
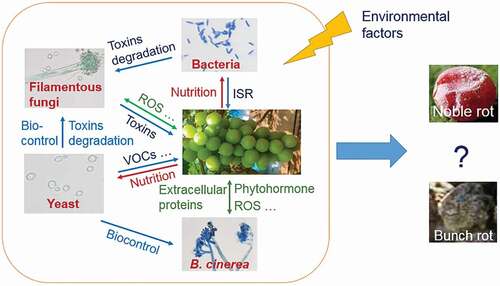
ABSTRACT
Botrytized wines are dessert wines made from overripe grapes infected by Botrytis cinerea known as “botrytized grapes” or “noble rot”. They are favored by human for their health promoting antioxidants and pleasant flavor. However, their production is low as natural botrytized grapes are rare in most vineyards worldwide. That is, not all B. cinerea infected grapes become botrytized grapes. The formation of botrytized grapes is determined by the interactions between grapes and the microbiota, including B. cinerea, under specific environmental conditions. Similarly, the interactions among the microorganisms in the grape must during the fermentation processes are also crucial for the quality of botrytized wines. Therefore, unraveling the roles of microbiota in the formation of botrytized grapes and wines can contribute to a better understanding of the determinants in this process, and provide a theoretical basis for improving the quality of botrytized wines.
GraphicalAbstract
Los vinos botritizados son vinos de postre elaborados con uvas sobremaduradas infectadas por Botrytis cinerea; se las conoce como “uvas botritizadas” o “podredumbre noble”. Dichos vinos son muy apreciados por sus antioxidantes, que favorecen la salud, así como por su agradable sabor. Sin embargo, su producción es limitada, ya que en la mayoría de los viñedos del mundo las uvas botritizadas naturales son escasas. Esto significa que no todas las uvas infectadas por B. cinerea se convierten en uvas botritizadas. Su formación es determinada por las interacciones entre las uvas y la microbiota, incluida B. cinerea, en condiciones ambientales específicas. Del mismo modo, para asegurar la calidad de los vinos botritizados son cruciales las interacciones entre los microorganismos del mosto de uva durante los procesos de fermentación. Por lo tanto, profundizar en la función que cumple la microbiota en la formación de uvas y vinos botritizados puede contribuir a comprender mejor los factores determinantes de este proceso, y proporcionar una base teórica que permita mejorar la calidad de los vinos botritizados.
1. Introduction
Botrytized wine (also known as noble wine) is a type of natural high-value dessert wine made from botrytized grapes. It is rich in antioxidants and aromatic compounds. The polyphenolic antioxidants have numerous health benefits (Thakur, Citation2018), while the complex and abundant fruity aroma, including a honey-like taste further contribute to its uniqueness (Magyar, Citation2011). However, the production of botrytized wine is extremely low owing to limited raw materials which is called “noble rot” or “botrytized grapes”. Botrytized grapes refer to overripe grapes infected by Botrytis cinerea, an ascomycetous fungus invading more than 1000 plant species (Weiberg et al., Citation2013). Not all grapes infected by B. cinerea become botrytized grapes. In fact, B. cinerea infected grape berries usually turn out to be bunch rot (grey mold), a fungal disease resulting in the rotting of grapes in most cases (Williamson et al., Citation2007). Only under special environmental conditions like sunny dry days and cold humid nights, botrytized grapes may be formed (Fournier et al., Citation2013). During this process, both B. cinerea and other microorganisms play important roles. As B. cinerea infects, the grape skin cracks and water evaporates, causing shriveling of the berry and an increased concentration of glucose, acids and other metabolites (Lovato et al., Citation2019). Simultaneously, fungal infection results in the perforation of grape epidermis, making grape nutrients accessible to other microorganisms on the fruit surface. Nutrient utilization by various microorganisms such as yeasts and bacteria results in the natural fermentation of nutrients in grape berries, and the generation of rich and complex flavors (Koenig et al., Citation2009). During winemaking, wine flavor is primarily determined by Saccharomyces cerevisiae. However, other microorganisms such as non-Saccharomyces yeast and bacteria may actively contribute to the unique flavor of wine (Eugenia Rodriguez et al., Citation2010). Thus, the difference in flavor may be closely related to the local microbial species and their interactions both on grape berries and during fermentation.
Although many aspects like the roles of B. cinerea and the diversity of microbial species in botrytized wine have recently been reviewed by experienced researchers (Magyar, Citation2011; Magyar & Soós, Citation2016; Thakur, Citation2018), the multivariate interactions among different microorganisms on grape berries or during fermentation need better understanding. While summarizing the species and roles of microbiota contributing to the formation of botrytized grapes and wines, this review aims to give a view of the interactions between microbes and grapes as well as interactions among microbial species in the formation of botrytized grapes and wines.
2. B. cinerea is essential in the formation of noble rot
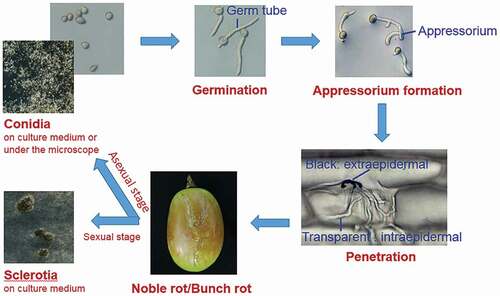
B. cinerea belongs to the Botrytis spp., which also includes other species such as Botrytis pseudocinerea. However, only B. cinerea proved to be essential for the formation of noble rot (Fournier et al., Citation2013). Up to 164 genotypes of B. cinerea had been isolated from both bunch rot and noble rot, indicating that both these may be caused by B. cinerea (Fournier et al., Citation2013). The formation of bunch rot or noble rot depends primarily on the interaction between B. cinerea and grape berries, and the environment plays an essential role in these interactions. B. cinerea is an important filamentous fungus that infects various plants. In its life cycle, there are sexual and asexual stages (). Under harsh environmental conditions, B. cinerea can survive by producing sclerotia, black sexual spores. When the environment is suitable, a large number of conidia are produced (Schumacher, Citation2016). Conidia are asexual spores, which are easily spread by wind, insects, and other media (Williamson et al., Citation2007). Conidia that land on the plant surface germinate under suitable conditions to produce germ tubes. These germ tubes elongate and form appressoria at their tips when suitable invasion sites are encountered (Gourgues et al., Citation2004). An appressorium is an important organ that has the ability to infect host plants with the help of extracellular hydrolases (Williamson et al., Citation2007). Following infection, the disease spot may form and expand at the infection site, or the infection may be limited by the host plant (Segmueller et al., Citation2008). Grape berries are more easily infected by B. cinerea during the ripening or storage stage (Haile et al., Citation2016). The infection of B. cinerea on grape berries plays a dual role. In a warm and humid environment, B. cinerea grow and propagate in large numbers as grey mold (bunch rot), which brings huge economic losses to the grape industry worldwide (Weiberg et al., Citation2013). Alternatively, on sunny dry days and cold humid nights, grapes infected by B. cinerea can shrink and form botrytized grapes (noble rot), which can be used as raw materials to produce botrytized wine (Fournier et al., Citation2013).
Figure 1. The life cycle of Botrytis cinerea.
Figura 1. El ciclo de vida de Botrytis cinerea.
2.1. B. cinerea-grape interaction for noble rot development
The interactions between B. cinerea and grape berries are the most important factors in botrytized grape formation. If B. cinerea win, bunch rot appears, otherwise latent infection or noble rot occurs (Haile et al., Citation2017). The interaction depends on the developmental stages of grape berries. Immature fruit secretes numerous tannins and other substances to limit the attack of B. cinerea. Overripe grapes are more likely to form noble rot (Magyar, Citation2011). During this stage, the interaction weapons between B. cinerea and grapes primarily involve reactive oxygen species (ROS), phytohormones, and extracellular proteins ().
Figure 2. The interactions between Botrytis cinerea and grape berries. ROS, reactive oxygen species.
Figura 2. Interacciones entre Botrytis cinerea y las bayas de uva. ERO, especies reactivas de oxígeno
ROS are widely used by plants as defense against invading phytopathogens at the initial stage of infection. The expression of ROS response genes in infected grapes were higher than that of the control grapes (Lovato et al., Citation2019). ROS generated at the infection site may induce programmed cell death (PCD) of surrounding cells, limiting nutrient absorption and expansion of the biotrophic fungus. However, this will not work for necrotrophic fungi such as B. cinerea, which can not only produce ROS scavenging enzyme to clear ROS, but also produce ROS (Li et al., Citation2018). For example, during the quiescent infection phase, the upregulation of ROS detoxification enzymes of B. cinerea were detected. During the infection stage, both ROS detoxification enzymes and ROS generating enzymes were upregulated in B. cinerea (Haile et al., Citation2019). NADPH oxidase (Nox) is an important ROS-generating enzyme in B. cinerea. It is a multi-subunit complex that primarily consists of NoxA, NoxB, and NoxR. Moreover, it has been shown that deletion mutants of the catalytic subunit NoxB in B. cinerea exhibited delayed infection, while the deletion mutant of catalytic subunit NoxA showed a defect in lesion expansion and pathogenicity after primary infection (Segmueller et al., Citation2008). Deleting the regulatory subunit NoxR damaged both primary infection and lesion expansion (Li et al., Citation2016). These results showed that ROS played an essential role in the interactions between B. cinerea and grape berries.
B. cinerea also facilitate its infection through interference of phytohormones such as ethylene, salicylate acid (SA) and jasmonate acid (JA). The pathogen can produce ethylene in response to light induction (Zhu et al., Citation2012). Ethylene participates in important physiological processes such as grape ripening, senescence, and stress. Fruit ripening is accompanied by the softening of the fruit structure and the exudation of carbohydrates (Cantu et al., Citation2008). The soft fruit structure is more susceptible to infection by B. cinerea and other microorganisms, and the exudation of nutrients plays an important role in the survival of microorganisms on the fruit surface (Koenig et al., Citation2009). Previous researches showed that B. cinerea infected fruit consists of more surface microorganisms than uninfected ones (Nisiotou & Nychas, Citation2007). Also, SA and JA play a role in resisting invasion of B. cinerea. For example, SA deficient Arabidopsis and tobacco plants were more resistant to Botrytis than wild-type plants, while JA deficient Arabidopsis plants were more sensitive (Rossi et al., Citation2011).
Extracellular metabolites are essential for B. cinerea to invade the grape cell. Although, appressoria are important to penetrate plant cell wall by physical means in many phytopathogenic fungi (Zhao et al., Citation2005), the appressorium of B. cinerea lacks membrane structure, which is not strong enough for penetration (Choquer et al., Citation2007). Thus, extracellular hydrolases such as endo-β-1,4-xylanases (Kars et al., Citation2005), that can degrade plant cell wall components, are extremely important for B. cinerea to breach cell walls. Various extracellular hydrolases with duplicate functions have been identified in B. cinerea (Li et al., Citation2020). Their abundance changed according to the extracellular pH and were regulated by the ROS generating system (B. Q. Li et al., Citation2012). Grapes also possess functional metabolites such as resveratrol. Resveratrol is a stilbenoid and can be converted into fungitoxic compounds by a specific laccase in B. cinerea which damages fungi (Schouten et al., Citation2010). Resveratrol is beneficial for human health because it is able to regulate cutaneous functions (Wen et al., Citation2020) and enhance cerebrovascular and cognitive functions (Wong et al., Citation2013). During noble rot formation, the resveratrol content was reported to be enriched (Negri et al., Citation2017), hence improving the functional component of botrytized wine.
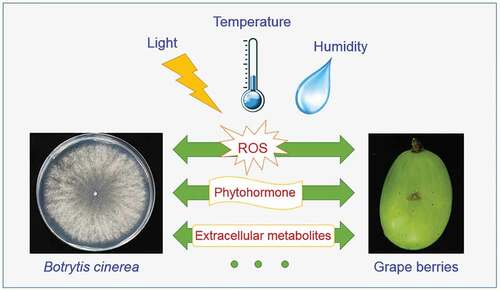
2.2. Environmental effects on B. cinerea-grape interaction
B. cinerea alters its own metabolism by responding to the environment, thereby affecting its interaction with the host grape. Through a detailed investigation of the factors causing grey mold or noble rot, Fournier et al. (Citation2013) proposed that the switch between them strongly depends on microclimate conditions. The microclimate includes light, temperature, and humidity conditions in the vineyard ().
B. cinerea possesses more than 11 photoreceptors from the long wavelength of far red to the short wavelength of near ultraviolet light (Schumacher, Citation2017). The transcription of these photoreceptors is controlled by the white collar complex (WCC) complex. Upon receiving light signals, the WCC complex promotes the transcription of different photoreceptors (Smith et al., Citation2010). Different photoreceptors transfer the signals to different signal pathways. In B. cinerea, deletion of BcSakA (a MAPK pathways protein) resulted in low expression of light transcription faction LTF2, which further led to the loss of the ability to produce conidia that would otherwise be produced under light (Schumacher, Citation2017). Altering the cAMP level also affects light-related phenotypes (Casas-Flores et al., Citation2006). Through signal pathways, the light signal finally transducts to functional proteins to regulate development, secondary metabolism, or stress response of fungi (Schumacher et al., Citation2014). The most evident developmental phenotype regulated by light is the generation of conidia or sclerotia. Conidia are asexual spores responsible for dispersal, and sclerotia are sexual spores primarily responsible for survival. The generation of conidia require light, whereas sclerotia are produced in the absence of light. More specifically, conidia generation occurs in the dark; however, it requires photoinduction (Schumacher, Citation2017).
Following conidia generation, temperature and humidity play an important role in its abjunction from sporophores, conidial germination, and appressorium infection. A rapid increase in temperature and a decrease in humidity in the morning would causes drying and twisting of conidiophores and abjunction of conidia (Williamson et al., Citation2007). Thereafter, the conidia may spread by air currents or insects (Fillinger & Elad, Citation2016). Conidia tend to germinate between 10°C and 30°C. The suitable germination temperature is 20°C; low or high temperature result in a delayed germination rate. In a dry environment, conidia have multiple germination sites and do not secrete extracellular substances. Under moist conditions, they exhibit only one or two germination sites and secrete extracellular substances continuously (Ciliberti et al., Citation2015). Extracellular substances may further facilitate the infection of the appressorium, which forms at the infection site of the germ tube of conidia. Normal infections were observed between 5°C and 30°C, with the highest infection incidence at 20°C. Apart from suitable temperature, short exposure to humidity is also a prerequisite infection by B. cinerea (Ciliberti et al., Citation2015).
2.3. Roles of B. cinerea in noble wine production
The genome of B. cinerea can be detected at all stages during fermentation of botrytized grapes. Its content decreases gradually from the beginning towards the end of fermentation. However, no viable B. cinerea can be recovered on yeast extract peptone dextrose plates during the mid-fermentation stage, which proves that the genome of B. cinerea detected at the mid and final fermentation stage are from dead cells (Lleixà et al., Citation2018). Thus, the influence of B. cinerea on botrytized wine is mainly on the formation of botrytized grapes and the early stage of fermentation.
The roles of B. cinerea on the fermentation of botrytized wines were previously reported. B. cinerea prefers to metabolize glucose in the grape, resulting in a high fructose/glucose ratio that adversely influences yeast growth and fermentation (Tronchoni et al., Citation2009). Moreover, botrytized grapes possess more polysaccharides of high molecular weight, which may cause problems during fermentation processes (Blanco-Ulate et al., Citation2015). Additionally, laccase secreted by B. cinerea can catalyze the oxidation of phenolic compounds into the corresponding quinones, which react further to dark-colored polymers to form brown pigments, a biomarker for B. cinerea infection (Ployon et al., Citation2020). B. cinerea was responsible for some unwanted wine off flavors such as earthy mushroom aroma (La Guerche et al., Citation2006). Alternatively, various beneficial substances are generated during the interaction among B. cinerea and grapes. Botrytized wines have lots of beneficial substances such as a higher content of fructose, organic acid, and minerals; and Tokaji Aszu´ wine was once thought to possess curative properties to various health conditions (Magyar, Citation2011). Besides, botrytized wines contain several antioxidants, such as resveratrol derivatives, polyphenols, and proanthocyanidins (Blanco-Ulate et al., Citation2015; Gindro et al., Citation2017). The level of some resveratrol derivatives in botrytised wine are reported to be higher than resveratrol following methoxylation, glycosylation, or oxidative oligomerization reactions catalyzed by the secretome of B. cinerea (Gindro et al., Citation2017). Botrytized wines are characterized by high polyphenol values as contributed by the biotransformation of stilbene, lignin, and anthocyanins by B. cinerea (Blanco-Ulate et al., Citation2015). Moreover, the noble wines are rich in pleasant flavors, including fruity, floral, and toasty aromas (Lopez Pinar et al., Citation2017).
3. The role of other filamentous fungi on grapes and botrytized wines
A variety of filamentous fungi have been detected on the surface of grape berries. The compositions of the fungal community are different according to grape varieties, regions, and the environment. The abundance of filamentous fungi on Malbec and Chardonnay grapes in Argentina and Brazil range from 1.3 × 104 to 5.4 × 106 CFU g−1 (Rosa et al., Citation2002). On Pinot Noir grapes from vineyards in Burgundy, the number was 6.2 × 103 to 2 × 106 CFU g−1 (Rousseaux et al., Citation2014). The mostly predominant genera of filamentous fungi identified include Botrytis, Penicillium, Aspergillus, Rhizopus, Plasmopara, and Uncinula (Jones & Mcmanus, Citation2017; Latorre et al., Citation2002; Lorenzini et al., Citation2016; Pithan et al., Citation2021). However, most filamentous are either pathogenic or secrete mycotoxins ().
Table 1. Mycotoxins secreted by filamentous fungi and the mycotoxin-degrading microorganisms.
Tabla 1. Micotoxinas secretadas por hongos filamentosos y microorganismos degradadores de micotoxinas
3.1. The interaction of filamentous fungi on the surface of grapes
Approximately 59 species of Penicillium have been isolated from grape vineyards globally (Lorenzini et al., Citation2016). The predominant species were based on grape varieties and location. For example, Penicillium expansum (P. expansum) is the most frequently isolated species in Portugal vineyards (Abrunhosa et al., Citation2001) and France vineyards (Bejaoui et al., Citation2006), while Penicillium chrysogenum is the most frequently isolated species from Argentinian vineyards (Magnoli et al., Citation2003). The Penicillium genus was reported to be the causal agent of green mold on mature berries (Z. Zhang et al., Citation2021). The infection of Penicillium altered the metabolic pathway of grape berries. Post-harvest proteomics of grapes showed that the infection of P. expansum had an impact on inducing energy metabolism and pathogenesis-related proteins (Lorenzini et al., Citation2016). Moreover, the infection of Penicillium crustosum induced various proteins related to redox homeostasis (Lorenzini et al., Citation2016). The interruption of redox homeostasis and the induction of hydrolases may play a synergistic role with B. cinerea to infect grape berries. In fact, P. expansum was proved to enhance the growth of B. cinerea in grape juice medium (Morales et al., Citation2013). Some Penicillium species such as P. expansum produce mycotoxins on grapes, such as patulin or citrinin (Abrunhosa et al., Citation2001). Patulin is a lactone molecule that is carcinogenic in animals. However, the accumulation patulin in grape juice medium can be prevented by B. cinerea (Morales et al., Citation2013).
Aspergillus spp. is the causal agent of black rot on grape berries (Ghuffar et al., Citation2020). Its virulence on grapes is less severe than that of B. cinerea or Penicillium spp. and can be controlled by good agricultural practices and fungicide application (Magista et al., Citation2021). The biggest threat of Aspergillus on the grape industry is the production of mycotoxins. Approximately 36 species of Aspergillus have been reported to be isolated from grape vineyards worldwide, with more than 16 species of these secreting Ochratoxin A (OTA) (Rousseaux et al., Citation2014). OTA is one of the primary mycotoxins discovered in human food, and has been reported to have nephrotoxic, hepatotoxic, and carcinogenic effect on animals (X. Huang et al., Citation2020). OTA has been detected in multiple grape producing areas in Portugal (Serra et al., Citation2006), France (Sage et al., Citation2002), Italy (Battilani et al., Citation2006), and China (X. Huang et al., Citation2020). For humans, wine is thought to be the second major source of OTA, just after cereals. Moreover, its maximum allowable amount was set by the European Commission to be 10 mg kg−1 in dried grapes and 2 mg kg−1 in wine (Regulation 123/2005). Fortunately, B. cinerea has the ability to degrade OTA in grape-like medium, leaving the botrytized grapes and wines with a low OTA content (Valero et al., Citation2008).
Rhizopus is a common fungus in the environment, which can be isolated from grape berries (Latorre et al., Citation2002). Rhizopus is usually used to saccharify starch to produce alcohol. It can produce fumaric acid, lactic acid, other organic acids, and aromatic esters.
Plasmopara viticola and Uncinula necator (syn. Erysiphe necator) are important grapevine fungal pathogens and causal agents of grape downy and powdery mildew, respectively (Jones & Mcmanus, Citation2017; Pithan et al., Citation2021). Resistance to P. viticola and U. necator in the grapevine is associated with the accumulation of compounds such as stilbenoids, which help to prevent the invasion of other fungi (Malacarne et al., Citation2011; Qiu et al., Citation2015). U. necator can also incur PCD-mediated resistance at the invasion site of the grapevine (Qiu et al., Citation2015). PCD-mediated resistance limits the infection of other biotrophic fungi, and contributes to the propagation of necrotrophic fungi such as B. cinerea. Moreover, during pathogen-grapevine interactions, both P. viticola and U. necator produce substances with an odor such as (Z)-1,5-octadien-3-one (Darriet et al., Citation2002; A. Pons et al., Citation2018).
3.2. The role of filamentous fungi on the fermentation of botrytized wine
Filamentous fungi account for a large proportion of the first stage of fermentation. The main filamentous fungi in the fermentation broth of healthy grapes are Rhizopus, whereas those in the fermentation broth of botrytized grapes are B. cinerea. Fusarium was only detected in the fermentation broth of healthy grapes, and Penicillium was greater in the fermentation broth of botrytized grapes than in healthy grapes. With the progress of fermentation, the proportion of filamentous fungi decreases, and the content of yeast significantly increases. At the late stage of fermentation, filamentous fungi could not be detected (Lleixà et al., Citation2018).
During fermentation, the bad odor and toxins generated by filamentous fungi can affect the quality of botrytized wines. The bad odor such as a moldy aroma is a major organoleptic defect of wines, which usually comes from some low-level aromatic compounds associated with the activity of filamentous fungi (Fontana, Citation2012; Siegmund & PoLlinger-Zierler, Citation2006). The compounds 1-octen-3-one, fenchol, and fenchone are frequently detected in grape wines and juices contaminated with filamentous fungi (La Guerche et al., Citation2006). The compound 1-octen-3-one was reported to originate from Penicillium brevicompactum and U. necator, whereas fenchol originated from Penicillium spp., Aspergillus spp., or Rhizopus nigricans (Darriet et al., Citation2002; La Guerche et al., Citation2006; M. Pons et al., Citation2011). However, mycotoxins, such as OTA produced from various fungi () are more prevalent in wines, and others like patulin can be degraded during fermentation (Abrunhosa et al., Citation2001).
4. Yeast in botrytized grapes and wines
Yeast is the most common microorganism isolated from the surface of the grape. The abundance and species of yeast are dynamic and closely related to the region and the nutritional status of the grape surface. Approximately 17 yeast species from eight genera were detected in four representative grape-growing areas in China. The dominant species were Hanseniaspora uvarum and Callista flavescens (S. S. Li et al., Citation2010). Moreover, 14 species from nine yeast genera were identified in Slovakia. Among them, H. uvarum and Metschnikowia pulcherrima were the most common species (Kántor et al., Citation2017). Yeasts with low nutritional requirements like Cryptococcus spp., Rhodotorula spp., and Sporobolomyces spp. are the most isolated species prior to veraison. With the ripening and softening of grapes, cracks appear on the intact surface and nutrition becomes more abundant, promoting the increase of aerobic and weak fermentative strains like Candida spp., Hanseniaspora spp., and Metschnikowia spp. When the surface of the grape is destroyed by microorganisms, high fermenting yeast groups such as Zygoascus hellenicus, Zygosaccharomyces spp., Torulaspora spp. and S. cerevisiae appear on the surface of grape berries (Barata et al., Citation2012).
The yeast population on botrytized grapes is three orders of magnitude higher than that on healthy grapes. Upon B. cinerea invasion, grape berries are usually colonized by yeasts whose activities modify the composition of grape juice. Pre-harvest grape yeasts are particularly rich in non-Saccharomyces, which forms the starting microflora of the fermenting grape must (Sipiczki, Citation2019).
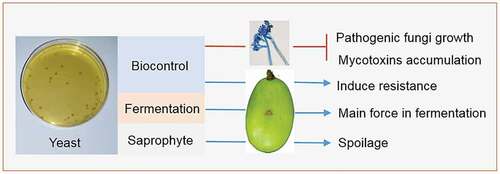
4.1. Yeasts antagonize other microorganism on grape berries
Yeast on the surface of grapes can be divided into S. cerevisiae and non-Saccharomyces. Both species play essential roles in botrytized grapes and wine formation (). S. cerevisiae primarily uses the nutrients on the surface of grapes, and is rarely detected on intact grapes (Kántor et al., Citation2017). Non-Saccharomyces includes some saprophytic and antagonistic yeasts. Antagonistic yeasts affect botrytized grape formation by inhibiting the growth of pathogens and inducing resistance of host cells.
Figure 3. Roles of yeast in the formation of botrytized grapes and wine.
Figura 3. Funciones de la levadura en la formación de uvas y vinos botritizados
Antagonistic yeasts control proliferation of pathogens through nutrition/space competition and gene modulation. For example, exogenous addition of trehalose showed enhanced biocontrol function of H. uvarum on grape berry rot caused by Aspergillus tubingensis and Penicillium commune (Apaliya et al., Citation2018). Pichia spp. compete for space and limit the growth of other microorganisms by film formation (Cabral et al., Citation2014). Metschnikowia species can modulate the genes in ethylene-associated immune responses in B. cinerea, resulting in a reduced virulence of this fungus on fruits (Tang et al., Citation2019). Moreover, Hanseniaspora osmophila in dual culture with Gluconobacter cerinus reported to inhibit B. cinerea, hence bunch rot and summer gray rot (Olivera et al., Citation2021).
Volatile organic compounds are also utilized by biocontrol yeast to combat other microorganisms. For example, volatile organic compounds generated by Aureobasidium pullulans function in reducing the growth rates and infection of B. cinerea, P. expansum, P. italicum, Colletotrichum acutatum, and Penicillium digitatum (Di Francesco et al., Citation2015). The compounds 2-ethyl-1-hexanol, 1,3,5,7-cyclooctatetraene, 3-methyl-1-butanol, 2-nonanone, and phenylethyl alcohol from Candida spp. were produced during the interaction between yeast and B. cinerea, and 2-ethyl-1-hexanol proved to inhibit the mycelial growth and spore germination of B. cinerea (Huang et al., Citation2012, Citation2011).
Finally, biocontrol yeast also induces the resistance of host cells, intervening in the interaction between pathogen and host cells, and secreting favorable flavor substances. Yeasts such as S. cerevisiae, Candida oleophila, and Metschnikowia species can induce a systemic immune response against various pathogens on the grape surface (Angelini et al., Citation2019; Droby et al., Citation2002; Hershkovitz et al., Citation2012).
4.2. Yeasts as the main force in botrytized wine fermentation
Undoubtedly, yeasts play an important role in wine fermentation. The diversity of yeast in the fermentation broth of botrytized grapes are reported to be higher than that of healthy grapes (Nisiotou et al., Citation2007). In the early and middle stage of fermentation, the fermentation broth of healthy grapes is primarily composed of H. uvarum, while the fermentation broth of botrytized grapes is composed of H. uvarum and Hanseniaspora osmophila (Lleixà et al., Citation2018). In addition, Lleixà et al. (Citation2018) detected Zygosasaccharomyces bailii, Issatchenkia spp., Kluyveromyces dobzhanskii, and Kazachstania spp. However, in the later stage of fermentation, S. cerevisiae gradually became the dominant strain (Lleixà et al., Citation2018).
During fermentation, yeast plays various roles to produce wine of desirable characteristics. First, fermentation is carried out in an environment of high sugar and increasing ethanol. The high sugar and ethanol tolerance of yeasts ensure completion of the fermentation process. For example, the existence of H. uvarum plays an important role in the fermentation of polysaccharides (Antunovics et al., Citation2005). S. cerevisiae and S. paradoxus have high sugar and strong ethanol tolerance, which plays an important role in the later stage of fermentation (Williams et al., Citation2015), whereas Candida apicola and Candida zemplinina are capable of growing in 14% ethanol (Tofalo et al., Citation2009). Secondly, yeasts contribute to the flavor and functional polyphenol formation. Terpenes such as linalool, citronellol, and linalool oxide are components that contribute to wine flavor and aroma profile (Yang et al., Citation2020). Co-fermentation of grape must with S. cerevisiae and glucosidase active non-Saccharomyces such as Issatchenkia terricola, Pichia kudriavzevii, or M. pulcherrima produced wine with higher terpene concentration (W. Zhang et al., Citation2020). Moreover, fermentation of grape must with proteolytic active yeasts contribute to increased amino acids and assimilable nitrogen in fermenting wine and facilitate extraction of polyphenols from grapes (Gaspar et al., Citation2019). Thirdly, yeast can antagonize filamentous fungi. It is reported that S. cerevisiae can antagonize the growth of Alternaria alternate (Prendes et al., Citation2021), B. cinerea (Lemos Junior & Nadai, Citation2016), Aspergillus carbonarius and A. niger (Ponsone et al., Citation2011). In addition, yeasts can degrade mycotoxins and antagonize a variety of toxins produced by Alternaria (Prendes et al., Citation2021) and the carcinogen OTA (Ponsone et al., Citation2011). Therefore, yeast plays an important role in improving the safety of botrytized grape wines.
5. The roles of bacteria in botrytized wine formation
Similar to yeast, species of bacteria are associated with the location and ripening stage of grape berries. A total of 44 species of bacteria from 21 genera were isolated from the wine producing area of Libourne in southwest France by medium culture and 16S rRNA sequencing (Martins et al., Citation2012). Approximately 51 bacterial species belonging to 22 genera were identified from Slovakian, Nitra, and Tokaj wine regions, among which Pseudomonas spp. was the most frequent, followed by Micrococcus spp. (Kántor et al., Citation2017). Bacteria from 16 bacterial phyla and 36 genera were identified in Xinjiang, China, and the dominant species were proteobacteria (Gao et al., Citation2019). Although there are apparent differences in bacterial distribution on different grape cultivar from different areas, Pantoea, Pseudomor, and Buchnera spp. were commonly abundant. Collectively, bacteria play both positive and negative roles in the formation of botrytized grapes and noble wines ().
Figure 4. Roles of bacteria in the formation of botrytized grapes and wines.
Figura 4. Funciones de la bacteria en la formación de uvas y vinos botritizados
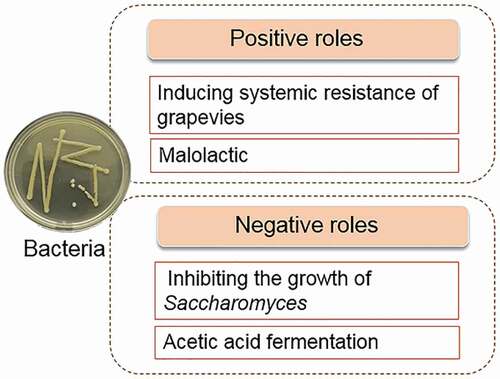
5.1. Bacteria help grapes to inhibit pathogenic fungi
Some bacteria proved to play roles in inducing systemic resistance (ISR) for grapevines to combat invading pathogens. ISR refers to the activation of the host defense system prior to pathogen attack, leading to improved resistance in the whole plant upon pathogen attack (B. W. M. Verhagen et al., Citation2010). Various bacteria showing ISR against other pathogens have been isolated from different parts of the grapevine. Pseudomonas fluorescens PTA-CT2, Pantoea agglomerans PTA-AF1, Acinetobacter lwoJJii AI-113, Nonas fluorescens PfCT2, and Burkholderia phytofirmans PsJN isolated from the rhizosphere or phyllosphere, all demonstrated protection on the grapevine against B. cinerea by ISR (Trotel-Aziz et al., Citation2008; B. W. M. Verhagen et al., Citation2010). Alternatively, antagonistic activities to protect grapevine against B. cinerea (Trotel-Aziz et al., Citation2008) by A. lwoJJii AI-113 and Nonas fluorescens PfCT2 induced oxidative burst and enhanced production of phytoalexins such as transresveratrol and trans-e-viniferin in the grapevine (B. Verhagen et al., Citation2011). B. phytofirmans PsJN forms a biofilm around the mycelium of B. cinerea to restrict it, induces oxidative bursts, and modulates changes of carbohydrate metabolism in grapevine leaves to resist B. cinerea. The existence of P. fluorescens PTA-CT2 strengthened the resistance of grapevines to both B. cinerea and P. viticola, which causes downy mildew in grapes. P. fluorescens mediated ISR against P. viticola through the SA signaling pathways and phytoalexin synthesis; however not through the JA and ET pathways. While ISR against B. cinerea mediated by P. viticola in grapevine primarily depended on improving the JA and ET signaling pathways, weakening hypersensitive cell death and inducing the accumulation of stilbenoids such as resveratrol, ε-viniferin, and δ-viniferin (Lakkis et al., Citation2019). Moreover, in an in vitro study, Lorenzini and Zapparoli (Citation2020) showed that Bacillus spp. isolated from withered grapes displayed an antagonistic effect on grape rotting fungi such as B. cinerea, P. expansum, and Aspergillus uvarum.
5.2. Bacteria participate in the fermentation of botrytized wines
Various bacteria participate in the process of wine fermentation, and 96 genera of bacteria were identified in wine samples by Piao et al. (Citation2015). During fermentation, bacteria first increased and then decreased, indicating that bacteria are active in this process. However, bacteria identified by Nicholas in botrytized wine were essentially unchanged (Bokulich et al., Citation2012). The most abundant bacteria in botrytized wine made in 2008–2010 from Dolce Winery was Rhodospirillales, and the second abundant bacteria were Lactobacillales (Bokulich et al., Citation2012). Moreover, Bokulich et al. (Citation2012) revealed that the content of bacteria in the botrytized wine fermentation were different from that in the fermentation broth of healthy grapes.
Bacteria are not the primary driving force of fermentation; however, they play an important role in fermentation. Lactic acid bacteria such as Oenococcus oeni contribute to malolactic fermentation and wine sensory quality (Piao et al., Citation2015). Acetic acid bacteria such as Acetobacter spp. and Gluconobacter spp. cause wine spoilage by generating undesirable volatile compounds such as acetic acid (Bartowsky, Citation2009). Moreover, Gluconobacter can inhibit the growth of Saccharomyces (Bokulich et al., Citation2012).
6. Conclusions
The formation of botrytized grapes is a complex process, during which both the microbial community and climate play important roles. The microorganisms primarily involved include filamentous fungi, yeast, and bacteria. B. cinerea is the most important filamentous fungi in the formation of botrytized grapes. Environmental factors such as light, temperature, and humidity are important in regulating formation of noble rot. Sunny dry days and cold humid nights are inclined to generate botrytized grapes, whereas heavy rainfall and high humidity tend to form bunch rot, a serious disease on grapes. The infection of B. cinerea considerably contributes to increased microbial content on grape berries, perhaps due to nutrition leakage during infection. Filamentous fungi such as Penicillium and Aspergillus are pathogenic fungi that secrete toxins and off-flavor substances on grapes. However, B. cinerea and some yeasts have the ability to degrade these toxins. Overall, microbiota involved in formation of botrytized grapes and fermentation can generate, increase, or modify various beneficial aromatic compound characteristics of noble wine. In addition, bacteria were also reported to induce the ISR of grapevines and influence the flavor of botrytized wine in the fermentation process. Therefore, the revealed roles of microorganisms contribute to improve our knowledge on the formation of botrytized grapes and wines, which may provide cues in optimizing the quality of fruit wines.
Disclosure statement
No potential conflict of interest was reported by the authors.
Additional information
Funding
References
- Abrunhosa, L., Ines, A., Rodrigues, A. I., Guimaraes, A., Pereira, V. L., Parpot, P., Mendes-Faia, A., & Venancio, A. (2014). Biodegradation of ochratoxin A by Pediococcus parvulus isolated from Douro wines. International Journal of Food Microbiology, 188, 45–52. https://doi.org/https://doi.org/10.1016/j.ijfoodmicro.2014.07.019
- Abrunhosa, L., Paterson, R. R. M., Kozakiewicz, Z., Lima, N., & Venancio, A. (2001). Mycotoxin production from fungi isolated from grapes. Letters in Applied Microbiology, 32(4), 240–242. https://doi.org/https://doi.org/10.1046/j.1472-765X.2001.00897.x
- Angelini, R. M. D. M., Rotolo, C., Gerin, D., Abate, D., Pollastro, S., & Faretra, F. (2019). Global transcriptome analysis and differentially expressed genes in grapevine after application of the yeast-derived defense inducer cerevisane. Pest Management Science, 75(7), 2020–2033. https://doi.org/https://doi.org/10.1002/ps.5317
- Antunovics, Z., Irinyi, L., & Sipiczki, M. (2005). Combined application of methods to taxonomic identification of Saccharomyces strains in fermenting botrytized grape must. Journal of Applied Microbiology, 98(4), 971–979. https://doi.org/https://doi.org/10.1111/j.1365-2672.2005.02543.x
- Apaliya, M. T., Zhang, H., Zheng, X., Yang, Q., Mahunu, G. K., & Kwaw, E. (2018). Exogenous trehalose enhanced the biocontrol efficacy of Hanseniaspora uvarum against grape berry rots caused by Aspergillus tubingensis and Penicillium commune. Journal of the Science of Food and Agriculture, 98(12), 4665–4672. https://doi.org/https://doi.org/10.1002/jsfa.8998
- Barata, A., Malfeito-Ferreira, M., & Loureiro, V. (2012). The microbial ecology of wine grape berries. International Journal of Food Microbiology, 153(3), 243–259. https://doi.org/https://doi.org/10.1016/j.ijfoodmicro.2011.11.025
- Bartowsky, E. J. (2009). Bacterial spoilage of wine and approaches to minimize it. Letters in Applied Microbiology, 48(2), 149–156. https://doi.org/https://doi.org/10.1111/j.1472-765X.2008.02505.x
- Battilani, P., Giorni, P., Bertuzzi, T., Formenti, S., & Pietri, A. (2006). Black aspergilli and ochratoxin A in grapes in Italy. International Journal of Food Microbiology, 111Suppl 1(4), S53–S60. https://doi.org/https://doi.org/10.1016/j.ijfoodmicro.2006.03.006
- Bejaoui, H., Mathieu, F., Taillandier, P., & Lebrihi, A. (2006). Black aspergilli and ochratoxin A production in French vineyards. International Journal of Food Microbiology, 111(2), S46–S52. https://doi.org/https://doi.org/10.1016/j.ijfoodmicro.2006.03.004
- Blanco-Ulate, B., Amrine, K. C., Collins, T. S., Rivero, R. M., Vicente, A. R., Morales-Cruz, A., Doyle, C. L., Ye, Z., Allen, G., Heymann, H., Ebeler, S. E., & Cantu, D. (2015). Developmental and metabolic plasticity of white-skinned grape berries in response to Botrytis cinerea during noble rot. Plant Physiology, 169(4), 2422–2443. https://doi.org/https://doi.org/10.1104/pp.15.00852
- Bokulich, N. A., Joseph, C. M., Allen, G., Benson, A. K., & Mills, D. A. (2012). Next-generation sequencing reveals significant bacterial diversity of botrytized wine. PloS One, 7(5), e36357. https://doi.org/https://doi.org/10.1371/journal.pone.0036357
- Bragulat, M. R., Abarca, M. L., & Cabaes, F. J. (2008). Low occurrence of patulin- and citrinin-producing species isolated from grapes. Letters in Applied Microbiology, 47(4), 286–289. https://doi.org/https://doi.org/10.1111/j.1472-765X.2008.02422.x
- Cabral, V., Znaidi, S., Walker, L. A., Martin-Yken, H., Dague, E., Legrand, M., Lee, K., Chauvel, M., Firon, A., Rossignol, T., Richard, M. L., Munro, C. A., Bachellier-Bassi, S., & d’Enfert, C. (2014). Targeted changes of the cell wall proteome influence Candida albicans ability to form single- and multi-strain biofilms. PLoS Pathogens, 10(12), e1004542. https://doi.org/https://doi.org/10.1371/journal.ppat.1004542
- Cantu, D., Vicente, A. R., Greve, L. C., Dewey, F. M., Bennett, A. B., Labavitch, J. M., & Powell, A. L. T. (2008). The intersection between cell wall disassembly, ripening, and fruit susceptibility to Botrytis cinerea. Proceedings of the National Academy of Sciences of the United States of America, 105(3), 859–864. https://doi.org/https://doi.org/10.1073/pnas.0709813105
- Casas-Flores, S., Rios-Momberg, M., Rosales-Saavedra, T., Martínez-Hernández, P., Olmedo-Monfil, V., & Herrera-Estrella, A. (2006). Cross talk between a fungal blue-light perception system and the cyclic amp signaling pathway. Eukaryotic Cell, 5(3), 499–506. https://doi.org/https://doi.org/10.1128/ec.5.3.499-506.2006
- Chen, Y. H., Sheu, S. C., Mau, J. L., & Hsieh, P. C. (2011). Isolation and characterization of a strain of Klebsiella pneumoniae with citrinin-degrading activity. World Journal of Microbiology & Biotechnology, 27(3), 487–493. https://doi.org/https://doi.org/10.1007/s11274-010-0478-4
- Choquer, M., Fournier, E., Kunz, C., Levis, C., Pradier, J.-M., Simon, A., & Viaud, M. (2007). Botrytis cinerea virulence factors: New insights into a necrotrophic and polyphageous pathogen. FEMS Microbiology Letters, 277(1), 1–10. https://doi.org/https://doi.org/10.1111/j.1574-6968.2007.00930.x
- Ciliberti, N., Fermaud, M., Languasco, L., & Rossi, V. (2015). Influence of fungal strain, temperature, and wetness duration on infection of grapevine inflorescences and young berry clusters by Botrytis cinerea. Phytopathology, 105(3), 325–333. https://doi.org/https://doi.org/10.1094/PHYTO-05-14-0152-R
- Darriet, P., Pons, M., Henry, R., Dumont, O., Findeling, V., Cartolaro, P., Calonnec, A., & Dubourdieu, D. (2002). Impact odorants contributing to the fungus type aroma from grape berries contaminated by powdery mildew (Uncinula necator); incidence of enzymatic activities of the yeast Saccharomyces cerevisiae. Journal of Agricultural and Food Chemistry, 50(11), 3277–3282. https://doi.org/https://doi.org/10.1021/jf011527d
- Di Francesco, A., Ugolini, L., Lazzeri, L., & Mari, M. (2015). Production of volatile organic compounds by Aureobasidium pullulans as a potential mechanism of action against postharvest fruit pathogens. Biological Control, 81, 8–14. https://doi.org/https://doi.org/10.1016/j.biocontrol.2014.10.004
- Díaz, G., Torres, R., Vega, M., & Latorre, B. A. (2009). Ochratoxigenic Aspergillus species on grapes from Chilean vineyards and Aspergillus threshold levels on grapes. International Journal of Food Microbiology, 133(1–2), 195–199. https://doi.org/https://doi.org/10.1016/j.ijfoodmicro.2009.04.018
- Diaz, G. A., Yanez, L., & Latorre, B. A. (2011). Low occurrence of patulin-producing strains of Penicillium in grapes and patulin degradation during winemaking in Chile. American Journal of Enology and Viticulture, 62(4), 542. https://doi.org/https://doi.org/10.5344/ajev.2011.11034
- Diaz, M. A., Pereyra, M. M., Santander, F. F. S., Perez, M. F., Cordoba, J. M., Alhussein, M., Karlovsky, P., & Dib, J. R. (2020). Protection of citrus fruits from postharvest infection with Penicillium digitatum and degradation of patulin by biocontrol yeast Clavispora lusitaniae 146. Microorganisms, 8(10), 1477. https://doi.org/https://doi.org/10.3390/microorganisms8101477
- Droby, S., Vinokur, V., Weiss, B., Cohen, L., Daus, A., Goldschmidt, E. E., & Porat, R. (2002). Induction of resistance to Penicillium digitatum in grapefruit by the yeast biocontrol agent Candida oleophila. Phytopathology, 92(4), 393–399. https://doi.org/https://doi.org/10.1094/phyto.2002.92.4.393
- Eugenia Rodriguez, M., Lopes, C. A., Barbagelata, R. J., Barda, N. B., & Caballero, A. C. (2010). Influence of Candida pulcherrima Patagonian strain on alcoholic fermentation behaviour and wine aroma. International Journal of Food Microbiology, 138(1–2), 19–25. https://doi.org/https://doi.org/10.1016/j.ijfoodmicro.2009.12.025
- Ferenczi, S., Cserhati, M., Krifaton, C., Szoboszlay, S., Kukolya, J., Szoke, Z., Koszegi, B., Albert, M., Barna, T., Mezes, M., Kovacs, K. J., & Kriszt, B. (2014). A new ochratoxin A biodegradation strategy using Cupriavidus basilensis Or16 strain. PloS One, 9(10), e109817. https://doi.org/https://doi.org/10.1371/journal.pone.0109817
- Fillinger, S., & Elad, Y. (2016). Botrytis: The fungus, the pathogen and its management in agricultural systems. Springer.
- Fontana, A. R. (2012). Analytical methods for determination of cork-taint compounds in wine. Trends in Analytical Chemistry, 37, 135–147. https://doi.org/https://doi.org/10.1016/j.trac.2012.03.012
- Fournier, E., Gladieux, P., & Giraud, T. (2013). The ‘Dr Jekyll and Mr Hyde fungus’: Noble rot versus gray mold symptoms of Botrytis cinerea on grapes. Evolutionary Applications, 6(6), 960–969. https://doi.org/https://doi.org/10.1111/eva.12079
- Fredj, S., & Chebil, S. (2009). Isolation and characterization of ochratoxin A and aflatoxin B1 producing fungi infecting grapevines cultivated in Tunisia. African Journal of Microbiology Research, 3(9), 523–527. https://doi.org/https://doi.org/10.3732/ajb.1200157
- Fuchs, E., Binder, E. M., Heidler, D., & Krska, R. (2002). Structural characterization of metabolites after the microbial degradation of type A trichothecenes by the bacterial strain BBSH 797. Food Additives and Contaminants, 19(4), 379–386. https://doi.org/https://doi.org/10.1080/02652030110091154
- Gao, F. F., Chen, J. L., Xiao, J., Cheng, W. D., Zheng, X. J., Wang, B., & Shi, X. W. (2019). Microbial community composition on grape surface controlled by geographical factors of different wine regions in Xinjiang, China. Food Research International, 122(AUG.), 348–360. https://doi.org/https://doi.org/10.1016/j.foodres.2019.04.029
- Gaspar, L. M., Machado, A., Coutinho, R., Sousa, S., Santos, R., Xavier, A., Figueiredo, M., Teixeira, M. D. F., Centeno, F., & Simões, J. (2019). Development of potential yeast protein extracts for red wine clarification and stabilization. Frontiers in Microbiology, 10, 2310. https://doi.org/https://doi.org/10.3389/fmicb.2019.02310
- Ghuffar, S., Ahmed, M. Z., Irshad, G., Zeshan, M. A., Qadir, A., Anwaar, H. A., Mansha, M. Z., Asadullah, H. M., Abdullah, A., & Farooq, U. (2020). First report of Aspergillus niger causing black rot of grapes in Pakistan. Plant Disease, 104, 1–3. https://doi.org/https://doi.org/10.1094/pdis-06-20-1390-pdn
- Gindro, K., Schnee, S., Righi, D., Marcourt, L., Nejad Ebrahimi, S., Codina, J. M., Voinesco, F., Michellod, E., Wolfender, J.-L., & Queiroz, E. F. (2017). Generation of antifungal stilbenes using the enzymatic secretome of Botrytis cinerea. Journal of Natural Products, 80(4), 887–898. https://doi.org/https://doi.org/10.1021/acs.jnatprod.6b00760
- Gourgues, M., Simon, A., Lebrun, M., & Levis, C. (2004). The tetraspanin BcPls1 is required for appressorium-mediated penetration of Botrytis cinerea into host plant leaves. Molecular Microbiology, 51(3), 619–629. https://doi.org/https://doi.org/10.1046/j.1365-2958.2003.03866.x
- Haile, Z. M., Malacarne, G., Pilati, S., Sonego, P., Moretto, M., Masuero, D., Vrhovsek, U., Engelen, K., Baraldi, E., & Moser, C. (2019). Dual transcriptome and metabolic analysis of Vitis vinifera cv. Pinot Noir Berry and Botrytis cinerea during quiescence and egressed infection. Frontiers in Plant Science, 10, 1704. https://doi.org/https://doi.org/10.3389/fpls.2019.01704
- Haile, Z. M., Pilati, S., Sonego, P., Malacarne, G., Vrhovsek, U., Engelen, K., Tudzynski, P., Zottini, M., Baraldi, E., & Moser, C. (2017). Molecular analysis of the early interaction between the grapevine flower and Botrytis cinerea reveals that prompt activation of specific host pathways leads to fungus quiescence. Plant, Cell & Environment, 40(8), 1409–1428. https://doi.org/https://doi.org/10.1111/pce.12937
- Haile, Z. M., Sonego, P., Engelen, K., Vrhovsek, U., Tudzynski, P., Baraldi, E., & Moser, C. (2016). Characterizing the interaction between Botrytis cinerea and grapevine inflorescences. In A. Ippolito, S. M. Sanzani, M. Wisniewski, & S. Droby (Eds.), Iii International Symposium on Postharvest Pathology: Using Science to Increase Food Availability (Vol.1144, pp. 29–35).
- Hershkovitz, V., Ben-Dayan, C., Raphael, G., Pasmanik-Chor, M., Liu, J., Belausov, E., Aly, R., Wisniewski, M., & Droby, S. (2012). Global changes in gene expression of grapefruit peel tissue in response to the yeast biocontrol agent Metschnikowia fructicola. Molecular Plant Pathology, 13(4), 338–349. https://doi.org/https://doi.org/10.1111/j.1364-3703.2011.00750.x
- Huang, R., Che, H. J., Zhang, J., Yang, L., Jiang, D. H., & Li, G. Q. (2012). Evaluation of Sporidiobolus pararoseus strain YCXT3 as biocontrol agent of Botrytis cinerea on post-harvest strawberry fruits. Biological Control, 62(1), 53–63. https://doi.org/https://doi.org/10.1016/j.biocontrol.2012.02.010
- Huang, R., Li, G. Q., Zhang, J., Yang, L., Che, H. J., Jiang, D. H., & Huang, H. C. (2011). Control of postharvest Botrytis fruit rot of strawberry by volatile organic compounds of Candida intermedia. Phytopathology, 101(7), 859–869. https://doi.org/https://doi.org/10.1094/phyto-09-10-0255
- Huang, X., Xiao, Z., Kong, F., Chen, A. J., Perrone, G., Wang, Z., Wang, J., & Zhang, H. (2020). Diversity and ochratoxin A-fumonisin profile of black Aspergilli isolated from grapes in China. World Mycotoxin Journal, 13(2), 225–233. https://doi.org/https://doi.org/10.3920/wmj2019.2505
- Jones, D. S., & Mcmanus, P. (2017). Susceptibility of cold-climate wine grape cultivars to downy mildew, powdery mildew, and black rot. Plant Disease, 101(7), 1077–1085. https://doi.org/https://doi.org/10.1094/PDIS-01-17-0022-RE
- Kanpiengjai, A., Mahawan, R., Lumyong, S., & Khanongnuch, C. (2016). A soil bacterium Rhizobium borbori and its potential for citrinin-degrading application. Annals of Microbiology, 66(2), 807–816. https://doi.org/https://doi.org/10.1007/s13213-015-1167-1
- Kántor, A., Mareček, J., Ivanišová, E., Terentjeva, M., & Kačániová, M. (2017). Microorganisms of grape berries. Proceedings of the Latvian Academy of Sciences. Section B. Natural, Exact, and Applied Sciences. National Academy of Sciences.
- Kars, I., Krooshof, G. H., Wagemakers, L., Joosten, R., Benen, J. A. E., & van Kan, J. A. L. (2005). Necrotizing activity of five Botrytis cinerea endopolygalacturonases produced in Pichia pastoris. The Plant Journal, 43(2), 213–225. https://doi.org/https://doi.org/10.1111/j.1365-313X.2005.02436.x
- Koenig, H., Unden, G., & Froehlich, J. (2009). Biology of microorganisms on grapes, in must and in wine. Springer.
- La Guerche, S., Dauphin, B., Pons, M., Blancard, D., & Darriet, P. (2006). Characterization of some mushroom and earthy off-odors microbially induced by the development of rot on grapes. Journal of Agricultural and Food Chemistry, 54(24), 9193–9200. https://doi.org/https://doi.org/10.1021/jf0615294
- Lakkis, S., Trotel-Aziz, P., Rabenoelina, F., Schwarzenberg, A., Nguema-Ona, E., Clement, C., & Aziz, A. (2019). Strengthening grapevine resistance by Pseudomonas fluorescens PTA-CT2 relies on distinct defense pathways in susceptible and partially resistant genotypes to downy mildew and gray mold diseases. Frontiers in Plant Science, 10, 1112. https://doi.org/https://doi.org/10.3389/fpls.2019.01112
- Latorre, B. A., Viertel, S. C., & Spadaro, I. (2002). Severe outbreaks of bunch rots caused by Rhizopus stolonifer and Aspergillus niger on table grapes in Chile. Plant Disease, 86(7), 815. https://doi.org/https://doi.org/10.1094/pdis.2002.86.7.815c
- Lemos Junior, W. J., & Nadai, C. (2016). Biocontrol ability and action mechanism of Starmerella bacillaris (synonym Candida zemplinina) isolated from wine musts against gray mold disease agent Botrytis cinerea on grape and their effects on alcoholic fermentation. Frontiers in Microbiology, 7, 1249. https://doi.org/https://doi.org/10.3389/fmicb.2016.01249
- Li, B. Q., Wang, W. H., Zong, Y. Y., Qin, G. Z., & Tian, S. P. (2012). Exploring pathogenic mechanisms of Botrytis cinerea secretome under different ambient pH based on comparative proteomic analysis. Journal of Proteome Research, 11(8), 4249–4260. https://doi.org/https://doi.org/10.1021/pr300365f
- Li, H., Chen, Y., Zhang, Z. Q., Li, B. Q., Qin, G. Z., & Tian, S. P. (2018). Pathogenic mechanisms and control strategies of Botrytis cinerea causing post-harvest decay in fruits and vegetables. Food Quality and Safety, 2(3), 111–119. https://doi.org/https://doi.org/10.1093/fqsafe/fyy016
- Li, H., Zhang, Z., Qin, G., He, C., Li, B., & Tian, S. (2020). Actin is required for cellular development and virulence of Botrytis cinerea via the mediation of secretory proteins. Msystems, 5(1), e00732–00719. https://doi.org/https://doi.org/10.1128/mSystems.00732-19
- Li, H., Zhang, Z. Q., He, C., Qin, G. Z., & Tian, S. P. (2016). Comparative proteomics reveals the potential targets of BcNoxR, a putative regulatory subunit of NADPH oxidase of Botrytis cinerea. Molecular Plant-Microbe Interactions, 29(12), 990–1003. https://doi.org/https://doi.org/10.1094/MPMI-11-16-0227-R
- Li, S. S., Cheng, C., Li, Z., Chen, J. Y., Yan, B., Han, B. Z., & Reeves, M. (2010). Yeast species associated with wine grapes in China. International Journal of Food Microbiology, 138(1–2), 85–90. https://doi.org/https://doi.org/10.1016/j.ijfoodmicro.2010.01.009
- Lleixà, J., Kioroglou, D., Mas, A., & del Carmen Portillo, M. (2018). Microbiome dynamics during spontaneous fermentations of sound grapes in comparison with sour rot and Botrytis infected grapes. International Journal of Food Microbiology, 281, 36–46. https://doi.org/https://doi.org/10.1016/j.ijfoodmicro.2018.05.016
- Lopez Pinar, A., Rauhut, D., Ruehl, E., & Buettner, A. (2017). Effects of bunch rot (Botrytis cinerea) and powdery mildew (Erysiphe necator) fungal diseases on wine aroma. Frontiers in Chemistry, 5, 20. https://doi.org/https://doi.org/10.3389/fchem.2017.00020
- Lorenzini, M., Mainente, F., Zapparoli, G., Cecconi, D., & Simonato, B. (2016). Post-harvest proteomics of grapes infected by Penicillium during withering to produce Amarone wine. Food Chemistry, 199, 639–647. https://doi.org/https://doi.org/10.1016/j.foodchem.2015.12.032
- Lorenzini, M., & Zapparoli, G. (2020). Epiphytic bacteria from withered grapes and their antagonistic effects on grape-rotting fungi. International Journal of Food Microbiology, 319, 108505. https://doi.org/https://doi.org/10.1016/j.ijfoodmicro.2019.108505
- Lovato, A., Zenoni, S., Tornielli, G. B., Colombo, T., Vandelle, E., & Polverari, A. (2019). Specific molecular interactions between Vitis vinifera and Botrytis cinerea are required for noble rot development in grape berries. Postharvest Biology and Technology, 156, 110924. https://doi.org/https://doi.org/10.1016/j.postharvbio.2019.05.025
- Magista, D., Cozzi, G., Gambacorta, L., Logrieco, A. F., Solfrizzo, M., & Perrone, G. (2021). Studies on the efficacy of electrolysed oxidising water to control Aspergillus carbonarius and ochratoxin A contamination on grape. International Journal of Food Microbiology, 338(2), 108996. https://doi.org/https://doi.org/10.1016/j.ijfoodmicro.2020.108996
- Magnoli, C., Astoreca, A., Ponsone, L., Combina, M., Palacio, G., Rosa, C. A. R., & Dalcero, A. M. (2010). Survey of mycoflora and ochratoxin A in dried vine fruits from Argentina markets. Letters in Applied Microbiology, 39(4), 326–331. https://doi.org/https://doi.org/10.1111/j.1472-765X.2004.01583.x
- Magnoli, C., Violante, M., Combina, M., Palacio, G., & Dalcero, A. (2003). Mycoflora and ochratoxin-producing strains of Aspergillus section Nigri in wine grapes in Argentina. Letters in Applied Microbiology, 37(2), 179–184. https://doi.org/https://doi.org/10.1046/j.1472-765X.2003.01376.x
- Magyar, I. (2011). Botrytized wines. Advances in Food and Nutrition Research, 63, 147–206. https://doi.org/https://doi.org/10.1016/B978-0-12-384927-4.00006-3
- Magyar, I., & Soós, J. (2016). Botrytized wines–current perspectives. International Journal of Wine Research, 8, 29–39. https://doi.org/https://doi.org/10.2147/ijwr.s100653
- Malacarne, G., Vrhovsek, U., Zulini, L., Cestaro, A., Stefanini, M., Mattivi, F., Delledonne, M., Velasco, R., & Moser, C. (2011). Resistance to Plasmopara viticola in a grapevine segregating population is associated with stilbenoid accumulation and with specific host transcriptional responses. BMC Plant Biology, 11(1), 114-114. https://doi.org/https://doi.org/10.1186/1471–2229-11-114
- Martins, G., Miot-Sertier, C., Lauga, B., Claisse, O., Lonvaud-Funel, A., Soulas, G., & Masneuf-Pomarède, I. (2012). Grape berry bacterial microbiota: Impact of the ripening process and the farming system. International Journal of Food Microbiology, 158(2), 93–100. https://doi.org/https://doi.org/10.1016/j.ijfoodmicro.2012.06.013
- Mikusova, P., Ritieni, A., Santini, J., & Srobarova, G. (2010). Contamination by moulds of grape berries in Slovakia. Food Additives & Contaminants. Part A, Chemistry, Analysis, Control, Exposure & Risk Assessment, 27(5), 738–747. https://doi.org/https://doi.org/10.1080/19440040903571754
- Mor, S., Puniya, A. K., & Singh, K. (2003). Degradation of preformed aflatoxins by detoxifying organisms. Indian Journal of Animal Sciences, 73(3), 315–318. https://doi.org/https://doi.org/10.1016/j.talanta.2014.03.053
- Morales, H., Paterson, R. R. M., Venancio, A., & Lima, N. (2013). Interaction with Penicillium expansum enhances Botrytis cinerea growth in grape juice medium and prevents patulin accumulation in vitro. Letters in Applied Microbiology, 56(5), 356–360. https://doi.org/https://doi.org/10.1111/lam.12056
- Negri, S., Lovato, A., Boscaini, F., Salvetti, E., Torriani, S., Commisso, M., Danzi, R., Ugliano, M., Polverari, A., Tornielli, G. B., & Guzzo, F. (2017). The induction of noble rot (Botrytis cinerea) infection during postharvest withering changes the metabolome of grapevine berries (vitis vinifera L., cv. garganega). Frontiers in Plant Science, 8, 1002. https://doi.org/https://doi.org/10.3389/fpls.2017.01002
- Nisiotou, A. A., & Nychas, G.-J. E. (2007). Yeast populations residing on healthy or Botrytis-infected grapes from a vineyard in Attica, Greece. Applied and Environmental Microbiology, 73(8), 2765–2768. https://doi.org/https://doi.org/10.1128/AEM.01864-06
- Nisiotou, A. A., Spiropoulos, A. E., & Nychas, G.-J. E. (2007). Yeast community structures and dynamics in healthy and Botrytis-affected grape must fermentations. Applied and Environmental Microbiology, 73(21), 6705–6713. https://doi.org/https://doi.org/10.1128/aem.01279-07
- Olivera, M., Delgado, N., Cádiz, F., Riquelme, N., Montenegro, I., Seeger, M., Bravo, G., Barros-Parada, W., Pedreschi, R., & Besoain, X. (2021). Diffusible compounds produced by Hanseniaspora osmophila and Gluconobacter cerinus help to control the causal agents of gray rot and summer bunch rot of table grapes. Antibiotics, 10(6), 664. https://doi.org/https://doi.org/10.3390/antibiotics10060664
- Patharajan, S., Reddy, K. R. N., Karthikeyan, V., Spadaro, D., Lore, A., Gullino, M. L., & Garibaldi, A. (2011). Potential of yeast antagonists on invitro biodegradation of ochratoxin A. Food Control, 22(2), 290–296. https://doi.org/https://doi.org/10.1016/j.foodcont.2010.07.024
- Piao, H., Hawley, E., Kopf, S., DeScenzo, R., Sealock, S., Henick-Kling, T., & Hess, M. (2015). Insights into the bacterial community and its temporal succession during the fermentation of wine grapes. Frontiers in Microbiology, 6, 809. https://doi.org/https://doi.org/10.3389/fmicb.2015.00809
- Pithan, P. A., Ducati, J. R., Garrido, L. R., Arruda, D. C., Thum, A. B., & Hoff, R. (2021). Spectral characterization of fungal diseases downy mildew, powdery mildew, black-foot and Petri disease on Vitis vinifera leaves. International Journal of Remote Sensing, 42(15), 5680–5697. https://doi.org/https://doi.org/10.1080/01431161.2021.1929542
- Ployon, S., Attina, A., Vialaret, J., Walker, A. S., Hirtz, C., & Saucier, C. (2020). Laccases 2 & 3 as biomarkers of Botrytis cinerea infection in sweet white wines. Food Chemistry, 315(2), 126233. https://doi.org/https://doi.org/10.1016/j.foodchem.2020.126233
- Pons, A., Mouakka, N., Deliere, L., Crachereau, J. C., Davidou, L., Sauris, P., Guilbault, P., & Darriet, P. (2018, January 15). Impact of Plasmopara viticola infection of Merlot and Cabernet Sauvignon grapes on wine composition and flavor. Food Chemistry, 239, 102. https://doi.org/https://doi.org/10.1016/j.foodchem.2017.06.087
- Pons, M., Dauphin, B., Guerche, S. L., Pons, A., Lavigne-Cruege, V. R., Shinkaruk, S., Bunner, D., Richard, T., Monti, J. P., & Darriet, P. (2011). Identification of impact odorants contributing to fresh mushroom off-flavor in wines: Incidence of their reactivity with nitrogen compounds on the decrease of the olfactory defect. Journal of Agricultural and Food Chemistry, 59(7), 3264–3272. https://doi.org/https://doi.org/10.1021/jf104215a
- Ponsone, M. L., Chiotta, M. L., Combina, M., Dalcero, A., & Chulze, S. (2011). Biocontrol as a strategy to reduce the impact of ochratoxin A and Aspergillus section Nigri in grapes. International Journal of Food Microbiology, 151(1), 70–77. https://doi.org/https://doi.org/10.1016/j.ijfoodmicro.2011.08.005
- Ponsone, M. L., Combina, M., Dalcero, A., & Chulze, S. (2007). Ochratoxin A and ochratoxigenic Aspergillus species in Argentinean wine grapes cultivated under organic and non-organic systems. International Journal of Food Microbiology, 114(2), 131–135. https://doi.org/https://doi.org/10.1016/j.ijfoodmicro.2006.07.001
- Prendes, L. P., Merin, M. G., Zachetti, V. G. L., Pereyra, A., Ramirez, M. L., & Morata de Ambrosini, V. I. (2021). Impact of antagonistic yeasts from wine grapes on growth and mycotoxin production by Alternaria alternata. Journal of Applied Microbiology, 131(2), 833–843. https://doi.org/https://doi.org/10.1111/jam.14996
- Qiu, W., Feechan, A., & Dry, I. (2015). Current understanding of grapevine defense mechanisms against the biotrophic fungus (Erysiphe necator), the causal agent of powdery mildew disease. Horticulture Research, 2(1), 15020. https://doi.org/https://doi.org/10.1038/hortres.2015.20
- Risa, A., Krifaton, C., Kukolya, J., Kriszt, B., Cserhati, M., & Tancsics, A. (2018). Aflatoxin B1 and zearalenone-detoxifying profile of Rhodococcus type strains. Current Microbiology, 75(7), 907–917. https://doi.org/https://doi.org/10.1007/s00284-018-1465-5
- Rosa, C. A. D., Palacios, V., Combina, M., Fraga, M. E., Rekson, A. D., Magnoli, C. E., & Dalcero, A. M. (2002). Potential ochratoxin A producers from wine grapes in Argentina and Brazil. Food Additives and Contaminants, 19(4), 408–414. https://doi.org/https://doi.org/10.1080/02652030110092748
- Rossi, F. R., Gárriz, A., Marina, M., Romero, F. M., Gonzalez, M. E., Collado, I. G., & Pieckenstain, F. L. (2011). The sesquiterpene botrydial produced by Botrytis cinerea induces the hypersensitive response on plant tissues and its action is modulated by salicylic acid and jasmonic acid signaling. Molecular Plant-Microbe Interactions, 24(8), 888–896. https://doi.org/https://doi.org/10.1094/mpmi-10-10-0248
- Rousseaux, S., Diguta, C. F., Radoï-Matei, F., Alexandre, H., & Guilloux-Bénatier, M. (2014). Non-Botrytis grape-rotting fungi responsible for earthy and moldy off-flavors and mycotoxins. Food Microbiology, 38, 104–121. https://doi.org/https://doi.org/10.1016/j.fm.2013.08.013
- Sage, L., Krivobok, S., Delbos, E., Seigle-Murandi, F., & Creppy, E. E. (2002). Fungal flora and ochratoxin A production in grapes and musts from France. Journal of Agricultural and Food Chemistry, 50(5), 1306–1311. https://doi.org/https://doi.org/10.1021/jf011015z
- Schouten, A., Wagemakers, L., Stefanato, F. L., Kaaij, R. M. V. D., & Kan, J. A. L. V. (2010). Resveratrol acts as a natural profungicide and induces self-intoxication by a specific laccase. Molecular Microbiology, 43(4), 883–894. https://doi.org/https://doi.org/10.1046/j.1365-2958.2002.02801.x
- Schumacher, J. (2016). DHN melanin biosynthesis in the plant pathogenic fungus Botrytis cinerea is based on two developmentally regulated key enzyme (PKS)-encoding genes. Molecular Microbiology, 99(4), 729–748. https://doi.org/https://doi.org/10.1111/mmi.13262
- Schumacher, J. (2017). How light affects the life of Botrytis. Fungal Genetics and Biology, 106(26–41), 26–41. https://doi.org/https://doi.org/10.1016/j.fgb.2017.06.002
- Schumacher, J., Simon, A., Cohrs, K. C., Viaud, M., & Tudzynski, P. (2014). The transcription factor BcLTF1 regulates virulence and light responses in the necrotrophic plant pathogen Botrytis cinerea. PLoS Genetics, 10(1), e1004040. https://doi.org/https://doi.org/10.1371/journal.pgen.1004040
- Segmueller, N., Kokkelink, L. G., Sabine, O., Van Kan, D. J., & Tudzynski, P. (2008). NADPH Oxidases are involved in differentiation and pathogenicityin Botrytis cinerea. Molecular Plant-Microbe Interactions, 21(6), 808–819. https://doi.org/https://doi.org/10.1094/MPMI-21-6-0808
- Serra, R., Braga, A., & Venancio, A. (2005). Mycotoxin-producing and other fungi isolated from grapes for wine production, with particular emphasis on ochratoxin A. Research in Microbiology, 156(4), 515–521. https://doi.org/https://doi.org/10.1016/j.resmic.2004.12.005
- Serra, R., Lourenco, A., Alipio, P., & Venancio, A. (2006). Influence of the region of origin on the mycobiota of grapes with emphasis on Aspergillus and Penicillium species. Mycological Research, 110(8), 971–978. https://doi.org/https://doi.org/10.1016/j.mycres.2006.05.010
- Shang, L., Bai, X., Chen, C., Liu, L., Li, M., Xia, X., & Wang, Y. (2019). Isolation and identification of a Bacillus megaterium strain with ochratoxin A removal ability and antifungal activity. Food Control, 106, 106743. https://doi.org/https://doi.org/10.1016/j.foodcont.2019.106743
- Shukla, S., Park, J. H., & Kim, M. (2020). Efficient, safe, renewable, and industrially feasible strategy employing Bacillus subtilis with alginate bead composite for the reduction of ochratoxin A from wine. Journal of Cleaner Production, 242, 118344. https://doi.org/https://doi.org/10.1016/j.jclepro.2019.118344
- Siegmund, B., & PoLlinger-Zierler, B. (2006). Odor thresholds of microbially induced off-flavor compounds in apple juice. Journal of Agricultural and Food Chemistry, 54(16), 5984–5989. https://doi.org/https://doi.org/10.1021/jf060602n
- Sipiczki, M. (2019). Yeasts in Botrytized wine making. Springer.
- Smith, K. M., Sancar, G., Dekhang, R., Sullivan, C. M., Li, S., Tag, A. G., Sancar, C., Bredeweg, E. L., Priest, H. D., McCormick, R. F., Thomas, T. L., Carrington, J. C., Stajich, J. E., Bell-Pedersen, D., Brunner, M., & Freitag, M. (2010). Transcription factors in light and circadian clock signaling networks revealed by genomewide mapping of direct targets for Neurospora white collar complex. Eukaryotic Cell, 9(10), 1549–1556. https://doi.org/https://doi.org/10.1128/ec.00154-10
- Somma, Stefania, Perrone, & Giancarlo (2012). Diversity of black Aspergilli and mycotoxin risks in grape, wine and dried vine fruits. Phytopathologia Mediterranea, 51, 131–147. https://doi.org/https://doi.org/10.14601/Phytopathol_Mediterr-9888
- Tang, Q., Zhu, F., Cao, X., Zheng, X., Yu, T., & Lu, L. (2019). Cryptococcus laurentii controls gray mold of cherry tomato fruit via modulation of ethylene-associated immune responses. Food Chemistry, 278, 240–247. https://doi.org/https://doi.org/10.1016/j.foodchem.2018.11.051
- Thakur, N. S. (2018). Botrytized Wines: A Review. International Journal of Food and Fermentation Technology, 8(1), 1–13. https://doi.org/https://doi.org/10.30954/2277-9396.01.2018.1
- Tofalo, R., Chaves-Lopez, C., Di Fabio, F., Schirone, M., Felis, G. E., Torriani, S., Paparella, A., & Suzzi, G. (2009). Molecular identification and osmotolerant profile of wine yeasts that ferment a high sugar grape must. International Journal of Food Microbiology, 130(3), 179–187. https://doi.org/https://doi.org/10.1016/j.ijfoodmicro.2009.01.024
- Tronchoni, J., Gamero, A., Arroyo-López, F., Barrio, E., & Querol, A. (2009). Differences in the glucose and fructose consumption profiles in diverse Saccharomyces wine species and their hybrids during grape juice fermentation. International Journal of Food Microbiology, 134(3), 237–243. https://doi.org/https://doi.org/10.1016/j.ijfoodmicro.2009.07.004
- Trotel-Aziz, P., Couderchet, M., Biagianti, S., & Aziz, A. (2008). Characterization of new bacterial biocontrol agents Acinetobacter, Bacillus, Pantoea and Pseudomonas spp. mediating grapevine resistance against Botrytis cinerea. Environmental and Experimental Botany, 64(1), 21–32. https://doi.org/https://doi.org/10.1016/j.envexpbot.2007.12.009
- Valero, A., Sanchis, V., Ramos, A. J., & Marin, S. (2008). Brief in vitro study on Botrytis cinerea and Aspergillus carbonarius regarding growth and ochratoxin A. Letters in Applied Microbiology, 47(4), 327–332. https://doi.org/https://doi.org/10.1111/j.1472-765X.2008.02434.x
- Verhagen, B., Trotel-Aziz, P., Jeandet, P., Baillieul, F., & Aziz, A. (2011). Improved resistance against Botrytis cinerea by grapevine-associated bacteria that induce a prime oxidative burst and phytoalexin production. Phytopathology, 101(7), 768–777. https://doi.org/https://doi.org/10.1094/phyto-09-10-0242
- Verhagen, B. W. M., Trotel-Aziz, P., Couderchet, M., Hoefte, M., & Aziz, A. (2010). Pseudomonas spp.-induced systemic resistance to Botrytis cinerea is associated with induction and priming of defence responses in grapevine. Journal of Experimental Botany, 61(1), 249–260. https://doi.org/https://doi.org/10.1093/jxb/erp295
- Wei, C., Yu, L., Qiao, N., Wang, S., Tian, F., Zhao, J., Zhang, H., Zhai, Q., & Chen, W. (2020). The characteristics of patulin detoxification by Lactobacillus plantarum 13M5. Food and Chemical Toxicology, 146, 111787. https://doi.org/https://doi.org/10.1016/j.fct.2020.111787
- Weiberg, A., Wang, M., Lin, F. M., Zhao, H., Zhang, Z., Kaloshian, I., Huang, H. D., & Jin, H. L. (2013). Fungal small RNAs suppress plant immunity by hijacking host RNA interference pathways. Science, 342(6154), 118–123. https://doi.org/https://doi.org/10.1126/science.1239705
- Wen, S., Zhang, J., Yang, B., Elias, P. M., & Man, M.-Q. (2020). Role of resveratrol in regulating cutaneous functions. Evidence-Based Complementary and Alternative Medicine, 2020(2), 2416837. https://doi.org/https://doi.org/10.1155/2020/2416837
- Williams, K. M., Liu, P., & Fay, J. C. (2015). Evolution of ecological dominance of yeast species in high‐sugar environments. Evolution, 69(8), 2079–2093. https://doi.org/https://doi.org/10.1111/evo.12707
- Williamson, B., Tudzynski, B., Tudzynski, P., & Van Kan, J. A. (2007). Botrytis cinerea: The cause of grey mould disease. Molecular Plant Pathology, 8(5), 561–580. https://doi.org/https://doi.org/10.1111/j.1364-3703.2007.00417.x
- Wong, R. H. X., Coates, A. M., Buckley, J. D., & Howe, P. R. C. (2013). Evidence for circulatory benefits of resveratrol in humans. In K. Brown & O. Vang (Eds.), Resveratrol and Health (Vol. 1290, pp. 52–58). Wiley-Blackwell.
- Xu, L., Sun, X., Wan, X., Li, H., Yan, F., Han, R., Li, H., Li, Z., Tian, Y., Liu, X., Kang, X., & Wang, Y. (2020). Identification of a Bacillus amyloliquefaciens H6 thioesterase involved in zearalenone detoxification by transcriptomic analysis. Journal of Agricultural and Food Chemistry, 68(37), 10071–10080. https://doi.org/https://doi.org/10.1021/acs.jafc.0c03954
- Yang, H., Cai, G., Lu, J., & Plaza, E. G. (2020). The production and application of enzymes related to the quality of fruit wine. Critical Reviews in Food Science and Nutrition, (2020(3), 1–11. https://doi.org/https://doi.org/10.1080/10408398.2020.1763251
- Yiwen, S., Lina, Z., Xiangfeng, Z., Zhen, L., Qiaofei, L., Yangyang, C., & Hongyin, Z. (2020). The possible mechanisms involved in degradation of patulin by Sporidiobolus pararoseus. Journal of Food Science and Biotechnology, 39(2), 16–23. https://doi.org/https://doi.org/10.3969/j.issn.1673-1689.2020.02.003
- Zhang, W., Zhuo, X., Hu, L., & Zhang, X. (2020). Effects of crude β-Glucosidases from Issatchenkia terricola, Pichia kudriavzevii, Metschnikowia pulcherrima on the flavor complexity and characteristics of wines. Microorganisms, 8(6), 953. https://doi.org/https://doi.org/10.3390/microorganisms8060953
- Zhang, Z., Li, S., Sun, D., Yang, Y., Wei, Z., Wang, C., & Lu, L. (2021). Cultivation of Rhodosporidium paludigenum in gluconic acid enhances effectiveness against Penicillium digitatum in citrus fruit. Postharvest Biology and Technology, 172, 111374. https://doi.org/https://doi.org/10.1016/j.postharvbio.2020.111374
- Zhao, X. H., Kim, Y., Park, G., & Xu, J. R. (2005). A mitogen-activated protein Kinase Cascade regulating infection-related morphogenesis in Magnaporthe grisea. The Plant Cell, 17(4), 1317–1329. https://doi.org/https://doi.org/10.1105/tpc.104.029116
- Zheng, X., Wei, W., Rao, S., Gao, L., Li, H., & Yang, Z. (2020). Degradation of patulin in fruit juice by a lactic acid bacteria strain Lactobacillus casei YZU01. Food Control, 112, 107147. https://doi.org/https://doi.org/10.1016/j.foodcont.2020.107147
- Zhu, P., Xu, L., Zhang, C., Toyoda, H., & Gan, -S.-S. (2012). Ethylene produced by Botrytis cinerea can affect early fungal development and can be used as a marker for infection during storage of grapes. Postharvest Biology and Technology, 66, 23–29. https://doi.org/https://doi.org/10.1016/j.postharvbio.2011.11.007