
Abstract
Amaranth sprouts are an edible food with good nutritional qualities and potential biological activities of their proteins. The chemical composition, angiotensin converting enzyme inhibitory activity and antioxidant activity of the sprouts were determined. Sprouts showed a protein content similar to the seeds’ on a dry basis (16%) and a high fiber content (17%). Amaranth sprout proteins presented a capacity to inhibit angiotensin converting enzyme activity similar to other plant proteins (IC50 = 0.9 ± 0.6 mg/mL). This capacity increased after in vitro gastrointestinal digestion (IC50 = 0.26 ± 0.07 mg/mL). Besides other non protein molecules, the amaranth sprout proteins also presented ABTS+. scavenging activity (TEAC = 0.32 ± 0.05 μmol/mg) that increased after in vitro gastrointestinal digestion (TEAC = 0.72 ± 0.08 μmol/mg) and oxygen radical antioxidant capacity. According to these results amaranth sprouts are a nutritive food with potential health promoting properties.
INTRODUCTION
Amaranth (Amaranthus hypochondriacus) is a pseudocereal with great nutritional potential due to its high content of good quality protein and it is an alternative to gluten-containing grains in diets of people with celiac disease.[Citation1] In addition to its nutritive value, amaranth grain contains bioactive compounds with health promoting effects[Citation2] that make amaranth a promising plant as a high-quality food source.
Germination can improve the nutritional quality of a grain by eliminating or inactivating some antinutritional factors, and increasing the digestibility of proteins and starches.[Citation3] Germination may also improve the physiological benefits of the grain by producing peptides from protein mobilization and by causing changes in secondary metabolite distribution.[Citation4] The practice of sprouting has become very popular and sprouts of legumes and cereal grains are offered as new vegetable foods. Sprouts present the advantage of being consumed fresh; they can also be used in many different food preparations including salads, soups, breakfast items, pasta, and baked products.
The production, chemical composition, and nutritive value of amaranth grain sprouts have been studied. Colmenares de Ruiz and Bressani[Citation5] have reported a decrease of lipids and phytic acid and an increase of digestibility and vitamins in sprouts as compared with the seeds. Several studies have shown that phenolic compounds, anthocyanins, and flavonoids increased upon germination of amaranth seeds under certain conditions. The solvent extracts of the germinated seeds showed an increased antioxidant activity.[Citation6,Citation7] Some authors have found that the antioxidant activity correlated with the presence of the polyphenolic compounds,[Citation2] whereas others proposed that this activity may also be ascribed to the presence of other compounds such as peptides.[Citation7] Other physiological activities, besides the antioxidants which are present in amaranth seeds, have not been investigated in germinated amaranth seeds. Many of those activities are developed by peptide free or released after hydrolysis, as is the case of antihypertensive activity. Amaranth protein hydrolysates and peptides develop antihypertensive properties, as shown by in vivo and in vitro tests.[Citation8–Citation10] The mobilization of proteins during germination generates peptides that might present physiological activity. Aphalo et al.[Citation11] have shown that amaranth protein-encrypted peptides are released mainly from albumin and globulin fractions, after 48 h of seed imbibition.
The advantages of seed germination from the point of view of nutritional quality, and the fact that allows obtaining peptides at lower cost than conventional hydrolysis processes, makes germination be an interesting process to study its potential as a supplier of bioactive peptides. In the present work we aimed to investigate the potential of amaranth sprouts as an ingredient with biological activity in particular as a source of antioxidant and angiotensin converting enzyme (ACE) inhibitor peptides, two activities of utmost importance for consumer health.
MATERIALS AND METHODS
Chemicals and Reagents
Porcine pepsin and pancreatin (4x-USP) were purchased from MP Biomedicals Inc., Solon Ohio, USA), polypeptide SDS-PAGE (denaturant polyacrylamide gel electrophoresis) molecular marker standards (MW 1.42–26.6 kDa) BIO-RAD Hercules California USA, and gel filtration calibration kit (GE-Healthcare, Buckinghamshire, UK). For antioxidant determinations: 2,2´azinobis-(3-ethylbenzotiazoline-6-sulfonic acid (ABTS) and 6-hydroxyl-2,5,7,8-tetramethylchroman-2–carboxylic acid (TROLOX) from Fluka-Sigma-Aldrich Chemie Steinheim, Germany), fluorescein sodium from Fluka (Steinheim, Germany) and 2,2´-azo-bis(2-methylpropionamidine (AAPH) dihydrochloride from Aldrich (Wisconsin, USA).
Angiotensin converting enzyme (ACE; from rabbit lungs), hippuryl-histidyl-leucine (HHL), cyanuric chloride and N-[¨(S)-3-mercapto-2-methylpropionil]-L-proline (captopril) were provided by Fluka-Sigma-Aldrich Chemie Steinheim, Germany) for ACE inhibitory capacity assay. All chemicals used (analytical grade) were provided by Sigma-Aldrich (Steinheim, Germany).
Seeds and Flour
Seeds from Amaranthus hypochondriacus, Antorcha variety (harvest 2011) were grown in Facultad de Ciencias Agrarias—Universidad Nacional de Río Cuarto, Córdoba, Argentina. Seeds were ground and screened by 0.092 mm mesh, in Facultad de Ciencias Agrarias y Forestales, Universidad Nacional de La Plata. The flour was defatted with hexane at 4ºC for 24 h (100 g/L suspension), dried at room temperature and stored at 4ºC until used.
Sprouting
Seeds were soaked in 0.02% (w/v) NaClO for 20 min, washed several times with distilled water, and then spread on absorbent paper wetted with distilled water in covered plastic boxes. After standing at 37°C for 48 h, the seeds germinated into sprouts of approximately 50 mm. Finally, sprouts were dried at 42°C for 1 h and stored at –80ºC until use. The germination experiments were carried out after harvest in three different batches.
Protein Extraction
Seed isolate (I) was prepared from defatted flour according to Martínez and Añón.[Citation12] Briefly, flour was suspended in water (10% w/v) and pH adjusted to 9.0 with NaOH 2 mol/L. Suspensions were stirred for 30 min at room temperature and centrifuged at 9000 × g for 20 min at 15ºC. Supernatants were adjusted to pH 5.0 with HCl 2mol/L and then were centrifuged at 9000 × g for 20 min at 4ºC. The precipitate (seed isolate) was suspended in water, neutralized with NaOH 0.1 mol/L, and freeze-dried.
The sprout isolate (SI) was prepared from a sprout homogenate obtained by soaking the sprouts in water in a ratio of 1/10 (w/v) with Ultraturrax homogeneizer (Ultraturrax T-25. Janke & Kunkel IKA-Labortechnik-Germany). SI was prepared using the same procedure described previously for seed isolate. After precipitation at pH 5.0 with HCl 2N, the supernatant and precipitate were neutralized and freeze-dried. The precipitate and supernatant were labelled as SI and as sprout supernatant (SS), respectively. Protein extraction was done by triplicate.
Simulated Gastrointestinal Protein Digestion
I, SI, and SS were subjected to simulated gastrointestinal digestion according to Roesler and Gururaj[Citation13] with minor modifications. Sample dispersions (20 mg/mL) were adjusted to pH 2 with 0.1 mol/L HCl and incubated at 37°C with porcine pepsin in 0.03 mmol/L NaCl at pH 2 at a 1:10 enzyme:protein ratio for 60 min. Afterwards, the mixture was adjusted to pH 7.0 10 with 0.1 M NaOH and pancreatin in 0.1 mol/L NaHCO3 was added at a 1:300 enzyme:protein ratio. After 60 min incubation at 37°C the reaction was stopped by heating at 90°C for 10 min. The mixtures were centrifuged at 10,000 × g for 20 min at 20ºC and supernatants were analyzed. Each sample was digested at least twice.
Chemical Composition
The chemical composition of the sprouts was carried out according to the following determinations: protein content using Kjeldahl method (f = 5.85) and insoluble dietary fiber following the AOAC methods,[Citation14] moisture and ashes were done using AOCS methods.[Citation15] All the determinations were done by duplicate.
Tricine-SDS polyacrylamide gel electrophoresis
Runs were performed in minislabs (Bio Rad Protean II Model) according to Schägger.[Citation16] The stacking, spacer, and separating gels of 4, 10, and 16% w/v acrylamide/6 M urea, respectively, were used. Freeze-dried protein samples (10 mg/mL) were dissolved in a reducing sample buffer (12% w/v SDS, 6% v/v mercaptoethanol, 30% w/v glycerol, 0.05% w/v Coomasie G 250 and 150 mM Tris-HCl pH 7.0) and centrifuged at 10,000 × g for 15 min at 20ºC; the supernatants used to load the gel (10 µL/lane). Polypeptide SDS-PAGE molecular weight standards were used as molecular markers. Gels were fixed in 50% v/v methanol, 10% v/v acetic acid, and water for 30 min and stained with Coomasie Brillant Blue. Duplicate runs of each sample from five experiments were performed.
Activity Antioxidant Determinations
Scavenging of ABTS+ radical
ABTS radical cation (ABTS+.) decoloration assay was performed according to Siddhuraju et al.[Citation17] with some modifications. ABTS+. was produced by reaction of 7 mmol/L ABTS∙+ (2,2´-azinobis-(3-ethylbenzothiazoline-6- sulfonic acid) in 50 mmol/L Tris pH 8.0 with 2.45 mmol/L potassium persulfate (final concentration) in the dark at room temperature for more than 16 h before use. Prior to the assay, the solution was diluted in 50 mmol/L Tris pH 8.0 and equilibrated at room temperature to give an absorbance of 0.70 ± 0.02 at 734 nm in a 1 cm cuvette (Beckman DU 650 spectrophotometer). The samples (10 µL) of different protein concentration (1–5 mg/mL) were mixed with 990 µL of ABTS∙+ solution, and absorbance at 734 nm was measured at different times (0, 1, 2, 5, 10, 15, 20, 30, 45, 60, and 120 minutes). TROLOX (6–hydroxyl—2, 5, 7, 8—tetramethylchroman—2-carboxylic acid was used as a positive control of the scavenging activity. Scavenging activity sample and TROLOX (%S) were calculated as follows:
where, AbCo: absorbance at 734 nm of the control without sample at t = 0 min and Abct: at t = 30 min. AbSt: absorbance of sample at t = 30 min. The protein content of the samples was determined by Lowry method.[Citation18] The %S was plotted as a function of the protein content of the sample or mM concentration of TROLOX to obtain the concentration (as mg/mL of protein or mM TROLOX) necessary to reduce absorbance by 50% (IC50). Results were expressed as TROLOX equivalent antioxidant capacity (TEAC) and were estimated as:
The scavenging capacity of each sample was measured at least in triplicate.
Oxygen radical absorbance capacity assay (ORAC)
This method was performed according to Orsini et al.[Citation19] Each protein sample was evaluated on triplicate and diluted as follows: 1:5; 1:10; 1:20; 1:40; 1:100; and 1:200 in buffer phosphate pH 8. Briefly, differential net areas under fluorescein decay curves were evaluated between the background sample (without AAPH) and the assayed samples. Additionally, a standard TROLOX curve (6–100 µmol/L) was done for each dilution.
ORAC value was expressed as: mg TROLOX /mg protein
Extraction of Hydrophobic Molecules
To extract molecules with potential antioxidant activity, amaranth isolate (I), SI, and SS were sequentially extracted with methanol-0.16 mol/L HCl for 1 h and 70% acetone for 2 h according to the method of Pasko et al.[Citation6] for extracts preparation. Supernatants were stored at 4ºC until used. The volume of each extract was referred to the amount of protein of the corresponding sample. The extraction of hydrophobic molecules was carried out twice.
ACE-Inhibitory Activity Assay
ACE-inhibitory activity was assayed according to Hurst and Lovell-Smith[Citation20] with modifications. The reaction mixture (200 μL) containing 1.25 mU ACE, 10 mmol/L substrate HHL, 0.2 mol/L borate buffer pH 8.3, 2.0 M NaCl and 10 μL sample was incubated at 37ºC for 30 min and stopped by heating at 90ºC for 15 min. Afterwards, 600 μL of 0.2 M potassium dihydrogen phosphate buffer, pH 8.3 was added followed by 515 μL of 3% w/v cyanuric chloride in dioxane. The hippurate released, which reacted with the cyanuric chloride, was quantified at 382 nm. Captopril N-[(S)-3-mercapto-2-methylpropionil]-L-proline was used as positive control of ACE inhibition. The IC50 sample value was defined as the protein concentration (mg/mL) in the reaction mixture necessary to inhibit ACE activity by 50%. It was calculated by linear regression of the ACE inhibition (%) versus log peptide concentration (mg/mL) plot. Assayed samples of different protein concentration (0.1–1 mg/mL) were determined by Lowry method.[Citation18] Each sample activity was assayed at least in triplicate according to the following equation:
where, Abmáx: absorbance at 382 nm of the control without sample and Abs: absorbance of sample at 382 nm. BAbs and BAbmáx: background absorbance at 382 nm without enzyme (ACE).
Fast Protein Liquid Chromatography (FPLC) Gel Filtration
Samples were analyzed at room temperature in a Superdex 75 column using a FPLC System (Pharmacia LKB, Uppsala, Sweden). Samples, 0.2 mL containing 4 mg of protein in 0.2 mol/L borate buffer pH 8.3–0.4 mol/L NaCl, were eluted with the same buffer at a flow rate of 0.2 mL/min. Elution profiles at 280 nm were obtained. The column was calibrated with gel filtration calibration kit.
Statistical Analysis
Multifactor analysis of variance (ANOVA) of variables was performed by means of Statgraphics Plus, a software package from Statgraphics Corp. (Rockville, MD, USA). Fisher-LSD test was used to compare the mean of different variables (p < 0.05), results are expressed as mean ± SD. Different letters were used to indicate significant differences.
RESULTS AND DISCUSSION
The chemical composition of the amaranth sprouts was compared with that of the seeds (); the sprouts presented a content of moisture higher than that of the seeds but the amount of protein on a dry basis was similar in both samples (values in brackets). The sprouts showed a high content of fiber, higher than that reported by[Citation5] in 48 h staged sprouts.
TABLE 1 Amaranth sprouts and seeds composition
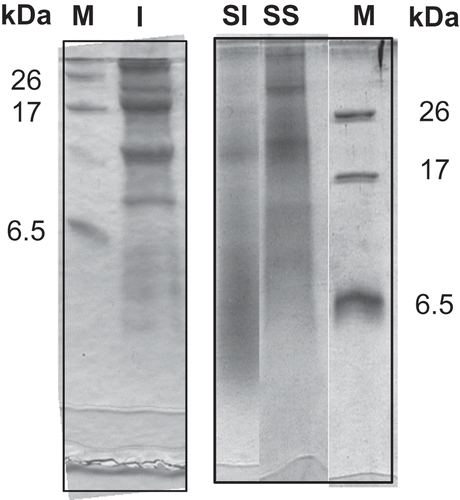
The protein content of amaranth seeds remained constant after 48 h germination which was in accordance with Colmenares de Ruiz and Bressani[Citation5] information. Nevertheless, the protein composition would probably have changed and new housekeeping proteins might have been synthesized from the storage proteins degradation products. This hypothesis is supported by results that showed the storage globulins hydrolysis and an increase of albumins after 48 h of amaranth seed germination.[Citation5,Citation11] According to the values shown in , the sum of the non-fiber carbohydrates and the lipids of the sprouts calculated by difference had decreased. They are used by the seed as a fuel and to produce new molecules. The protein extracted at pH 9 from the sprouts was divided in two fractions, the protein isolate, precipitated at pH 5 (55% protein), and the supernatant, containing the proteins which are soluble at pH 5 (32% protein; materials and methods section).
The proteins of these two fractions were analyzed by electrophoresis and compared with a seed protein isolate (). The SI pattern (, lane SI) showed a higher proportion of polypeptides lower than 6.5 kDa than the seed isolate (, lane I). These results are an indication of the proteolysis that took place during germination. The supernatant fraction pattern (, lane SS) presented a proportion of polypeptides larger than 17 kDa higher than those shown in the SI pattern. The proteins of the supernatant fraction comprise albumins and housekeeping proteins which have been probably less hydrolyzed than the storage globulins.
FIGURE 1 Tricine-SDS-PAGE. I: Seed isolate; SI: sprout isolate; SS: sprout supernatant; M: standard proteins; MW: standard molecular masses.
The chromatographic results of the proteins soluble at pH 8 are shown in . According to the seed and SI chromatograms, the proteins with a molecular weight lower than 6.5 kDa (area between lines A and B) are in a similar proportion in the SI (55%) than in the seed isolate (50%).These results, that are rather different from the electrophoretic results, may be explained considering that most of the new small peptides of the SI shown in the electrophoretic pattern, were aggregated in the chromatographic sample at pH 8 without the disrupting agent SDS.
FIGURE 2 Gel filtration chromatography. I: Seed isolate; SI: sprout isolate; SS: sprout supernatant. Molecular markers elution volumes are indicated with arrows (29 kDa and 6.5 kDa). Percentages under the chromatograms correspond to each profile peaks area between the A and B lines.
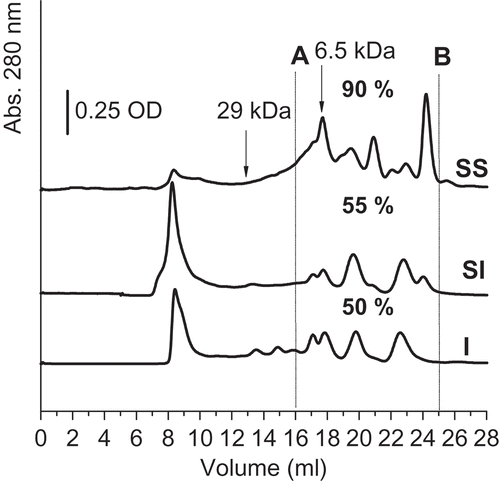
On the other hand, the supernatant profile showed species smaller and different from those of the other two fractions (area of SS between A and B: 90%). This result is also different from that of the electrophoresis, which showed the supernatant peptides of higher size than those of the isolate. The reason for these differences could be the fact that the isolates contain globulins, which have disulfide bridges, whereas the supernatant proteins present few or no disulfide bridges.[Citation11,Citation12] The isolate globulins were dissociated in the electrophoresis and aggregated in the chromatography.
ACE Inhibitory Capacity
As a first attempt to explore the possible antihypertensive activity of the sprout proteins, the samples and the products of their in vitro gastrointestinal digestion were tested as possible inhibitors of the ACE activity. As shown in the activity of the sprout proteins (isolate and supernatant) were similar to that of the seed isolate (sample I). They showed a low activity but higher than an A. cruentus protein isolate (IC50 = 12 mg/mL).[Citation21] After the in vitro gastrointestinal digestion the three samples improved their activity showing a lower IC50, which suggests that the hydrolytic action of the gastrointestinal enzymes released peptides with ACE inhibitor activity.
The presence of ACE inhibitory compounds in the amaranth sprouts allows us to consider them as a promising health promoting food. The ACE inhibitory activity shown by the SI and supernatant suggested the presence of bioactive compounds, probably peptides. ACE inhibitory compounds were not described in amaranth and quinoa sprouts until now. On the other hand, peptides with ACE inhibitory activity were described in a fermented product coming from buckwheat sprouts,[Citation22] they showed an IC50 of 0.69–56 mmol/L. Peptides with ACE inhibitory activity were also isolated from hydrolysates of other plant sprouts as pea, walnut, and lentil.[Citation23–Citation25] In this work we demonstrated that the ACE inhibitory activity increased after the hydrolysis of the amaranth SI and supernatant showing IC50 similar to those informed for other plants SI hydrolysates (pea sprout trypsin hydrolysate IC50 0.24 mg/mL and pea sprout pepsin hydrolysate IC50 1.59 mg/mL).[Citation24] Our results suggested that ACE inhibitory peptides were released from the amaranth sprout proteins after the digestion.
The release of ACE inhibitory peptides by the in vitro gastrointestinal digestion was also shown in an amaranth seed isolate, some amaranth protein fractions and in other species proteins.[Citation8,Citation26,Citation27] The digested proteins of this work presented IC50 values similar to those reported for other digested seed isolates (amaranth IC50 = 0.439 ± 0.018 mg/mL,[Citation21] soybean IC50 = 0.37 ± 0.02 mg/mL[Citation28]) although lower than that for synthetic drugs such as captopril (IC50 2.3 × 10–Citation3 mg/mL) used as positive control of our trials. This behavior, so far, is characteristic of natural inhibitors of this enzyme reported in different studies.
Antioxidant Capacity
The neutralization of free radicals is one of the most important mechanisms by which peptides control oxidative reactions in foods.[Citation29] In this work the potential antioxidant capacity of the samples was analyzed by means of their ABTS+ radical scavenging activities Results () indicate that both, seed and SIs, presented similar antioxidant activity. These activities were higher than that of an A. mantegazzianus isolate[Citation19] and similar to those of other plants proteins.[Citation30] On the other hand, the SS showed a higher activity than the isolates. The antioxidant activity of the three samples after in vitro gastrointestinal digestion increased, the digested supernatant activity was higher than those of the other samples. These results suggested that the in vitro digestion had released peptides with ABTS radical scavenging capacity. These activities were similar to those of digested A. mantegazzianus proteins and to the activities of other plant proteins subjected to hydrolytic treatments.[Citation19,Citation30,Citation31]
TABLE 2 Amaranth sprouts in vitro biological activity
The antioxidant capacity of the samples was also analyzed using the ORAC assay. Results obtained () indicate that seeds and sprouts isolates presented a similar ability to absorb oxygen radicals while the SS showed greater activity. This behavior is similar to that obtained for the scavenging of ABTS+ radical. After the simulated gastrointestinal digestion, seeds, and sprouts isolates showed no variation in their oxygen radical antioxidant capacity. While the activity corresponding to the SS decreased after digestion although remains higher than that of seeds and sprouts digested and undigested.
To search for the presence of non-protein molecules with antioxidant capacity in the samples, the isolates and supernatant were treated with solvents and the ABTS+ radical scavenging activity of the extracts were tested. According to the results (), all the samples contained some hydrophobic molecules, which may include some hydrophobic peptides, with antioxidant activity. The isolate extracts presented about 20% of the sample activity, whereas the supernatant extract showed 45% of that sample activity (). This work results suggested that those compounds would be present in the protein samples and that both, the hydrophobic and protein molecules, would be responsible for the antioxidant activity.
TABLE 3 ABTS+ protein and extract scavenging capacity*
Concerning the antioxidant activity, this work results demonstrated that the two sprout protein fractions presented ABTS radical scavenging activity and oxygen radical antioxidant capacity and that the supernatant fraction presented the highest activities. Nevertheless, these fraction activities cannot be assigned only to proteins because, according to the fraction protein content, they may contain other molecules, like polyphenols, with ABTS radical scavenging activity. Different amount of phenolic compounds in amaranth seeds have been described,[Citation32,Citation33] and other authors[Citation6] have reported the increase of phenolic compounds with antioxidant activity in germinated amaranth seeds.
The increase of the ABTS radical scavenging activity of the samples after the in vitro digestion may be assigned to the release of peptides with antioxidant activity and/or the increase of the antioxidant activity of compounds (polyphenols) that were attached to the proteins,[Citation31–Citation33] while we cannot associate the absorption of oxygen radicals to polyphenols or proteins. Further studies are needed to identify the compound or compounds responsible for both activities. The reported antioxidant activity of amaranth, quinoa, and buckwheat sprouts was until now assigned only to non-protein molecules.[Citation6,Citation7] Nevertheless, peptides coming from other plant sprout hydrolysates were found to present antioxidant activity.[Citation24] Our results indicated that non-protein molecules in the sprout fractions showed part of the total antioxidant activity and, therefore, antioxidant peptides would also be present in the samples.
CONCLUSIONS
According to this work results we may consider A. hypochondriacus sprouts as a health promoting food; they presented a high content of fiber and are a source of proteins with potential antihypertensive and antioxidant activities. It was demonstrated that the sprout proteins presented a capacity to inhibit ACE activity similar to other plant proteins and lower to the synthetic drugs such as captopril. This capacity increased after in vitro gastrointestinal digestion. Results also showed that, besides other non-protein molecules, the amaranth sprout proteins presented ABTS+ radical scavenging activity that increased after in vitro gastrointestinal digestion and ability to absorb oxygen radicals decreases or remains unchanged after in vitro gastrointestinal digestion. Though in vivo analyses are needed to confirm the mentioned activities and also is needed broaden and deepen the study of these and other activities, this work results allowed us to present the amaranth sprouts as a potential functional food. It should also be noted that the germination process is less expensive than others used in the production of functional peptides and which is also a simple process from the point of view of process technical specialization required.
ACKNOWLEDGMENTS
The authors would like to thank Eng. Guillermo Peiretti from Facultad de Ciencias Agrarias, Universidad Nacional de Río Cuarto, Córdoba, Argentina for providing the amaranth seeds (Antorcha variety).
Related Research Data
REFERENCES
- Ballabio, C.; Uberti, F.; Di Lorenzo, C.; Brandolini, A.; Penas, E.; Restani, P. Biochemical and Immunological Characterization of Different Varieties of Amaranth (Amaranthus L. Ssp) as a Safe Ingredient for Gluten Free Products. Journal of Agricultural and Food Chemistry 2011, 50(24), 12969–12974.
- Huerta-Ocampo, J.A.; Barba de la Rosa, A.P.; Amaranth: A Pseudocereal with Nutraceutical Properties. Current Nutrition and Food Science 2011, 7(1), 1–9.
- Urbano, G.; Aranda, P.; Vílchez, A.; Aranda, C.; Cabrera, L.; Porres, J.M.; López-Jurado, M. Effects of Germination on the Composition and Nutritive Value of Proteins in Pisum Sativum L. Food Chemistry 2005, 93, 671–679.
- Mora-Escobedo, R.; Robles-Ramírez, M.C.; Ramón-Gallegos, E.; Reza-Alemán, R. Effect of Protein Hydrolysates from Germinated Soybean on Cancerous Cells of Human Cervix: An In Vitro Study. Plant Foods for Human Nutrition 2009, 64, 271–278.
- Colmenares de Ruiz, A.S.; Bressani, R. Effect of Germination on the Chemical Composition and Nutritive Value of Amaranth Grain. Cereal Chemistry 1990, 67(2), 519–522.
- Paśko, P.; Bartoń, H.; Zagrodzki, P.; Gorinstein, S.; Folta, M.; Zachwieja, Z. Anthocyanins, Total Polyphenols, and Antioxidant Activity in Amaranth and Quinoa Seeds and Sprouts During Their Growth. Food Chemistry 2009, 115, 994–998.
- Caselato-Sousa, V.; Amaya-Farfán, J. State of Knowledge on Amaranth Grain: A Comprehensive Review. Journal of Food Science 2012, 77, R93–R104.
- Tovar-Pérez, E.G.; Guerrero-Legarreta, I.; Farrés-González, A.; Soriano-Santos, J. Angiotensin I Converting Enzyme-Inhibitory Peptide Fractions from Albumin 1 and Globulin as Obtained of Amaranth Grain. Food Chemistry 2009, 116, 437–444.
- Vecchi, B.; Añón, M.C. ACE-Inhibitory Tetrapeptides from Amaranthus hypochondriacus 11S Globulin. Phytochemistry 2009, 70, 864–870.
- Fritz, M.; Vecchi, B.; Rinaldi, G.; Añón, M.C. Amaranth Seed Protein Hydrolysates Have In Vivo and In Vitro Antihypertensive Activity. Food Chemistry 2011, 126, 878–884.
- Aphalo, P.; Martínez, E.N.; Añón, M.C. Structural Modifications of Amaranth Proteins During Germination. Protein Journal 2009, 28, 131–138.
- Martínez, E.N.; Añón, M.C. Composition and Structural Characterization of Amaranth Protein Isolates: An Electrophoretic and Calorimetric Study. Journal of Agricultural and Food Chemistry 1996, 44, 2523–2530.
- Roesler, K.R.; Gururaj, R.A. Rapid Gastric Fluid Digestion and Biochemical Characterization of Engineered Proteins Enriched in Essential Amino Acids. Journal of Agricultural and Food Chemistry 2001, 49, 3443–3451.
- Association of Official Analytical Chemists. Official Methods of Analysis (methods 954.01 and 991.43), 15th Ed; 3rd Supplement. AOAC International: Arlington, VA, 1992.
- American Oil Chemists’ Society. Official Methods and Recommended Practices of the AOCS (Methods Ac 2-41 and Ba 5a-49), 6th Ed; 3rd printing. AOCS Press: Urbana, IL, 2009.
- Schägger, H. Tricine-SDS-PAGE. Nature Protocols 2006, 1(1), 16–21.
- Siddhuraju, P. The Antioxidant Activity and Free Radical-Scavenging Capacity of Phenolics of Raw and Dry Heated Moth Bean (Vigna aconitifolia; Jacq.) Marechal Seed Extracts. Food Chemistry 2006, 99, 149–157.
- Lowry, O.; Rosebrough, N.; Farr, A.; Randall, R. Protein Measurement with the Folin Phenol Reagent. Journal of Biological Chemistry 1951, 193, 265–275.
- Orsini Delgado, M.C.; Tironi, V.A.; Añón, M.C. Antioxidant Activity of Amaranth Protein or Their Hydrolysates under Simulated Gastrointestinal Digestion. LWT–Food Science and Technology 2011, 44, 1752–1760.
- Hurst, P.L.; Lovell-Smith, C.J. Optimized Assay for Serum Angiotensin-Converting Enzyme Activity. Clinical Chemistry 1981, 27, 2048–2052.
- Tiengo, A.; Faria, M.; Netto, F.M. Characterization and ACE-Inhibitory Activity of Amaranth Proteins. Journal of Food Science 2009, 74, H121–H126.
- Koyama, M.; Hattori, S.; Amano, Y.; Watanabe, M.; Nakamura, K. Blood Pressure—Lowering Peptides from Neo-Fermented Buckwheat Sprouts: A New Approach to Estimating ACE-Inhibitory Activity. PLOS One 2014, 9(9), e105802.
- Bamdad, F.; Dokhani, S.; Keramat, J.; Zaneie, R. The Impact of Germination and In Vitro Digestion on the Formation of Angiotensin Converting Enzyme (ACE) Inhibitory Peptides from Lentil Proteins Compared to Whey Proteins. World Academy Science Engineering and Technology 2009, 37, 36–46.
- Jakubczyc, A.; Karaś, M.; Beraniak, M. Antyhipertensive and Antioxidative Activity of Peptides Derived from Pea Sprouts (Pisum sativum) Protein Hydrolysates. Annales Universitatis Mariae–Curie Skłodowska, Sectio DDD Pharmacia 2011, 24(3), 175–182.
- Gu, X.; Hou, Y.; Li, D.; Wang, J.-Z.; Wang, F. Separation, Purification, and Identification of Angiotensin I-Converting Enzyme Inhibitory Peptides from Walnut (Junglans regia L.) Hydrolysate. International Journal of Food Properties 2015, 18(2), 266–276.
- Torruco-Uco, J.; Chel-Guerrero, L.; Martínez-Ayala, A.; Dávila-Ortíz, G.; Betancur-Ancona, D. Angiotensin-I Converting Enzyme Inhibitory and Antioxidant Activities of Protein Hydrolysates from Phaseolus lunatus and Phaseolus vulgaris Seeds. LWT–Food Science and Technology 2009, 42, 1597–1604.
- Jamdar, S.N.; Rajalakshmi, V.; Pednekar, M.D.; Juan, F.; Yardi, D.; Sharma, A. Influence of Degree of Hydrolysis on Functional Properties, Antioxidant Activity, and ACE-Inhibitory Activity of Peanut Protein Hydrolysate. Food Chemistry 2010, 121, 178–184.
- Lo, W.M.Y.; Farnworth, E.R.; Li-Chan, E.C.Y. Angiotensin I-Converting Enzyme Inhibitory Activity of Soy Protein Digestion: A Dynamic Model System Simulating the Upper Gastrointestinal Tract. Journal of Food Science 2006, 71(3), S231–S237.
- Pinchuk, I.; Shoval, H.; Dotan, Y.; Lichtenberg, D. Evaluation of Antioxidants: Scope, Limitations, and Relevance of Assays. Chemistry and Physics of Lipids 2012, 165, 638–647.
- Samaranayaka, A.G.P.; Li-Chan, E.C.Y. Food-Derived Peptidic Antioxidants a Review of Their Production, Assessment, and Potential Applications. Journal of Functional Foods 2011, 3, 229–254.
- Tounkara, F.; Bashari, M.; Wei Le, G.; Shi, Y.-H. Antioxidant Activities of Roselle (Hibiscus sabdariffa L.) Seed Protein Hydrolysates and Its Derived Peptide Fractions. International Journal of Food Properties 2014, 17(9), 1998–2011.
- Repo-Carrasco-Valencia, R.; Hellstrom, J.K.; Pihlava, J.M.; Mattila, P.H. Flavonoids and Other Phenolic Compounds in Andean Indigenous Grains: Quinoa (Chenopodium quinoa), Kaniwa (Chenopodium pallidicaule), and Kiwicha (Amaranthus caudatus). Food Chemistry 2010, 120, 128–133.
- Tironi, V.A.; Añón, M.C. Amaranth Proteins as a Source of Antioxidant Peptides: Effect of Proteolysis. Food Research International 2010, 43, 315–322.