
Abstract
Polyphenols are an important class of phytochemicals, and several lines of evidence have demonstrated their beneficial effects in the context of a number of pathologies including neurodegenerative disorders such as Alzheimer’s and Parkinson’s disease. In this report, we review the studies on the effects of polyphenols on neuronal survival, growth, proliferation and differentiation, and the signaling pathways involved in these neurotrophic actions. Several polyphenols including flavonoids such as baicalein, daidzein, luteolin, and nobiletin as well as nonflavonoid polyphenols such as auraptene, carnosic acid, curcuminoids, and hydroxycinnamic acid derivatives including caffeic acid phentyl ester enhance neuronal survival and promote neurite outgrowth in vitro, a hallmark of neuronal differentiation. Assessment of underlying mechanisms, especially in PC12 neuronal-like cells, reveals that direct agonistic effect on tropomyosin receptor kinase (Trk) receptors, the main receptors of neurotrophic factors including nerve growth factor (NGF) and brain-derived neurotrophic factor (BDNF) explains the action of few polyphenols such as 7,8-dihydroxyflavone. However, several other polyphenolic compounds activate extracellular signal-regulated kinase (ERK) and phosphoinositide 3-kinase (PI3K)/Akt pathways. Increased expression of neurotrophic factors in vitro and in vivo is the mechanism of neurotrophic action of flavonoids such as scutellarin, daidzein, genistein, and fisetin, while compounds like apigenin and ferulic acid increase cyclic adenosine monophosphate response element-binding protein (CREB) phosphorylation. Finally, the antioxidant activity of polyphenols reflected in the activation of Nrf2 pathway and the consequent upregulation of detoxification enzymes such as heme oxygenase-1 as well as the contribution of these effects to the neurotrophic activity have also been discussed. In conclusion, a better understanding of the neurotrophic effects of polyphenols and the concomitant modulations of signaling pathways is useful for designing more effective agents for management of neurodegenerative diseases.
Introduction
Polyphenolic compounds
Polyphenols are an important group of phytochemicals that are abundantly present in food sources.Citation1 Several lines of evidence have demonstrated beneficial effects of these compounds in the context of several pathologies,Citation2–Citation8 and it has been repeatedly shown that consumption of foods rich in phenolic compounds can lower the risk of several diseases.Citation9,Citation10
Polyphenols have been reported to be of therapeutic value in neurodegenerative diseases,Citation11–Citation13 hypertensionCitation14 and other cardiovascular diseases,Citation15 cancer,Citation16–Citation18 inflammation,Citation19,Citation20 diabetes,Citation21 dyslipidemia,Citation22–Citation24 allergy, and immune system diseases.Citation25,Citation26 They have also been reported to have a role in the prevention of different types of cancer ranging from liver,Citation4 prostate,Citation27 and colorectalCitation28 cancer to lymphoblastic leukemia.Citation29
Neurodegenerative disorders such as Alzheimer’s, Parkinson’s, and Huntington’s disease represent rapidly growing causes of disability and death, which have profound economic and social implications; nonetheless, only few effective disease-modifying therapies are available for these diseases.Citation30–Citation32 Recent research has shown that certain polyphenols may have considerable neuroprotective effects in different brain pathologies including neurodegenerative diseases.Citation8,Citation11,Citation33,Citation34 Polyphenols from grape juice administered to older adult subjects have been shown to ameliorate the mild cognitive impairment.Citation35 Similarly, Witte et alCitation36 have shown that administration of resveratrol, a polyphenol present in wine, in older adults significantly improves memory performance, indicating potential strategies to maintain brain health during aging. Assessment of cognitive performance in middle-aged individuals has indicated that consumption of different polyphenols such as catechins, flavonols, and hydroxybenzoic acids is strongly associated with language and verbal memory.Citation37 Other studies have also shown similar effects of other polyphenols on cognitive function and memory in older individuals, who are at risk for neurodegenerative diseases.Citation38,Citation39 In another human study, epigallocatechin–gallate (EGCG) significantly improved cognitive deficits in individuals with Down syndrome.Citation40
Several studies have demonstrated that different polyphenolics with neuroprotective activity, such as EGCG,Citation41,Citation42 epicatechin,Citation43 curcumin,Citation44 resveratrol,Citation45 quercetin,Citation46 and citrus flavonoids (naringenin and hesperetin),Citation47 are able to cross the blood–brain barrier and therefore indicate that these compounds localize within the brain tissue and may well exert neuroprotective and neuromodulatory actions in the settings of different brain pathologies.
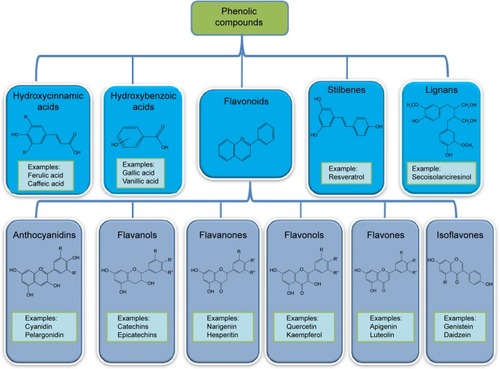
Polyphenolic compounds can be classified into different groups as described in .Citation1 Among them, flavonoids constitute a major subgroup, which are highly present in fruits and vegetables and possess several biological activities.Citation1
Figure 1 Main classes of polyphenols and their chemical structures.
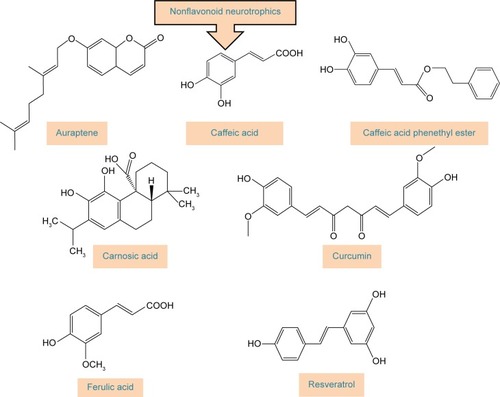
Polyphenols have been extensively studied for their antioxidant actionCitation48–Citation52 and some of their beneficial effects, including neuroprotective activity, have been attributed to this capacity.Citation53–Citation55 The neuroprotective effect of polyphenols against the diseases of nervous system has also been addressed in a number of studies.Citation56–Citation59 The aim of this review, however, is to extensively address the alterations of signaling pathways involved in the neurotrophic action of polyphenols that lead to neuronal survival, growth, proliferation, and differentiation. The other aspects of neuroprotective activity of these compounds, which are mainly ascribed to the antioxidant action and mitigation of oxidative stress, have been reviewed elsewhereCitation5,Citation7,Citation8,Citation60–Citation62 and are only briefly mentioned here. It should also be mentioned that in in vitro studies, which are the main focus of this report, supraphysiological doses of polyphenols may have been used and, therefore, caution should be exerted in extrapolation of these data to in vivo conditions.Citation56 The structures of some of the most highly studied neurotrophic polyphenols are depicted in and .
Figure 2 Chemical structures of flavonoids with neurotrophic activity.
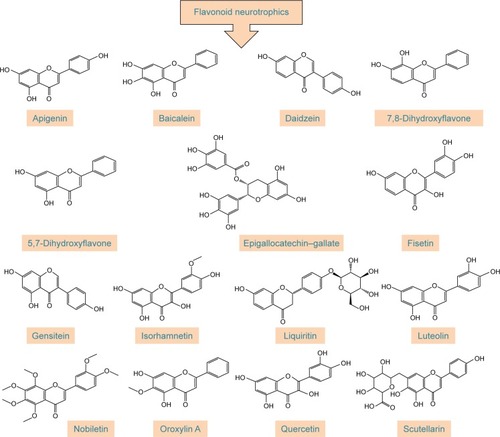
Figure 3 Chemical structures of polyphenols with neurotrophic activity that do not belong to the group of flavonoids.
Neurotrophic effects of polyphenols: Enhancement of survival in neuronal cells
Survival signaling is important to suppress the cell death machinery and counterbalance apoptotic signaling in the nervous system.Citation63 Several studies have shown that polyphenolic compounds enhance neuronal survival in serum-deprived conditions. Lin et alCitation64 showed that in neuronal-like PC12 cells, luteolin (3′,4′,5,7-tetrahydroxyflavone, ) attenuates serum withdrawal-induced cytotoxicity. Other examples of neuroprotectants that exerted prosurvival action in PC12 cells include curcuminoids, the predominant polyphenolic compounds of Curcuma longa Linn.;Citation65 EGCG (), the major polyphenol of green tea extract;Citation66 caffeic acid phenethyl ester (CAPE, ), an active component of propolis;Citation67 5-hydroxy-3,6,7,8,3′,4′-hexamethoxyflavone (HHMF) from the Citrus genus; and nobiletin (), the most abundant polymethoxyflavone in orange peel extract.Citation68 Furthermore, other investigators have shown that treatment with genistein and daidzein (), isoflavones present in soybeans and soy products, enhances proliferation and survival of the hippocampal neuronal cells.Citation69 In addition, application of 7,8-dihydroxyflavone (7,8-DHF, ), a flavone, promotes survival of cultured motoneurons, spiral ganglion neurons (SGNs), and hippocampal neuronal cells.Citation70 In another study, the protective ability of quercetin (3,3′,4′,5,7-pentahydroxyflavone, ) on P19-derived neurons was determined.Citation71 Ferulic acid (4-hydroxy-3-methoxycinnamic acid, ) was able to significantly increase the survival of neural stem/progenitor cells (NSC/NPCs) cultured from rat embryo, and also increased the number and size of secondary formed neurospheres.Citation72
Promotion of neurite outgrowth in neuronal cells
Neurite outgrowth is a crucial step in the differentiation of neurons, which begins at the cell body and extends outward to form functional synapses. In response to extracellular signals, the growth of neurite processes starts, involving the addition of new plasma membranes, generation of new cytoplasm, and the continued expansion and modification of the cytoskeleton.Citation73 A number of neurotypic proteins have been associated with neurite outgrowth, including growth-associated protein 43 (GAP-43), microtubule-associated protein (MAP) and tau, as well as presynaptic membrane-associated proteins, such as synaptophysin and synapsin.Citation74 Many polyphenolic compounds from natural and synthetic sources have been demonstrated to induce neurite outgrowth in various primary neuronal cultures and neuronal cell lines ().
Table 1 Induction of neurite outgrowth by polyphenols and involvement of neurotrophic signaling pathways
Several studies have shown that different polyphenols including flavonoids such as genistein,Citation75 quercetin, liquiritin from Glycyrrhizae radix plant (),Citation76 isorhamnetin (a flavonolaglycone from Ginkgo biloba plant, ),Citation77 and acetylated flavonoid glycosides from ScopariadulcisCitation78 as well as the stilbenoid compound resveratrol (a polyphenol present in grapes and red wine, )Citation79 cause a significant enhancement of neurotrophin (nerve growth factor [NGF] and brain-derived neurotrophic factor [BDNF])-mediated neurite outgrowth in PC12 cells.
Some of the phenolic compounds are capable of enhancing the expression levels of the differentiation markers (GAP-43, neurofilament light subunit, synaptophysin, synapsin, etc). Flavonoids that are capable of causing such an effect include luteolin,Citation64 daidzein,Citation80 7,8-DHF,Citation81 citrus HHMF,Citation68 puerarin (an isoflavone),Citation82 CAPE,Citation67 curcuminoids,Citation65 tectoridin (an isoflavone from Belamcandachinensis plant), hesperidin (a flavanone glycoside from the genus Citrus of plants), and also flavonols including kaempferol, quercetin, and isorhamnetin.Citation77
The neuropathy induced by chemotherapeutic agents such as cisplatin is an important side effect of these drugs.Citation83,Citation84 Two polyphenolic compounds, in separate studies, have been tested to block or reverse the cisplatin-induced neurite toxicity in PC12 cells. Phenoxodiol (2H-1-benzopyran-7-0,1,3-[4-hydroxyphenyl]), a compound related to genistein, showed significant neurite-protective effects against cisplatin at 1 μM, a concentration that was not toxic to the PC12 cells.Citation85,Citation86 Curcumin () has also shown similar protective effects against cisplatin-induced neurite toxicity in PC12 cells. Moreover, curcumin did not interfere with the cisplatin’s antitumor mode of action as assessed in vitro in HepG2 cells.Citation87
Activation of tropomyosin receptor kinases
Mammalian neurotrophins including NGF, BDNF, neurotrophin 3 (NT3), and neurotrophin 4 (NT4) play major roles in development, maintenance, repair, and survival of specific neuronal populations.Citation88–Citation90 Several lines of evidence indicate that decreased functioning of neurotrophins and their receptors can lead to neuronal injury and contribute to the pathogenesis of neurodegenerative diseases.Citation30,Citation91,Citation92 Thus, neurotrophins and bioactive compounds capable of activation of neurotrophin receptors have great potential for management of neurodegenerative diseases and other neurological disorders.Citation93,Citation94
Neurotrophins interact with two principal receptor types: p75NTR and the tropomyosin receptor kinase (Trk) receptors consisting of three receptors of TrkA, TrkB, and TrkC in mammals (also known as Ntrk1, Ntrk2, and Ntrk3).Citation95 Different patterns of Trk receptors’ expression exist throughout the mammalian brain and peripheral nervous system. TrkA is highly expressed in cholinergic neurons in the basal forebrain and also the peripheral nervous system, while TrkB and TrkC are highly expressed in the hippocampus.Citation96 These transmembrane receptors belonging to the group of receptor tyrosine kinases (RTKs) and the larger family of catalytic receptors include an extracellular neurotrophin-binding domain and an intracellular tyrosine kinase domain.Citation92,Citation97 NGF has a higher binding rate with TrkA, while BDNF and NT-4/5 bind with TrkB, and finally, NT-3 is the main ligand for TrkC receptor, with a lower affinity for TrkA and TrkB.Citation98 The neurotrophins-induced dimerization of the Trk receptors leads to activation through transphosphorylation of the cytoplasmic domain kinases and stimulates three major signaling pathways: phoshpatidyloinositol-3-kinase (PI3K)/Akt, mitogen-activated protein kinase (MAPK), and phospholipase C-γ1.Citation92 Downstream signaling principally promotes survival, growth, and neuronal differentiation and mediates neurogenesis and plasticity in many neuronal populations.Citation99–Citation101
There are only few polyphenolic compounds that act as direct agonists of Trk receptors and mimic the binding of neurotrophins, while several others stimulate the more downstream pathways leading to the neurotrophic effect (discussed in the following sections). 7,8-DHF provokes TrkB dimerization and tyrosine phosphorylation and activates downstream Akt and extracellular signal-regulated kinase (ERK) as potently as BDNF. 7,8-DHF also inhibits neuronal death in T48, a stably transfected TrkB murine cell line, and hippocampal neurons, and its activity can be inhibited by K252a, a Trk receptor antagonist.Citation102 In another study, 7,8-DHF strongly activated TrkB receptor and its downstream Akt and ERK1/2 pathways, prevented cell death, and promoted neuritogenesis in the retinal ganglion cells.Citation103 It was also shown in a later report that 7,8-DHF as a BDNF agonist causes phosphorylation of TrkB receptors and stimulates survival and neurite growth of cultured motoneurons: PI3K/Akt but not MAPK pathway was responsible for the survival and growth promoting effects of 7,8-DHF,Citation70 which is apparently different from MAPK activation mediated by 7,8-DHF in hippocampal neurons.Citation102
Furthermore, 7,8,3′-trihydroxyflavone, a compound related to 7,8-DHF, has shown similar effects on the survival of SGNs, phosphorylation of TrkB receptor, and ERK.Citation104 Diosmetin, another polyphenol compound belonging to flavonoids class of polyphenols, has also shown antiapoptotic effects, but it induces only weak phosphorylation of TrkB. Diosmetin also increased the phosphorylation of Akt and ERK.Citation102 Another flavonoid, epicatechin, restores TrkA phosphorylation in diabetic animals and reduces diabetes-induced neuronal cell death. It also blocks diabetes-induced p75NTR expression and p75NTR apoptotic pathway in vivo and in Müller cells.Citation105
Modulation of expression of neurotrophic factors and Trk receptors
Several polyphenols increase the expression levels of neurotrophins and Trk receptors (). Scutellarin, isolated from the traditional Chinese herb, Erigeron breviscapus (), alpinetin of the genus Alpinia, and also luteolin, calycosin, genistein, and isorhamnetin effectively upregulate the synthesis and release of NGF, glial cell line-derived neurotrophic factor (GDNF), and BDNF.Citation106,Citation107 The capacity of luteolin, genistein, calycosin, and isorhamnetin in increasing the expression of neurotrophins was mediated through estrogen signaling pathways. In the estrogen receptor (ER)-dependent pathway, the phosphorylated ER dimer was demonstrated to elevate the BDNF messenger RNA levels.Citation108 Scutellarin induced the expression of neurotrophins’ messenger RNAs and proteins through cyclic adenosine monophosphate response element-binding protein (P-CREB) and p-Akt signaling in primary rat astrocytes.Citation106
Table 2 Increased expression of neurotrophic factors by polyphenols in cell and animal models
Several polyphenolic compounds have been reported to increase BDNF levels; different flavonoids such as baicalein (5,6,7-trihydroxyflavone, ),Citation109 butein and fisetin of Rhusverniciflua (),Citation110 chrysin (5,7-dihydroxyflavone, ),Citation111 daidzein and genistein,Citation112 3,5,6,7,8,3′,4′-heptamethoxyflavone (HMF),Citation113 oroxylin A (5,7-dihidrixy-6-methoxyflavone, ),Citation114 puerarin,Citation115 and quercetin,Citation116 as well as other polyphenols such as CAPE,Citation117 curcumin,Citation118 and resveratrol,Citation79,Citation119–Citation122 have been reported to possess this property.
The aforementioned studies have examined BDNF levels under various experimental conditions. Some of the polyphenolic compounds, including fisetin and baicalin, one of the major flavonoids isolated from the roots of Scutellaria Baicalensis Georgi could activate the ERK-CREB pathway and exhibited memory-enhancing effects through the activation of CREB-BDNF pathway in the hippocampus of a mouse model of amnesia induced by scopolamine.Citation109,Citation110 In another study, chrysin was shown to rescue memory impairment and BDNF reduction caused by aging in mice.Citation111
Other studies have demonstrated the increased BDNF expression and neuroprotective effects of polyphenolic compounds in the settings of in vitro and in vivo models of ischaemia-, 6-hydroxydopamine (6-OHDA)-, or amyloid-β (Aβ)-mediated injury.Citation113,Citation115,Citation118,Citation123,Citation124 For example, HMF enhances BDNF production in astrocytes and induces neurogenesis in the hippocampus after brain ischemia, which is mediated by the activation of ERK1/2 and CREB pathways.Citation113 Puerarin has also a protective effect in 6-OHDA-lesioned rats by modulating BDNF expression and activation of the nuclear factor E2-related factor 2/antioxidant response element (Nrf2/ARE) signaling pathway.Citation115 Curcumin, on the other hand, was able to block BDNF reduction in the Aβ-infused rats through Akt/GSK-3β signaling pathway.Citation118 Antiamnesic and protective effects of luteolin against Aβ toxicity in mice was associated with the increase of BNDF as well as TrkB expression in the cerebral cortex of mice.Citation123
Many polyphenolic compounds have antidepressant effects that are associated with increased BDNF levels.Citation125,Citation126 Xiaochaihutang, a mixture of seven Chinese herbs that is traditionally used in People’s Republic of China for treatment of depressive-like symptoms, considerably increased BDNF, NGF, TrkB, and TrkA expressions in the hippocampus and also improved depression-like behaviors in chronic unpredictable mild stress (CUMS) model in rats.Citation127 A flavanonol compound, astilbin, reverses depressive-like behaviors in a mouse model of depression and its effect is mediated by the upregulation of BDNF and activation of ERK and Akt pathways.Citation128 Antidepressant effects of baicalein, resveratrol, and rosmarinic acid (a caffeic acid ester) are also mediated by BDNF-ERK-mediated neurotrophic action.Citation129–Citation131 Another polyphenol, ferulic acid, not only increases the proliferation of NSC/NPCs in vitro and in vivo but also produces an additive antidepressant-like effect in corticosterone-treated mice, mediated by CREB-BDNF signaling pathway.Citation72 Also, chrysin improves age-related memory decline in mice, an effect that is probably related to the modulation of BDNF production and free radical scavenging action of this compound.Citation111
ERK pathway activation
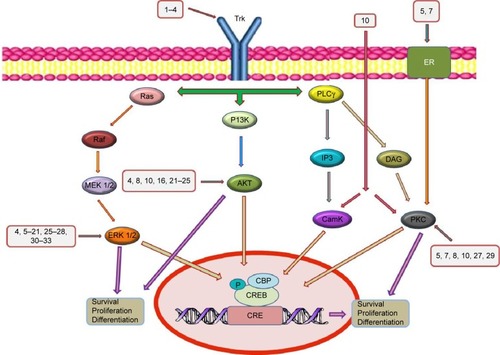
Modulation of neuronal survival signaling pathways may represent a promising approach to the management of central nervous system diseases. Several pathways including ERK1/2 and PI3K promote cell survival in the nervous system as well as other tissues.Citation132,Citation133 Recently, the ERK pathway, a part of MAPKs, has been implicated in several physiological functions of neurons, including proliferation, differentiation, survival, and regulation of response to various growth factors.Citation134–Citation138 The activation of ERK1/2 requires phosphorylation of threonine and tyrosine residues that is carried out by the upstream activator kinase, mitogen-activated protein kinase kinase (MEK). Activated ERK1/2 then changes its localization and phosphorylates different target molecules, including transcription regulators and cytoskeletal proteins ().Citation139,Citation140
Figure 4 Main signaling pathways that mediate the neurotrophic effects of various polyphenols.
Abbreviations: CaMK, Ca2+-calmodulin kinase; CREB, cyclic adenosine monophosphate response element-binding protein; CBP, CREB-binding protein; DAG, diacylglycerol; ER, estrogen receptor; ERK, extracellular signal-regulated kinase; IP3, inositol trisphosphate; MAPK, mitogen-activated protein kinase; MEK, mitogen-activated protein kinase kinase; PI3K, phosphatidylinsoitol-3-kinase; PKC, protein kinase C; PLCγ, phospholipase Cγ; Trk, tropomyosin receptor kinase.
ERK1/2 activation by some of the polyphenolic compounds promotes survival and antiapoptotic signaling in various cell lines (). It has been shown that ERK1/2 activation by luteolin protects PC12 cells against apoptosis. Furthermore, the cell viability of PC12 cells that were pretreated with U0126, a specific inhibitor of ERK1/2 kinase, was significantly reduced.Citation64
Table 3 Neuronal survival and differentiation induced by polyphenols and contribution of different neurotrophic pathways to these effects
Activation of the ERK1/2 pathway has mediated the neuroprotective activity against damaging insults.Citation132 Aβ peptide can cause a significant decrease in cell viability and neurite outgrowth and can induce apoptosis in neuronal cells. Liquiritin’s protection against Aβ-induced neuronal apoptosis and its effect on the differentiation of rats’ primary cultured hippocampal neurons were inhibited with a MAPK inhibitor.Citation141 Similarly, in another study, the decreased cell viability induced by Aβ25–35 was reported to be blocked by icaritin (a prenyl flavonoid derivative from Chinese tonic herb Epimedium). A blocker of ERK/MAPK pathway weakened this protective effect, which implied that ERK1/2 pathway is involved in the neuroprotective action of icaritin.Citation142 Likewise, rutin (3,3′,4,5,7-pentahydroxyflavone-3-rhamnoglucoside) has also shown beneficial effects against Aβ-induced neurotoxicity in rats through the activation of MAPK and BDNF.Citation143
As already mentioned, ERK1/2 signaling is crucial for neuronal differentiation and activation of associated cytoskeletal and synaptic proteins. It has been found that luteolin increases neurite outgrowth and expression of GAP-43 protein, a neuronal differentiation biomarker, and also heme oxygenase-1 (HO-1) expression in PC12 cells. All these effects could be blocked by pharmacological inhibition of ERK1/2.Citation64 In another report, it was established that luteolin induced microRNA-132 expression in PC12 cells and induced neurite outgrowth, while these effects were suppressed by protein kinase A (PKA) and MEK1/2 inhibitors, but not by protein kinase C (PKC) inhibitors.Citation144
Lin et alCitation64 have shown the involvement of both ERK and PKC signaling pathways in PC12 neurite outgrowth. Similarly, the involvement of the ERK and PKC signaling pathways in the process of neuritogenesis in PC12 cells was demonstrated in response to curcuminoids.Citation65 In another study, artepillin C-induced neurite outgrowth of PC12m3 cells has been shown to be inhibited by the ERK and p38 MAPK inhibitors. On the other hand, inhibition of ERK by U0126 entirely blocked artepillin C-mediated p38 MAPK phosphorylation of PC12m3 cells. It was suggested that the activation of p38 MAPK through the ERK signaling pathway is responsible for the artepillin C-induced neurite outgrowth of PC12m3 cells.Citation145 Finally, fisetin has been very effective in induction of PC12 cell differentiation, while MEK inhibitors considerably decreased fisetin-induced ERK activation and neurite outgrowth.Citation146 The different neurotrophic effects of polyphenols involving the ERK pathway are listed in –.
Table 4 Improvement of memory and modulation of other brain functions in animal models by polyphenols and involvement of neurotrophic signaling pathways
Table 5 Inhibition of neurotoxin-induced damage by polyphenols and neurotrophic signaling pathways that possibly contribute to this effect
PI3K pathway activation
A number of studies have demonstrated that PI3K and its downstream effector Akt are involved in neuronal survival and increased neurite outgrowth as well as other neurotrophic effects of polyphenolsCitation82,Citation106,Citation118,Citation128,Citation147 ( –). In this regard, Hoppe et alCitation118 showed that PI3K/Akt pathway is involved in curcumin-mediated neuroprotection in Aβ-induced cognitive impairment in rats. Similarly, scutellarin is reported to protect neurons against hypoxia by increasing neurotrophins through pCREB and pAkt signaling, but not MAPKs.Citation106 In cultured primary cortical neurons, methyl 3,4-dihydroxybenzoate, a phenolic acid derivative, promoted neuronal survival and neurite outgrowth via PI3K/Akt signaling pathway, and these effects could be inhibited by a PI3K-specific inhibitor.Citation147 In another report, evidence was provided that oroxylin A increased BDNF production and neuronal differentiation in primary cortical neurons by activation of the Akt pathway. Puerarin also protected dopaminergic cells and potentiated the effect of NGF on neuritogenesis in PC12 cells via activation of the ERK1/2 and PI3K/Akt pathways.Citation82 Finally, astilbin, a natural flavonoid, has been demonstrated to reduce depressive-like behaviors in mice models of depression by activation of the MAPK/ERK and PI3K/Akt pathways that are the downstream signaling pathways of BDNF.Citation128
Phosphorylation of CREB
Cyclic AMP response element (CRE) sequence is found in the regulatory region of several genes. CREB is a transcription regulator that recognizes CRE sequence and activates gene transcription, a process believed to play an important role in learning and memory in the brain. In neuronal cells, activation of the CREB pathway by neurotrophic factors such as NGF leads to the expression of genes that regulate survival, growth, synaptic plasticity, differentiation, dendritic spine formation, and long-term memory.Citation148,Citation149
Mantamadiotis et alCitation150 provided evidence that mice lacking CREB function in the brain showed neurodegenerative process in the hippocampus and dorsolateral striatum. Considering the importance of this pathway in the nervous system, we hereby summarize the studies on some phenolic compounds that activate CREB pathway and discuss their possible therapeutic effects in neurodegenerative diseases ().
Luteolin,Citation144 auraptene (a coumarin derivative, ),Citation151 curcumin and demethoxycurcumin,Citation65 nobiletin,Citation152 hesperetin (a flavonoid),Citation153 and citrus HHMFCitation68 have all been shown to increase CREB phosphorylation in PC12 cells. Chai et alCitation106 have also suggested that one of the signaling pathways related to neuroprotective effect of scutellarin is pCREB, which stimulates the production and release of neurotrophic factors in primary rat astrocyte cultures.
Resveratrol has shown neurotrophic effect in dopaminergic neuronsCitation79 and antidepressant-like effects in rats via activation of CREB in the hippocampus and amygdala.Citation130 In a study by Li et al,Citation154 green tea catechins-treated mice showed significantly higher CREB phosphorylation than the aged control mice. Ferulic acid has also been reported to increase CREB phosphorylation in the hippocampus of corticosterone-treated mice.Citation72 In an animal model of traumatic brain injury, 7,8-DHF restores the levels of significantly reduced CREB phosphorylation.Citation81 Baicalein increased pCREB in the hippocampus of miceCitation109 and also in NPC.Citation155 Administration of baicalein to rats with cognitive impairment induced by corticosterone significantly improved memory-associated decrease in the expression levels of BDNF and CREB proteins in the hippocampus of these animals.Citation156 Activation of CREB by apigenin (4′,5,7-trihydroxyflavone, )Citation157 and pinocembrin (5,7-dihydroxyflavanone)Citation158 was seen in transgenic Alzheimer’s disease mouse model. Finally, rutin and fustin flavonoids have been demonstrated to increase the expression of CREB in the hippocampi of rats and to attenuate Aβ-induced learning impairment in mice, respectively.Citation143,Citation159
Antioxidant activity
The CNS is unique in its high susceptibility to oxidative stress, which is a result of its high oxygen consumption and also the presence of large amounts of fatty acids and metals.Citation160 Several lines of evidence suggest that oxidative stress is involved in the pathogenesis of neurodegenerative disorders such as Alzheimer’s and Parkinson’s disease.Citation61,Citation161 The antioxidant effect of polyphenols has long been studied as a mechanism of their neuroprotection against neurodegenerative disordersCitation61,Citation162 and other neurological diseases.Citation163 Although the direct interaction of polyphenols with reactive oxygen species (ROS) does not seem to be very likely to happen in vivo as a main mechanism of action, their antioxidant effects are most probably exerted through other mechanisms such as activation of Nrf2 pathway, upregulation of antioxidant enzymes, induction of hypoxia signal transduction (HIF-1-α pathway), and interaction with metal ions as sources of ROS.Citation12,Citation161,Citation164
For example, EGCGCitation165 and resveratrolCitation166 have been reported to exert their neuroprotective action through activation of the HIF-1 pathway. Mangiferin and morin (3,5,7,2′,4′-pentahydroxyflavone) revealed considerable antioxidant and antiapoptotic properties through the activation of antioxidant enzymes.Citation167
Protection against damage induced by neurotoxins
Cellular, biochemical, and animal studies have shown that Aβ is a crucial factor in the pathogenesis of Alzheimer’s disease.Citation168,Citation169 There are several reports suggesting that some polyphenols protect neuronal cells against Aβ-induced neuronal cell death or other forms of neuronal injury (). For instance, icaritin was shown to protect primary rat cortical neuronal cells against apoptosis induced by Aβ25–35 insult.Citation142 Similarly, Ushikubo et alCitation170 demonstrated that 3,3′,4′,5,5′-pentahydroxyflavone prevents Aβ fibril formation and that lowering fibril formation decreases Aβ-induced cell death in rat hippocampal neuronal cells. In another study, ursolic acid, p-coumaric acid, and gallic acid extracted from Cornifructus plant were shown to attenuate apoptotic features such as morphological nuclear changes, DNA fragmentation, and cell blebbing induced by Aβ peptide in PC12 cells.Citation171 The major flavonoids of cocoa, epicatechin and catechin, protect PC12 cells from Aβ-induced neurotoxicity.Citation172 Carnosic acid (), a highly bioactive phenolic compound found in rosemary (Rosmarinus officinalis), and liquiritin have shown a protective effect against Aβ in SH-SY5Y human neuroblastoma cells and primary cultured hippocampal neurons, respectively.Citation141,Citation173
6-OHDA is a compound used to induce a Parkinson-like pathology in vitro and in vivo. It has been reported that baicaleinCitation174 and also ferulic acid and caffeic acid derivatives, both belonging to hydroxycinnamic acid family (),Citation60 protect neuronal SH-SY5Y cells against 6-OHDA.
Hydrogen peroxide is an ROS that has been shown to induce neuronal cell damage in experimental models.Citation175 It has been shown that several polyphenols such as quercetin in cultured neuronal precursor cells,Citation176 7,8-DHF in retinal ganglion and RGC-5 cells,Citation103 and caffeic acid esters in PC12 cellsCitation53 are able to exert protective effects against ROS. In addition, other authors suggest that the neuroprotective effects of 7,8-DHF are mediated by its ability to scavenge ROS and increase cellular glutathione levels.Citation55
Other neurotoxins have also been applied to produce experimental models to assess the neuroprotective capacity of polyphenolic compounds. CAPE protects PC12 cells against dopaminergic neurotoxin MPP+ (1-methyl-4-phenylpyridinium).Citation67 Administration of 7,8-DHF prevented neuronal death in mouse brain induced by kainic acid.Citation102Icariin is another polyphenol compound that protects primary cultured rat hippocampal neuronal cells against corticosterone-induced apoptosis.Citation177 In addition, baicalein was also shown to inhibit necrotic cell death damage in NPCs and to attenuate impairment of hippocampal neurogenesis caused by irradiation.Citation155
Polyphenols have also shown therapeutic potential in animal models of neurodegenerative diseases induced by various neurotoxins. In an Aβ-induced amnesia model in mice, oral administration of luteolin mitigated learning and memory impairment.Citation123 Curcumin has also been shown to be effective in preventing neuroinflammation, tau hyper-phosphorylation, and behavioral impairments, triggered by Aβ in vivo.Citation118
Activation of the Nrf2 signaling pathway
One of the elegant mechanisms that neuronal cells have adapted to protect themselves against oxidative stress and other insults is Nrf2 pathway and the binding of this master transcriptional regulator with ARE in the regulatory region of many genes, which leads to the expression of several enzymes with antioxidant and detoxification capacities ().Citation178 The main enzymes that are transcribed under the control of ARE include γ-glutamylcysteine synthetase, glutathione peroxidase, glutathione S-transferase, HO-1, NADPH quinine oxidoreductase 1, peroxiredoxin, sulfiredoxin, thioredoxin, and thioredoxin reductase all of which have important roles in the protection of cells.Citation179–Citation181 The pivotal finding that Nrf2-knockout mice exhibit a severe deficiency in the coordinated regulation of gene expression and their susceptibility to oxidative damage indicate the crucial role of Nrf2 in maintaining intracellular redox homeostasis and antioxidant defense mechanism.Citation182 The role of this pathway and the detoxification enzymes that it regulates has been especially emphasized in CNS diseases.Citation181
Figure 5 Polyphenols activate Keap1/Nrf2/ARE pathway and increase the expression of detoxification/antioxidant enzymes.
Abbreviations: ARE, antioxidant response element; CAT, catalase; EGCG, epigallocatechin–gallate; ERK, extracellular signal-regulated kinase; GCL, γ-glutamylcysteine synthetase; GPx, glutathione peroxidase; GST, glutathione S-transferase; HO-1, heme oxygenase-1; Nrf2, Nuclear factor E2-related factor 2; PKC, protein kinase C; PRX, peroxiredoxin; SOD, superoxide dismutase; Trx, thioredoxin.
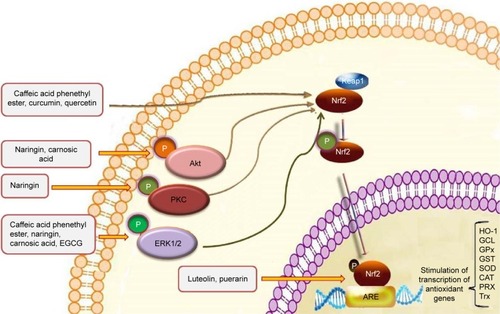
Some phenolic compounds such as luteolinCitation64 and puerarinCitation82 in PC12 cells and also CAPE in dopaminergic neuronsCitation117 enhance the binding of Nrf2 to ARE and increase the expression of HO-1. In cultured neurons, EGCG was able to protect cells against oxidative stress by increasing HO-1 expression via activation of the transcription factor Nrf2.Citation183 Li et alCitation184 evaluated the neuroprotective effect of puerarin on lesioned substantia nigra induced by 6-OHDA. Their findings showed that puerarin effectively protects neurons in substantia nigra by modulating brain-derived neurotrophic factor (BDNF) expression and also by activating the Nrf2/ARE pathway.Citation115 Carnosic acid and common sage (Salvia officinalis) plants also showed neuroprotective effects by activating Nrf2 in PC12h cells.Citation184
Polyphenols and activation of other neurotrophic pathways
Polyphenols having multiple beneficial effects on the nervous system could provide an important resource for the development of new drugs for management of neurodegenerative diseases. In addition to the aforementioned signaling pathways involved in the neurotrophic action of polyphenols, other mechanisms may also be involved. Daidzein has caused significant axonal outgrowth via upregulation of GAP-43 expression in hippocampal neurons in culture. Interestingly, daidzein-promoted phosphorylation of PKC, and GAP-43 was abolished by pretreatment with ER and PKC antagonist. These results suggest that ER-mediated PKC phosphorylation of GAP-43 may play a role in daidzein-mediated axonal outgrowth.Citation80 Hesperetin can also exhibit multiple neurotrophic effects via ER- and TrkA-mediated parallel pathways.Citation153
The Na+/K+/2Cl cotransporter (NKCC) is a member of the cation–chloride cotransporter family and is involved in the transport of chloride ion(s) coupled with cation(s) across the plasma membrane.Citation185 A previous study has shown that NGF treatment of PC12D cells increased the expression of NKCC1 protein.Citation186 In another report, it has been demonstrated that knockdown of NKCC1 strongly inhibits NGF induced-neurite outgrowth in PC12 cells. Interestingly, quercetin also promoted NGF-induced neurite outgrowth by increasing Cl−, and knockdown of NKCC1 inhibited this stimulatory effect. In these cells, intracellular Cl− affects microtubule polymerization via modulation of intrinsic GTPase activity of tubulin.Citation187
A2A subtype of adenosine receptors has been reported to elevate the expression of BDNF as well as synaptic actions of BDNF.Citation188,Citation189 This receptor also activates TrkB receptor and Akt signaling pathway, which induces neuronal survival and modulates neurite outgrowth in several different cell types.Citation190–Citation192 Jeon et alCitation114 have recently shown that oroxylin A could induce BDNF production in cortical neurons via activation of A2A receptor, which induced cellular survival, synapse formation, and neurite outgrowth. In another report, the adenosine A2A receptor inhibitor could block methyl 3,4-dihydroxybenzoate-induced neuronal survival and neurite outgrowth in cultured primary cortical neurons.Citation147
Conclusion
Polyphenols with multiple beneficial effects in the nervous system could provide an important resource for the discovery of new neurotrophic agents. Agonistic action on Trk receptors, activation of the ERK, PI3Kinase/Akt and CREB pathways, activation of the Nrf2 pathway and upregulation of antioxidant and detoxification enzymes, as well as several other mechanisms underlie the neurotrophic action of different polyphenols. A better understanding of the neurotrophic effects and the molecular mechanisms of action of these compounds could help design better agents for management of neurodegenerative diseases and other disorders of the nervous system.
Acknowledgments
The authors thank the support provided by the vice chancellor for research, Shiraz University of Medical Sciences.
Disclosure
The authors report no conflict of interest in this work.
References
- ManachCScalbertAMorandCRémésyCJiménezLPolyphenols: food sources and bioavailabilityAm J Clin Nutr200479572774715113710
- LinkABalaguerFGoelACancer chemoprevention by dietary polyphenols: promising role for epigeneticsBiochem Pharmacol201080121771179220599773
- WengCJYenGCChemopreventive effects of dietary phytochemicals against cancer invasion and metastasis: phenolic acids, monophenol, polyphenol, and their derivativesCancer Treat Rev2012381768721481535
- DarveshASBishayeeAChemopreventive and therapeutic potential of tea polyphenols in hepatocellular cancerNutr Cancer201365332934423530632
- AnnuzziGBozzettoLCostabileGDiets naturally rich in polyphenols improve fasting and postprandial dyslipidemia and reduce oxidative stress: a randomized controlled trialAm J Clin Nutr201499346347124368433
- ShayJElbazHALeeIZielskeSPMalekMHHuttemannMMolecular mechanisms and therapeutic effects of (−)-epicatechin and other polyphenols in cancer, inflammation, diabetes, and neurodegenerationOxid Med Cell Longev2015201518126026180580
- DaiJMumperRJPlant phenolics: extraction, analysis and their antioxidant and anticancer propertiesMolecules201015107313735220966876
- PocernichCBLangeMLSultanaRButterfieldDANutritional approaches to modulate oxidative stress in Alzheimer’s diseaseCurr Alzheimer Res20118545246921605052
- VauzourDVafeiadouKRodriguez-MateosARendeiroCSpencerJPThe neuroprotective potential of flavonoids: a multiplicity of effectsGenes Nutr200833–411512618937002
- ScalbertAManachCMorandCRémésyCJiménezLDietary polyphenols and the prevention of diseasesCrit Rev Food Sci Nutr200545428730616047496
- BhullarKSRupasingheHPolyphenols: multipotent therapeutic agents in neurodegenerative diseasesOxid Med Cell Longev20132013
- EbrahimiASchluesenerHNatural polyphenols against neurodegenerative disorders: potentials and pitfallsAgeing Res Rev201211232934522336470
- LiHZhangYCaoLCurcumin could reduce the monomer of TTR with Tyr114Cys mutation via autophagy in cell model of familial amyloid polyneuropathyDrug Des Devel Ther2014821212128
- LarsonAJSymonsJDJaliliTTherapeutic potential of quercetin to decrease blood pressure: review of efficacy and mechanismsAdv Nutr201231394622332099
- QuiñonesMMiguelMAleixandreABeneficial effects of polyphenols on cardiovascular diseasePharmacol Res201368112513123174266
- FrescoPBorgesFMarquesMDinizCThe anticancer properties of dietary polyphenols and its relation with apoptosisCurr Pharm Des201016111413420214622
- NishiumiSMiyamotoSKawabataKDietary flavonoids as cancer-preventive and therapeutic biofactorsFront Biosci2010313321362
- MartinMAGoyaLRamosSPotential for preventive effects of cocoa and cocoa polyphenols in cancerFood Chem Toxicol20135633635123439478
- ConteEFagoneEFrucianoMGiliEIemmoloMVancheriCAnti-inflammatory and antifibrotic effects of resveratrol in the lungHistol Histopathol201430552352925515609
- GonzalezRBallesterILopez-PosadasREffects of flavonoids and other polyphenols on inflammationCrit Rev Food Sci Nutr201151433136221432698
- BabuPVALiuDGilbertERRecent advances in understanding the anti-diabetic actions of dietary flavonoidsJ Nutr Biochem201324111777178924029069
- AssiniJMMulvihillEEHuffMWCitrus flavonoids and lipid metabolismCurr Opin Lipidol2013241344023254473
- YaoZZhangLJiGEfficacy of polyphenolic ingredients of Chinese herbs in treating dyslipidemia of metabolic syndromesEur J Integr Med2014123135146
- IslamBSharmaCAdemAAburawiEOjhaSInsight into the mechanism of polyphenols on the activity of HMGR by molecular dockingDrug Des Devel Ther2015949434951
- ChirumboloSDietary assumption of plant polyphenols and prevention of allergyCurr Pharm Des201420681183923701556
- CuevasASaavedraNSalazarLAAbdallaDSModulation of immune function by polyphenols: possible contribution of epigenetic factorsNutrients2013572314233223812304
- CiminoSSortinoGFavillaVPolyphenols: key issues involved in chemoprevention of prostate cancerOxid Med Cell Longev20122012
- JinHLengQLiCDietary flavonoid for preventing colorectal neoplasmsCochrane Libr20128CD009350
- ZandHChemopreventive and chemosensitization potential of flavonoids in acute lymphoblastic leukemiaJ Pediatr Biochem2012211521
- QuerfurthHWLaFerlaFMAlzheimer’s diseaseNew Engl J Med2010362432934420107219
- CitronMAlzheimer’s disease: strategies for disease modificationNat Rev Drug Discov20109538739820431570
- MangialascheFSolomonAWinbladBMecocciPKivipeltoMAlzheimer’s disease: clinical trials and drug developmentLancet Neurol20109770271620610346
- ShayJElbazHALeeIZielskeSPMalekMHHüttemannMMolecular mechanisms and therapeutic effects of (−)-epicatechin and other polyphenols in cancer, inflammation, diabetes, and neurodegenerationOxid Med Cell Longev20152015
- AlbarracinSLStabBCasasZEffects of natural antioxidants in neurodegenerative diseaseNutr Neurosci20121511922305647
- KrikorianRBoespflugELFleckDEConcord grape juice supplementation and neurocognitive function in human agingJ Agric Food Chem201260235736574222468945
- WitteAVKertiLMarguliesDSFlöelAEffects of resveratrol on memory performance, hippocampal functional connectivity, and glucose metab olism in healthy older adultsJ Neurosci201434237862787024899709
- Kesse-GuyotEFezeuLAndreevaVATotal and specific polyphenol intakes in midlife are associated with cognitive function measured 13 years laterJ Nutr20121421768322090468
- LetenneurLProust-LimaCLe GougeADartiguesJ-FBarberger-GateauPFlavonoid intake and cognitive decline over a 10-year periodAm J Epidemiol2007165121364137117369607
- CimrováBBudáčSMelicherováUJergelováMJaglaFElectrophysiological evidence of the effect of natural polyphenols upon the human higher brain functionsNeuro Endocrinol Lett201132446446821876511
- TorreRSolaSPonsMEpigallocatechin-3-gallate, a DYRK1A inhibitor, rescues cognitive deficits in Down syndrome mouse models and in humansMol Nutr Food Res201458227828824039182
- GundimedaUMcNeillTHSchiffmanJEHintonDRGopalakrishnaRGreen tea polyphenols potentiate the action of nerve growth factor to induce neuritogenesis: possible role of reactive oxygen speciesJ Neurosci Res201088163644365520936703
- SuganumaMOkabeSOniyamaMTadaYItoHFujikiHWide distribution of [3H](−)-epigallocatechin gallate, a cancer preventive tea polyphenol, in mouse tissueCarcinogenesis19981910177117769806157
- El MohsenMMAKuhnleGRechnerARUptake and metabolism of epicatechin and its access to the brain after oral ingestionFree Radic Biol Med200233121693170212488137
- RossiLMazzitelliSArcielloMCapoCRotilioGBenefits from dietary polyphenols for brain aging and Alzheimer’s diseaseNeurochem Res200833122390240018415677
- VingtdeuxVGilibertoLZhaoHAMP-activated protein kinase signaling activation by resveratrol modulates amyloid-β peptide metabolismJ Biol Chem2010285129100911320080969
- YoudimKAQaiserMZBegleyDJRice-EvansCAAbbottNJFlavonoid permeability across an in situ model of the blood–brain barrierFree Radic Biol Med200436559260414980703
- YoudimKADobbieMSKuhnleGProteggenteARAbbottNJRice-EvansCInteraction between flavonoids and the blood–brain barrier: in vitro studiesJ Neurochem200385118019212641740
- FiruziOLacannaAPetrucciRMarrosuGSasoLEvaluation of the antioxidant activity of flavonoids by “ferric reducing antioxidant power” assay and cyclic voltammetryBiochim Biophys Acta200517211–317418415652192
- FiruziOMladěnkaPPetrucciRMarrosuGSasoLHypochlorite scavenging activity of flavonoidsJ Pharm Pharmacol200456680180715231046
- DueñasMGonzález-ManzanoSGonzález-ParamásASantos- BuelgaCAntioxidant evaluation of O-methylated metabolites of catechin, epicatechin and quercetinJ Pharm Biomed Anal201051244344919442472
- LemmensKJvan de WierBVaesNThe flavonoid 7-mono-O-(β-hydroxyethyl)-rutoside is able to protect endothelial cells by a direct antioxidant effectToxicol in Vitro201428453854324412621
- MartínMÁFernández-MillánERamosSBravoLGoyaLCocoa flavonoid epicatechin protects pancreatic beta cell viability and function against oxidative stressMol Nutr Food Res201458344745624115486
- GarridoJGasparAGarridoEMAlkyl esters of hydroxycinnamic acids with improved antioxidant activity and lipophilicity protect PC12 cells against oxidative stressBiochimie201294496196722210493
- GasparAMartinsMSilvaPDietary phenolic acids and derivatives. Evaluation of the antioxidant activity of sinapic acid and its alkyl estersJ Agric Food Chem20105821112731128020949946
- ChenJChuaK-WChuaCCAntioxidant activity of 7,8-dihydroxyflavone provides neuroprotection against glutamate-induced toxicityNeurosci Lett2011499318118521651962
- VauzourDDietary polyphenols as modulators of brain functions: biological actions and molecular mechanisms underpinning their beneficial effectsOxid Med Cell Longev2012201291427322701758
- Gutierrez-MerinoCLopez-SanchezCLagoaRSamhan-AriasAKBuenoCGarcia-MartinezVNeuroprotective actions of flavonoidsCurr Med Chem20111881195121221291366
- HwangS-LShihP-HYenG-CNeuroprotective effects of citrus flavonoidsJ Agric Food Chem201260487788522224368
- VenkatesanRJiEKimSYPhytochemicals that regulate neurode-generative disease by targeting neurotrophins: a comprehensive reviewBiomed Res Int2015201581406826075266
- SilvaTBravoJSummavielleTBiology-oriented development of novel lipophilic antioxidants with neuroprotective activityRSC Adv20155211580015811
- DarveshASCarrollRTBishayeeAGeldenhuysWJVan der SchyfCJOxidative stress and Alzheimer’s disease: dietary polyphenols as potential therapeutic agentsExpert Rev Neurother201010572974520420493
- FiruziOMiriRTavakkoliMSasoLAntioxidant therapy: current status and future prospectsCurr Med Chem201118253871388821824100
- YuanJYanknerBAApoptosis in the nervous systemNature2000407680580280911048732
- LinC-WWuM-JLiuIY-CSuJ-DYenJ-HNeurotrophic and cytoprotective action of luteolin in PC12 cells through ERK-dependent induction of Nrf2-driven HO-1 expressionJ Agric Food Chem20105874477448620302373
- LiaoK-KWuM-JChenP-YCurcuminoids promote neurite outgrowth in PC12 cells through MAPK/ERK-and PKC-dependent pathwaysJ Agric Food Chem201160143344322145830
- ReznichenkoLAmitTYoudimMMandelSGreen tea polyphenol (−)-epigallocatechin-3-gallate induces neurorescue of long-term serum-deprived PC12 cells and promotes neurite outgrowthJ Neurochem20059351157116715934936
- dos SantosNAGMartinsNMde Barros SilvaRFerreiraRSSistiFMdos SantosACCaffeic acid phenethyl ester (CAPE) protects PC12 cells from MPP+ toxicity by inducing the expression of neuron-typical proteinsNeuroToxicology20144513113825454720
- LaiH-CWuM-JChenP-YNeurotrophic effect of citrus 5-hydroxy-3, 6, 7, 8, 3′, 4′-hexamethoxyflavone: promotion of neurite outgrowth via cAMP/PKA/CREB pathway in PC12 cellsPLoS One2011611e2828022140566
- YangS-HLiaoC-CChenYSyuJ-PJengC-JWangS-MDaidzein induces neuritogenesis in DRG neuronal culturesJ Biomed Sci2012198022931352
- TsaiTKlausmeyerAConradR7,8-Dihydroxyflavone leads to survival of cultured embryonic motoneurons by activating intracellular signaling pathwaysMol Cell Neurosci201356182823500004
- TangsaengvitNKitphatiWTadtongSBunyapraphatsaraNNukoolkarnVNeurite outgrowth and neuroprotective effects of quercetin from Caesalpinia mimosoides Lamk. on cultured P19-derived neuronsEvid Based Complement Alternat Med2013201383805123840266
- YabeTHiraharaHHaradaNFerulic acid induces neural progenitor cell proliferation in vitro and in vivoNeuroscience2010165251552419837139
- TangBLProtein trafficking mechanisms associated with neurite outgrowth and polarized sorting in neuronsJ Neurochem200179592393011739603
- DasKPFreudenrichTMMundyWRAssessment of PC12 cell differentiation and neurite growth: a comparison of morphological and neurochemical measuresNeurotoxicol Teratol200426339740615113601
- NakajimaK-INiisatoNMarunakaYGenistein enhances the NGF-induced neurite outgrowthBiomed Res201132535135622033305
- ChenZ-AWangJ-LLiuR-TLiquiritin potentiate neurite outgrowth induced by nerve growth factor in PC12 cellsCytotechnology2009601–312513219789989
- XuSLChoiRCZhuKYIsorhamnetin, a flavonol aglycone from Ginkgo biloba L., induces neuronal differentiation of cultured PC12 cells: potentiating the effect of nerve growth factorEvid Based Complement Alternat Med2012201227827322761636
- LiYChenXSatakeMOshimaYOhizumiYAcetylated flavonoid glycosides potentiating NGF action from scoparia dulcisJ Nat Prod200467472572715104516
- ZhangFWangY-YLiuHResveratrol produces neurotrophic effects on cultured dopaminergic neurons through prompting astroglial BDNF and GDNF releaseEvid Based Complement Alternat Med2012201293760523304227
- WangPJengC-JChienC-LWangS-MSignaling mechanisms of daidzein-induced axonal outgrowth in hippocampal neuronsBiochem Biophys Res Commun2008366239340018062921
- AgrawalRNobleETyagiEZhuangYYingZGomez-PinillaFFlavonoid derivative 7,8-DHF attenuates TBI pathology via TrkB activationBiochim Biophys Acta20151852586287225661191
- ZhaoJChengYYFanWBotanical drug puerarin coordinates with nerve growth factor in the regulation of neuronal survival and neuritogenesis via activating ERK1/2 and PI3K/Akt signaling pathways in the neurite extension processCNS Neurosci Ther2015211617025310912
- TaLEEspesetLPodratzJWindebankAJNeurotoxicity of oxaliplatin and cisplatin for dorsal root ganglion neurons correlates with platinum–DNA bindingNeurotoxicology2006276992100216797073
- QuasthoffSHartungHPChemotherapy-induced peripheral neuropathyJ Neurol2002249191711954874
- KleinRBrownDTurnleyAMPhenoxodiol protects against Cisplatin induced neurite toxicity in a PC-12 cell modelBMC Neurosci2007816117672914
- KamsteegMRutherfordTSapiEPhenoxodiol – an isoflavone analog – induces apoptosis in chemoresistant ovarian cancer cellsOncogene200322172611262012730675
- DikshitPGoswamiAMishraACatterjeeMJanaNRCurcumin induces stress response, neurite outgrowth and prevent NF-κB activation by inhibiting the proteasome functionNeurotox Res200691293716464750
- LevyYSGilgun-SherkiYMelamedEOffenDTherapeutic potential of neurotrophic factors in neurodegenerative diseasesBio Drugs200519297127
- HymanCHoferMBardeYABDNF is a neurotrophic factor for dopaminergic neurons of the substantia nigraNature199135063152302322005978
- MaisonpierrePCBelluscioLFriedmanBNT-3, BDNF, and NGF in the developing rat nervous system: parallel as well as reciprocal patterns of expressionNeuron1990545015091688327
- SchindowskiKBelarbiKBueeLNeurotrophic factors in Alzheimer’s disease: role of axonal transportGenes Brain Behav20087s1435618184369
- LongoFMMassaSMSmall-molecule modulation of neurotrophin receptors: a strategy for the treatment of neurological diseaseNat Rev Drug Discov201312750752523977697
- NagaharaAHTuszynskiMHPotential therapeutic uses of BDNF in neurological and psychiatric disordersNat Rev Drug Discov201110320921921358740
- AllenSJWatsonJJShoemarkDKBaruaNUPatelNKGDNF, NGF and BDNF as therapeutic options for neurodegenerationPharmacol Ther2013138215517523348013
- HuangEJReichardtLFTrk receptors: Roles in neuronal signal transductionAnnu Rev Biochem2012722003609642
- WebsterNJPirrungMCSmall molecule activators of the Trk receptors for neuroprotectionBMC Neuroscience20089Suppl 2S119090982
- ChaoMVNeurotrophins and their receptors: a convergence point for many signalling pathwaysNat Rev Neurosci20034429930912671646
- UltschMHWiesmannCSimmonsLCCrystal structures of the neurotrophin-binding domain of TrkA, TrkB and TrkCJ Mol Biol1999290114915910388563
- HuangEJReichardtLFTrk receptors: roles in neuronal signal transductionAnnu Rev Biochem200372160964212676795
- KaplanDRMillerFDNeurotrophin signal transduction in the nervous systemCurr Opin Neurobiol200010338139110851172
- PatapoutianAReichardtLFTrk receptors: mediators of neurotrophin actionCurr Opin Neurobiol200111327228011399424
- JangS-WLiuXYepesMA selective TrkB agonist with potent neurotrophic activities by 7,8-dihydroxyflavoneProc Natl Acad Sci U S A201010762687269220133810
- GuptaVKYouYLiJCKlistornerAGrahamSLProtective effects of 7,8-dihydroxyflavone on retinal ganglion and RGC-5 cells against excitotoxic and oxidative stressJ Mol Neurosci20134919610423054592
- YuQChangQLiuXProtection of spiral ganglion neurons from degeneration using small-molecule TrkB receptor agonistsJ Neurosci20133332130421305223926258
- Al-GayyarMMatragoonSPillaiBAliTAbdelsaidMEl-RemessyAEpicatechin blocks pro-nerve growth factor (proNGF)-mediated retinal neurodegeneration via inhibition of p75 neurotrophin receptor proNGF expression in a rat model of diabetesDiabetologia201154366968021136036
- ChaiLGuoHLiHScutellarin and caffeic acid ester fraction, active components of Dengzhanxixin injection, upregulate neurotrophins synthesis and release in hypoxia/reoxygenation rat astrocytesJ Ethnopharmacol2013150110010724012966
- XuSLBiCWChoiRCFlavonoids induce the synthesis and secretion of neurotrophic factors in cultured rat astrocytes: a signaling response mediated by estrogen receptorEvid Based Complement Alternat Med2013201312707523878590
- SohrabjiFMirandaRToran-AllerandCDIdentification of a putative estrogen response element in the gene encoding brain-derived neurotrophic factorProc Natl Acad Sci U S A1995922411110111147479947
- ParkSJKimDHKimJMMismatch between changes in baicalein-induced memory-related biochemical parameters and behavioral consequences in mouseBrain Res2010135514115020691671
- ChoNLeeKYHuhJCognitive-enhancing effects of Rhus verniciflua bark extract and its active flavonoids with neuroprotective and anti-inflammatory activitiesFood Chem Toxicol20135835536123688860
- SouzaLCAntunesMSBorges FilhoCFlavonoid Chrysin prevents age-related cognitive decline via attenuation of oxidative stress and modulation of BDNF levels in aged mouse brainPharmacol Biochem Behav2015134223025931267
- PanMHanHZhongCGengQEffects of genistein and daidzein on hippocampus neuronal cell proliferation and BDNF expression in H19-7 neural cell lineJ Nutr Health Aging201216438939422499464
- OkuyamaSShimadaNKajiMHeptamethoxyflavone, a citrus flavonoid, enhances brain-derived neurotrophic factor production and neurogenesis in the hippocampus following cerebral global ischemia in miceNeurosci Lett2012528219019522985518
- JeonSJBakHSeoJOroxylin A induces BDNF expression on cortical neurons through adenosine A2A receptor stimulation: a possible role in neuroprotectionBiomol Ther201220127
- LiRLiangTXuLZhengNZhangKDuanXPuerarin attenuates neuronal degeneration in the substantia nigra of 6-OHDA-lesioned rats through regulating BDNF expression and activating the Nrf2/ARE signaling pathwayBrain Res201315231923747813
- LiuPZouDYiLQuercetin ameliorates hypobaric hypoxia-induced memory impairment through mitochondrial and neuron function adaptation via the PGC-1α pathwayRestor Neurol Neurosci201533214315725588463
- KurauchiYHisatsuneAIsohamaYMishimaSKatsukiHCaffeic acid phenethyl ester protects nigral dopaminergic neurons via dual mechanisms involving haem oxygenase-1 and brain-derived neurotrophic factorBr J Pharmacol201216631151116822224485
- HoppeJBCoradiniKFrozzaRLFree and nanoencapsulated curcumin suppress β-amyloid-induced cognitive impairments in rats: involvement of BDNF and Akt/GSK-3β signaling pathwayNeurobiol Learn Mem201310613414423954730
- ZhangFLuY-FWuQLiuJShiJ-SResveratrol promotes neurotrophic factor release from astrogliaExp Biol Med20122378943948
- RahvarMNiksereshtMShafieeSMEffect of oral resveratrol on the BDNF gene expression in the hippocampus of the rat brainNeurochem Res201136576176521221775
- YuanHZhangJLiuHLiZThe protective effects of resveratrol on Schwann cells with toxicity induced by ethanol in vitroNeurochem Int201363314615323770283
- MoriyaJChenRYamakawaJ-ISasakiKIshigakiYTakahashiTResveratrol improves hippocampal atrophy in chronic fatigue mice by enhancing neurogenesis and inhibiting apoptosis of granular cellsBiol Pharm Bull201134335435921372384
- LiuRGaoMQiangG-FThe anti-amnesic effects of luteolin against amyloid β 25–35 peptide-induced toxicity in mice involve the protection of neurovascular unitNeuroscience200916241232124319442706
- LeeYWKimDHJeonSJNeuroprotective effects of salvianolic acid B on an Aβ 25–35 peptide-induced mouse model of Alzheimer’s diseaseEur J Pharmacol20137041707723461850
- DonatoFde GomesMGGoesATRHesperidin exerts antidepressant-like effects in acute and chronic treatments in mice: Possible role of L-arginine-NO-cGMP pathway and BDNF levelsBrain Res Bull2014104192624709058
- WangZGuJWangXAntidepressant-like activity of resveratrol treatment in the forced swim test and tail suspension test in mice: the HPA axis, BDNF expression and phosphorylation of ERKPharmacol Biochem Behav201311210411024125781
- SuGYYangJYWangFAntidepressant-like effects of Xiaochaihutang in a rat model of chronic unpredictable mild stressJ Ethnopharmacol2014152121722624440317
- LvQ-QWuW-JGuoX-LAntidepressant activity of astilbin: involvement of monoaminergic neurotransmitters and BDNF signal pathwayBiol Pharm Bull201437698799524681540
- XiongZJiangBWuP-FAntidepressant effects of a plant-derived flavonoid baicalein involving extracellular signal-regulated kinases cascadeBiol Pharm Bull201134225325921415537
- LiuDXieKYangXResveratrol reverses the effects of chronic unpredictable mild stress on behavior, serum corticosterone levels and BDNF expression in ratsBehav Brain Res201426491624503118
- JinXLiuPYangFZhangY-HMiaoDRosmarinic acid ameliorates depressive-like behaviors in a rat model of CUS and up-regulates BDNF levels in the hippocampus and hippocampal-derived astrocytesNeurochem Res20133891828183723756732
- HetmanMGozdzARole of extracellular signal regulated kinases 1 and 2 in neuronal survivalEur J Biochem2004271112050205515153093
- HetmanMXiaZSignaling pathways mediating anti-apoptotic action of neurotrophinsActa Neurobiol Exp2000604531546
- SatohYKobayashiYTakeuchiAPagèsGPouysségurJKazamaTDeletion of ERK1 and ERK2 in the CNS causes cortical abnormalities and neonatal lethality: Erk1 deficiency enhances the impairment of neurogenesis in Erk2-deficient miceJ Neurosci20113131149115521248139
- ParmarMSJaumotteJDWyrostekSLZigmondMJCavanaughJEThe role of ERK1, 2, and 5 in dopamine neuron survival during agingNeurobiol Aging201435366967924411019
- LiZTheusMHWeiLRole of ERK 1/2 signaling in neuronal differentiation of cultured embryonic stem cellsDev Growth Differ200648851352317026715
- LiQChenMLiuHYangLYangTHeGThe dual role of ERK signaling in the apoptosis of neuronsFront Biosci20131914111417
- MunshiARameshRMitogen-activated protein kinases and their role in radiation responseGenes Cancer201349–1040140824349638
- MoustafaKAbuQamarSJarrarMAl-RajabAJTrémouillaux-GuillerJMAPK cascades and major abiotic stressesPlant Cell Rep20143381217122524832772
- KomisGIllésPBeckMŠamajJMicrotubules and mitogen-activated protein kinase signallingCurr Opin Plant Biol201114665065721839668
- YangYBianGLuQNeuroprotection and neurotrophism effects of liquiritin on primary cultured hippocampal cellsJ Chin Mater Med2008338931935
- WangZZhangXWangHQiLLouYNeuroprotective effects of icaritin against beta amyloid-induced neurotoxicity in primary cultured rat neuronal cells via estrogen-dependent pathwayNeuroscience2007145391192217321691
- MoghbelinejadSNassiri-AslMFarivarTNRutin activates the MAPK pathway and BDNF gene expression on beta-amyloid induced neurotoxicity in ratsToxicol Lett2014224110811324148604
- LinL-FChiuS-PWuM-JChenP-YYenJ-HLuteolin induces microRNA-132 expression and modulates neurite outgrowth in PC12 cellsPloS One201278e4330422916239
- KanoYHorieNDoiSArtepillin C derived from propolis induces neurite outgrowth in PC12m3 cells via ERK and p38 MAPK pathwaysNeurochem Res20083391795180318338254
- SagaraYVanhnasyJMaherPInduction of PC12 cell differentiation by flavonoids is dependent upon extracellular signal-regulated kinase activationJ Neurochem20049051144115515312169
- ZhangZCaiLZhouXMethyl 3, 4-dihydroxybenzoate promote rat cortical neurons survival and neurite outgrowth through the adenosine A2a receptor/PI3K/Akt signaling pathwayNeuroReport201526636737325807175
- LonzeBEGintyDDFunction and regulation of CREB family transcription factors in the nervous systemNeuron200235460562312194863
- BossVRobackJDYoungANNerve growth factor, but not epidermal growth factor, increases Fra-2 expression and alters Fra-2/JunD binding to AP-1 and CREB binding elements in pheochromocytoma (PC12) cellsJ Neurosci2001211182611150315
- MantamadiotisTLembergerTBleckmannSCDisruption of CREB function in brain leads to neurodegenerationNat Genet2002311475411967539
- FurukawaYWatanabeSOkuyamaSNakajimaMNeurotrophic effect of Citrus auraptene: neuritogenic activity in PC12 cellsInt J Mol Sci20121355338534722754300
- NagaseHOmaeNOmoriANobiletin and its related flavonoids with CRE-dependent transcription-stimulating and neuritegenic activitiesBiochem Biophys Res Commun200533741330133616253614
- HwangS-LLinJ-AShihP-HYehC-TYenG-CPro-cellular survival and neuroprotection of citrus flavonoid: the actions of hesperetin in PC12 cellsFood Funct20123101082109022767158
- LiQZhaoHZhangZLong-term administration of green tea catechins prevents age-related spatial learning and memory decline in C57BL/6 J mice by regulating hippocampal cyclic amp-response element binding protein signaling cascadeNeuroscience200915941208121519409206
- OhSBParkHRJangYJChoiSYSonTGLeeJBaicalein attenuates impaired hippocampal neurogenesis and the neurocognitive deficits induced by γ-ray radiationBr J Pharmacol2013168242143122891631
- LeeBSurBShimILeeHHahmD-HBaicalin improves chronic corticosterone-induced learning and memory deficits via the enhancement of impaired hippocampal brain-derived neurotrophic factor and cAMP response element-binding protein expression in the ratJ Nat Med201468113214323749289
- ZhaoLWangJ-LLiuRLiX-XLiJ-FZhangLNeuroprotective, anti-amyloidogenic and neurotrophic effects of apigenin in an Alzheimer’s disease mouse modelMolecules20131889949996523966081
- LiuRLiJ-ZSongJ-KPinocembrin improves cognition and protects the neurovascular unit in Alzheimer related deficitsNeurobiol Aging20143561275128524468471
- JinCHShinEJParkJBFustin flavonoid attenuates β-amyloid (1–42)-induced learning impairmentJ Neurosci Res200987163658367019533734
- BasliASouletSChaherNWine polyphenols: potential agents in neuroprotectionOxid Med Cell Longev20122012
- SasoLFiruziOPharmacological applications of antioxidants: lights and shadowsCurr Drug Targets201415131177119925341421
- AmaraFBerbenniMFragniMNeuroprotection by cocktails of dietary antioxidants under conditions of nerve growth factor deprivationOxid Med Cell Longev2015201521725826236423
- PanickarKSAndersonRAEffect of polyphenols on oxidative stress and mitochondrial dysfunction in neuronal death and brain edema in cerebral ischemiaInt J Mol Sci201112118181820722174658
- Razzaghi-AslNGarridoJKhazraeiHBorgesFFiruziOAntioxidant properties of hydroxycinnamic acids: A review of structure-activity relationshipsCurr Med Chem201320364436445023834166
- WeinrebOAmitTYoudimMBA novel approach of proteomics and transcriptomics to study the mechanism of action of the antioxidant–iron chelator green tea polyphenol (−)-epigallocatechin-3-gallateFree Radic Biol Med200743454655617640565
- HarikumarKBAggarwalBBResveratrol: a multitargeted agent for age-associated chronic diseasesCell Cycle2008781020103518414053
- Campos-EsparzaMRSanchez-GomezMVMatuteCMolecular mechanisms of neuroprotection by two natural antioxidant polyphenolsCell Calcium200945435836819201465
- SelkoeDJClearing the brain’s amyloid cobwebsNeuron200132217718011683988
- VillemagneVLBurnhamSBourgeatPAmyloid β deposition, neurodegeneration, and cognitive decline in sporadic Alzheimer’s disease: a prospective cohort studyLancet Neurol201312435736723477989
- UshikuboHWatanabeSTanimotoY3,3′,4′,5, 5′-Pentahydroxyflavone is a potent inhibitor of amyloid β fibril formationNeurosci Lett20125131515622343025
- HongS-YJeongW-SJunMProtective effects of the key compounds isolated from Corni fructus against β-amyloid-induced neurotoxicity in PC12 cellsMolecules2012179108311084522964500
- HeoHJLeeCYEpicatechin and catechin in cocoa inhibit amyloid β protein induced apoptosisJ Agric Food Chem20055351445144815740021
- MengPYoshidaHTanjiKCarnosic acid attenuates apoptosis induced by amyloid-1–42 or 1–43 in SH-SY5Y human neuroblastoma cellsNeurosci Res2015941925510380
- MuXHeGChengYLiXXuBDuGBaicalein exerts neuroprotective effects in 6-hydroxydopamine-induced experimental parkinsonism in vivo and in vitroPharmacol Biochem Behav200992464264819327378
- TavakkoliMMiriRJassbiARCarthamus, Salvia and Stachys species protect neuronal cells against oxidative stress-induced apoptosisPharm Biol201452121550155725243877
- SajadMZarganJZargarMAQuercetin prevents protein nitration and glycolytic block of proliferation in hydrogen peroxide insulted cultured neuronal precursor cells (NPCs): Implications on CNS regenerationNeurotoxicology201336243323396185
- LiuBZhangHXuCNeuroprotective effects of icariin on corticosterone-induced apoptosis in primary cultured rat hippocampal neuronsBrain Res20111375596721182828
- KenslerTWWakabayashiNBiswalSCell survival responses to environmental stresses via the Keap1-Nrf2-ARE pathwayAnnu Rev Pharmacol Toxicol2007478911616968214
- ChungSWLiuXMaciasAABaronRMPerrellaMAHeme oxygenase-1–derived carbon monoxide enhances the host defense response to microbial sepsis in miceJ Clin Invest2008118123918060048
- ZhangDDMechanistic studies of the Nrf2-Keap1 signaling pathwayDrug Metab Rev200638476978917145701
- ZhangMAnCGaoYLeakRKChenJZhangFEmerging roles of Nrf2 and phase II antioxidant enzymes in neuroprotectionProg Neurobiol2013100304723025925
- ChoH-YJedlickaAEReddySPRole of NRF2 in protection against hyperoxic lung injury in miceAm J Respir Cell Mol Biol200226217518211804867
- RomeoLIntrieriMD’AgataVThe major green tea poly-phenol, (−)-epigallocatechin-3-gallate, induces heme oxygenase in rat neurons and acts as an effective neuroprotective agent against oxidative stressJ Am Coll Nutr200928sup4492S499S20234037
- KosakaKMimuraJItohKRole of Nrf2 and p62/ZIP in the neurite outgrowth by carnosic acid in PC12h cellsJ Biochem20101471738119762340
- RussellJMSodium-potassium-chloride cotransportPhysiol Rev200080121127610617769
- NakajimaK-IMiyazakiHNiisatoNMarunakaYEssential role of NKCC1 in NGF-induced neurite outgrowthBiochem Biophys Res Commun2007359360461017548052
- NakajimaK-INiisatoNMarunakaYQuercetin stimulates NGF-induced neurite outgrowth in PC12 cells via activation of Na+/K+/2Cl-cotransporterCell Physiol Biochem201128114715621865857
- DiógenesMJFernandesCCSebastiaoAMRibeiroJAActivation of adenosine A2A receptor facilitates brain-derived neurotrophic factor modulation of synaptic transmission in hippocampal slicesJ Neurosci200424122905291315044529
- TebanoMMartireAPotenzaRAdenosine A2A receptors are required for normal BDNF levels and BDNF-induced potentiation of synaptic transmission in the mouse hippocampusJ Neurochem2008104127928618005343
- WieseSJablonkaSHoltmannBAdenosine receptor A2A-R contributes to motoneuron survival by transactivating the tyrosine kinase receptor TrkBProc Natl Acad Sci U S A200710443172101721517940030
- ChengH-CShihH-MChernYEssential role of cAMP-response element-binding protein activation by A2A adenosine receptors in rescuing the nerve growth factor-induced neurite outgrowth impaired by blockage of the MAPK cascadeJ Biol Chem200227737339303394212114502
- FlajoletMWangZFutterMFGF acts as a co-transmitter through adenosine A2A receptor to regulate synaptic plasticityNat Neurosci200811121402140918953346
- PalazzoloGHorvathPZenobi-WongMThe flavonoid isoquercitrin promotes neurite elongation by reducing RhoA activityPLoS One2012711e4997923209630
- Bora-TatarGErdem-YurterHInvestigations of curcumin and resveratrol on neurite outgrowth: perspectives on spinal muscular atrophyBiomed Res Int2014201470910825105137
- DasguptaBMilbrandtJResveratrol stimulates AMP kinase activity in neuronsProc Natl Acad Sci U S A2007104177217722217438283
- LiMTsangKSChoiSTLiKShawPCLauKFNeuronal differentiation of C17. 2 neural stem cells induced by a natural flavonoid, baicalinChembiochem201112344945621290546
- WooKWKwonOWKimSYPhenolic derivatives from the rhizomes of Dioscorea nipponica and their anti-neuroinflammatory and neuroprotective activitiesJ Ethnopharmacol201415521164117024973689
- GuanSZhangX-LGeDLiuT-QMaX-HCuiZ-FProtocatechuic acid promotes the neuronal differentiation and facilitates survival of phenotypes differentiated from cultured neural stem and progenitor cellsEur J Pharmacol2011670247147821946114
- LinC-YNiC-CYinM-CLiiC-KFlavonoids protect pancreatic beta-cells from cytokines mediated apoptosis through the activation of PI3-kinase pathwayCytokine2012591657122579112
- El OmriAHanJYamadaPKawadaKAbdrabbahMBIsodaHRosmarinus officinalis polyphenols activate cholinergic activities in PC12 cells through phosphorylation of ERK1/2J Ethnopharmacol2010131245145820633629
- NishinaAKimuraHTsukagoshiHNeurite outgrowth of PC12 cells by 4′-O-β-d-glucopyranosyl-3′,4-dimethoxychalcone from Brassica rapa L. ‘hidabeni’ was enhanced by pretreatment with p38MAPK inhibitorNeurochem Res201338112397240724057400
- ZeniALBZomkowskiADEMaraschinMRodriguesALSTascaCIInvolvement of PKA, CaMKII, PKC, MAPK/ERK and PI3K in the acute antidepressant-like effect of ferulic acid in the tail suspension testPharmacol Biochem Behav2012103218118622960128
- MaherPAkaishiTAbeKFlavonoid fisetin promotes ERK-dependent long-term potentiation and enhances memoryProc Natl Acad Sci U S A200610344165681657317050681
- MendoncaLMda Silva MachadoCTeixeiraCCCde FreitasLAPBianchiMdLPAntunesLMGCurcumin reduces cisplatin-induced neurotoxicity in NGF-differentiated PC12 cellsNeurotoxicology20133420521123036615
- LevitesYAmitTMandelSYoudimMBNeuroprotection and neurorescue against Aβ toxicity and PKC-dependent release of nonamyloidogenic soluble precursor protein by green tea polyphenol (−)-epigallocatechin-3-gallateFASEB J200317895295412670874
- LevitesYAmitTYoudimMBMandelSInvolvement of protein kinase C activation and cell survival/cell cycle genes in green tea polyphenol (−)-epigallocatechin 3-gallate neuroprotective actionJ Biol Chem200227734305743058012058035