
Abstract
This study explores the correlation between human carbonic anhydrase (CA, EC 4.2.1.1) isoforms I and II (hCA I, II) and the inhibitory features of some spirobisnaphthalene derivatives. A group of spirobisnaphthalenes was synthesized and their hCA I and II inhibitory effects was investigated. The Ki values were similar for both CA isoenzymes, the compounds showing good inhibitory activity. Ki values ranged between 1.60 and 460.42 µM for hCA I and between 0.39 and 419.42 µM for hCA II, respectively. The spirobisnaphthalenes derivatives might be useful for designing CA inhibitors belonging to novel chemotypes compared to the highly investigated sulfonamides, sulfamates or coumarins.
Introduction
The spirobisnaphthalenes exhibit a wide range of biological effects such as antifungal and antibacterialCitation1–4, antimitoticCitation5, antileishmanialCitation6 and antitumorCitation5,Citation7 activities. The antitumor and antimitotic effect may result from the phospholipase DCitation8–10, DNA gyraseCitation11 or thioredoxin-reductaseCitation12–14 inhibition. The palmarumycins belong to a relatively new and rare family of bioactive natural products and have a unique structural feature. They consist of a 1,8-dihydroxynaphthalene unit and a partially reduced naphthalene unit, which are connected to each other by a spiroacetal. The first representative, called bipendensin, was obtained in very small amounts from wood samples of Afzelia bipendensisCitation15. However, it is assumed that a fungus living in this plantCitation16 produced this compound. Meanwhile, a large number of such spiro compounds were isolated mostly from endophytic fungi and named diepoxinesCitation1, Sch plus numberCitation8,Citation17,Citation18, CJ-plus numberCitation11, cladosporinsCitation19–21, palmarumycinsCitation3,Citation22–24, sphaerolonesCitation25, decaspironesCitation4,Citation17 and deoxypreussomerinCitation26.
The interesting structures, various biological activities and potential applications in agriculture, medicine and the food industry attracted the attention of many research groups with respect to the synthesis of spirobisnaphthalenes. In the last few years, many investigations have been carried out and several natural compounds of this family have been identified, analyzed and synthesizedCitation27.
Carbonic anhydrase (CA, EC 4.2.1.1) is a metalloenzyme and is fairly ubiquitous, being found in animals, plants and in the microorganisms. CAs found in animal cells were shown to be different from CAs of plants or other oranismsCitation28–30. Indeed, the CAs possess different structures, activities and isoforms, with six distinct genetic CA families (α-, β-, γ-, δ-, ζ- and η-CAs) being described nowadaysCitation30–32.
The human CAs belongs to α-class. Until now, 16 isozymes have been determined. These 16 isozymes differ by molecular specifications, oligomeric arrangement, cellular localization, kinetic properties and tissues, expression levels, distribution. Some studies showed important roles of CAs in a diversity of physiological processes, and demonstrated that activities or unusual levels of these enzymes have been often associated with various human diseasesCitation33–35.
Human catalytically active α-CA have different subcellular localization, where CA I, II, III, VII and XIII are in cytosol, CA IV, IX, XII and XIV are membrane-associated and CA VB and VA are in mitochondria. CA VI is secreted in milk and salivaCitation36–39.
CA is a well-characterized pH-regulatory metalloenzyme widely found in many tissues including red blood cells, gastrointestinal tract, kidneys, lungs, etc.Citation40,Citation41 It quickly catalyzes the hydration of carbon dioxide to form bicarbonate, as well as the reversible dehydration reaction of bicarbonate which generates CO2Citation42,Citation43.
Inhibitors of carbonic anhydrases (CAIs) have been developed and used for the treatment of various conditions. Some inhibitors are used clinically as antiglaucoma agents; for the management of neurological disorders, as diuretics, in the treatment of osteoporosis or as antiepileptic agents. Ultimately a sulfonamide CA inhibitor entered Phase I clinical trials as an antitumor agentCitation44. Besides on these applications of CA inhibitors, there are continued efforts in developing diverse agents for the management of cancer, obesity and other diseases in which the activity of these enzymes is disregulatedCitation45–49.
This group has reported some inhibition studies of several CA isozymes with sulfamides. As sulfamide exhibits important biological activities, CA inhibitory properties of some sulfamide derivatives will be useful for further biological studies. It has been shown that this simple compound behaves as a weak inhibitor against the cytosolic isoforms CA I and II. Sulfonamides have R-SO2NH2 general chemical structure. According to the recent studies, sulfonamides have high affinity for CA. Especially for hCA isoenzymes inhibitor studies increased in recent years. At present, strong CAIs have been discovered using modified moieties such as sulfonamide or sulfamate among othersCitation50.
The synthesis of spirobisnaphthalenes derivatives (1–16) was performed as described previouslyCitation51. In this study, we have examined the inhibition effects of spirobisnaphthalenes derivatives (1–16) against the both human cytosolic CA isoforms (hCA I and II). Also, the inhibition results were compared to acetazolamide (AZA) and dorzolamide (DRZ).
Materials and methods
The human erythrocytes were centrifuged at 10 000× g for 0.5 h. Then precipitate and the serum were separated. The serum pH was adjusted with solid Tris (pH 8.7)Citation52–55. As well as sepharose-4B-tirozyne-sulfanylamide affinity column balanced with Tris–HCl/Na2SO4 (25 mM/0.1 M, pH 8.7). Then, this sample was applied to the Sepharose-4B-tirozyne-sulfanylamide affinity column and equilibrated with Tris–HCl/Na2SO4 (25 mM/22 mM, pH 8.7). Finally, HCA I and II isozymes were eluted with 1.0 M NaCl/25 mM sodium phosphate (pH 6.3) and 0.1 M sodium acetate/0.5 M NaClO4 (pH 5.6), respectively. All of enzyme studies were performed at 4 °CCitation56–60. The protein content during the purification steps, Bradford method was usedCitation61. Bovine serum albumin was used as standardCitation62–64.
The purity of both hCA I and II was controlled by SDS polyacrylamide gel electrophoresis (SDS–PAGE)Citation65. This method had been described in previous studiesCitation66,Citation67. The running and separating gels contained SDS (0.1%) and 3% and 10% acrylamide, respectivelyCitation68,Citation69. Before loading into the gel, samples were added 1% SDS and 10% 2-mercaptoethanol and then were waited in boiling water bath for 5 minCitation70. The gel was painted in coloring solution after bands were detected. This coloring solution is prepared in (10%) acetic acid and methanol (50%), which contain Coomassie Brilliant Blue (R-250, 0.1%)Citation71.
During the isoenzyme purification and inhibition process, esterase activity studies were performed according to the Verpoorte procedureCitation72. Both CA isoenzymes activities were determined by following the change in absorbance at 348 nmCitation73. The Ki values were calculated from experiments using three different spirobisnaphthalenes derivative (1–16) concentrations and NPA as the substrate at five different concentrations to create Lineweaver–Burk curves reported previouslyCitation74–76.
Results and discussion
CA purification and activity assay
First, hCA I and II isoenzymes were purified by sepharose-4B-l-thyrosine-sulfanilamide chromatography. Then, the purity of the enzymes was detected by SDS–PAGECitation53,Citation54,Citation77 and single band was observed for each isoenzyme. We studied the inhibition effects of spirobisnaphthalenes derivatives against hCA I and II. For this purpose, the esterase activity method was performedCitation78. In this activity determination method, the phenyl acetate is hydrolyzed and the resulting product gives the absorbance at 348 nmCitation56,Citation57,Citation79.
In the last decades, many valuable bioactive secondary metabolites, which show a great variety of biological activities, have been successfully isolated from the endophytic fungi. The spirobisnaphthalenes are a growing group of fungal secondary metabolites, which contain two 1,8-dihydroxynaphthalene-derived units bridged through a spiroketal linkage. This group could be mainly classified as spairoxin, preussomerin, palmarumycin and urnucratin-type spirobisnaphthalenes, according to their structural featuresCitation51,Citation80. The chemical formula of spirobisnaphthalenes, which are used in this study, are given in .
Figure 1. The chemical structures of synthesized spirobisnaphthalenes derivatives (1–16).
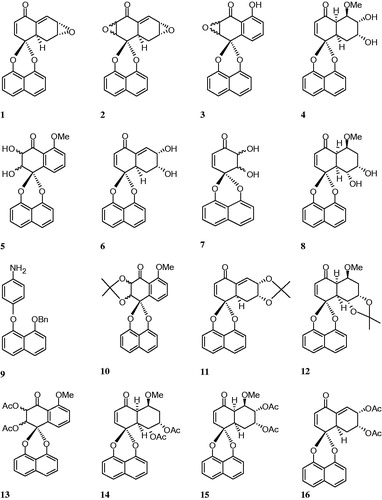
The CA isoenzymes have become an interesting target for the design of activators or inhibitors with biomedical applications. With this purpose, we have investigated the inhibitory effects of many compounds on the CA isoenzymes. In this circumstance, Lineweaver–Burk graphs were drawn for determination of inhibition effect of each spirobisnaphthalenes derivate. Then, the average of IC50 and Ki values were calculated from drawn graphs (). A first step, we report the inhibitory effects of derivatives spirobisnaphthalenes on the esterase activity of hCA I and II under the in vitro conditions. The results were summarized in .
Table 1. Human carbonic anhydrase I and II isoenzymes (hCA I and II) inhibition values of spirobisnaphthalenes derivatives (1–16), by an esterase assay with NPA.
Spirobisnaphthalenes were given to be effect inhibitors of hCA I, II (). Spirobisnaphthalenes derivatives (1–16) were all determined to inhibit hCA I, with Ki values ranging of 1.60–460.42 µM for spirobisnaphthalenes derivatives. Spirobisnaphthalenes derivatives (1–16) were all found to inhibit hCA II, with Ki values ranging of 0.39–419.42 µM for spirobisnaphthalenes derivatives.
These results demonstrated that hCA I and II are inhibited by spirobisnaphthalenes derivatives in the micromolar range. The mechanism by which these compounds inhibit the enzyme is not well understood at this moment and work is in progress to decipher it.
Conclusion
Carbonic anhydrase inhibitory properties of spirobisnaphthalenes derivatives (1–16) have been evaluated. These biologically active compounds generally demonstrated effective inhibition against both hCA I and II isoenzymes. Spirobisnaphthalenes 1–16 behaved as micromolar inhibitors of hCA I and II and may be used for generating more potent hCA I and II inhibitors for the treatment of glaucoma, as diuretics, for the management of mountain sickness, epilepsy, neurological disorders, gastric and duodenal ulcers, osteoporosis conditions.
Declaration of interest
The authors have declared no conflict of interest.
References
- Schlingmann G, West RR, Milne L, et al. Diepoxins, novel fungal metabolites with antibiotic activity. Tetrahedron Lett 1993;34:7225–8
- Krohn K, Michel A, Flörke U, et al. Palmarumycins CP1–CP4 from Coniothyrium palmarum: isolation, structure elucidation, and biological activity. Liebigs Ann Chem 1994;11:1093–7
- Krohn K, Michel A, Flörke U, et al. Palmarumycins C1–C16 from Coniothyrium sp.: isolation, structure elucidation, and biological activity. Liebigs Ann Chem 1994;11:1099–108
- Jiao P, Swenson DC, Gloer JB, et al. Bioactive spirodioxynaphthalenes from the freshwater aquatic fungus Decaisnella thyridioides. J Nat Prod 2006;69:1667–71
- Lazo JS, Tamura AZ, Vogt A, et al. Antimitotic actions of a novel analog of the fungal metabolite palmarumycin CP1. J Pharmacol Exp Ther 2001;296:364–71
- Martinnez-Luis S, Della-Togna G, Coley PD, et al. Antileishmanial constituents of the Panamanian endophytic fungus Edenia sp. J Nat Prod 2008;71:2011–14
- Chu M, Truumees I, Patel MG, et al. Sch 50673 and Sch 50676, two novel antitumor fungal metabolites. Antibiotics 1995;48:329–31
- Chu M, Patel MG, Pai MG, et al. Sch 53823 and sch 53825, novel fungal metabolites with phospholipase D inhibitory activity. Bioorg Med Chem Lett 1996;6:579–84
- Chu M, Truumers I, Patel GM, et al. 2 New phospholipase-d inhibitors, sch-49211 and sch-49212, produced by the fungus Nattrassia-mangiferae. Bioorg Med Chem Lett 1994;4:1539–42
- Pai JK, Frank EA, Blood C, Chu M. Novel ketoepoxides block phospholipase D activation and tumor cell invasion. Anticancer Drug Des 1994;9:363–72
- Sakemi S, Inagaki T, Kaneda K, et al. CJ-12,371 and CJ-12,372, two novel DNAgyrase inhibitors. Fermentation, isolation, structural elucidation and biological activities. J Antibiot 1995;27:134–42
- Wipf P, Hopkins TD, Jung JJ, et al. New inhibitors of the thioredoxin–thioredoxin reductase system based on a naphthoquinone spiroketal natural product lead. Bioorg Med Chem Lett 2001;11:2637–41
- Wipf P, Lynch SM, Birmingham A, et al. Natural product based inhibitors of the thioredoxin–thioredoxin reductase system. Org Biomol Chem 2004;2:1651–8
- Powis G, Wipf P, Lynch SM, et al. Molecular pharmacology and antitumor activity of palmarumycin based inhibitors of thioredoxin reductase. Mol Cancer Ther 2006;5:630–6
- Xiong Z, Corey EJ. Simple total synthesis of the pentacyclic Cs-symmetric structure attributed to the squalenoid glabrescol and three Cs-symmetric diastereomers compel structural revision. J Am Chem Soc 2000;122:4831–2
- Prajoubklang A, Sirithunyalug B, Charoenchai P, et al. Bioactive deoxypreussomerins and dimeric naphthoquinones from Diospyros ehretioides fruits: deoxypreussomerins may not be plant metabolites but may be from fungal epiphytes or endophytes. Chem Biodivers 2005;2:1358–67
- Chu M, Truumees I, Patel MG, et al. Structure of Sch 49209: a novel antitumor agent from the fungus Nattrassia mangiferae. J Org Chem 1994;59:1222–3
- Singh SB, Zink DL, Liesch JM, et al. Preussomerins and deoxypreussomerins: novel inhibitors of ras farnesyl-protein transferase. J Org Chem 1994;59:6296–302
- Thiergardt R, Hug P, Rihs G, Peter HH. Cladospirone bisepoxide: definite structure assignment including absolute configuration and selective chemical transformations. Tetrahedron 1995;51:733–42
- Petersen F, Moerker T, Vanzanella FPHH. Production of cladosporine bisepoxide, a new fungal metabolite. J Antibiot 1994;47:1098–103
- Bode HB, Walker M, Zeeck A. Cladospirones B to I from Sphaeropsidales sp. F-24′707 by variation of culture conditions. Eur J Org Chem 2000;18:3185–93
- Krohn K, Beckmann K, Flörke U, et al. Biologically active metabolites from fungi, 9 new palmarumycins CP4a and CP5 from Coniothyrium palmarum: structure elucidation, crystal structure analysis and determination of the absolute configuration by CD calculations. Tetrahedron 1997;53:3101–10
- Ishizaki M, Ozaki K, Kanematsu A, et al. Synthetic approaches toward spiro[2,3-dihydro-4H-l-benzopyran-4,1′-cyclohexan]-2-oDnerivatives via radical reactions: total synthesis of (f)-lycoramine. J Org Chem 1993;58:3877–85
- Dai J, Krohn K, Elsässer B, et al. Metabolic products of the endophytic fungus Microsphaeropsis sp. from Larix decidua. Eur J Org Chem 2007;29:4845–54
- Bode HB, Zeeck A. Sphaerolone and dihydrosphaerolone, two bisnaphthyl-pigments from the fungus Sphaeropsidales sp. F-24'707. Phythochemistry 2000;54:597–601
- Ravindranath N, Reddy MR, Mahender G, et al. Deoxypreussomerins from Jatropha curcas: are they also plant metabolites? Phytochemistry 2004;65:2387–90
- Krohn K. Natural products derived from naphthalenoid precursors by oxidative dimerization. In: Herz W, Falk H, Kirby GW, et al., eds. Progress in the chemistry of organic natural products. Vol. 85. Wien (NY): Springer; 2003:1–49
- Akbaba Y, Bastem E, Topal F, et al. Synthesis and carbonic anhydrase inhibitory effects of novel sulfamides derived from 1-aminoindanes and anilines. Arch Pharm 2014;347:950–7
- Göksu S, Naderi A, Akbaba Y, et al. Carbonic anhydrase inhibitory properties of novel benzylsulfamides using molecular modeling and experimental studies. Bioorg Chem 2014;56:75–82
- Hisar O, Beydemir Ş, Gülçin İ, et al. Effect of low molecular weight plasma inhibitors of rainbow trout (Oncorhyncytes mykiss) on human erythrocytes carbonic anhydrase-II isozyme activity in vitro and rat erythrocytes in vivo. J Enzyme Inhib Med Chem 2005;20:35–9
- Yıldırım A, Atmaca U, Keskin A, et al. N-Acylsulfonamides strongly inhibit human carbonic anhydrase isoenzymes I and II. Bioorg Med Chem 2015;23:2598--605
- Boztaş M, Çetinkaya Y, Topal M, et al. Synthesis and carbonic anhydrase isoenzymes I, II, IX, and XII inhibitory effects of dimethoxy-bromophenol derivatives incorporating cyclopropane moieties. J Med Chem 2015;58:640–50
- Şentürk M, Gülçin İ, Daştan A, et al. Carbonic anhydrase inhibitors. Inhibition of human erythrocyte isozymes I and II with a series of antioxidant phenols. Bioorg Med Chem 2009;17:3207–11
- Arabaci B, Gülçin İ, Alwasel S. Capsaicin: a potent inhibitor of carbonic anhydrase isoenzymes. Molecules 2014;19:10103–14
- Supuran CT. Carbonic anhydrases – an overview. Curr Pharm Des 2008;14:603–14
- Innocenti A, Öztürk Sarıkaya SB, Gülçin İ, Supuran CT. Carbonic anhydrase inhibitors. Inhibition of mammalian isoforms I–XIV with a series of natural product polyphenols and phenolic acids. Bioorg Med Chem 2010;18:2159–64
- Şentürk M, Gülçin İ, Beydemir Ş, et al. In vitro inhibition of human carbonic anhydrase I and II isozymes with natural phenolic compounds. Chem Biol Drug Des 2011;77:494–9
- Supuran CT. Carbonic anhydrases: novel therapeutic applications for inhibitors and activators. Nat Rev Drug Discov 2008;7:168–81
- Supuran CT, Scozzafava A, Conway J. Carbonic anhydrase: its inhibitors and activators. Boca Raton (FL): CRC Press; 2004
- Öztürk Sarıkaya SB, Topal F, Şentürk M, et al. In vitro inhibition of α-carbonic anhydrase isozymes by some phenolic compounds. Bioorg Med Chem Lett 2011;21:4259–62
- Güney M, Coşkun A, Topal F, et al. Oxidation of cyanobenzocycloheptatrienes: synthesis, photooxygenation reaction and carbonic anhydrase isoenzymes inhibition properties of some new benzotropone derivatives. Bioorg Med Chem 2014;22:3537–43
- Innocenti A, Gülçin İ, Scozzafava A, Supuran CT. Carbonic anhydrase inhibitors. Antioxidant polyphenol natural products effectively inhibit mammalian isoforms I–XV. Bioorg Med Chem Lett 2010;20:5050–3
- Topal M, Gülçin İ. Rosmarinic acid: a potent carbonic anhydrase isoenzymes inhibitor. Turk J Chem 2014;38:894–902
- Göçer H, Akıncıoğlu A, Göksu S, et al. Carbonic anhydrase and acetylcholine esterase inhibitory effects of carbamates and sulfamoylcarbamates. J Enzyme Inhib Med Chem 2015;30:316–320
- Nar M, Çetinkaya Y, Gülçin İ, Menzek A. (3,4-Dihydroxyphenyl) (2,3,4-trihydroxyphenyl)methanone and its derivatives as carbonic anhydrase isoenzymes inhibitors. J Enzyme Inhib Med Chem 2013;28:402–6
- Scozzafava A, Mastrolorenzo A, Supuran CT. Modulation of carbonic anhydrase activity and its applications in therapy. Expert Opin Ther Patents 2004;14:667–702
- Pastorekova S, Parkkila S, Pastorek J, Supuran CT. Carbonic anhydrases: current state of the art, therapeutic applications and future prospects. J Enzyme Inhib Med Chem 2004;19:199–229
- Supuran CT, Scozzafava A, Casini A. Carbonic anhydrase inhibitors. Med Res Rev 2003;23:146–89
- Gülçin İ, Beydemir S. Phenolic compounds as antioxidants: carbonic anhydrase isoenzymes inhibitors. Mini Rev Med Chem 2013;13:408–30
- Alterio V, Fiore DA, D’Ambrosio K, et al. Multiple binding modes of ınhibitors to carbonic anhydrases: how to design specific drugs targeting 15 different isoforms? J Am Chem Soc 2012;112:4421–68
- Aslan A, Altun S, Ahmed I, et al. Synthesis of biologically active nonnatural palmarumycin derivatives. Eur J Org Chem 2011;6:1176–88
- Gülçin İ, Beydemir Ş, Büyükokuroğlu ME. In vitro and in vivo effects of dantrolene on carbonic anhydrase enzyme activities. Biol Pharm Bull 2004;27:613–16
- Aras Hisar S, Hisar O, Beydemir S, et al. Effect of vitamin E on carbonic anhydrase enzyme activity in rainbow trout (Oncorhynchus mykiss) erythrocytes in vitro and in vivo. Acta Vet Hung 2004;52:413–22
- Beydemir S, Gülcin I. Effect of melatonin on carbonic anhydrase from human erythrocyte in vitro and from rat erythrocyte in vivo. J Enzyme Inhib Med Chem 2004;19:193–7
- Hisar O, Beydemir Ş, Gülçin İ, et al. The effect of melatonin hormone on carbonic anhydrase enzyme activity in rainbow trout (Oncorhynchus mykiss) erythrocytes in vitro and in vivo. Turk J Vet Anim Sci 2005;29:841–5
- Çoban TA, Beydemir S, Gülçin I, Ekinci D. Morphine inhibits erythrocyte carbonic anhydrase in vitro and in vivo. Biol Pharm Bull 2007;30:2257–61
- Çoban TA, Beydemir S, Gülçin I, Ekinci D. The inhibitory effect of ethanol on carbonic anhydrase isoenzymes: in vivo and in vitro studies. J Enzyme Inhib Med Chem 2008;23:266–70
- Gülçin İ, Beydemir Ş, Çoban TA, Ekinci D. The inhibitory effect of dantrolene sodium and propofol on 6-phosphogluconate dehydrogenase from rat erythrocyte. Fresen Environ Bull 2008;17:1283–7
- Öztürk Sarikaya SB, Gülçin İ, Supuran CT. Carbonic anhydrase inhibitors: inhibition of human erythrocyte isozymes I and II with a series of phenolic acids. Chem Biol Drug Des 2010;75:515–20
- Şentürk M, Gülçin İ, Daştan A, et al. Carbonic anhydrase inhibitors: inhibition of human erythrocyte isozymes I and II with a series of antioxidant phenols. Bioorg Med Chem 2009;17:3207–11
- Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein dye binding. Anal Biochem 1976;72:248–51
- Coban TA, Beydemir S, Gülçin İ, et al. Sildenafil is a strong activator of mammalian carbonic anhydrase isoforms I–XIV. Bioorg Med Chem 2009;17:5791–5
- Çetinkaya Y, Göçer H, Gülçin İ, Menzek A. Synthesis and carbonic anhydrase isoenzymes inhibitory effects of brominated diphenylmethanone and its derivatives. Arch Pharm 2014;347:354–9
- Gülçin İ, Küfrevioğlu Öİ, Oktay M. Purification and characterization of polyphenol oxidase from nettle (Urtica dioica L.) and inhibition effects of some chemicals on the enzyme activity. J Enzyme Inhib Med Chem 2005;20:297–302
- Laemmli DK. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970;227:6805
- Köksal E, Gülçin İ. Purification and characterization of peroxidase from cauliflower (Brassica oleracea L.) buds. Protein Peptide Lett 2008;15:320–6
- Şentürk M, Gülçin İ, Çiftci M, Küfrevioğlu Öİ. Dantrolene inhibits human erythrocyte glutathione reductase. Biol Pharm Bull 2008;31:2036–9
- Şişecioğlu M, Çankaya M, Gülçin İ, Özdemir H. The inhibitory effect of propofol on lactoperoxidase. Protein Peptide Lett 2009;16:46–9
- Şişecioğlu M, Çankaya M, Gülçin İ, Özdemir H. Interactions of melatonin and serotonin with lactoperoxidase enzyme. J Enzyme Inhib Med Chem 2010;25:779–83
- Taslimi P, Gulcin İ, Ozgeris B, et al. The human carbonic anhydrase isoenzymes I and II (hCA I and II) inhibition effects of trimethoxyindane derivatives. J Enzyme Inhib Med Chem 2015. doi:10.3109/14756366.2015.1014476
- Göçer H, Gülçin İ. Caffeic acid phenethyl ester (CAPE): a potent carbonic anhydrase isoenzymes inhibitor. Int J Acad Res 2013;5:150–5
- Verpoorte JA, Mehta S, Edsall JT. Esterase activities of human carbonic anhydrases B and C. J Biol Chem 1967;242:4221–9
- Çetinkaya Y, Göçer H, Göksu S, Gülçin İ. Synthesis and carbonic anhydrase isoenzymes inhibitory effects of novel benzylamine derivatives. J Enzyme Inhib Med Chem 2014;29:168–74
- Akıncıoğlu A, Akbaba Y, Göçer H, et al. Novel sulfamides as potential carbonic anhydrase isoenzymes inhibitors. Bioorg Med Chem 2013;21:1379–85
- Akbaba Y, Akıncıoğlu A, Göçer H, et al. Carbonic anhydrase inhibitory properties of novel sulfonamide derivatives of aminoindanes and aminotetralins. J Enzyme Inhib Med Chem 2014;29:35–42
- Akıncıoğlu A, Topal M, Gülçin İ, Göksu S. Novel sulfamides and sulfonamides incorporating tetralin scaffold as carbonic anhydrase and acetylcholine esterase inhibitors. Arch Pharm 2014;347:68–76
- Gocer H, Topal F, Topal M, et al. Acetylcholinesterase and carbonic anhydrase isoenzymes I and II from human red blood cell inhibition profiles of taxifolin. J Enzyme Inhib Med Chem 2015. doi.org/10.3109/14756366.2015.1036051
- Scozzafava A, Kalın P, Supuran CT, et al. The impact of hydroquinone on acetylcholine esterase and certain human carbonic anhydrase isoenzymes (hCA I, II, IX, and XII). J Enzyme Inhib Med Chem 2015. doi:10.3109/14756366.2014.999236
- Aksu K, Nar M, Tanç M, et al. The synthesis of sulfamide analogues of dopamine related compounds and their carbonic anhydrase inhibitory properties. Bioorg Med Chem 2013;21:2925–31
- Krohn, K, Wang S, Ahmed I, et al. Flexible route to palmarumycin CP1 and CP2 and CJ-12.371 methyl ether. Eur J Org Chem 2010;23:4476–81