Abstract
Stress relaxation in two varieties of cooked potato tubers was studied using the sigmoid rate controlled model (RCM). The model parameters C 1, C 2 and K were rationalised in dimensionless form (C 1′ and C 2′) or as a ratio to the initial stress (K′). The activation volume and some other helpful parameters were calculated from the model parameters. The activation volume behaved similarly as activation volume obtained for raw tubers, but variety differences are very low. Variety differences were observed for the model parameter N:2.34 ± 0.42 for ‘Nicola’ and 3.36 ± 0.57 for ‘Panda’. Other variety differences were observed for i) ratio of activation volumes of the cooked and raw tubers and ii) ratio of f-values obtained at the same test parameters for the varieties to be compared.
INTRODUCTION
Texture of cooked potato is usually evaluated using sensory panels that give their sensory impressions of textural attributes Citation[1]. These are consistency, disintegration, humidity, mealiness, and structure Citation[2]. The consistency and mealiness are the most important quality elements of cooked potatoes, well representing more than 95 per cent of their variation Citation[3]. Since the 1950s several attempts have been made to replace the sensory panels by objective methods of measuring the cooked potato texture. One of the methods is based on careful analysis of short time stress relaxation in cooked potato tissue Citation[4].
An important role in the analysis plays the mathematical model that is used for a description of the process. The selected model parameters can serve as measures of the texture parameters and especially of the consistency. Davis and Leung Citation[4] used the generalised Maxwell function and the differences between potato texture that was observed for Russet Burbank potatoes of different density were expressed by different values of the model Young's moduli and viscosities. The generalised Maxwell model belongs to the special type of models that can be described by the following Eq. Equation5:
Among the lot of different mathematical functions that can be used as the source functions the special role is played by those that take part in so called rate controlled models (RCM) – Citation[5], e.g. logarithmic model Citation[5], Citation[7], with source f-function:
In previous paper Citation[6], the logarithmic RCM was modified. New model is termed sigmoid RCM and is given by the following source f-function:
In this paper, sigmoid RCM was used to study the stress relaxation in cooked tubers of two varieties with different texture. Special attention was paid to pre-stress part of real relaxation curves, where un-controllable stress relaxation processes take part.
MATERIALS AND METHODS
Experimental Procedure
The potatoes (varieties ‘Nicola’ and ‘Panda’) were supplied by the Potato Research Institute at Havlíčkův Brod, located in the east part of Bohemia. Traditional salad variety ‘Nicola’ has a lower starch content (16–17%) in comparison to ‘Panda’ (19–21%). The tubers were harvested in October 1998 and kept in cold store (at temperature 6°C and 95% relative humidity) until the end of January and then tested.
Compression tests were carried out on cylindrical specimens, 15 mm diameter and 23 mm long cut from the central parts of tubers by a cork borer in such a manner that the specimen axis was approximately parallel to the tuber axis. The specimens were vacuum packed in plastic bags (Fatranex, Fatra a.s., Czech Republic) and cooked in the water bath 20 minutes at the temperature (95 ± 0.2)°C. Then the specimens were removed and cooled rapidly in another water bath at temperature (25 ± 0.2)°C. All tests were performed in an Instron universal testing machine, type 4464.
Twenty-five specimens were used for the usual stress relaxation test in compression; 20 at pre-strain rate 0.167 mm s−1 (5 specimens per every different pre-strain level: 5%, 10%, 15%, and 20%), and 5 specimens at level pre-strain rate 0.8333 mm s−1 (pre-strain level 15%). The real relaxation time was 10 minutes. Deformation force was registered ten times per second and the obtained data, 6000 per every specimen, were used for numerical calculation. This number was reduced in some numerically difficult calculations up to 70 points. In this case, the force interval between the initial and final points was covered by the selected data equidistantly. All the data obtained for relaxation times lower than 0.3 second were excluded from the evaluation procedures.
Pre-strain parameters, e.g. pre-strain stress and strain, and also all the computed quantities, are used here in so-called “true” or Hencky's representation Citation[8], Citation[6]. For incompressible media the true stress is approximated by σ0t = σ0(1−ε0) with σ0 and ε0 are the engineering stress and strain, respectively. A similar simple equation ε0t = − ln (1 − ε0) holds for the true strain.
Model
The sigmoid RCM expressed by the sigmoid source f-function given by Eq. Equation3 was used for evaluation of the obtained stress relaxation curves.
Determination of the Model Parameters
The first time derivative of stress relaxation curve was numerically determined as the first derivative of the Lagrangian interpolation polynomial of degree 2 in 3 successive points. These values were related to the middle of the three neighbour points so that the derivative was determined in every point (the first and last points were excluded) of each stress relaxation curve. The model parameters were determined by minimisation of Chi-square function SFC (standard fitting coefficient), defined by the following equation
Minimisation was performed using the FORTRAN programme MINUIT Citation[9]. The minimisation process led to the model parameters that gave the best approximation of the experimental curves.
RESULTS AND DISCUSSION
Source f-Functions and Model Parameters
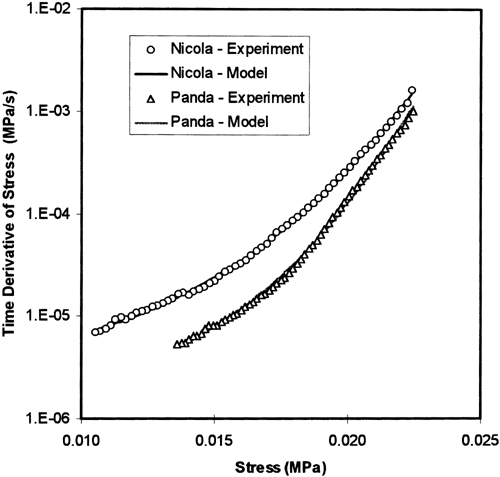
Typical source f-functions of our stress relaxation curves are given in Figure . The experimental data were obtained as negative values of the first time derivative of stress. The functions are approximated by the sigmoid source f-functions giving the best fits. The stress relaxation started at the initial stress σ0t displayed at the right side of the source f-functions plotted in Figure . During the test both the quantities plotted in Figure (the stress and negative value of time derivative of the stress) decreased continuously. During ten minutes the stress decreased up to approximately one half of the initial value, but in the same time the decrease of the negative time derivative of stress is represented by reduction about two orders. The experimental data well determine the part of the source f-function for stresses higher than 0.5 σ0t . This part of the source f-function is very useful for good estimation of the model parameters K, C 1, and N. Not so far for the parameter C 2, which strongly depends on the initial part of the source f-function, where no data were obtained in our experiments. The minimisation procedures with C 2 as a free parameter led to results with very high variation not only of C 2-values, but also of all the other model parameters. This is why we fixed parameter C 1 at the mean values of the preliminary results: 2.5/σ0t for all our experiments. The sigmoid f-function is characterised also by one point of inflection Citation[6], usually calculated for stresses well below the minimum values observed in our tests (σ it = 0.1–0.3 σ0t ).
Figure 1. Typical f-functions determined directly for both the varieties after 5 per cent pre-strain. The best model approximations of the data by the sigmoid source f-function are also plotted in the figure (SFC: 4.33% – ‘Nicola’, 7.58% – ‘Panda’).
Mean values of the model parameters that were obtained for different variations of the experiments are given in Table . The results were obtained with approximately the same level of precision (SFC ≈ 5–8% in most cases) in contrary to stress relaxation in raw potato tubers, where precision of approximation increased with increasing pre-strain Citation[6]. Table contains parameters C 1 and C 2 in the dimensionless normalised form (C 1′ = C 1σ0t N and C 2′ = C 2σ0t ), which did not vary with change of the test conditions.
Table 1. Mean Values of Model Parameters (Sigmoid Model), LSD – Least Significant Difference
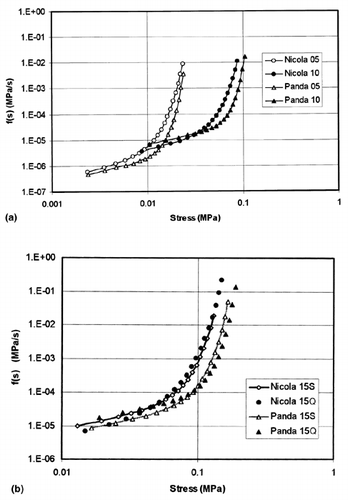
Mean values of the model parameters given in Table were used to calculate the model source f-functions – see Figure . Source f-functions of ‘Panda’ had lower f-values than the corresponding source f-functions of ‘Nicola’ in the initial parts of relaxation curves (i.e. at high stresses). This difference is eliminated at low stresses close to the end of relaxation test; in some cases, crossing of both the source f-functions for ‘Nicola’ and ‘Panda’ was observed. The source functions calculated for 15% pre-strain are only slightly sensitive to the pre-strain rate. For both the tested varieties, the source f-functions calculated for slow and quick pre-strain are very closed one to another (Figure b). The main difference between them consists in higher initial stress values observed at quicker pre-strains.
Figure 2. The model sigmoid source f-functions calculated using the mean values of the model parameters (Table ). Symbols at variety denote test details (the pre-strain level: 05 (5%), 10 (10%), 15 (15%), the pre-strain rate S (Slow −0.167 mm s−1), Q (Quick −0.833 mm s−1). a. f-functions for 5% and 10% pre-strain levels, b. f-functions for 5% and 10% pre-strain levels.
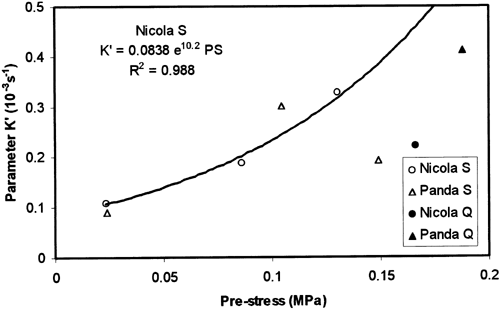
The parameter K′ equalled K/σ0t increased with increasing pre-stress level (Figure ) in contrary to the results obtained at raw potato tubers, where the totally opposite trend of the change was observed Citation[6]. It means that the initial slope of the source f-functions increased with increasing pre-stress more quickly than linearly.
Figure 3. Parameter K′ = K/σ0t plotted against the pre-stress σ0t . Symbols S and Q at variety title denote pre-strain rates 0.167 mm s−1 and 0.833 mm s−1, respectively.
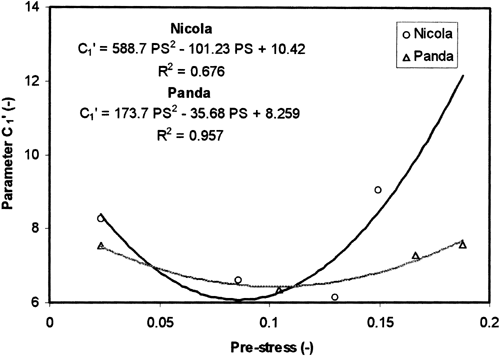
The parameter C 1′ depended on the pre-strain rate and pre-stress level. More information on these relations brings Figure . The observed dependence C 1′ on pre-stress level is generally non-monotonous, with minimum close to 0.1 MPa pre-stress value. The dependence is well described by polynomial of the second order. This dependence is more expressive for ‘Nicola’, while for ‘Panda’ only very flat curve was observed.
Figure 4. Parameter C 1′ = C 1σ0t n plotted against pre-stress. The experimental points were approximated by polynomial of the second order.
Significant differences between both the tested varieties were observed for exponent N (Table ). These differences are approximately the same for different pre-stress levels and different strain rates, which was used for pre-straining (Figure ).
Figure 5. Parameter N(MV ± SD) plotted against the pre-stress value.
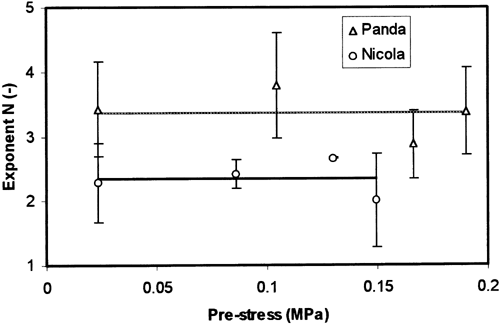
The other model parameters, significantly dependent on some test parameter (Table ), will not be discussed here. Their relations to the test parameters are either trivial or they are not experimentally well supported (e.g. position of the point of inflection located in such a part of source f-function where the experimental data were not obtained).
Activation Volume
Two parameters: activation energy and activation volume play very important role in every RCM Citation[10]. The relaxation rate is studied at different stress levels in our case and it makes possible to calculate activation volume using the following formula Citation5-6:
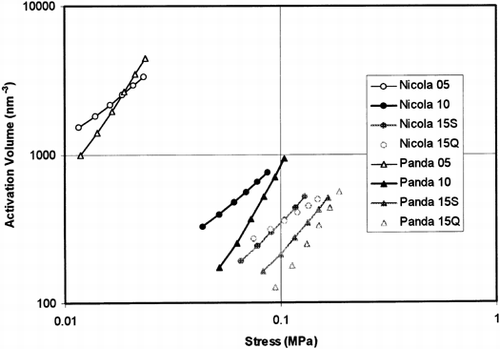
The activation volumes that were calculated from the model parameters under the exact model formulas (Eq. Equation5) are plotted against the actual stress in Figure . This figure shows that the activation volume decreased with increasing pre-strain and also decreased with decreasing stress value in a separate test. This is very close to the results that were obtained previously Citation[6] for raw potato tissue. It means that the RCM cannot be understood as a simple function of stress, but the source f-function has to contain some relation to the other parameters such as time, the pre-stress and pre-strain values, etc. The influence of pre-strain rate on the activation volume is very limited similarly as was observed for the raw potato tissue Citation[6]. Contrary to the previous experiments with raw potato tissue, the influence of variety on the activation volume is limited (Figure ).
Figure 6. Activation volume calculated using Eq. Equation5 for stresses 0.5σ0t , 0.6σ0t , 0.7σ0t , 0.8σ0t , 0.9σ0t , and σ0t is plotted against stress level for all combinations of the test parameters (two varieties, different pre-strain levels, T = 293 K). For symbols see Figure .
The differences between the activation volumes obtained for cooked and raw potato tubers can be expressed by ratio of these quantities that were obtained at the same test conditions. Figure contains this ratio calculated from the model parameters by Eq. Equation5 plotted against relative stress. The results can be well approximated by the power formula as is illustrated there for the results obtained at the lowest pre-stress. The power formula between the activation volume ratio and the relative stress can be also given by approximated formula – see Eq. Equation6. Table contains parameters of the power formula given by both the methods: i) regression analysis of the exact results plotted against relative stress, and ii) by approximated formula – Eq. Equation6: a = σ0tr C 1c ′ Nc /σ0tc C 1r ′ Nr and m = Nc −Nr . Figure shows that for the high degree of relaxation at relative stress about 0.5 and lower pre-strain values (less than about 10%) the activation volume ratio in Figure is much higher for ‘Nicola’ than for ‘Panda’.
Figure 7. Ratio of the activation volumes for cooked potato tubers and for raw potato tissue Equation6 calculated from the model parameters as determined at the same test parameters using Eq. Equation5. The calculated ratio is plotted against the relative stress (ratio of the actual stress to the pre-stress value). The values obtained at 5% pre-strain were approximated by power formula.
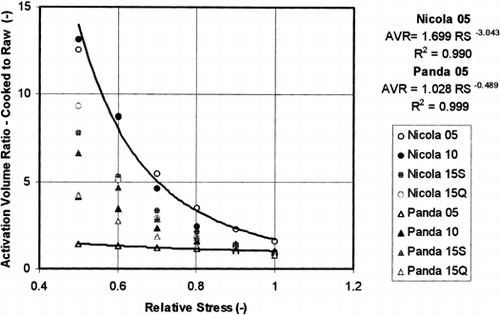
Table 2. Parameters of Power Relation Between Ratio of Activation Volumes (for Cooked and Raw Potato Tubers) and Relative Stress AVR = a.RSm (see Figure )
Variety Differences
The differences in model parameters that were observed for ‘Nicola’ and ‘Panda’ would be used as general measures of differences in cooked potato tubers of lower starch content (‘Nicola’) and of higher starch content (‘Panda’). The main differences between both the varieties consists in different shapes of the f(σ) curves, i.e., plots of the source f-function on stress (Figures . The f(σ) curves for ‘Panda’ were observed steeper and more curved than f(σ) curves for ‘Nicola’. This difference consists mainly in different values of the parameter N of the sigmoid RCM. The parameter N was observed significantly higher at ‘Panda’ than at ‘Nicola’ (Figure and Table ). This difference between both the varieties seems to be independent on pre-strain and pre-strain rate levels. This cannot be observed by Davis and Leung Citation[4], because they used short-time relaxation tests (10 seconds), that was evaluated using the generalised Maxwell model giving unreliable course of source f-function, that are too far from the observed sigmoid source f-functions at low stresses Citation[6].
Another difference between both the tested varieties was observed in the ratio of activation volumes (as determined for cooked and raw tubers) – see Figure – that were obtained at the same pre-strain parameters. The differences are most expressive at lower pre-strains and low relative stress values (close to the end of relaxation curve).
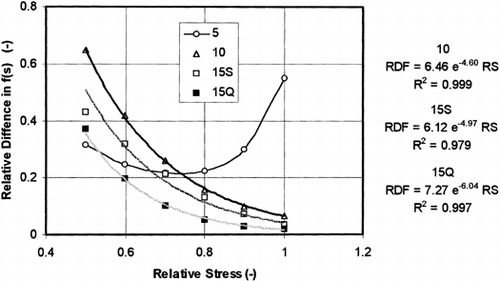
The variety difference of f (σ) shape can be expressed by ratio of f (σ) of ‘Panda’ and f(σ) of ‘Nicola’, both determined at the same stress value. The ratio based on our data is plotted in Figure against the relative stress observed in ‘Nicola’. The plot can be generally non-monotonous (e.g. 5% pre-strain), but for higher pre-strain values is nearly exponential. The higher the numerical coefficient in the exponent of the regression equation (values 4.80, 4.97, and 6.04 in Figure ), the bigger is the difference in texture of the varieties to be observed.
Figure 8. Ratio of the f (σ) values of ‘Panda’ and ‘Nicola’, as obtained at the same stress, plotted against the relative stress in ‘Nicola’. The data obtained at 10% and 15% pre-strains are approximated by exponential equations.
Relation to Texture Parameters
In Maxwellian body Citation[8], source f-function is in relation with fictive model viscosity η given by equation η = Eσ/f, where E is Young's modulus and f corresponding source f-function value. Most important differences between both the tested varieties were observed at higher relaxation times, i.e., at lower stresses. At this stage of relaxation the higher stresses and the lower f-function values were observed in ‘Panda’ in comparison to ‘Nicola’ (Figures and ). This behaviour can be understood as an indication of higher “viscosity” and/or “mealines” of cooked ‘Panda’ tubers. The mealiness correlates also with the “initial elasticity” and “fracturability to Young's modulus ratio”, the objective texture parameters defined in our previous paper Citation[11]. The exact relations between the stress relaxation parameters and both groups of texture parameters (objective and sensoric) have to be determined in future research, in which stress relaxation and texture of cooked potato tubers will be studied in parallel series.
CONCLUSIONS
Stress relaxation curves of cooked potato tubers can be well approximated by sigmoid rate controlled model with a source f-function of σ-shape termed here as the sigmoid source f-function. The model parameters C 1, C 2 and K can be rationalised in dimensionless form (C 1′ and C 2′) or as a ratio to the initial stress (K′). Activation volume behaves similarly as activation volume obtained in previous paper for the raw tubers, but the variety differences are lower in cooked tubers. The model parameter N does not depend significantly on parameters of pre-stress part of the test but there is variety dependent: values 2.34 ± 0.42 for ‘Nicola’ and 3.36 ± 0.57 for ‘Panda’ were obtained. The higher observed parameter N, the higher is curvature of the source f-function in semi-logarithmic scale. The variety difference can be also expressed by i) ratio of activation volumes of the cooked and raw tubers and ii) ratio of f-values obtained at the same test parameters for the varieties to be compared, especially at higher times of relaxation. The last possibility can probably serve as an objective indicator of tuber viscosity that is usually termed as “mealiness” in texture vocabulary.
ACKNOWLEDGMENTS
The authors thank Mr. J. Vacek, the Potato Research Institute at Havlíčkův Brod for experimental material.
REFERENCES
- 1977 . “ Anonymous ” . In Methods of Assessment for Potatoes and Potato Products Wageningen : EAPR Pudoc .
- Lugt , C. 1959 . Report on the Third International Meeting on Potato Quality Research held at Zürich-Oerlikon, Switzerland on February 5th, 6th and 7th, 1959 . Eur. Potato J. , 2 : 84 – 85 .
- Marle , J. T. , van de Vries , R. , Wilkinson , E. C. and Yuksel , D. 1997 . Sensory Evaluation of the Texture of Steam-cooked Table Potatoes . Potato Res. , 40 : 79 – 90 .
- Davis , D. C. and Leung , H. K. 1987 . Standardizing Measurement of Rheological Properties of Cooked Russet Burbank Potatoes . Trans. ASAE , 30 : 554 – 559 .
- Dotsenko , V. I. 1979 . Stress Relaxation in Crystals . Physica Status Solidi , 93 : 11 – 43 .
- Blahovec , J. Improved Rate Controlled Model for Stress Relaxation in Vegetable Tissue . Submitted to International Agrophysics ,
- Blahovec , J. 1996 . Stress Relaxation Phenomena in Vegetable Tissue Experimental Results . Journal of Materials Science , 31 : 1729 – 1734 .
- Mohsenin , N. N. 1970 . Physical Properties of Plant and Animal Materials Vol. 1 , New York : Gordon and Breach .
- James , F. and Roos , M. 1975 . Minuit – A System for Function Minimization and Analysis of the Parameter Errors and Correlations . Comp. Physics Comm. , 10 : 343 – 367 .
- Daniels , F. and Alberty , R. A. 1961 . Physical Chemistry New York : John Wiley & Sons .
- Blahovec , J. , Esmir , A. A.S. and Valentova , H. 2000 . Simplified Texture Profile Analysis of Cooked Potatoes . Int. J. Food Properties , 3 : 193 – 206 .