
Abstract
Pathogenic bacteria produce virulence factors called effectors, which are important components of the infection process. Effectors aid in pathogenesis by facilitating bacterial attachment, pathogen entry into or exit from the host cell, immunoevasion, and immunosuppression. Effectors also have the ability to subvert host cellular processes, such as hijacking cytoskeletal machinery or blocking protein translation. However, host cells possess an evolutionarily conserved innate immune response that can sense the pathogen through the activity of its effectors and mount a robust immune response. This “effector triggered immunity” (ETI) was first discovered in plants but recent evidence suggest that the process is also well conserved in metazoans. We will discuss salient points of the mechanism of ETI in metazoans from recent studies done in mammalian cells and invertebrate model hosts.
The immune system consists of two evolutionarily divergent arms: the sophisticated and specific adaptive immunity and the more generic innate immunity. Although the adaptive immune system confers long lasting and protective immunity, it takes several weeks to develop a sustained response and the majority of organisms lack this acquired immune system.Citation1 The innate immune system on the other hand, involves a population of cells and signaling pathways that constitutively function to respond rapidly to pathogens at the site of infection. Innate immune system thus forms the first line of defense, suppressing pathogens or keeping them at bay before the adaptive immune system takes over. This primitive form of immunity is present across multicellular organisms as disparate as nematodes, flies and vertebrates.Citation2 However, not all innate immunity signaling pathways are conserved among metazoans, the nuclear factor kappa-light-chain-enhancer of activated B cells (NFκB) pathway is conserved in vertebrates and flies but not worms whereas the mitogen-activated protein kinases (MAPK) pathway is conserved among all the three. The focus of this review will be on innate immunity, particularly on “effector triggered immunity” (ETI), a process by which bacterial toxins or secreted proteins initiate a protective immune response in the host. During infection, pathogens secrete a broad array of virulence factors called “effector proteins”, which subvert the host cellular processes, including hijacking cytoskeletal machinery, blocking translation and suppressing the immune response.Citation3,Citation4 Non-professional immune cells such as epithelial cells depend on ETI to respond quickly and robustly to pathogens, especially since these cells are constantly exposed to a barrage of microbes including those that form the microbiota. In contrast, professional immune cells like macrophages, which normally reside inside tissues, are less dependent on ETI and respond against all microbes that violate the sanctity of the tissues, irrespective of whether or not they are pathogenic. However, studies done on macrophages infected with Legionella pneumophila also suggest that TLR signals and ETI activation possibly work in concert as a two-signal infection response, which leads to transcriptional upregulation of cytokines and activation of the adaptive immune system.Citation5,Citation6
Detection of Pathogens by the Host
The cells of the innate immune system rely on their pattern recognition receptors (PRR) to recognize conserved pathogen-associated molecular patterns (PAMPs) and microbe-associated molecular patterns (MAMPs) such as microbial nucleic acids, lipoproteins, and carbohydrates that are expressed only in pathogens and not in the host.Citation7 PRRs can be categorized into four families, Toll-like receptors (TLRs), C-type lectin receptors (CLRs), (RIG)-I-like receptors (RLRs), and NOD-like receptors (NLRs).Citation8 TLRs, the best characterized receptors among the PRRs, are transmembrane proteins that recognize lipoprotein, lipopolysaccharide, double stranded RNA, and other ligands associated with diverse pathogens such as bacteria, viruses, and protozoa.Citation9,Citation10 RLRs and NLRs are localized to the cytoplasm and recognize viral nucleic acids and bacterial peptides. PRRs can also recognize cellular damage by binding with products of cellular and tissue degradation, or damage-associated molecular patterns (DAMPs).Citation11 Damaged or necrotic cells release factors such as high mobility group box-1 (HMGB1), serum amyloid A (SAA), and S100A8, which initiate an immune response by engaging TLRs.Citation12-Citation14 DAMPs trigger formation of inflammasomes, which are multimeric protein complexes consisting of caspase 1. Inflammasome formation results in caspase 1 activation, followed by the activation of cytokines IL-1β and IL-18, which induce inflammation.Citation15 Binding with ligands activates the PRRs, which oligomerize and trigger a defense response including activation of NFκB, IRF, and MAPK pathways, signaling the presence of an infection (). This signaling cascade leads to secretion of antimicrobial peptides and attracts cells of the innate and adaptive immune system.Citation16
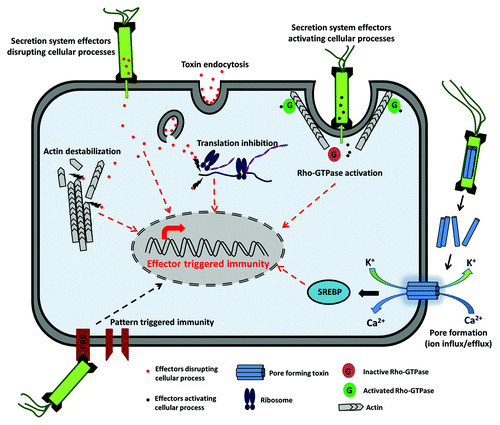
Figure 1. Effector triggered immunity (ETI). ETI can be triggered by toxins that are either directly injected into the host by bacterial secretion systems or internalized from the extracellular environment by endocytosis. Effectors are directly capable of triggering an immune response through transcriptional regulation. Effectors can also disrupt cellular processes such as protein translation and cytoskeletal remodeling, which will trigger an immune response. Some bacterial effectors activate Rho-GTPases, which facilitate bacterial entry and can also trigger ETI. Pore-forming toxins form membrane channels, and the resulting influx/efflux of ions also triggers a protective response.
Beneficial microbes, including commensal bacteria, also possess MAMPs. Therefore, mounting an immune response specifically against harmful pathogens is dependent on the recognition of both the pathogen and the associated host cell damage caused by the pathogen, through MAMPs and DAMPs respectively.Citation11 A decision checkpoint used by phagocytes before amplifying an immune response is the detection of live intracellular bacteria. Following phagocytosis, bacterial mRNA is released only by live bacteria, which is detected by cytosolic PRRs, signaling microbial life to the innate immune system.Citation17 Non-professional immune cells such as intestinal epithelial cells, which are constantly exposed to microbes, detect the presence of pathogens through their cytosolic PRRs and by a polarized distribution of PRRs at the apical and basolateral surfaces. Activation of the PRRs in the cytosol or the basolateral surface will indicate an epithelial cell, or cell surface breach, and attract professional immune cells.Citation18 Pathogens have evolved multiple strategies to avoid detection by modifying MAMPs or subverting PRR signaling.Citation19 Therefore, a rapid immune defense response can be initiated from also monitoring for perturbations in a few core pathways and essential cellular activities, which enables the host to indirectly sense the pathogen instead of evolving specific PRRs for each pathogen or damage-associated molecule.
Effector Triggered Immunity
The defense phenomenon ETI was first observed in plants and our understanding of this phenomenon has evolved starting from the “gene-for-gene theory”, which describes the association between plants and their pathogens through the interaction of pathogen-derived avirulence (Avr) genes and plant derived resistance (R) genes.Citation20 Bacterial effectors are secreted by six distinct secretion systems classified as Type I–VIII.Citation21,Citation22 Pathogenic bacteria deliver their effectors into the plant cells through the type III secretion systems (TTSS) to interfere with plant PAMP-triggered immunity (PTI) and facilitate pathogen survival and dispersal. Plants respond to these challenges by activation of ETI, which triggers release of antimicrobial molecules and hydrolytic enzymes and causes encasement of pathogens and deposition of callus at the infection site.Citation23 For many years, ETI was also speculated to exist in animals, but the cellular mechanisms of ETI activation in metazoans have only just been identified in recent decades.Citation5,Citation24-Citation34 In contrast to the fairly straightforward system for ETI activation in plants, ETI in metazoans can be more broadly defined to encompass host response to cellular damage and to bacterial virulence factors (including effectors) that manipulate central host processes, independent of PAMP and MAMP recognition. ETI in metazoans is triggered by the activation of MAPK and NFκB signaling pathways and there are several excellent reviews describing the downstream effect of this activation.Citation35-Citation37
Blockade of Protein Translation
During an infection, bacterial pathogens cause damage to host cells either directly through their toxins or indirectly by eliciting an adverse immune reaction. The host responds to these challenges by initiating a damage response to maintain cellular integrity and an immune response to restrict bacterial growth.Citation38,Citation39 Bacterial effectors and toxins can blunt these responses by blocking several steps of the host translation machinery.Citation40 However, blockade of protein translation activates ETI and augments the initial immune response (). Legionella pneumophila invades macrophages by translocating over 200 effectors into the host cell through the type IV secretion system to create an intracellular niche ideal for the pathogen to survive and replicate.Citation41,Citation42 The effectors Lgt1, Lgt2, Lgt3, SidI, and SidL inhibit host translation through inactivation of the host elongation factor eEF1a, which activates the NFκB pathway and promotes transcription of stress response genes and pro-inflammatory cytokines.Citation32 Infection of macrophages with L. pneumophila also activates the MAPK pathway.Citation43 Further insights into the effect of translational inhibition on innate immunity was gained from studies done in Caenorhabditis elegans, which has emerged as a popular model for studying host-pathogen relationship and drug discovery.Citation44,Citation45 RNAi-mediated disruption of translation and other essential processes in C. elegans was reported to induce expression of innate immune response genes.Citation46 In C. elegans infected with Pseudomonas aeruginosa, bacterial exotoxin ToxA is internalized by host cells through endocytosis. ToxA blocks translation by inhibiting the host elongation factor 2 (EF-2), triggering expression of the transcription factor ZIP-2, which elicits a protective transcriptional response.Citation30,Citation31,Citation47
In contrast to the above studies, intracellular pathogens such as Shigella flexneri and Salmonella Typhimurium are not known to translocate toxins that directly block host translation. Instead, they cause an overall downregulation of protein synthesis. Infection of epithelial cells with S. flexneri triggers acute amino acid starvation. The resulting induction of the amino acid stress pathway triggers the activation of the GCN2, eIF2α, and ATF3-dependent reprogramming of the transcriptional response in response to invasive pathogens.Citation48 Infection with S. Typhimurium also triggers a similar, albeit transient amino acid stress response, suggesting that the pathogen has evolved strategies to subvert host metabolic stress response pathways. Amino acid starvation can also trigger autophagy, a highly conserved cellular process that is turned on during starvation stress.Citation49 Autophagy functions to recycle damaged cellular organs and complexes in order to maintain levels of essential nutrients such as amino acids.Citation50 Activation of autophagy response against bacteria (xenophagy), and the subsequent activation of the amino acid starvation pathways, represents an important link between innate immunity and metabolic pathways.
Reorganization of the Host Cytoskeleton
The eukaryotic cytoskeleton plays a pivotal role in several cellular processes, including endocytosis, adhesion, migration, phagocytosis, and formation of the immunological synapse.Citation51-Citation54 Genetic mutations, such as those causing Wiskott Aldrich syndrome, affect cytoskeletal regulation and lead to immune deficiency due to impaired function of phagocytes.Citation55-Citation58 Pathogenic bacteria have evolved multiple strategies to manipulate the host cytoskeleton to facilitate intercellular entry and tissue invasion.Citation59 In addition, pathogens have evolved effectors that manipulate the host cytoskeleton to facilitate evasion from the host immune response. In fact, a large number of bacterial effectors have been identified that have an immune inhibitory activity.Citation60 However, when pathogens interfere with the host cytoskeleton for the purpose of immune-evasion, paradoxically, they can also trigger an immune response against the pathogen (). For example, the type III secretion system in pathogenic Yersinia spp. translocates the Yop effectors across the eukaryotic plasma membrane and into the cytosol, where they disrupt key functions of the host cell.Citation61 YopJ inhibits MAPK kinases and MAPKK kinases and also activates Caspase-1, which disrupts the intestinal barrier and promotes dissemination of the bacteria.Citation28 YopE paralyzes the phagocytic functions in macrophages by disrupting the actin microfilament structure.Citation62 The treatment of cultured intestinal epithelial cells with cytochalasin D or latrunculin B, which disrupts the actin cytoskeleton, can cause activation of the p38 MAP kinase and NFκB pathways, showing that subversion of the host actin cytoskeleton components can activate an immune response.Citation63 This was further evident from studies on the effect of Clostridium difficile toxin A on colonic CaCo-2 cells.Citation64 Toxin A causes disruption of the actin cytoskeleton by monoglucosylation of Rho-GTPases Rho, Rac, and Cdc42, which triggers transcriptional upregulation of the p38 MAP kinase pathway. Additionally, Salmonella Typhimurium binds to the surface of epithelial cells and uses type III secretion system effectors to cause a burst of actin polymerization, which induces membrane ruffling and facilitates bacterial entry into the host cell.Citation65 The Salmonella effectors SopE, SopE2, and SopB trigger these events by activating the Rho-GTPases of the host cell in a redundant manner, which in turn can cause a defense response by activating the MAPK and NFκB signaling cascade.Citation29 The CNF1 toxin produced by E. coli is another example of a bacterial protein, which activates Rho-GTPase and can elicit activation of immune pathways.Citation66 Interestingly, studies done in Drosophila and mammalian cells suggest that CNF1 is not injected into the host cell but internalized by receptor-mediated endocytosis into the cytosol, where it covalently modifies the Rho-GTPase Rac 2, triggering protective immunity by activating the Rip kinase signaling pathway.Citation34,Citation67 Enteropathogenic E. coli (EPEC) and enterohemorrhagic E. coli (EHEC) use the type III secretion system to translocate the WxxxE effectors Map, EspM, and EspT into the host cell. These effectors subvert the actin cytoskeleton by mimicking guanine nucleotide exchange factors (GEFs) for activation of Rho-GTPases, which in turn triggers the MAPK and NFκB pathways.Citation68
Pore Forming Toxins
Bacterial pathogens produce virulence factors called pore-forming toxins (PFTs), which attack the cellular membranes of eukaryotic cells. Host proteases recognize and cleave PFTs, which aggregate into oligomeric structures that insert into the membrane to form ionic pores ().Citation69 At high concentrations, PFTs cause rapid death of the host cell due to membrane disruption, leakage of intracellular contents and lysis. However, when PFTs are present in low, sublytic concentration, host cells respond to the damage by activating the ETI response, as seen from studies done in C. elegans and mammalian cells.Citation25,Citation26,Citation70,Citation71 Exposure to PFTs can lead to the activation of NFκB and MAPK pathways, in addition to the unfolded protein response (UPR) and increased autophagy. In epithelial cells, osmotic stress induced by membrane destabilization leads to phosphorylation of p38 MAP kinase. Relieving the osmotic stress by addition of dextran or cellulose can block phosphorylation and activation of p38 MAP kinase.Citation72 The mechanism of activation of immune pathways possibly involves the efflux of potassium ions, as well as an influx of calcium ionsCitation73,Citation74 Treatment of chinese hamster ovary (CHO) cells with aerolysin, a toxin produced by Aeromonas hydrophilia, induces K+ efflux, which subsequently leads to activation and assembly of caspase-1 inflammasome.Citation26 Caspase-1 then induces the activation of the sterol regulatory element binding proteins (SREBPs), which in turn activate the MAPK pathway.Citation75 A role for K+ efflux was similarly proposed for activation of p38 MAPK in HaCaT cells treated with Staphylococcus aureus α-toxin, in which case the immune activation could be neutralized by high concentrations of extracellular K+.Citation73 Mammalian cells subjected to mechanical stress, or toxin-mediated plasma membrane insult cause Ca2+ influx, which triggers a rapid repair process.Citation76 The repair mechanism possibly involves both removal of the damaged area and resealing the membrane by a combination of endocytosis and exocytosis. C. elegans exposed to low doses of the Bacillus thuringiensis toxin Cry5B, resulted in transcriptional upregulation of the genes in the JNK and MAPK pathways.Citation25 The involvement of these pathways in host defense was further demonstrated when C. elegans strains defective in these pathways were more susceptible than the wild type strain to exposure to low doses of Cry5B. A similar response was seen when nematodes were treated with the B. thuringiensis toxin Cry21A. The p38 MAPK pathway is also induced by sublytic concentrations of the cholesterol binding cytolysin (CDC) PFTs such as anthrolysin O (Bacillus anthracis), vaginolysin (Gardnerella vaginalis), pneumolysin (Streptococcus pneumonia), and streptolysin O (Streptococcus aureus).Citation77-Citation80 CDC toxins bind to the cholesterol in the eukaryotic cell membrane to form oligomeric structures that form membrane pores.
Conclusion
Unlike the adaptive immune system, the innate immune system does not need specialized immune cells to mount a protective response. Microbes and their components have a unique molecular signature, which allows early and rapid detection through pattern recognition receptors. However, a system that mounts an antimicrobial response based solely on the molecular signature of microbial components is not only inefficient, but also runs the risk of killing the beneficial commensals that reside in our body. In this respect, effector-triggered immunity (ETI) is an ideal compensatory mechanism that relies primarily on detecting the damage inflicted on the host cell from the microbe, which allows distinguishing between pathogen and commensal. The examples listed in this review provide clear evidence that ETI is a key component of the innate immune response, which is supported by the fact that it is evolutionary conserved across phylogeny in divergent species such as plants, nematodes, and mammals.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
References
- Hoebe K, Janssen E, Beutler B. The interface between innate and adaptive immunity. Nat Immunol 2004; 5:971 - 4; http://dx.doi.org/10.1038/ni1004-971; PMID: 15454919
- Sansonetti PJ. The innate signaling of dangers and the dangers of innate signaling. Nat Immunol 2006; 7:1237 - 42; http://dx.doi.org/10.1038/ni1420; PMID: 17110939
- Mattoo S, Lee YM, Dixon JE. Interactions of bacterial effector proteins with host proteins. Curr Opin Immunol 2007; 19:392 - 401; http://dx.doi.org/10.1016/j.coi.2007.06.005; PMID: 17662586
- Ribet D, Cossart P. Pathogen-mediated posttranslational modifications: A re-emerging field. Cell 2010; 143:694 - 702; http://dx.doi.org/10.1016/j.cell.2010.11.019; PMID: 21111231
- Fontana MF, Banga S, Barry KC, Shen X, Tan Y, Luo ZQ, Vance RE. Secreted bacterial effectors that inhibit host protein synthesis are critical for induction of the innate immune response to virulent Legionella pneumophila. PLoS Pathog 2011; 7:e1001289; http://dx.doi.org/10.1371/journal.ppat.1001289; PMID: 21390206
- Fontana MF, Vance RE. Two signal models in innate immunity. Immunol Rev 2011; 243:26 - 39; http://dx.doi.org/10.1111/j.1600-065X.2011.01037.x; PMID: 21884165
- Akira S, Uematsu S, Takeuchi O. Pathogen recognition and innate immunity. Cell 2006; 124:783 - 801; http://dx.doi.org/10.1016/j.cell.2006.02.015; PMID: 16497588
- Takeuchi O, Akira S. Pattern recognition receptors and inflammation. Cell 2010; 140:805 - 20; http://dx.doi.org/10.1016/j.cell.2010.01.022; PMID: 20303872
- Jin MS, Kim SE, Heo JY, Lee ME, Kim HM, Paik SG, Lee H, Lee JO. Crystal structure of the TLR1-TLR2 heterodimer induced by binding of a tri-acylated lipopeptide. Cell 2007; 130:1071 - 82; http://dx.doi.org/10.1016/j.cell.2007.09.008; PMID: 17889651
- Barbalat R, Lau L, Locksley RM, Barton GM. Toll-like receptor 2 on inflammatory monocytes induces type I interferon in response to viral but not bacterial ligands. Nat Immunol 2009; 10:1200 - 7; http://dx.doi.org/10.1038/ni.1792; PMID: 19801985
- Matzinger P. The danger model: a renewed sense of self. Science 2002; 296:301 - 5; http://dx.doi.org/10.1126/science.1071059; PMID: 11951032
- Park JS, Gamboni-Robertson F, He Q, Svetkauskaite D, Kim JY, Strassheim D, Sohn JW, Yamada S, Maruyama I, Banerjee A, et al. High mobility group box 1 protein interacts with multiple Toll-like receptors. Am J Physiol Cell Physiol 2006; 290:C917 - 24; http://dx.doi.org/10.1152/ajpcell.00401.2005; PMID: 16267105
- Vogl T, Tenbrock K, Ludwig S, Leukert N, Ehrhardt C, van Zoelen MA, Nacken W, Foell D, van der Poll T, Sorg C, et al. Mrp8 and Mrp14 are endogenous activators of Toll-like receptor 4, promoting lethal, endotoxin-induced shock. Nat Med 2007; 13:1042 - 9; http://dx.doi.org/10.1038/nm1638; PMID: 17767165
- He RL, Zhou J, Hanson CZ, Chen J, Cheng N, Ye RD. Serum amyloid A induces G-CSF expression and neutrophilia via Toll-like receptor 2. Blood 2009; 113:429 - 37; http://dx.doi.org/10.1182/blood-2008-03-139923; PMID: 18952897
- Latz E, Xiao TS, Stutz A. Activation and regulation of the inflammasomes. Nat Rev Immunol 2013; 13:397 - 411; http://dx.doi.org/10.1038/nri3452; PMID: 23702978
- Medzhitov R. Approaching the asymptote: 20 years later. Immunity 2009; 30:766 - 75; http://dx.doi.org/10.1016/j.immuni.2009.06.004; PMID: 19538928
- Sander LE, Davis MJ, Boekschoten MV, Amsen D, Dascher CC, Ryffel B, Swanson JA, Müller M, Blander JM. Detection of prokaryotic mRNA signifies microbial viability and promotes immunity. Nature 2011; 474:385 - 9; http://dx.doi.org/10.1038/nature10072; PMID: 21602824
- Abreu MT. Toll-like receptor signalling in the intestinal epithelium: how bacterial recognition shapes intestinal function. Nat Rev Immunol 2010; 10:131 - 44; http://dx.doi.org/10.1038/nri2707; PMID: 20098461
- Roy CR, Mocarski ES. Pathogen subversion of cell-intrinsic innate immunity. Nat Immunol 2007; 8:1179 - 87; http://dx.doi.org/10.1038/ni1528; PMID: 17952043
- Spoel SH, Dong X. How do plants achieve immunity? Defence without specialized immune cells. Nat Rev Immunol 2012; 12:89 - 100; http://dx.doi.org/10.1038/nri3141; PMID: 22273771
- Tseng TT, Tyler BM, Setubal JC. Protein secretion systems in bacterial-host associations, and their description in the Gene Ontology. BMC Microbiol 2009; 9:Suppl 1 S2; http://dx.doi.org/10.1186/1471-2180-9-S1-S2; PMID: 19278550
- Desvaux M, Hébraud M, Talon R, Henderson IR. Secretion and subcellular localizations of bacterial proteins: a semantic awareness issue. Trends Microbiol 2009; 17:139 - 45; http://dx.doi.org/10.1016/j.tim.2009.01.004; PMID: 19299134
- Jones JD, Dangl JL. The plant immune system. Nature 2006; 444:323 - 9; http://dx.doi.org/10.1038/nature05286; PMID: 17108957
- Stuart LM, Paquette N, Boyer L. Effector-triggered versus pattern-triggered immunity: how animals sense pathogens. Nat Rev Immunol 2013; 13:199 - 206; http://dx.doi.org/10.1038/nri3398; PMID: 23411798
- Huffman DL, Abrami L, Sasik R, Corbeil J, van der Goot FG, Aroian RV. Mitogen-activated protein kinase pathways defend against bacterial pore-forming toxins. Proc Natl Acad Sci U S A 2004; 101:10995 - 1000; http://dx.doi.org/10.1073/pnas.0404073101; PMID: 15256590
- Gurcel L, Abrami L, Girardin S, Tschopp J, van der Goot FG. Caspase-1 activation of lipid metabolic pathways in response to bacterial pore-forming toxins promotes cell survival. Cell 2006; 126:1135 - 45; http://dx.doi.org/10.1016/j.cell.2006.07.033; PMID: 16990137
- Bebien M, Hensler ME, Davanture S, Hsu LC, Karin M, Park JM, Alexopoulou L, Liu GY, Nizet V, Lawrence T. The pore-forming toxin β hemolysin/cytolysin triggers p38 MAPK-dependent IL-10 production in macrophages and inhibits innate immunity. PLoS Pathog 2012; 8:e1002812; http://dx.doi.org/10.1371/journal.ppat.1002812; PMID: 22829768
- Meinzer U, Barreau F, Esmiol-Welterlin S, Jung C, Villard C, Léger T, Ben-Mkaddem S, Berrebi D, Dussaillant M, Alnabhani Z, et al. Yersinia pseudotuberculosis effector YopJ subverts the Nod2/RICK/TAK1 pathway and activates caspase-1 to induce intestinal barrier dysfunction. Cell Host Microbe 2012; 11:337 - 51; http://dx.doi.org/10.1016/j.chom.2012.02.009; PMID: 22520462
- Bruno VM, Hannemann S, Lara-Tejero M, Flavell RA, Kleinstein SH, Galán JE. Salmonella Typhimurium type III secretion effectors stimulate innate immune responses in cultured epithelial cells. PLoS Pathog 2009; 5:e1000538; http://dx.doi.org/10.1371/journal.ppat.1000538; PMID: 19662166
- McEwan DL, Kirienko NV, Ausubel FM. Host translational inhibition by Pseudomonas aeruginosa Exotoxin A Triggers an immune response in Caenorhabditis elegans. Cell Host Microbe 2012; 11:364 - 74; http://dx.doi.org/10.1016/j.chom.2012.02.007; PMID: 22520464
- Dunbar TL, Yan Z, Balla KM, Smelkinson MG, Troemel ER. C. elegans detects pathogen-induced translational inhibition to activate immune signaling. Cell Host Microbe 2012; 11:375 - 86; http://dx.doi.org/10.1016/j.chom.2012.02.008; PMID: 22520465
- Fontana MF, Shin S, Vance RE. Activation of host mitogen-activated protein kinases by secreted Legionella pneumophila effectors that inhibit host protein translation. Infect Immun 2012; 80:3570 - 5; http://dx.doi.org/10.1128/IAI.00557-12; PMID: 22851749
- Raymond B, Crepin VF, Collins JW, Frankel G. The WxxxE effector EspT triggers expression of immune mediators in an Erk/JNK and NF-κB-dependent manner. Cell Microbiol 2011; 13:1881 - 93; http://dx.doi.org/10.1111/j.1462-5822.2011.01666.x; PMID: 21848814
- Boyer L, Magoc L, Dejardin S, Cappillino M, Paquette N, Hinault C, Charriere GM, Ip WK, Fracchia S, Hennessy E, et al. Pathogen-derived effectors trigger protective immunity via activation of the Rac2 enzyme and the IMD or Rip kinase signaling pathway. Immunity 2011; 35:536 - 49; http://dx.doi.org/10.1016/j.immuni.2011.08.015; PMID: 22018470
- Arthur JS, Ley SC. Mitogen-activated protein kinases in innate immunity. Nat Rev Immunol 2013; 13:679 - 92; http://dx.doi.org/10.1038/nri3495; PMID: 23954936
- Hayden MS, West AP, Ghosh S. NF-kappaB and the immune response. Oncogene 2006; 25:6758 - 80; http://dx.doi.org/10.1038/sj.onc.1209943; PMID: 17072327
- Vallabhapurapu S, Karin M. Regulation and function of NF-kappaB transcription factors in the immune system. Annu Rev Immunol 2009; 27:693 - 733; http://dx.doi.org/10.1146/annurev.immunol.021908.132641; PMID: 19302050
- Buchon N, Broderick NA, Chakrabarti S, Lemaitre B. Invasive and indigenous microbiota impact intestinal stem cell activity through multiple pathways in Drosophila. Genes Dev 2009; 23:2333 - 44; http://dx.doi.org/10.1101/gad.1827009; PMID: 19797770
- Chatterjee M, Ip YT. Pathogenic stimulation of intestinal stem cell response in Drosophila. J Cell Physiol 2009; 220:664 - 71; http://dx.doi.org/10.1002/jcp.21808; PMID: 19452446
- Chakrabarti S, Liehl P, Buchon N, Lemaitre B. Infection-induced host translational blockage inhibits immune responses and epithelial renewal in the Drosophila gut. Cell Host Microbe 2012; 12:60 - 70; http://dx.doi.org/10.1016/j.chom.2012.06.001; PMID: 22817988
- Zhu W, Banga S, Tan Y, Zheng C, Stephenson R, Gately J, Luo ZQ. Comprehensive identification of protein substrates of the Dot/Icm type IV transporter of Legionella pneumophila. PLoS One 2011; 6:e17638; http://dx.doi.org/10.1371/journal.pone.0017638; PMID: 21408005
- Isberg RR, O’Connor TJ, Heidtman M. The Legionella pneumophila replication vacuole: making a cosy niche inside host cells. Nat Rev Microbiol 2009; 7:13 - 24; http://dx.doi.org/10.1038/nrmicro1967; PMID: 19011659
- Shin S, Case CL, Archer KA, Nogueira CV, Kobayashi KS, Flavell RA, Roy CR, Zamboni DS. Type IV secretion-dependent activation of host MAP kinases induces an increased proinflammatory cytokine response to Legionella pneumophila. PLoS Pathog 2008; 4:e1000220; http://dx.doi.org/10.1371/journal.ppat.1000220; PMID: 19043549
- Desalermos A, Muhammed M, Glavis-Bloom J, Mylonakis E. Using C. elegans for antimicrobial drug discovery. Expert Opin Drug Discov 2011; 6:645 - 52; http://dx.doi.org/10.1517/17460441.2011.573781; PMID: 21686092
- Rajamuthiah R, Fuchs BB, Jayamani E, Kim Y, Larkins-Ford J, Conery A, Ausubel FM, Mylonakis E. Whole animal automated platform for drug discovery against multi-drug resistant Staphylococcus aureus. PLoS One 2014; 9:e89189; http://dx.doi.org/10.1371/journal.pone.0089189; PMID: 24586584
- Melo JA, Ruvkun G. Inactivation of conserved C. elegans genes engages pathogen- and xenobiotic-associated defenses. Cell 2012; 149:452 - 66; http://dx.doi.org/10.1016/j.cell.2012.02.050; PMID: 22500807
- Hubber A, Roy CR. Modulation of host cell function by Legionella pneumophila type IV effectors. Annu Rev Cell Dev Biol 2010; 26:261 - 83; http://dx.doi.org/10.1146/annurev-cellbio-100109-104034; PMID: 20929312
- Tattoli I, Sorbara MT, Vuckovic D, Ling A, Soares F, Carneiro LA, Yang C, Emili A, Philpott DJ, Girardin SE. Amino acid starvation induced by invasive bacterial pathogens triggers an innate host defense program. Cell Host Microbe 2012; 11:563 - 75; http://dx.doi.org/10.1016/j.chom.2012.04.012; PMID: 22704617
- Ghislat G, Patron M, Rizzuto R, Knecht E. Withdrawal of essential amino acids increases autophagy by a pathway involving Ca2+/calmodulin-dependent kinase kinase-β (CaMKK-β). J Biol Chem 2012; 287:38625 - 36; http://dx.doi.org/10.1074/jbc.M112.365767; PMID: 23027865
- Yang Z, Klionsky DJ. Mammalian autophagy: core molecular machinery and signaling regulation. Curr Opin Cell Biol 2010; 22:124 - 31; http://dx.doi.org/10.1016/j.ceb.2009.11.014; PMID: 20034776
- Bokoch GM. Regulation of innate immunity by Rho GTPases. Trends Cell Biol 2005; 15:163 - 71; http://dx.doi.org/10.1016/j.tcb.2005.01.002; PMID: 15752980
- Thrasher AJ, Burns SO. WASP: a key immunological multitasker. Nat Rev Immunol 2010; 10:182 - 92; http://dx.doi.org/10.1038/nri2724; PMID: 20182458
- Misra A, George B, Rajmohan R, Jain N, Wong MH, Kambadur R, Thanabalu T. Insulin receptor substrate protein 53kDa (IRSp53) is a negative regulator of myogenic differentiation. Int J Biochem Cell Biol 2012; 44:928 - 41; http://dx.doi.org/10.1016/j.biocel.2012.02.020; PMID: 22465711
- Misra A, Rajmohan R, Lim RP, Bhattacharyya S, Thanabalu T. The mammalian verprolin, WIRE induces filopodia independent of N-WASP through IRSp53. Exp Cell Res 2010; 316:2810 - 24; http://dx.doi.org/10.1016/j.yexcr.2010.07.015; PMID: 20678498
- Lorenzi R, Brickell PM, Katz DR, Kinnon C, Thrasher AJ. Wiskott-Aldrich syndrome protein is necessary for efficient IgG-mediated phagocytosis. Blood 2000; 95:2943 - 6; PMID: 10779443
- Leverrier Y, Lorenzi R, Blundell MP, Brickell P, Kinnon C, Ridley AJ, Thrasher AJ. Cutting edge: the Wiskott-Aldrich syndrome protein is required for efficient phagocytosis of apoptotic cells. J Immunol 2001; 166:4831 - 4; http://dx.doi.org/10.4049/jimmunol.166.8.4831; PMID: 11290758
- Rajmohan R, Raodah A, Wong MH, Thanabalu T. Characterization of Wiskott-Aldrich syndrome (WAS) mutants using Saccharomyces cerevisiae. FEMS Yeast Res 2009; 9:1226 - 35; http://dx.doi.org/10.1111/j.1567-1364.2009.00581.x; PMID: 19817875
- Rajmohan R, Meng L, Yu S, Thanabalu T. WASP suppresses the growth defect of Saccharomyces cerevisiae las17Delta strain in the presence of WIP. Biochem Biophys Res Commun 2006; 342:529 - 36; http://dx.doi.org/10.1016/j.bbrc.2006.01.160; PMID: 16488394
- Lemichez E, Aktories K. Hijacking of Rho GTPases during bacterial infection. Exp Cell Res 2013; 319:2329 - 36; http://dx.doi.org/10.1016/j.yexcr.2013.04.021; PMID: 23648569
- Brodsky IE, Medzhitov R. Targeting of immune signalling networks by bacterial pathogens. Nat Cell Biol 2009; 11:521 - 6; http://dx.doi.org/10.1038/ncb0509-521; PMID: 19404331
- Cornelis GR, Wolf-Watz H. The Yersinia Yop virulon: a bacterial system for subverting eukaryotic cells. Mol Microbiol 1997; 23:861 - 7; http://dx.doi.org/10.1046/j.1365-2958.1997.2731623.x; PMID: 9076724
- Black DS, Bliska JB. The RhoGAP activity of the Yersinia pseudotuberculosis cytotoxin YopE is required for antiphagocytic function and virulence. Mol Microbiol 2000; 37:515 - 27; http://dx.doi.org/10.1046/j.1365-2958.2000.02021.x; PMID: 10931345
- Németh ZH, Deitch EA, Davidson MT, Szabó C, Vizi ES, Haskó G. Disruption of the actin cytoskeleton results in nuclear factor-kappaB activation and inflammatory mediator production in cultured human intestinal epithelial cells. J Cell Physiol 2004; 200:71 - 81; http://dx.doi.org/10.1002/jcp.10477; PMID: 15137059
- Gerhard R, Tatge H, Genth H, Thum T, Borlak J, Fritz G, Just I. Clostridium difficile toxin A induces expression of the stress-induced early gene product RhoB. J Biol Chem 2005; 280:1499 - 505; http://dx.doi.org/10.1074/jbc.M406014200; PMID: 15531589
- Hardt WD, Chen LM, Schuebel KE, Bustelo XR, Galán JES. S. typhimurium encodes an activator of Rho GTPases that induces membrane ruffling and nuclear responses in host cells. Cell 1998; 93:815 - 26; http://dx.doi.org/10.1016/S0092-8674(00)81442-7; PMID: 9630225
- Lerm M, Selzer J, Hoffmeyer A, Rapp UR, Aktories K, Schmidt G. Deamidation of Cdc42 and Rac by Escherichia coli cytotoxic necrotizing factor 1: activation of c-Jun N-terminal kinase in HeLa cells. Infect Immun 1999; 67:496 - 503; PMID: 9916051
- Lemonnier M, Landraud L, Lemichez E. Rho GTPase-activating bacterial toxins: from bacterial virulence regulation to eukaryotic cell biology. FEMS Microbiol Rev 2007; 31:515 - 34; http://dx.doi.org/10.1111/j.1574-6976.2007.00078.x; PMID: 17680807
- Hobbie S, Chen LM, Davis RJ, Galán JE. Involvement of mitogen-activated protein kinase pathways in the nuclear responses and cytokine production induced by Salmonella typhimurium in cultured intestinal epithelial cells. J Immunol 1997; 159:5550 - 9; PMID: 9548496
- Parker MW, Feil SC. Pore-forming protein toxins: from structure to function. Prog Biophys Mol Biol 2005; 88:91 - 142; http://dx.doi.org/10.1016/j.pbiomolbio.2004.01.009; PMID: 15561302
- Aroian R, van der Goot FG. Pore-forming toxins and cellular non-immune defenses (CNIDs). Curr Opin Microbiol 2007; 10:57 - 61; http://dx.doi.org/10.1016/j.mib.2006.12.008; PMID: 17234446
- Freche B, Reig N, van der Goot FG. The role of the inflammasome in cellular responses to toxins and bacterial effectors. Semin Immunopathol 2007; 29:249 - 60; http://dx.doi.org/10.1007/s00281-007-0085-0; PMID: 17805541
- Ratner AJ, Hippe KR, Aguilar JL, Bender MH, Nelson AL, Weiser JN. Epithelial cells are sensitive detectors of bacterial pore-forming toxins. J Biol Chem 2006; 281:12994 - 8; http://dx.doi.org/10.1074/jbc.M511431200; PMID: 16520379
- Kloft N, Busch T, Neukirch C, Weis S, Boukhallouk F, Bobkiewicz W, Cibis I, Bhakdi S, Husmann M. Pore-forming toxins activate MAPK p38 by causing loss of cellular potassium. Biochem Biophys Res Commun 2009; 385:503 - 6; http://dx.doi.org/10.1016/j.bbrc.2009.05.121; PMID: 19497299
- McNeil PL, Steinhardt RA. Plasma membrane disruption: repair, prevention, adaptation. Annu Rev Cell Dev Biol 2003; 19:697 - 731; http://dx.doi.org/10.1146/annurev.cellbio.19.111301.140101; PMID: 14570587
- Kotzka J, Müller-Wieland D, Roth G, Kremer L, Munck M, Schürmann S, Knebel B, Krone W. Sterol regulatory element binding proteins (SREBP)-1a and SREBP-2 are linked to the MAP-kinase cascade. J Lipid Res 2000; 41:99 - 108; PMID: 10627507
- Idone V, Tam C, Goss JW, Toomre D, Pypaert M, Andrews NW. Repair of injured plasma membrane by rapid Ca2+-dependent endocytosis. J Cell Biol 2008; 180:905 - 14; http://dx.doi.org/10.1083/jcb.200708010; PMID: 18316410
- Park JM, Ng VH, Maeda S, Rest RF, Karin M. Anthrolysin O and other gram-positive cytolysins are toll-like receptor 4 agonists. J Exp Med 2004; 200:1647 - 55; http://dx.doi.org/10.1084/jem.20041215; PMID: 15611291
- Gelber SE, Aguilar JL, Lewis KL, Ratner AJ. Functional and phylogenetic characterization of Vaginolysin, the human-specific cytolysin from Gardnerella vaginalis. J Bacteriol 2008; 190:3896 - 903; http://dx.doi.org/10.1128/JB.01965-07; PMID: 18390664
- Kim YJ, Shin HS, Lee JH, Jung YW, Kim HB, Ha UH. Pneumolysin-mediated expression of β-defensin 2 is coordinated by p38 MAP kinase-MKP1 in human airway cells. J Microbiol 2013; 51:194 - 9; http://dx.doi.org/10.1007/s12275-013-2579-x; PMID: 23625220
- Stassen M, Müller C, Richter C, Neudörfl C, Hültner L, Bhakdi S, Walev I, Schmitt E. The streptococcal exotoxin streptolysin O activates mast cells to produce tumor necrosis factor alpha by p38 mitogen-activated protein kinase- and protein kinase C-dependent pathways. Infect Immun 2003; 71:6171 - 7; http://dx.doi.org/10.1128/IAI.71.11.6171-6177.2003; PMID: 14573633