Abstract
The α-amylase was extracted from pure persimmon honey and purified by DEAE-Toyopearl 650M, CM-Toyopearl 650M, and Toyopearl HW-55F column chromatographies. Molecular weight of purified enzyme was estimated to be about 58 kDa by Toyopearl HW-55F gel chromatography and SDS-PAGE, respectively suggested that the purified enzyme was a monomer. Optimum pH of the enzyme was 6.0−7.0 and optimum temperature 40°C. The enzyme was extremely inactivated at pH was higher than 7.0 or lower than 5.0. Heat inactivation occurred at 40°C. This enzyme activated by Ca 2+ , Mn2+, PCMB, and DTNB, but inhibited by Ba2+, Fe3+, Hg2+, Mg2+, and iodoacetic acid. The purified enzyme was of α -type by TLC analysis. The relative rate of hydrolysis of the polymeric substance decreased with decreasing percentage of α -1,4-linkages and with increasing percentage of α -1,6-linkages in substrate similar to the results from commercially available honey.
Keywords:
INTRODUCTION
Honey market currently shows a tendency to establish geographical limits of production with the aim of protecting a production zone that has developed and marketed a particular standard of quality. Honey composition is closely associated with its botanical origin and, to some extent the geographical area in which it originated, because soil and climate characteristics determine melliferous floral.[Citation1] Honey is a natural complex product produced by honeybees from the nectar of blossoms or from exudates of trees and plants to produce nectar honeys or honeydews, respectively. Honey composition depends on the plants visited by the honeybees and on the climatic and environmental conditions. The strong sweetening capacity of honey is due to the presence of the monosaccharide's fructose and glucose as majority components about 60 to 85% and phenolic compounds, minerals, proteins, free amino acids, enzymes such as amylase (diastase), and vitamins as minor components.[Citation2,Citation3] Amino acids in honeys are attributable to the honeybees or to plant sources.
It has been known the presence of enzymes in honeys for many years. In particular, α-amylase is one of most important enzyme in honey species, although it contains a very small quantity of enzymes. The origin of α-amylase in honeys is commonly attributed to the honeybee. The nectar collected is mixed with secretions from the salivary and hypopharyngeal gland of foraging honeybees. α-Amylase was estimated to account for about only 2% of the total protein in the hypopharyngeal gland.[Citation4] This enzyme is largely used in Europe as a measure of honey freshness, because this activity decreases in old or heated honeys.
On the other hand, α-amylase is used in starch liquelaction to produce glucose, fructose, and maltose and in brewing, baking, textile, paper, detergent, and sugar industries.[Citation5] There are few report about α-amylase in honey species,[Citation6,Citation7] although this enzyme is beneficial to industrial use. The primary goal of this investigation was to purify and characterize amylase from persimmon honey, and to compare with the characteristics of amylase from other species.
MATERIALS AND METHODS
Materials
Fresh pure honey from persimmon was obtained from Inoue Yohojo Bee Farm Inc. (Hyogo, Japan) and used in this study. DEAE-Toyopearl 650M, CM-Toyopearl 650M, and Toyopearl HW-55F were purchased from Tosoh Co. (Tokyo, Japan). Marker proteins for gel chromatography were obtained from Boehringer Mannheim Co. (Tokyo, Japan). Marker proteins for electrophoresis and starch from rice were from Sigma Chemical Co. (St. Louis, MO) and from Amersham Biosciences UK Ltd. (UK). Coomassie brilliant blue R-250 was from Fluka Fine Chemicals Co., Ltd. (Tokyo, Japan). Pure α-amylase from Bacillus subtilis (20 U/mg), pure β-amylase from barley (32 U/mg), starch (corn, wheat, potato, and sweet potato), and bovine serum albumin (BSA) were purchased from Wako Chemicals Co., Ltd. (Osaka, Japan). Soluble starch was from Nacalai tesque Inc. (Kyoto, Japan). Neo. amylase test was purchased from Daiichi pure chemicals Co., Ltd. (Tokyo, Japan). Other chemicals were of analytical grade.
Measurement of α-Amylase Activity
α-Amylase activity was measured using blue starch as substrate. A 0.02 ml of honey sample solution was mixed with 0.784 ml of substrate solution [a tablet per ml of 0.1 M sodium phosphate buffer (pH 6.0)], and then incubated at 37°C for 30 min. After the reaction was stopped by the addition of 0.196 ml of 0.5 M NaOH, the suspension was centrifuged at 12,000 rpm for 5 min. The supernatants were measured by reading the absorbance at 620 nm. One enzyme unit was defined as the activity (IU/L): one unit (IU/L) = 1.85 × (Somogyi unit/dl). Distilled water was used as negative control.
Protein Determination
Protein concentration was measured as the method of Lowry et al.[Citation8] using BSA as standard protein.
Gel Filtration and Electrophoresis for Determination of Molecular Weight
Molecular weight of the purified enzyme was estimated using Toyopearl HW-55F (1.5 × 75 cm) gel filtration. Ferritin (450 kDa), catalase (240 kDa), aldolase (158 kDa), and albumin (68 kDa) were used as the standard marker proteins for gel filtration. Sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) was performed by the method of Laemmli[Citation9] using 10% gel. Myosin (205 kDa), β-galactosidase (116 kDa), phosphorylase (97.4 kDa), bovine serum albumin (66 kDa), ovalbumin (45 kDa), and carbonic anhydrase (29 kDa) were used as standard proteins. After electrophoresis, the gel was stained with Coomassie brilliant blue R-250 and destained with 25% ethanol and 7.5% acetic acid.
Thin-layer Chromatography (TLC) Analysis of Enzymatic Degradation Products
TLC was performed to investigate the reaction products of the purified amylase. Ten microliters of enzyme solution (pure α-amylase from Bacillus subtilis: 0.2 U/ml; pure β-amylase from barley: 0.04 U/ml; purified amylase from persimmon honey: 0.3 IU/ml) was incubated with 17 μl of 2% unmodified starch (from rice and sweet potato), 21 μl of 0.2 M sodium acetate buffer (pH 5.0), and 20 μl of 0.5 M NaCl. After at 40°C for 1 h, a 5 μl aliquot of each reaction mixture was loaded on a precoated K6F silica gel 60Å plate (size: 5 × 10 cm; layer thickness: 250 μm: Whatman International Ltd Maidstone England, UK). At the same time, 2% maltose and 1% unmodified starch (from rice and sweet potato) were included as controls. The hydrolysates were developed with butanol, ethanol, and water (5:3:2). The plates were sprayed with sulfuric acid-methanol (1:3) solution and then heated it in an oven at 100°C for 30 min to detect the distribution of sugars.
RESULTS AND DISCUSSION
Purification and Characterization of α-Amylase from Persimmon Honey
Pure honey from persimmon was dialyzed to remove a large amount of free sugars against distilled water at 4°C for 3 days by changing the water once a day. The dialysate was centrifuged at 50,000 × g for 1 h, and the supernatants were pooled and then freeze-dried. The dried sample (400 mg) was dissolved in 40 ml of 10 mM sodium phosphate buffer (pH 7.0). The enzyme solution was applied to a DEAE-Toyopearl 650M column (1.0 × 5.0 cm) previously equilibrated with the same buffer. The enzyme was not absorbed in the column. Next, the active fractions were collected, and were applied to a CM-Toyopearl 650M column (1.0 × 5.0 cm) previously equilibrated with the same buffer. Unfortunately, the enzyme was also not absorbed in this column. The active fractions were collected and concentrated to 0.5 ml using ultrafree-MC (10,000 NMWL Filter Unit: Millipore Co., USA). The concentrated enzyme solution was loaded on a Toyopearl HW-55F column (1.5 × 70 cm) which has been equilibrated with the same buffer, and the enzyme was eluted with the same buffer. Fractions of 3 ml were collected at a flow rate of 0.3 ml/min. This solution was separated into two protein fractions. The purified enzyme was stored at −85°C until used in the following experiments. The purification procedures are summarized in . Finally, α-amylase was purified up to 5.4-fold with a recovery rate of 46.8% as compared with crude extract.
Table 1 Summary of purification of α-amylase from persimmon honey
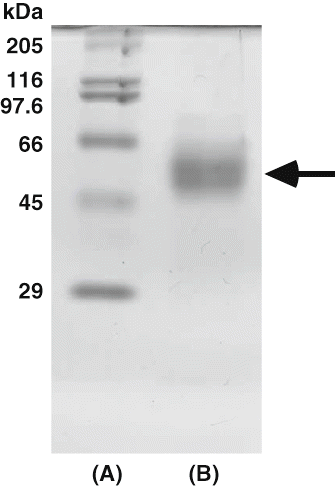
The molecular weight of the purified enzyme was estimated about 58 kDa by Toyopearl HW-55F (1.5 × 75 cm) gel filtration (data not shown). According to SDS-PAGE, the purified enzyme gave a single protein band of molecular weight about 58 kDa (). Babacan and Rand[Citation6] purified amylase from commercially available honey using ultrafiltration with a Hollow Fiber Cartridge Adapter System Model DH4 (Amicon Co., USA), ultracentrifugation, gel filtration, and ion-exchange chromatography. As a result, the amylase was purified up to 531.3-fold with a recovery rate of 3.9% in comparison to crude enzyme. The partial purified enzyme showed three protein bands by SDS-PAGE using 10% gel: the major band with 57 kDa and faint traces of two smaller component bands. Noman and others[Citation10] purified α-amylase from post-harvest Pachyrhizus erosus L. tuber by DEAE- and CM-Cellulose chromatographies. This enzyme was not absorbed in DEAE-Cellulose column, but was absorbed in CM-Cellulose column. These purification procedures yielded a pure amylase with 22.8% and the purity of the enzyme was 110-fold to crude enzyme. The molecular weight of the enzyme was estimated to 40 kDa. Tsvetkov and Emanuilova[Citation11] reported the purification and characterization of heat stable extracellular α-amylase from Bacillus brevis. Purification achieved was 8.1-fold from the crude extract with a yield of 30%. This had a molecular weight of about 58 kDa. Moreover, Ivanova and others[Citation12] were purified extracellular α-amylase having the molecular weight of 58 kDa produced by the Bacillus licheniformis. These values about molecular weight were similar to that from persimmon honey. On the other hand, Paquet and others[Citation13] purified the extracellular α-amylase from Clostridium acetobutylicum ATCC 824 using Mono Q ion-exchange chromatography and Superose 12 gel chromatography. Purification was 22.2-fold from crude extract with a yield of 23.2%. The purified enzyme showed the molecular weight about 84 kDa using SDS-PAGE. This value was similar to that of the extracellular enzyme from Streptococcus bovis JB1.[Citation14]
Figure 1 SDS-polyacrylamide gel electrophoresis of amylase from persimmon honey. (A) High molecular marker. Myosin (205 kDa), β-galactosidase (116 kDa), phosphorylase (97.4 kDa), bovine serum albumin (66 kDa), ovalbumin (45 kDa), and carbonic anhydrase (29 kDa) were used as standards; (B) Purified enzyme.
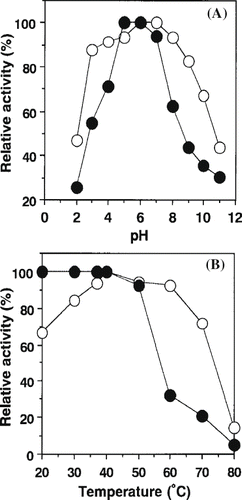
The influence of pH on enzymatic activity and stability is shown in A. The α-amylase from persimmon honey showed optimal activity from 6.0 to 7.0. This value was generally similar to those of B. brevis (pH 5.0 to 9.0),[11] C. acetobutylicum ATCC 824 (pH 5.6),[Citation13] S. bovis JB1 (pH 5.0 to 6.0),[14] and B. licheniformis (pH 6.0 to 6.5).[Citation12] The enzyme was most stable at pH 5.0 to 6.0, however it suddenly unstable when the pH was lower than 5.0 or higher than 7.0. This tendency was similar to that of S. bovis JB1 (pH 5.5 to 8.5),[Citation14] but was fairly different from those of C. acetobutylicum ATCC 824 (pH 3.0 to 5.5)[Citation13] and B. licheniformis (pH 6.5 to 8.0).[Citation12] The effect of temperature on enzymatic activity and stability is shown in B. The enzyme had a temperature optimum at 40°C and a stability below 40°C. Babacan and Rand[Citation7] reported that amylase from commercially available honey was optimal at pH 4.6 to 5.3 and at 55°C. The pH stability of this enzyme ranged from 7.0–8.0. These results indicate that optimal pH of amylase from persimmon honey was similar to that from commercially available honey, but optimal temperature was fairly different. Noman and others[Citation10] reported that α-amylase from P. erosus L. tuber exhibited optimum activity at pH 7.3 and at 37°C. This enzyme was stable between pH 6.0 and 8.0 for 24 h incubation at 4°C. The enzymatic activity decreased gradually at acidic pH or alkaline pH.
Figure 2 Effects of pH and temperature on activity and stability of amylase from persimmon honey. (A) For each pH, activity was measured as described under “Materials and Methods” and expressed as relative activity (○). The pH stability curve (•) represents the residual activity after a preincubation period of 1 day at the indicated pH. (B) Enzymatic activity was measured for each temperature (○). Thermal stability of the enzyme was incubated at different temperatures at pH 6.0 for 60 min. After cooling, the residual activity was measured (•).
Effects of metal ions and chemicals such as SH-blocking reagents at a final concentration of 1.0 mM each on the purified enzyme were determined under the standard assay and the results were indicated as relative activity. As a result, Mn2+ and PCMB and DTNB one of the sulfhydryl reagents strongly activated the enzymatic activity (). The enzyme was slightly activated by Na+, Ca2+, and Co2+. On the other hand, Ba2+, Cu2+, Zn2+, and glutathione had a slight inhibitory effects (about 10-57%) on the enzymatic activity. In particular, the enzyme strongly inhibited by Fe 3+ , Hg2+, CH2ICOOH. Among these chemicals Mg2+ completely inhibited the enzyme activity at a final concentration of 1.0 mM. Babacan and Rand[Citation6] investigated the effects of various metal ions on amylase from commercially available honey. The enzyme was inhibited by Cu+, Mg2+, and Hg2+, while Ca2+, Mn2+, and Zn2+ did not affect in this enzymatic activity. Noman and others[Citation10] measured enzymatic activity of amylase from P. erosus L. tuber against inhibition by different concentration of metal ions and chelating reagent. Most of ions, except for Na+, Ca2+, Fe2+, and Mn2+, strongly inhibited the enzymatic activity. The enzymatic activity was completely inhibited by addition of 100 mM EDTA. It was suggested that this amylase required metal ions for revelation of the activity. On the other hand, Ca2+ at a final concentration of 100 mM drastically enhanced the activity, suggesting the requirement of calcium for appearance of the activity and stability of the enzyme. From these results indicated that the inhibitory effects of other divalent cations might be due to the competition for calcium binding site while monovalent cations and Mg2+ might be poor competitors for calcium binding. Savchenko and others[Citation15] reported that calcium is required for α-amylase secretion and synthesis in Pyrococcus furiosus. Ranwala and Miller[Citation16] reported that Hg2+ and Ag+ at 2.0 mM completely inhibited the enzyme. Shaw and Ou-Lee[Citation17] reported α-amylase strongly inhibited by Cu2+ and moderately by Li+. It was reported that α-amylase from B. brevis was inhibited by Mg2+, Co2+, Cu2+, Ag+, and PMSF.[Citation12] Paquet and others[Citation13] reported that α-amylase from C. acetobutylicum ATCC 824 exhibited a slightly inhibition by Fe3+, Fe2+, Co2+, Mn2+, Zn2+, and Pb2+ at a final concentration of 1 mM (less than 35%) and totally inhibited by sulfhydryl oxidant metals such as Cu2+, Ag+, and Hg2+. Freer[Citation14] reported that α-amylase from S. bovis JB1 inhibited by Mn2+ (final concentration of 1.0 mM) and EDTA (10 mM) about 19 and 11%, respectively. This enzyme also moderately inhibited by Hg2+ about 19% and by PCMB about 88% at a final concentration of 1.0 mM each. In the same year, Ivanova and others[Citation12] reported that α-amylase from B. licheniformis was strongly inhibited by N-bromosuccinimide and EDTA, and the stability against temperature depended on the existence of Ca2+.
Table 2 Effects of metal ions and chemicals on persimmon honey α-amylase activity
Kinetic parameters of purified amylase from persimmon honey for starch as substrate were measured at optimal conditions (pH 6.0 and 40°C). This enzyme showed Michaelis-type kinetics when hydrolyzing starch. As calculated from Lineweaver-Burk plots, apparent Km and Vmax values were 2.81 mg and 0.532 (IU/L), respectively. There are many literatures about Km values of α-amylase as follows: 0.72 mg/ml (commercially available honey),[Citation7] 0.8 mg/ml (yeast Lipomyces kononenkoae),[Citation18] 0.9 mg/ml (B. licheniformis),[Citation12] 0.88 mg/ml (S. bovis),[Citation14] 3.6 mg/ml (C. acetobutylicum ATCC 824),[Citation13] 0.8 mg/ml (B. brevis 174),[Citation11] respectively. Noman and others[Citation10] reported Km and Vmax values were 0.29% and 0.37 μM/min/mg protein, respectively, for starch as substrate. Moreover, Abe and others[Citation19] also reported the same value in two allozymes from S. oryzae, however, Baker[Citation20] reported about double one in amylase extract from Rhyzopertha dominica.
Using several raw starches such as potato, sweet potato, wheat, and rice, soluble starch, polysaccharides such as amylose and amylopectin, substrate specificity of persimmon honey were investigated. As a result, amylose, high molecular weight substrate containing a large amount of α-1,4-linkage, was good substrate for this enzyme in comparison to soluble starch (). With increasing percentage of α-1,6-linkages and decreasing percentage of α-1,4-linkages in the substrate, the rate of hydrolysis of polymeric substrate decreased. Among these starches tested, starches from potato and sweet potato were good substrates, however the rate of hydrolysis did not reach to that of soluble starch. The relative activities of starches were as follows in the order: potato > sweet potato > wheat > rice. These tendencies of hydrolytic rate were similar to those of α-amylase from P. erosus L. tuber.[Citation10] Paquet and others[Citation13] studied the substrate specificity of α-amylase from C. acetobutylicum ATCC 824. As a result, amylose and amylopectin were the best substrates. The enzymatic activity increased from maltotriose to maltoheptaose, suggesting that the activity was higher in high molecular weight substrates than in low molecular weight. Freer[Citation14] investigated the substrate specificities of α-amylase from S. bovis JB1 using starches from potato and corn and polysaccharides such as amylose and amylopectin. The relative activities of this enzyme were higher in the following order: soluble corn starch > soluble potato starch = potato amylose (type III) > potato amylopectin > corn amylopectin.
Table 3 Substrate specificity of α-amylase from persimmon honey
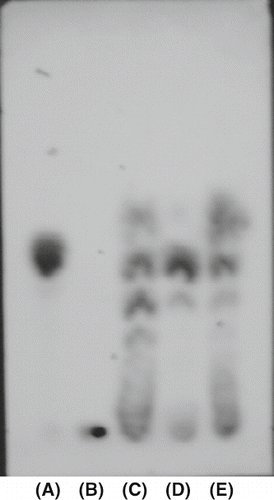
TLC analysis was performed to easily detect the enzymatic reaction products of purified amylase from persimmon honey and to identify type α- or β-amylase. α-Amylase from B. subtilis and α-amylase from Barley were used for comparison. As a result, α-amylase from Barley produced only maltose from rice starch (D). On the other hand, α-amylase from B. subtilis produced a large number of intermediate products (C). Purified amylase from persimmon honey not only produced maltose, but other intermediate products: this result also bore a striking resemblance to that from α-amylase from B. subtilis. In other words, purified enzyme from persimmon honey was an type α-amylase. On the other hand, corn starch was also degradated in the same manner. The results were shown in that was similar to those in rice starch. It suggests purified amylase from persimmon honey split the interior α-1,4-glycosidic bonds in a random manner.
Figure 3 Thin-layer chromatogram of the enzymatic degradated products of rice starch by pure α-amylase, β-amylase, and purified amylase from persimmon honey after incubation at pH 5.0 and at 40°C for 1 h. The reaction mixture was developted by butanol, ethanol, and water (5:3:2) and the sugars were detected by sulfuric acid : methanol (1:3) solution and heating at 100°C for 30 min. (A) maltose; (B) rice starch; (C) pure α-amylase; (D) pure β-amylase; (E) purified amylase from perssimon honey.
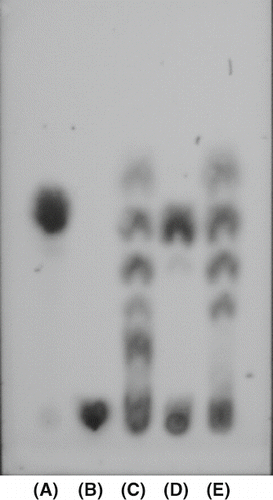
Figure 4 Thin-layer chromatogram of the enzymatic degradated products of corn starch by pure α-amylase, β-amylase, and purified amylase from persimmon honey. See sample nomenclature in except for (B) corn starch.
CONCLUSION
The α-amylase from pure persimmon honey was purified by some chromatographies and then was characterized. This enzyme was an type α-amylase that split the interior α-1,4-glycosidic bonds in a random manner.
Related Research Data
REFERENCES
- González Paramás , A.M. , Gómez Báres , J.A. , Garcia-Villanova , R.J. , Rivas Palá , T. , Ardunuy Albajar , R. and Sánchez , Sánchez, J. 2000 . Geographical discrimination of honeys by using mineral composition and common chemical quality parameters . J. Sci. Food Agric. , 80 : 157 – 165 .
- Ortiz Valbuena , A. and Fernandez Maeso , M.C. 1996 . “ Subra Muñoz dela Torre, E. Principales características de la miel de La Alcarria ” . In Investigación Agraria en Castilla la Mancha; Consejería de Agricultura y Medio Ambiente de Castilla la Mancha , 94 Spain : Castilla la Mancha .
- Conforti , P.A. , Lupano , C.E. , Malacalza , N.H. , Arias , V. and Castells , C.B. 2006 . Crystallization of honey at –20°C . Inter. J. Food Prop. , 9 : 99 – 107 .
- Ohashi , K. , Natori , S. and Kubo , T. 1999 . Expression of amylase and glucose oxidase in the hypopharyngeal gland with an age-dependent role change of the worker honeybee (Apis mellifera L.) . Eur. J. Biochem. , 265 : 127 – 133 .
- Crueger , W. and Crueger , A. 1990 . Biotechnology: a textbook of industrial microbiology , Edited by: Brock , T.D. 191 – 199 . Sunderland, IL, , USA : Sinauer Associate Inc .
- Babacan , S. and Rand , A.G. 2005 . Purification of amylase from honey . J. Food Sci. , 70 : C413 – 418 .
- Babacan , S. and Rand , A.G. 2007 . Characterization of honey amylase . J. Food Sci. , 72 : C50 – 55 .
- Lowry , O.H. , Rosebrough , N.J. , Farr , A.L. and Randall , R.J. 1951 . Protein measurement with the Folin phenol reagent . J. Biol. Chem. , 193 : 265 – 275 .
- Laemmli , U.K. 1970 . Cleavage of structural protein during the assembly of the head of bacteriophage T4 . Nature , 227 : 680 – 685 .
- Noman , A.S.M. , Hoque , M.A. , Sen , P.K. and Karim , M.R. 2006 . Purification some properties of α-amylase from post-harvest Pachyrhizus erosus L. tuber . Food Chemistry , 99, 444–449.
- Tsvetkov , V.T. and Emanuilova , E.I. 1989 . Purification and properties of heat stable α-amylase from Bacillus brevis . Appl. Microbiol. Biotechnol. , 31 : 246 – 248 .
- Ivanova , V.N. , Dobreva , E.P. and Emanuilova , E.I. 1993 . Purification and characterization of a thermostable alpha-amylase from Bacillus licheniformis . J. Biotechnol , 28 : 277 – 289 .
- Paquet , V. , Croux , C. , Goma , G. and Soucaille , P. 1991 . Purification and characterization of the extracellular α-amylase from Clostridium acetobutylicum ATCC 824 . Appl. Environ. Microbiol. , 57 : 212 – 218 .
- Freer , S.N. 1993 . Purification and characterization of the extracellular α-amylase from Streptococcus bovis JB1 . Appl. Environ. Microbiol. , 59 : 1398 – 1402 .
- Savchenko , A. , Vielle , C. , Kang , S. and Zeikus , G. 2002 . Pyrococcus furiosus α-amylase is stabilized by calcium and zinc . Biochemistry , 41 : 6193 – 6201 .
- Ranwala , A.P. and Miller , W.B. 2000 . Purification and characterization of an endoamylase from tulip (Tulipa gesneriana) bulbs . Physiologia Plantarum , 109 : 388
- Shaw , J.F. and Ou-Lee , T.M. 1984 . Simultaneous purification of α-amylase β-amylase from germinated rice seeds and some factors affecting activities of the purified enzyme . Bot. Bull. Acad. Sin. , 25 : 197 – 204 .
- Prieto , J.A. , Bort , B.R. , Martinez , J. , Randez-Gil , F. , Buesa , C. and Sanz , P. 1995 . Purification and characterization of new α-amylase of intermediate thermal stability from the yeast Lipomyces kononenkoae . Biochem. Cell. Biol. , 73 : 41 – 49 .
- Abe , R. , Chib , A.Y. and Nakajima , T. 2002 . Characterization of the functional module responsible for the low temperature optimum of rice α-amylase(Amy 3E) . Biologia Bratislava , 57 ( Suppl. 11 ) : 197 – 202 .
- Baker , J.E. 1991 . Purification and partial characterization of α-amylase allozymes from the lesser grain borer Rhyzopertha dominica . Insect Biochem. , 21 : 303 – 311 .