt2 MAP kinases in a precocious manner (10 min of treatment), and activation of the latter was abrogated in an AbHog1 deficient strain. Later (after 2–3 h of exposure), camalexin treatment is sensed as ER stress and subsequently activates UPR target genes, which may further compensate for membrane weakening and contribute to cell wall strengthening. Cell wall strengthening is indeed a potential strategy that occurs in fungal cells to prevent intracellular accumulation of antifungal metabolites. Sellam et al.Citation42 showed that some genes involved in melanin biosynthesis and cell wall maintenance were upregulated in response to camalexin in A. brassicicola, suggesting the reinforcement of a physical barrier against the cellular penetration of toxic compounds. This strategy to escape the host defense response is no longer possible in an UPR-deficient mutant. All of these results suggest that stress responses mediated by UPR and MAPK signaling cascades are coordinated to buffer cells against the toxicity of camalexin ().
Figure 4. Model illustration of fungal signaling pathways regulated by camalexin. Stress responses mediated by UPR and MAPK signaling cascades are coordinated to buffer cells against the toxicity of camalexin. Parallel studies with A. brassicicola knockout mutants showed that camalexin induced AbHog1-dependent phosphorylation of AbSlt2 MAP kinase.Citation43 The UPR signaling pathway is independently activated by production of spliced AbHacA mRNA (AbHacAi).Citation8 This compensatory response may lead to cell wall strengthening and membrane remodeling.
Conclusions and Future Perspectives: UPR Signaling as a Novel Target for Disease Control
UPR plays a major role in the virulence of fungal necrotrophs, in protecting cells against host metabolites and maintaining the ability of the fungus to assimilate nutrients required for growth in the host environment. This pathway is required for efficient leaf or silique tissue colonization. Thus, it does support vertical (i.e., seed transmission) and horizontal (i.e., in planta asexual sporulation) transmission of the pathogen, with both steps being a key element of the fungal disease spreading process. These results suggest that targeting the UPR pathway would be a promising strategy for disease control.
It is likely that this strategy could be applied not only to most necrotrophic fungi but also to biotrophic and hemibiotrophic plant pathogens. Rather than killing the host cells and deriving nutrients from dead cells, biotrophs establish a long-term feeding relationship with the living cells of their hosts. They invade only a few host cells and derive nutrients by differentiation of specialized infection structures termed haustoria.Citation3,Citation44 Other fungi, termed hemibiotrophs, behave first as biotrophs and then switch to a necrotrophic lifestyle.Citation45 For all of these pathogenic lifestyles, in planta secretion of fungal pathogen proteins, including hydrophobins, effector proteins, and other extracellular hydrolytic enzymes is critical for the nutrient acquisition strategy and disease progression. Most of these pathogenic proteins have an N-terminal signal peptide for translocation into the ER, and a functional UPR may be required for their processing in ER during host infection. In agreement with this hypothesis, Yi et al.Citation46 showed that the ER chaperone LHS1 and proper ER function play a role in virulence of the hemibiotroph Magnaporthe orizae. The Δlhs1 mutant showed a defect in the translocation of proteins across the ER membrane and reduced extracellular enzyme activities. Mutation of lhs1 notably impacts the function and secretion of the fungal avirulence effector AVR-Pita. The mutant is severely impaired in conidiation, host penetration, and biotrophic invasion in susceptible rice varieties lacking major gene resistance.
Moreover, we showed that loss of UPR leads to an increased fungal susceptibility to various fungicides commercially used to control fungal crop diseases, such as iprodione and azoxystrobin (unpublished data). Similarly, Richie et al.Citation10 reported that loss of UPR enhances A. fumigatus susceptibility to two major classes of antifungal drugs used for aspergillosis treatment. In response to these agents, fungi generally try to preserve cell integrity by activating cell wall and membrane repair systems. This demand may increase stress on the secretory system and require the UPR function. Moreover, hacA mutants exhibit a cell wall defect, which may participate in its hypersensitivity. These results suggest that pharmacologic inhibition of the UPR could synergize with current drugs in combination therapy. This strategy is a prospective way to decrease the amount of antifungal synthetic molecules in order to reduce the emergence of resistance and meet expectations related to their toxicity.
The next challenge to overcome is to find a therapeutic window to specifically modulate fungal UPR without targeting plant UPR. UPR signaling in plant cells has two pathway branches involving different classes of ER stress transducers. One branch is mediated by the membrane-associated transcription factors bZIP17 and bZIP28, which undergo proteolytic activation and target stress response genes in a similar manner to mammalian ATF6.Citation47 Another branch involves two IRE1 homologs, which catalyze unconventional splicing of mRNA encoding bZIP60, a basic leucine-zipper domain containing transcription factor.Citation48,Citation49 Spliced bZIP60 proteins translocate to the nucleus to activate UPR target genes in a manner similar to HAC1.Citation50,Citation51 IRE1 appears to be an obvious target to modulate the fungal UPR. Targeting kinases is indeed a common approach utilized by drug discovery companies, especially because of the presence of their ATP binding pocket which can be targeted by selective small molecules with favorable pharmaceutical properties. However, specific targeting of fungal IRE1 proteins is challenging as they share structural similarities with plants IRE1 orthologs. The C-terminal regions, which contain kinase and RNase domains, have indeed significant sequence identity with the plant homologous domains. On the contrary, the N-terminal regions, containing the sensor domains of unfolded proteins in ER lumen, are much less conserved among the homologs.Citation48,Citation49 Although the IRE1 ribonuclease domains are well conserved between plants and fungi, they may have specific functional features, which could be exploited for specific UPR inhibitor design. For instance, the yeast HAC1 RNA cannot be spliced in the Arabidopsis protoplasts.Citation48 On the other hand, Wiseman et al.Citation52 recently reported the surprising discovery of an allosteric site at the dimer interface of the yeast Ire1. This ligand-binding pocket could be targeted to modulate ribonuclease activity, providing interesting perspectives for drug development. Hac1 is also an attractive target for plant disease control. Indeed, although bZIP60 mRNA shares predicted secondary structure similarities with yeast HAC1, it does not show apparent sequence homology with HAC1 mRNA, suggesting that specific targeting of fungal UPR is possible to avoid any adverse side effects to the host.Citation53 As they possess an enzymatic activity that can be inhibited by small molecules, other components of the fungal UPR, such as chaperones or protein disulfide isomerases, could also be considered as promising targets. An approach similar to targeting the ATP binding pocket of kinases could be applied to molecules such as ER chaperones that require ATP binding. A powerful approach for the discovery of ATPase inhibitors consists in a high-throughput screening combined with X-ray crystallography and structure-based design.Citation54
Based on the increasing involvement of ER stress in the occurrence of diverse human pathological conditions, considerable recent effort has been devoted to identifying pharmacological modulators of the human UPRosome.Citation54,Citation55 Different approaches led to the identification of selective inhibitors of Ire1 endonuclease activity and which may have therapeutic value.Citation56-Citation58 In the future, these studies should help to identify selective inhibitors of the fungal UPR. Finally, although molecular components of the fungal UPR pathway have been the focus of extensive investigations in recent years, knowledge of HacA-independent pathways involved in ER homeostasis maintenance, i.e., RESS or PT-RESS mechanisms, is still scarce. Gaining insight into the molecular basis of these secretion stress responses should provide additional opportunities to identify new promising drug targets.
| Abbreviations: | ||
| ER | = | endoplasmic reticulum |
| UPR | = | unfolded protein response |
| USDA | = | United States Department of Agriculture |
| HST | = | host-specific toxins |
| NHSTs | = | non-host-specific toxins |
| ERAD | = | ER-associated degradation |
| RESS | = | repression under secretion stress |
| PT-RESS | = | posttranscriptional RESS |
| ORF | = | open reading frame |
| DTT | = | dithiothreitol |
| BFA | = | brefeldin A |
| PDA | = | potato dextrose agar |
| CWDE | = | cell wall degrading enzyme |
| AlITC | = | allyl-isothiocyanate |
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Acknowledgments
This work was partly supported by the French Région Pays de la Loire (QUALISEM research program). We thank David Manley for correcting the English version of the manuscript.
References
- Salo PM, Arbes SJ Jr., Sever M, Jaramillo R, Cohn RD, London SJ, Zeldin DC. Exposure to Alternaria alternata in US homes is associated with asthma symptoms. J Allergy Clin Immunol 2006; 118:892 - 8; http://dx.doi.org/10.1016/j.jaci.2006.07.037; PMID: 17030243
- Lawrence CB, Mitchell TK, Craven KD, Cho Y, Cramer RA, Kim K. At Death’s Door: Alternaria Pathogenicity Mechanisms. Plant Pathol J 2008; 24:101 - 11; http://dx.doi.org/10.5423/PPJ.2008.24.2.101
- Divon HH, Fluhr R. Nutrition acquisition strategies during fungal infection of plants. FEMS Microbiol Lett 2007; 266:65 - 74; http://dx.doi.org/10.1111/j.1574-6968.2006.00504.x; PMID: 17083369
- Cho Y, Cramer RA Jr., Kim KH, Davis J, Mitchell TK, Figuli P, Pryor BM, Lemasters E, Lawrence CB. The Fus3/Kss1 MAP kinase homolog Amk1 regulates the expression of genes encoding hydrolytic enzymes in Alternaria brassicicola. Fungal Genet Biol 2007; 44:543 - 53; http://dx.doi.org/10.1016/j.fgb.2006.11.015; PMID: 17280842
- Fan C, Köller W. Diversity of cutinases from plant pathogenic fungi: differential and sequential expression of cutinolytic esterases by Alternaria brassicicola. FEMS Microbiol Lett 1998; 158:33 - 8; http://dx.doi.org/10.1111/j.1574-6968.1998.tb12796.x
- Friesen TL, Faris JD, Solomon PS, Oliver RP. Host-specific toxins: effectors of necrotrophic pathogenicity. Cell Microbiol 2008; 10:1421 - 8; http://dx.doi.org/10.1111/j.1462-5822.2008.01153.x; PMID: 18384660
- Feng X, Krishnan K, Richie DL, Aimanianda V, Hartl L, Grahl N, Powers-Fletcher MV, Zhang M, Fuller KK, Nierman WC, et al. HacA-independent functions of the ER stress sensor IreA synergize with the canonical UPR to influence virulence traits in Aspergillus fumigatus. PLoS Pathog 2011; 7:e1002330; http://dx.doi.org/10.1371/journal.ppat.1002330; PMID: 22028661
- Joubert A, Simoneau P, Campion C, Bataillé-Simoneau N, Iacomi-Vasilescu B, Poupard P, François JM, Georgeault S, Sellier E, Guillemette T. Impact of the unfolded protein response on the pathogenicity of the necrotrophic fungus Alternaria brassicicola. Mol Microbiol 2011; 79:1305 - 24; http://dx.doi.org/10.1111/j.1365-2958.2010.07522.x; PMID: 21251090
- Kaufman RJ, Scheuner D, Schröder M, Shen X, Lee K, Liu CY, Arnold SM. The unfolded protein response in nutrient sensing and differentiation. Nat Rev Mol Cell Biol 2002; 3:411 - 21; http://dx.doi.org/10.1038/nrm829; PMID: 12042763
- Richie DL, Hartl L, Aimanianda V, Winters MS, Fuller KK, Miley MD, White S, McCarthy JW, Latgé JP, Feldmesser M, et al. A role for the unfolded protein response (UPR) in virulence and antifungal susceptibility in Aspergillus fumigatus. PLoS Pathog 2009; 5:e1000258; http://dx.doi.org/10.1371/journal.ppat.1000258; PMID: 19132084
- Schröder M, Kaufman RJ. ER stress and the unfolded protein response. Mutat Res 2005; 569:29 - 63; http://dx.doi.org/10.1016/j.mrfmmm.2004.06.056; PMID: 15603751
- Archer DB, Turner G. Genomics of protein secretion and hyphal growth in Aspergillus. In: Brown A, ed. The Mycota XIII, Fungal Genomics: Springer, 2006:75-96.
- Conesa A, Punt PJ, van Luijk N, van den Hondel CA. The secretion pathway in filamentous fungi: a biotechnological view. Fungal Genet Biol 2001; 33:155 - 71; http://dx.doi.org/10.1006/fgbi.2001.1276; PMID: 11495573
- Lubertozzi D, Keasling JD. Developing Aspergillus as a host for heterologous expression. Biotechnol Adv 2009; 27:53 - 75; http://dx.doi.org/10.1016/j.biotechadv.2008.09.001; PMID: 18840517
- Rüegsegger U, Leber JH, Walter P. Block of HAC1 mRNA translation by long-range base pairing is released by cytoplasmic splicing upon induction of the unfolded protein response. Cell 2001; 107:103 - 14; http://dx.doi.org/10.1016/S0092-8674(01)00505-0; PMID: 11595189
- Kohno K. Stress-sensing mechanisms in the unfolded protein response: similarities and differences between yeast and mammals. J Biochem 2010; 147:27 - 33; http://dx.doi.org/10.1093/jb/mvp196; PMID: 19942680
- Miyazaki T, Nakayama H, Nagayoshi Y, Kakeya H, Kohno S. Dissection of Ire1 functions reveals stress response mechanisms uniquely evolved in Candida glabrata. PLoS Pathog 2013; 9:e1003160; http://dx.doi.org/10.1371/journal.ppat.1003160; PMID: 23382685
- Kimmig P, Diaz M, Zheng J, Williams CC, Lang A, Aragón T, Li H, Walter P. The unfolded protein response in fission yeast modulates stability of select mRNAs to maintain protein homeostasis. Elife 2012; 1:e00048; http://dx.doi.org/10.7554/eLife.00048; PMID: 23066505
- Hoseki J, Ushioda R, Nagata K. Mechanism and components of endoplasmic reticulum-associated degradation. J Biochem 2010; 147:19 - 25; http://dx.doi.org/10.1093/jb/mvp194; PMID: 19923195
- Richie DL, Feng X, Hartl L, Aimanianda V, Krishnan K, Powers-Fletcher MV, Watson DS, Galande AK, White SM, Willett T, et al. The virulence of the opportunistic fungal pathogen Aspergillus fumigatus requires cooperation between the endoplasmic reticulum-associated degradation pathway (ERAD) and the unfolded protein response (UPR). Virulence 2011; 2:12 - 21; http://dx.doi.org/10.4161/viru.2.1.13345; PMID: 21217201
- Travers KJ, Patil CK, Wodicka L, Lockhart DJ, Weissman JS, Walter P. Functional and genomic analyses reveal an essential coordination between the unfolded protein response and ER-associated degradation. Cell 2000; 101:249 - 58; http://dx.doi.org/10.1016/S0092-8674(00)80835-1; PMID: 10847680
- Al-Sheikh H, Watson AJ, Lacey GA, Punt PJ, MacKenzie DA, Jeenes DJ, Pakula T, Penttilä M, Alcocer MJ, Archer DB. Endoplasmic reticulum stress leads to the selective transcriptional downregulation of the glucoamylase gene in Aspergillus niger. Mol Microbiol 2004; 53:1731 - 42; http://dx.doi.org/10.1111/j.1365-2958.2004.04236.x; PMID: 15341651
- Pakula TM, Laxell M, Huuskonen A, Uusitalo J, Saloheimo M, Penttilä M. The effects of drugs inhibiting protein secretion in the filamentous fungus Trichoderma reesei. Evidence for down-regulation of genes that encode secreted proteins in the stressed cells. J Biol Chem 2003; 278:45011 - 20; http://dx.doi.org/10.1074/jbc.M302372200; PMID: 12941955
- Guillemette T, van Peij NN, Goosen T, Lanthaler K, Robson GD, van den Hondel CA, Stam H, Archer DB. Genomic analysis of the secretion stress response in the enzyme-producing cell factory Aspergillus niger. BMC Genomics 2007; 8:158; http://dx.doi.org/10.1186/1471-2164-8-158; PMID: 17561995
- Leber JH, Bernales S, Walter P. IRE1-independent gain control of the unfolded protein response. PLoS Biol 2004; 2:E235; http://dx.doi.org/10.1371/journal.pbio.0020235; PMID: 15314654
- Pochon S, Terrasson E, Guillemette T, Iacomi-Vasilescu B, Georgeault S, Juchaux M, Berruyer R, Debeaujon I, Simoneau P, Campion C. The Arabidopsis thaliana-Alternaria brassicicola pathosystem: A model interaction for investigating seed transmission of necrotrophic fungi. Plant Methods 2012; 8:16; http://dx.doi.org/10.1186/1746-4811-8-16; PMID: 22571391
- Humpherson-Jones FM. The incidence of Alternaria spp. and Leptosphaeria maculans in commercial brassica seed in the United Kingdom. Plant Pathol 1985; 34:385 - 90; http://dx.doi.org/10.1111/j.1365-3059.1985.tb01377.x
- Kohl J, van Tongeren CAM, Groenenboom-de Haas BH, van Hoof RA, Driessen R, van der Heijden L. Epidemiology of dark leaf spot caused by Alternaria brassicicola and A. brassicae in organic seed production of cauliflower. Plant Pathol 2010; 59:358 - 67; http://dx.doi.org/10.1111/j.1365-3059.2009.02216.x
- Maude RB, Humpherson-Jones FM. Studies on the seed-borne phases of dark leaf spot (Alternaria brassicicola) and grey leaf spot (Alternaria brassicae) of brassicas. Ann Appl Biol 1980; 95:311 - 9; http://dx.doi.org/10.1111/j.1744-7348.1980.tb04752.x
- Bock CH, Thrall PH, Burdon JJ. Genetic structure of populations of Alternaria brassicicola suggests the occurrence of sexual recombination. Mycol Res 2005; 109:227 - 36; http://dx.doi.org/10.1017/S0953756204001674; PMID: 15839106
- Iacomi-Vasilescu B, Bataillé-Simoneau N, Campion C, Dongo A, Laurent E, Serandat I, et al. Effect of null mutations in the AbNIK1 gene on saprophytic and parasitic fitness of Alternaria brassicicola isolates highly resistant to dicarboximides fungicides. Plant Pathol 2008; 57:937 - 47; http://dx.doi.org/10.1111/j.1365-3059.2008.01864.x
- Guillemette T, Iacomi-Vasilescu B, Simoneau P. Conventional and real-time PCR-based assay for detecting pathogenic Alternaria brassicae in cruciferous seed. Plant Dis 2004; 88:490 - 6; http://dx.doi.org/10.1094/PDIS.2004.88.5.490
- Martínez IM, Chrispeels MJ. Genomic analysis of the unfolded protein response in Arabidopsis shows its connection to important cellular processes. Plant Cell 2003; 15:561 - 76; http://dx.doi.org/10.1105/tpc.007609; PMID: 12566592
- Cheon SA, Jung KW, Chen YL, Heitman J, Bahn YS, Kang HA. Unique evolution of the UPR pathway with a novel bZIP transcription factor, Hxl1, for controlling pathogenicity of Cryptococcus neoformans. PLoS Pathog 2011; 7:e1002177; http://dx.doi.org/10.1371/journal.ppat.1002177; PMID: 21852949
- Mulder HJ, Nikolaev I. HacA-dependent transcriptional switch releases hacA mRNA from a translational block upon endoplasmic reticulum stress. Eukaryot Cell 2009; 8:665 - 75; http://dx.doi.org/10.1128/EC.00131-08; PMID: 19181870
- Wimalasena TT, Enjalbert B, Guillemette T, Plumridge A, Budge S, Yin Z, Brown AJ, Archer DB. Impact of the unfolded protein response upon genome-wide expression patterns, and the role of Hac1 in the polarized growth, of Candida albicans. Fungal Genet Biol 2008; 45:1235 - 47; http://dx.doi.org/10.1016/j.fgb.2008.06.001; PMID: 18602013
- Bicknell AA, Babour A, Federovitch CM, Niwa M. A novel role in cytokinesis reveals a housekeeping function for the unfolded protein response. J Cell Biol 2007; 177:1017 - 27; http://dx.doi.org/10.1083/jcb.200702101; PMID: 17562790
- Sellam A, Iacomi-Vasilescu B, Hudhomme P, Simoneau P. In vitro antifungal activity of brassinin, camalexin and two isothiocyanates against the crucifer pathogens Alternaria brassicicola and Alternaria brassicae. Plant Pathol 2007; 56:296 - 301; http://dx.doi.org/10.1111/j.1365-3059.2006.01497.x
- Morrissey JP, Osbourn AE. Fungal resistance to plant antibiotics as a mechanism of pathogenesis. Microbiol Mol Biol Rev 1999; 63:708 - 24; PMID: 10477313
- Fahey JW, Zalcmann AT, Talalay P. The chemical diversity and distribution of glucosinolates and isothiocyanates among plants. Phytochemistry 2001; 56:5 - 51; http://dx.doi.org/10.1016/S0031-9422(00)00316-2; PMID: 11198818
- Pedras MS, Yaya EE, Glawischnig E. The phytoalexins from cultivated and wild crucifers: chemistry and biology. Nat Prod Rep 2011; 28:1381 - 405; http://dx.doi.org/10.1039/c1np00020a; PMID: 21681321
- Sellam A, Dongo A, Guillemette T, Hudhomme P, Simoneau P. Transcriptional responses to exposure to the brassicaceous defence metabolites camalexin and allyl-isothiocyanate in the necrotrophic fungus Alternaria brassicicola. Mol Plant Pathol 2007; 8:195 - 208; http://dx.doi.org/10.1111/j.1364-3703.2007.00387.x; PMID: 20507491
- Joubert A, Bataille-Simoneau N, Campion C, Guillemette T, Hudhomme P, Iacomi-Vasilescu B, Leroy T, Pochon S, Poupard P, Simoneau P. Cell wall integrity and high osmolarity glycerol pathways are required for adaptation of Alternaria brassicicola to cell wall stress caused by brassicaceous indolic phytoalexins. Cell Microbiol 2011; 13:62 - 80; http://dx.doi.org/10.1111/j.1462-5822.2010.01520.x; PMID: 20812995
- Glazebrook J. Contrasting mechanisms of defense against biotrophic and necrotrophic pathogens. Annu Rev Phytopathol 2005; 43:205 - 27; http://dx.doi.org/10.1146/annurev.phyto.43.040204.135923; PMID: 16078883
- Perfect SE, Green JR. Infection structures of biotrophic and hemibiotrophic fungal plant pathogens. Mol Plant Pathol 2001; 2:101 - 8; http://dx.doi.org/10.1046/j.1364-3703.2001.00055.x; PMID: 20572997
- Yi M, Chi MH, Khang CH, Park SY, Kang S, Valent B, Lee YH. The ER chaperone LHS1 is involved in asexual development and rice infection by the blast fungus Magnaporthe oryzae. Plant Cell 2009; 21:681 - 95; http://dx.doi.org/10.1105/tpc.107.055988; PMID: 19252083
- Liu J-X, Howell SH. Endoplasmic reticulum protein quality control and its relationship to environmental stress responses in plants. Plant Cell 2010; 22:2930 - 42; http://dx.doi.org/10.1105/tpc.110.078154; PMID: 20876830
- Noh SJ, Kwon CS, Chung WI. Characterization of two homologs of Ire1p, a kinase/endoribonuclease in yeast, in Arabidopsis thaliana. Biochim Biophys Acta 2002; 1575:130 - 4; http://dx.doi.org/10.1016/S0167-4781(02)00237-3; PMID: 12020828
- Koizumi N, Martinez IM, Kimata Y, Kohno K, Sano H, Chrispeels MJ. Molecular characterization of two Arabidopsis Ire1 homologs, endoplasmic reticulum-located transmembrane protein kinases. Plant Physiol 2001; 127:949 - 62; http://dx.doi.org/10.1104/pp.010636; PMID: 11706177
- Deng Y, Humbert S, Liu JX, Srivastava R, Rothstein SJ, Howell SH. Heat induces the splicing by IRE1 of a mRNA encoding a transcription factor involved in the unfolded protein response in Arabidopsis. Proc Natl Acad Sci U S A 2011; 108:7247 - 52; http://dx.doi.org/10.1073/pnas.1102117108; PMID: 21482766
- Nagashima Y, Mishiba K, Suzuki E, Shimada Y, Iwata Y, Koizumi N. Arabidopsis IRE1 catalyses unconventional splicing of bZIP60 mRNA to produce the active transcription factor. Sci Rep 2011; 1:29; http://dx.doi.org/10.1038/srep00029; PMID: 22355548
- Wiseman RL, Zhang Y, Lee KPK, Harding HP, Haynes CM, Price J, Sicheri F, Ron D. Flavonol activation defines an unanticipated ligand-binding site in the kinase-RNase domain of IRE1. Mol Cell 2010; 38:291 - 304; http://dx.doi.org/10.1016/j.molcel.2010.04.001; PMID: 20417606
- Iwata Y, Koizumi N. Plant transducers of the endoplasmic reticulum unfolded protein response. Trends Plant Sci 2012; 17:720 - 7; http://dx.doi.org/10.1016/j.tplants.2012.06.014; PMID: 22796463
- Healy SJ, Gorman AM, Mousavi-Shafaei P, Gupta S, Samali A. Targeting the endoplasmic reticulum-stress response as an anticancer strategy. Eur J Pharmacol 2009; 625:234 - 46; http://dx.doi.org/10.1016/j.ejphar.2009.06.064; PMID: 19835867
- Woehlbier U, Hetz C. Modulating stress responses by the UPRosome: a matter of life and death. Trends Biochem Sci 2011; 36:329 - 37; http://dx.doi.org/10.1016/j.tibs.2011.03.001; PMID: 21482118
- Cross BC, Bond PJ, Sadowski PG, Jha BK, Zak J, Goodman JM, Silverman RH, Neubert TA, Baxendale IR, Ron D, et al. The molecular basis for selective inhibition of unconventional mRNA splicing by an IRE1-binding small molecule. Proc Natl Acad Sci U S A 2012; 109:E869 - 78; http://dx.doi.org/10.1073/pnas.1115623109; PMID: 22315414
- Papandreou I, Denko NC, Olson M, Van Melckebeke H, Lust S, Tam A, Solow-Cordero DE, Bouley DM, Offner F, Niwa M, et al. Identification of an Ire1alpha endonuclease specific inhibitor with cytotoxic activity against human multiple myeloma. Blood 2011; 117:1311 - 4; http://dx.doi.org/10.1182/blood-2010-08-303099; PMID: 21081713
- Volkmann K, Lucas JL, Vuga D, Wang X, Brumm D, Stiles C, Kriebel D, Der-Sarkissian A, Krishnan K, Schweitzer C, et al. Potent and selective inhibitors of the inositol-requiring enzyme 1 endoribonuclease. J Biol Chem 2011; 286:12743 - 55; http://dx.doi.org/10.1074/jbc.M110.199737; PMID: 21303903