Abstract
Las altas presiones hidrostáticas (APH) se han propuesto como método cuarentenario alternativo para destruir las larvas de la mosca de la fruta Anastrepha ludens y Anastrepha obliqua que infestan al mango. Sin embargo, existe escasa información científica sobre el efecto de las APH en la fisiología poscosecha de frutos. El objetivo de este trabajo fue evaluar el efecto de la presurización en la fisiología del mango ‘Ataulfo’ durante su maduración a 25°C. Se obtuvieron mangos en madurez fisiológica, se presurizaron a 15, 30 y 60 MPa por 10 o 20 min (se mantuvo un testigo sin tratamiento). Se almacenaron durante 14 días y se evaluaron cambios fisiológicos y fisicoquímicos. Las APH modificaron la fisiología poscosecha de los frutos de forma determinante pero no inhibieron el proceso de maduración. El efecto dependió del nivel de presión, el tiempo de presurización y la interacción de ambos parámetros.
High hydrostatic pressure (HHP) has been proposed as a quarantine alternative method to destroy larvae from fruit flies Anastrepha ludens and Anastrepha obliqua that infest the mango. However, there is scarce scientific information on the effect of HHP on the postharvest physiology of target fruits. The aim of this study was to evaluate the effect of pressurization of low intensity in the physiology of the mango “Ataulfo” during ripening in storage at 25°C. Mangoes were obtained in a state of physiological maturity and pressurized to 15, 30, and 60 MPa for 10 or 20 min (fruits control were maintained without treatment). Mangoes were stored for 14 days and changes in physiological and physicochemical variables were evaluated. HHP changed the postharvest physiology of fruits but did not inhibit the ripening process; the effect was dependent on the level of pressure, pressurization time, and interaction of both parameters.
Palabras clave:
Introducción
En diferentes países se exige a los exportadores de frutas que éstas sean sometidas a tratamientos cuarentenarios de sanitización para prevenir que sus cultivares sean infectados con moscas de las fruta no nativas de su territorio (Gazit, Rossler, Wang, Tang, & Lurie, Citation2004). Los tratamientos aprobados incluyen fumigación con compuestos tóxicos, irradiación y tratamientos físicos que usan calor o frío (Armstrong & Mangan, 2007). Estos tratamientos físicos han sido más ampliamente empleados por haber demostrado que son efectivos para matar los huevos y las larvas desarrolladas dentro de los frutos hospederos, presentando pocos efectos negativos en la calidad organoléptica de la fruta durante sutratamiento o posterior almacenamiento, y porque no incrementan sustancialmente el costo del producto (Mangan & Hallman, 1998). Sin embargo, todos los métodos cuarentenarios inducen cambios en la calidad de las frutas que afectan su vida de anaquel, cambios que dependen del tipo de fruto y la variedad (Hallman, Citation2000). Es por ello que constantemente se buscan métodos cuarentenarios alternativos que permitan minimizar el efecto negativo sobre las frutas, sin afectar su valor o su comercialización. Los efectos negativos que pueden inducir los métodos cuarentenarios incluyen lesiones superficiales, picado, pérdida de aroma, decoloración interna y superficial, ablandamiento, senescencia, variación de peso, maduración no homogénea y predisposición al deterioro microbiano (Armstrong & Mangan, 2007; Mangan & Hallman, 1998; Sosa-Morales et al., Citation2009). Recientemente se ha estudiado la eficiencia como métodos cuarentenarios de algunas tecnologías emergentes que han probado su eficiencia en la destrucción de microorganismos en alimentos. Las principales tecnologías estudiadas incluyen: campos de pulsos eléctricos (Hallman & Zhang, Citation1997), radiofrecuencia (Wang, Monzon, Johnson, Mitcham, & Tang, Citation2007), tratamiento térmico en agua asistido con radio frecuencia (Monzon et al., Citation2007; Sosa-Morales et al., Citation2009, Sosa-Morales, Valerio-Junco, López-Malo, & García, Citation2010; Tiwari, Wang, Birla, & Tang, Citation2008), microondas (Varith, Sirikajornjaru, & Kiatsiriroat, Citation2007), y altas presiones hidrostáticas (APH) (Butz & Tauscher, Citation1995; Candelario-Rodríguez et al., Citation2009; Neven, Follet, & Raghubeer, Citation2007).
Diferentes estudios han demostrado la eficiencia de las APH para destruir insectos que infestan las frutas, incluyendo la mosca de la fruta del Mediterráneo (Ceratitis capitata Wiedemann) (Butz & Tauscher, Citation1995), la mosca de la fruta de la cereza Occidental (Rhagoletis indifferens Curran), polilla de la manzana (Cydia pomonella L.) (Neven et al., Citation2007), y la mosca Mexicana de la fruta (Anastrepha ludens Loew) (Candelario-Rodríguez et al., Citation2009). Sin embargo las condiciones requeridas para destruir la plaga (cercanas a los 150 MPa) afectaron negativamente la calidad de la fruta (Neven et al., Citation2007). Recientemente se ha demostrado que las APH combinadas con calor (50°C) o frío (0°C), pueden destruir los huevos y larvas de mosca de la fruta de Anastrepha ludens (Loew) y A. oblicua (Macquart) usando niveles de presurización por debajo de los 100 MPa (Velázquez et al., Citation2010a,b). Esto es importante porque se sabe que los mangos verdes de las variedades Florida presentan cambios mínimos externos cuando se someten a100 MPa por 5 min (Candelario-Rodríguez et al., Citation2009). Por otra parte, la mosca de la fruta mexicana (A. ludens) y de la India occidental (A. oblicua) son las dos plagas más importantes que infectan frutas en México, incluyendo los mangos destinados a exportación (Camargo, Odell, & Jirón, Citation1996; Hernández, Orozco, Breceda, & Domínguez, Citation2007).
Los mangos son frutas tropicales climatéricas y su producción y comercialización en los mercados internacionales representan una importante actividad comercial (Montalvo,García, Tovar, & Mata, Citation2007; Ragazzo-Sánchez et al., Citation2009). México es el cuarto productor de mango en el mundo y el principal exportador. El cultivo de mango ‘Ataulfo’ es importante en nuestro país debido a que es una de las principales variedades que se exportan (SAGARPA, Citation2007). Actualmente hay muy poca información sobre los cambios en la calidad del mango ‘Ataulfo’ al ser sometido a APH. El objetivo del presente estudio fue determinar el efecto que las APH ejercen sobre la fisiología postcosecha del mango ‘Ataulfo’ cosechado en estado de madurez fisiológica, para establecer la viabilidad de emplear esta tecnología emergente como método cuarentenario para eliminar la mosca de la fruta.
Materiales y métodos
Obtención de los mangos
Se utilizó mango (Mangifera indica L) ‘Ataulfo’ en estado de madurez fisiológica (verde claro, cavidad basal (pedúnculo) hundida y con hombros llenos), cosechado a 1.5 km del poblado 5 de Mayo, Municipio de Tepic Nayarit, de la temporada 2010. Los frutos se trasladaron al laboratorio, endonde se lavaron y se sumergieron por cinco minutos en una solución de 2-(4-tiazolil)-1H-bencimidazol (20 mg/L) para prevenir crecimiento de hongos. Posteriormente se dejaron se secar e inmediatamente se usaron para el presente estudio.
Presurización de los mangos
Los mangos se dividieron en siete grupos para ser sometidos presiones de 15, 30 ó 60 MPa, por 10 ó 20 min. Se mantuvo un grupo testigo sin tratamiento de presurización. Cada grupo estuvo formado por 50 frutos incluyendo el testigo. Los tratamientos con presión hidrostática se aplicaron usando una Prensa Isostática Modelo CIP42260 (Avure Autoclave Systems, Erie, PA) con una cámara de presurización de 101.6 mm de diámetro interno y 584.2 mm de longitud, la cual opera a una capacidad máxima de 400 MPa. Se usó una mezcla de 5:1 agua:lubricante anticorrosivo (Dietil-amino-etanol; Hydrolubric 120-B, E.F. Houghton y Co., Valley Forge, PA) como líquido presurizante. Latemperatura de la mezcla se ajustó a 25°C antes de presurizar. El tiempo requerido para alcanzar la presión hidrostática de 15, 30 y 60 MPa fue de 3, 10 y 28 s, respectivamente. El tiempo para alcanzar la liberación de la presión fue siempre menor de 30 s. La temperatura del fluido de presurización no fue mayor a 26°C al final de los tratamientos. Los frutos se cubrieron con una película de polietileno de alta densidad antes de introducirlos en la cámara de tratamiento para evitar el contacto directo con el líquido presurizante. Los mangos presurizados se almacenaron a 25 ± 1°C y 85–90% de humedad relativa (HR) hasta la senescencia.
Velocidad de respiración y producción de etileno
La velocidad de respiración (VR) y de producción de etileno (VPE) se determinaron usando el método descrito por Tovar, García, and Mata (Citation2001) y se reportaron en mL/kg/h de CO2 y μL/kg/h de etileno, respectivamente. Para medir la VR se colocaron dos frutos en forma individual por tratamiento en recipientes herméticos por 2 h y se analizó el microambiente generado dentro del recipiente. Esta prueba es no destructiva, por lo que se usaron siempre los mismos ejemplares durante el desarrollo del estudio hasta alcanzar la senescencia. Usando una jeringa se tomó 1 mL del aire presente en el espacio de cabeza de cada frasco y se analizó utilizando un cromatógrafo de gases marca HP modelo 6890. Para medir laVPE se colocaron dos frutos en forma individual por tratamiento en recipientes herméticos por 12 h; para eliminar el CO2 producido se colocó dentro del recipiente hermético un absorbedor de CO2 (200 ml de solución saturada de NaOH en recipiente de vidrio). Las condiciones empleadas en el cromatógrafo fueron: N2 como gas acarreador con un flujo de 7 mL/min, la cámara de inyección y el detector de CO2 se mantuvieron a 250°C, el horno se mantuvo a 50°C por 30 s y posteriormente se aplicó una rampa de 30°C/min hasta alcanzar 80°C, la velocidad de flujo del aire fue de 400 mL/minuto y del H2 de 30 mL/minuto; se usó una columna HP-plot (polímero de divinilbenceno estireno) de 15 m × 0.53 mm y 40 μm de espesor de película. Se empleó un detector de conductividad térmica para detectar CO2 conectado en serie con un detector de ionización de flama para detectar etileno. Para su cuantificación se empleó un patrón de calibración certificado Praxair® conteniendo 11.3 μmol/mol de etileno, 2.01 cmol/mol de CO2, 10.1cmol/mol O2 y N2 para balancear la mezcla.
Actividad de ACC oxidasa
La actividad de la enzima ácido 1-carboxílico -1-amino-ciclopropano oxidasa (ACC oxidasa) in vivo fue determinada usando 0.5 g de pulpa obtenida en forma de discos de 1 mm de espesor y 8 mm de diámetro. Los discos de la pulpa se incubaron con agitación en un vórtex en presencia o en ausencia de 0.5 mL del ácido 1-aminociclopropano-1-carboxílico (ACC) 0.1 mM. La producción de etileno se midió después de 4 h de incubación a 25 ± 1°C por cromatografía de gases empleando las condiciones anteriores para medir etileno. La cantidad de etileno encontrada en los viales sin ACC se restó a la encontrada en los viales con ACC y se expresó la actividad como nmol de etileno/g/h (Cua & Lizada, Citation1990; Mansour, Latché, Vaillant, Pech, & Reid, Citation1986).
Firmeza
La firmeza se determinó midiendo la resistencia de los mangos a la penetración, en la parte cercana a la cavidad basal, hombro dorsal y ventral y cerca del ápice del mango usando un puntal cilíndrico de 0.5 cm de diámetro con un medidor de firmeza marca Shimpo (Lincolnwood, Japón). Los resultados se expresaron en Newtons (N).
Pérdida fisiológica de peso
La pérdida de peso se obtuvo pesando diariamente 10 frutos en una balanza granataria Modelo 1500D Ohaus (Florham Park, NJ) y se reportó la relación existente entre el peso obtenido y el peso inicial, expresado en porcentaje.
Sólidos solubles
Los sólidos solubles totales se determinaron extrayendo el jugo presente en la pulpa de la fruta y analizándolo con un refractómetro Abbé DR-A1 (Atago, Co., Tokyo, Japan) (AOAC, 1984).
Color interno del fruto
Se midió usando la siguiente escala visual subjetiva continua de 10 puntos: 1. crema, 2. crema con áreas amarillas, 3. igual crema y amarillo, 4. amarillo con áreas crema, 5. amarillo, 6. amarillo con áreas naranja claro, 7. naranja claro, 8. naranja claro con áreas naranja intenso, 9. naranja intenso con áreas naranja claro, 10. naranja intenso. El color interno se monitoreó en cinco frutos. En la escala usada el número 1 siempre estuvo al inicio y el 10 al final. Esta evaluación se realizó sólo por el investigador.
Color externo del fruto
El color externo se evaluó visualmente en cinco frutos empleando la escala de la carta oficial de calidad de EMEX para mangos de exportación (EMEX, Citation1998), la cual es la siguiente: 1 = verde, 2 = verde con áreas amarillas, 3 = igual verde y amarillo, 4 = amarillo con áreas verdes, 5 = amarillo, 6 = amarillo con áreas naranja.
Análisis estadístico
Los datos se analizaron con un diseño de bloques completamente al azar con arreglo factorial 3 X 2 (presión hidrostática de 15, 30 y 60 MPa X el tiempo de tratamiento de 10 y 20 min), donde los bloques fueron los días de almacenamiento; para el análisis de varianza se uso el modelo general lineal (GLM) del SAS (The SAS System for WindowsTM. Version 6.11, SAS Institute, Inc., Cary, NC) y se realizó una prueba de medias con el método de Tukey (p ⩽ 0.05). Se realizaron dos réplicas y cada análisis se hizo por triplicado.
Resultados y discusión
Velocidad de respiración
El efecto de la presurización en la velocidad de respiración de los mangos se muestra en la . Los frutos testigo nopresurizados presentaron un incremento en la VR a partirdel día 3 y se encontraron dos máximos en el día 7(113,51 mL/kg/h de CO2) y en el día 9 (131,73 mL/kg/h de CO2) y posteriormente disminuyó ligeramente hasta alcanzar la madurez de consumo. En los mangos presurizados la VR se vio afectada significativamente (p ⩽ 0,01) por el nivel de presión, el tiempo de presurización y su interacción. En la se observa que hay un incremento en la VR desde los primeros días de almacenamiento en relación al testigo en todos los tratamientos y excepto por los mangos presurizados a 15 MPa, no se aprecia la presencia de un pico climatérico definido, aunque la máxima VR parece alcanzarse a los siete días, cuando se les dio el tratamiento a las tres presiones por 20 min. En los frutos tratados a las tres presiones por 10 min fue menor la VR que en los frutos tratados por 20 min, debido al mayor tiempo de exposición al estrés en los frutos por la presión alta. Ninguno de los frutos tratados presentó un patrón respiratorio similar o parecido al de los frutos testigo, en los frutos tratados por 15 o 30 MPa por 10 min la elevación de la VR fue en los primeros días de almacenamiento pero su valor máximo apenas alcanzó la mitad delencontrado en los frutos testigo en el pico climatérico. Este comportamiento provocó un adelanto en la maduración de las frutas de estos tratamientos y los valores bajos de VR en los frutos tratados con 60 MPa por 10 min ocasionaron que no maduraran.
Figura 1. Velocidad de respiración en mangos ‘Ataulfo’ testigo y tratados con altas presiones hidrostáticas por (A) 10 min y (B) 20 min. Los valores representan la media de tres repeticiones en dos muestras distintas. Las barras indican el error estándar.
Figure 1. Respiration rate in “Ataulfo” mango control and treated with high hydrostatic pressure for (A) 10 min and (B) 20 min. The values represent the mean of three replications in two different samples. The bars indícate the standard error.
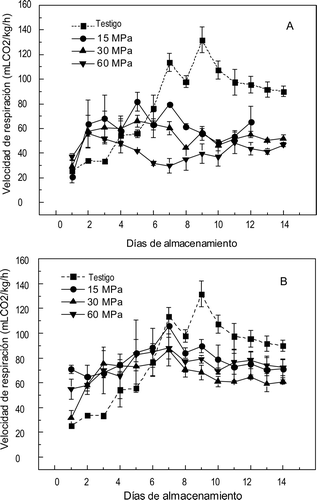
El incremento de la VR en el presente estudio señala que la presurización altera el metabolismo respiratorio de las frutas en función del nivel de presión y tiempo de tratamiento. La alteración puede deberse a cambios estructurales a nivel celular o proteico. Tangwongchai, Ledward, and Ames, (Citation2000) encontraron que a una presión de 200 MPa las células perdieron la estructura colapsando y causando pérdida de turgencia y firmeza en jitomate Cherry. La pérdida de estructura permitiría que el CO2 generado en la respiración saliera con mayor facilidad al exterior del fruto aumentando su concentración en el recipiente. Sin embargo es más factible que el aumento en la VR se deba a una respuesta fisiológica al estrés producido por la presurización como sucede en los mangos golpeados o maltratados (Rodríguez, Ruiz-Altisent, & De la Plaza, Citation1991).
Velocidad de producción de etileno
El efecto de los tratamientos de APH sobre la velocidad de producción de etileno (VPE), la hormona de la maduración se muestra en la . Los frutos no presurizados presentaron la VPE característica del mango ‘Ataulfo’, el etileno se detectó a partir del día 4 y de ahí se elevó hasta alcanzar un valor promedio de VPE de 2,84 μL/kg/h de etileno en el día 14, lo que coincide con Montalvo et al. (Citation2007). La VPE fue afectada significativamente (p ⩽ 0,01) por el nivel de presión, el tiempo de presurización y su interacción. Se ha reportado que a presiones de 100 MPa se activan algunas enzimas (Asaka, Aoyama, Ritsuko, & Hayashi, Citation1994; Jolibert, Tonello, Sagegh, & Raymond, Citation1994) si son enzimas monoméricas, mientras que presiones más elevadas generalmente las inactivan y esta inactivación depende fuertemente del tipo de enzima, pH, composición del medio y temperatura (Balny & Masson, Citation1993). En la producción de etileno participan muchas enzimas, las cuales pueden haber sido afectadas por las presiones utilizadas; así, a la presión de 15 MPa por 10 min la VPE entre los días 7 y 11 se encuentra mayor VPE que en el testigo, lo cual podría deberse a la elevación de la actividad de esas enzimas y esto es más evidente en el tratamiento de 15 MPa por 20 min entre los días 7 y 11, en donde la VPE fue mayor al testigo y a los otros tratamientos. Por otra parte, en los mangos sometidos a 30 y 60 MPa se disminuyó notablemente la producción de etileno en relación al testigo y posiblemente esto se deba a una inhibición parcial de la actividad de las enzimas que son responsables de la producción de etileno, ya que a la presión mayor hubo menor VPE.
Figura 2. Velocidad de producción de etileno en mangos ‘Ataulfo’ testigo y tratados con altas presiones hidrostáticas por (A) 10 min y (B) 20 min. Los valores representan la media de tres repeticiones en dos muestras distintas. Las barras indican el error estándar.
Figure 2. Production rate of ethylene in “Ataulfo” mango control and treated with high hydrostatic pressure for (A) 10 min and (B) 20 min. The values represent the mean of three replications in two different samples. The bars indícate the standard error.
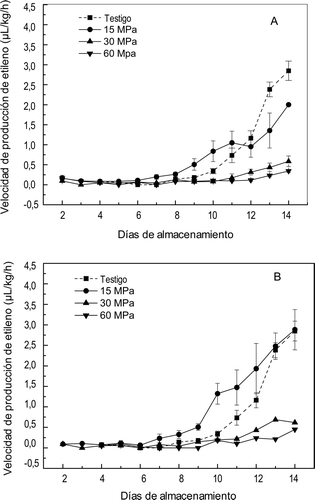
Actividad de ACC oxidasa
El efecto de las APH sobre la ACC oxidasa, enzima que cataliza la última etapa en la síntesis del etileno se muestra en la . Se observó un efecto significativo (p ⩽ 0,01) por efecto del nivel de presión y del tiempo, pero no de su interacción. En los frutos testigo el incremento de la actividad de la ACC oxidasa se correlacionó (coeficiente 0.997) con el aumento de la VPE (Figura 2). Se observó que las frutas presurizadas a 15 MPa presentaron un incremento en la actividad de la ACC oxidasa a partir del día 5, superior al de las muestras presurizadas a 30 y 60 MPA. Este incremento parece estar asociado con el aumento en la VPE que presentaron las frutas presurizadas a 15 MPa. También seencontraron correlaciones positivas entre la actividad de ACC oxidasa y la VPE para todos los tratamientos excepto el tratado con 60 MPa por 10 min. En los tratamientos dados durante 10 min la actividad de ACC oxidasa fue menor que la del testigo, lo cual sugiere que esta enzima se pudo inactivar parcialmente, como se menciona en el apartado anterior, mientras que en los tratamientos dados durante 20 min podría haber existido activación en los frutos tratados por 15 y 60 MPa al final del almacenamiento, ya que sus valores promedio fueron superiores al encontrado en el testigo. Sin embargo, se observa que en las muestras presurizadas a 30 y 60 MPa durante 20 min se incrementó la actividad de ACC oxidasa a partir del día 8, pero este incremento no se reflejó en un aumento en la VPE (Figura 3). También se ha reportado que los enlaces covalentes de las proteínas son más difíciles de cambiar por las presiones altas pero no así los enlaces de hidrógeno y los iónicos, los cuales pueden ayudar a estabilizar una estructura terciaria y loscuales pueden ocasionar cambios dramáticamente (Hendricks, Ludikhuyze, Van den Broeck, & Weemaes, Citation1998). Estos cambios podrían estar afectando la estabilidad de la ACC oxidasa.
Figura 3. Actividad de ACC oxidasa en mangos ‘Ataulfo’ testigo y tratados con altas presiones hidrostáticas por (A) 10 min y (B) 20 min. Los valores representan la media de tres repeticiones en dos muestras distintas. Las barras indican el error estándar.
Figure 3. Activity of ACC oxidase in mango fruit “Ataulfo” control and treated with high hydrostatic pressure for (A) 10 min and (B) 20 min. The values represent the mean of three replications in two different samples. The bars indícate the standard error.
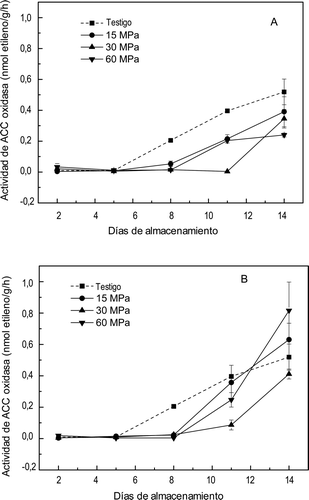
Firmeza
Los cambios de firmeza se muestran en la . Los frutos no tratados por APH mostraron mayores valores de firmeza durante el almacenamiento, que disminuyó gradualmente para alcanzar el mismo valor de firmeza el día 14 estando el fruto maduro. Las APH afectaron significativamente la firmeza (p ⩽ 0,01) en los mangos presurizadas por 20 min desde el primer día. Sin embargo la pérdida de firmeza dependió más del nivel de presión que del tiempo de presurización y no se encontró un efecto significativo en la interacción de ambos parámetros. Así en los frutos tratados por 60 MPa durante 10 o 20 min la firmeza fue mayor entre los días 5 y 11 que en los otros frutos tratados aunque menor que en el testigo; esto puede deberse a que a presión mayor pudieron inactivarse parcialmente las enzimas como la pectimetilesterasa (Hendricks et al., Citation1998). Se ha reportado que las APH modifican la actividad de la pectinmetilesterasa (Basak & Ramaswamy, Citation1996).
Figura 4. Firmeza en mangos ‘Ataulfo’ testigo y tratados con altas presiones hidrostáticas por (A) 10 min y (B) 20 min. Los valores representan la media de tres repeticiones en dos muestras distintas. Las barras indican el error estándar.
Figure 4. Firmness in mango “Ataulfo” control and treated with high hydrostatic pressure for (A) 10 min and (B) 20 min. The values represent the mean of three replications in two different samples. The bars indícate the standard error.
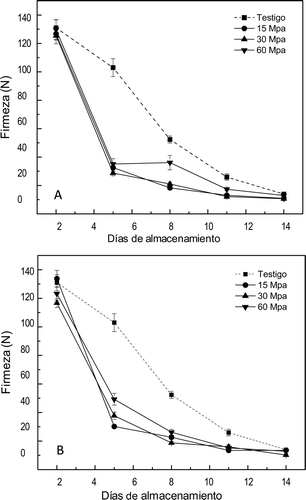
Los cambios en la textura inducidos por las APH pueden ocasionarse por un efecto físico directo, en el que se altera la macro o microestructura del tejido y de su pared celular por cambios fisicoquímicos en las proteínas que integran las membranas celulares afectando la permeabilidad y de las enzimas metabólicas o a la combinación de factores. Puesto que los cambios en la textura no fueron inmediatos, sino que se presentaron durante los primeros días de almacenamiento, el fenómeno debe estar asociado con cambios en la fisiología del fruto. A este respecto, Guerrero-Beltrán, Swanson, and Barbosa-Canovas (Citation2005) reportaron cambios en la pulpa de mango presurizada asociada con la descompartamentalización de las células. Es importante considerar que si las APH dañan los tejidos se pueden ocasionar cambios de permeabilidad en la membrana celular, facilitando el contacto de los sustratos con sus enzimas (Tangwongchai et al., Citation2000). Esto favorecería la actividad simultánea de la pectinmetilesterasa y la poligalacturonasa, afectando la firmeza del fruto, ya que la poligalacturonasa despolimeriza la pectina que ha sido desmetilada previamente por la pectinmetilesterasa (Knorr, Citation1995; Oey, Lille, Van Loey, & Hendrickx, Citation2008). La presencia de estos dos tipos de enzimas se han reportado en mango ‘Ataulfo’ (Montalvo, Adame, García, Tovar, & Mata, Citation2009; Ketsa, Chidtragool, Klein, & Lurie, Citation1999).
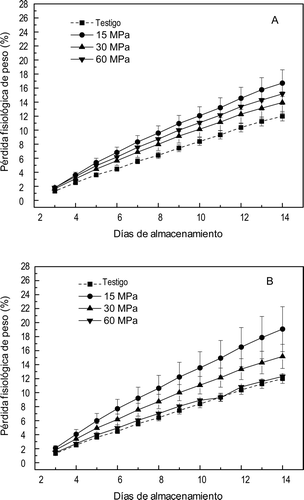
Pérdida fisiológica de peso
En la se muestra la evolución de la pérdida fisiológica de peso. En general los frutos testigo no presurizados presentaron los valores más bajos de pérdida de peso. Las frutas sometidas a APH presentaron cambios enla pérdida fisiológica de peso, afectadas por la presión y el tiempo. Los frutos tratados a menor presión y tiempo presentaron mayor pérdida de peso. Observándose que la pérdida de peso de los frutos tratados a 60 MPa por 20 min no fue diferente significativamente (p ⩽ 0,01) que la pérdida de los frutos no presurizados. Este comportamiento indica que los cambios observados en la pérdida de peso parecen estar relacionados con el daño en la firmeza asociada con la alteración de la estructura interna y cambios de permeabilidad (Figura 4) y el incremento en la VR (Figura 1) y la VPE (Figura 2).
Figura 5. Pérdida fisiológica de peso en mangos ‘Ataulfo’ testigo y tratados con altas presiones hidrostáticas por (A) 10 min y (B) 20 min. Los valores representan la media de tres repeticiones en dos muestras distintas. Las barras indican el error estándar.
Figure 5. Physiological weight loss in mango “Ataulfo” control and treated with high hydrostatic pressure for (A) 10 min and (B) 20 min. The values represent the mean of three replications in two different samples. The bars indícate the standard error.
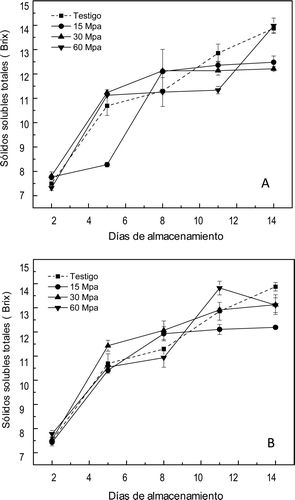
Sólidos solubles totales
La evolución de los sólidos solubles totales se muestra en la . En los sólidos solubles totales se encontró efecto altamente significativo (p ⩽ 0,01) por la presión y el tiempo, pero no se encontró efecto debido a la interacción de estos dos factores. El contenido de sólidos solubles a lo largo de la maduración de los frutos se mostró con un comportamiento distinto para los frutos testigo comparados con los frutos presurizados. Los frutos sometidos a APH mostraron un contenido menor de sólidos solubles que los frutos testigo, con excepción de los mangos sometidos a 60 MPa, que alcanzaron el mismo contenido de sólidos solubles que los frutos testigo durante la exposición por 10 minutos.
Figura 6. Sólidos solubles totales en mangos ‘Ataulfo’ testigo y tratados con altas presiones hidrostáticas por (A) 10 min y (B) 20 min a 25°C y almacenados hasta maduración completa a 25 ± 2°C. Los valores representan la media de tres repeticiones en dos muestras distintas. Las barras indican el error estándar.
Figure 6. Total soluble solids in mango “Ataulfo” control and treated with high hydrostatic pressure for (A) 10 min and (B) 20 min. The values represent the mean of three replications in two different samples. The bars indícate the standard error.
Color interno
El color interno del mango ‘Ataulfo’ se desarrolló en forma normal en los mangos testigo no presurizados, como se observa en la , pasando de un color amarillo en el día 11 (valor 5 en la escala usada) a un color naranja con tonos amarillos (valor 9) en el día 14. Las áreas amarillas estaban presentes cerca de la cáscara, ya que el proceso de maduración inicia de la semilla hacia la cáscara.
Tabla 1. Efecto de los tratamientos de Alta Presión Hidrostática en el color interno y externo del mango ‘Ataulfo’.
Table 1. Effect of high hydrostatic pressure treatments on the inner and external color of ‘Ataulfo’ Mango.
Los frutos sometidos a APH mostraron un desarrollo del color interno diferente al del los frutos no tratados. En el día 11 los mangos presurizados presentaron un color interno con mayor incidencia de tonalidades naranja (valores superiores a 6) excepto el mango presurizado a 60 MPa por 10 min, que fue amarillo (valor 5). Sin embargo en el día 14, los mangos presurizados tendieron a presentar menos incidencia de coloraciones naranja (valores menores a 9), comparados con el mango testigo. En los mangos presurizados el desarrollo del color interno fue irregular, presentándose alta variabilidad y por ello fue difícil establecer diferencias significativas. En los tratamientos de 30 y 60 MPa se presentó una mayor incidencia de áreas amarillas claras cerca a la cavidad basal, lo cual parece indicar que esta zona parece ser más sensible a cambios inducidos por alta presión.
Color externo
Los cambios en el color externo se muestran en la . Los frutos testigo mostraron una evolución normal en el desarrollo del color externo, pasando de un color verde amarillo en el día 11 (valores 3 y 4 en la escala), que pasó a un color amarillo (valor 5) en el día 14. Los mangos presurizados presentaron tonalidades más amarillas que los mangos testigo en el día 11. Sin embargo, el desarrollo del color varió entre frutos, por lo que no fue posible establecer una diferencia significativa. En el caso de los mangos presurizados a 60 MPa por 20 min se inhibió el desarrollo homogéneo de la coloración amarilla, presentando un valor de 1.9. En el día 14 se observa que los mangos presurizados por 20 min, o a 30 y 60 MPa por 10 min presentaron menor incidencia de tonalidades amarilla externas, pero debido a la variabilidad entre frutos, esta diferencia solo fue significativa para los mangos presurizados a 60 MPa por 20 min. El cambio de color en los frutos estuvo asociado con la VPE (Figura 2), ya que los mangos sometidos a mayor presión presentaron menor VPE y menor desarrollo de color. En el mango ‘Ataulfo’ al ser una fruta climatérica la maduración está regulada por el etileno. Esta hormona puede verse afectada en su biosíntesis y/o percepción por cambios debidos al desarrollo del fruto, exposición a medio ambiente adverso como la refrigeración o en este caso las APH (Lelièvre, Latché, Jones, Bouzayen, & Pech, Citation1997). Se ha demostrado que el etileno afecta la biosíntesis de los pigmentos en la fruta y acelera la degradación de la clorofila (Lelièvre et al., Citation1997; Montalvo et al., Citation2009). En el presente estudio se observó que la baja producción de etileno en los frutos presurizados, estuvo asociada con la alteración en el desarrollo del color amarillo-anaranjado característico de los mangos Ataulfo. Este fenómeno podría estar asociado con la baja producción de etileno o una modificación en la percepción del mismo, ya que ambos factores inhiben la biosíntesis de los carotenos.
Daño aparente
Los mangos presurizados presentaron cambios físicos internos y externos apreciables de manera evidente. Los mangos sometidos a APH desarrollaron internamente, a partir del día 11 un tejido esponjoso que implica la formación de cavidades o hendiduras (a). La formación del tejido esponjoso fue mayor en los mangos sometidos a tratamientos más drásticos de presurización (presión y tiempo). El desarrollo de tejido esponjoso es una alteración fisiológica que se manifiesta con frecuencia en los mangos de la variedad Alfonso y ha sido asociado con alteraciones en el metabolismo del almidón, provocando una disminución en la concentración de azúcares (Gupta, Lad, Chavan, & Salvi, Citation1985; Katrodia & Rane, Citation1988). El tejido esponjoso también se ha reportado en mango Tommy Atkins, asociado con el metabolismo del almidón (Oliveira-Lima, Bosco-Chitarra & Chitarra, Citation2001). Recientemente se ha relacionado la formación del tejido esponjoso con una disminución de la actividadde la ACC oxidasa en mango Alfonso (Nagamani, Shivashankara, & Roy, Citation2010). También se ha reportado el desarrollo de tejido esponjoso durante la maduración de mangos Alfonso sometidos a tratamientos cuarentenarios de irradiación (Mercado-Silva, Citation2011).
Figura 7. Efecto de la presurización en la (A) apariencia interna y (B) apariencia externa de mangos ‘Ataulfo’ presurizados a 60 MPa por 20 min.
Figure 7. Effect of pressurization on the (A) internal appearance and (B) external appearance of mango “Ataulfo” pressurized to 60 MPa for 20 min.
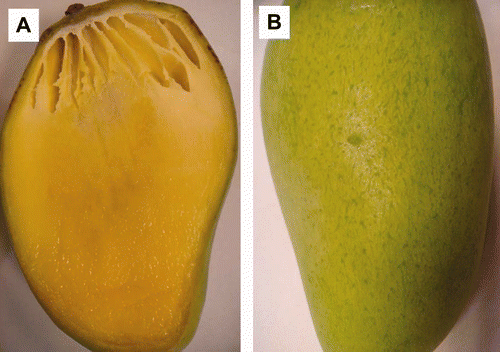
Los resultados del presente estudio indican que las APH pueden inducir la formación de tejido esponjoso (Figura 7), asociado con alteraciones metabólicas, entre ellas una disminución en la actividad de la ACC oxidasa (Figura 3) y su influencia en la producción de etileno (Figura 2), así como la alteración del metabolismo de los almidones y su transformación en azúcares (Figura 6).
Los frutos presurizados presentaron también alteraciones externas. Los frutos presurizados a 15 MPa por 10 y 20 min presentaron deformaciones en la forma a partir del día 7. En los frutos presurizados a 30 MPa por 10 min se observaron las deformaciones a partir del día 8. Los frutos presurizados a 30 MPa por 20 min y 60 MPa por 10 ó 20 min, presentaron en su superficie zonas verdes claras y oscuras inmediatamente después del tratamiento (Figura 7b), las cuales se oscurecieron cinco días después. Estos frutos presentaron también deformaciones a partir del día 8.
Conclusiones
La aplicación de APH ha sido propuesta como un método cuarentenario alternativo para el control de las moscas de la fruta Anastrepha ludens y A. oblicua que infectan los mangos. Los tratamientos propuestos han sido optimizados para poder destruir los huevos y larvas de estas plagas a 90 MPa por 9 min a 45°C ó 0°C (Ramírez, Castañón-Rodríguez, Velazquez, Montoya, & Vázquez, 2011). En el presente estudio, los resultados preeliminares señalan que el mango ‘Ataulfo’ se ve afectado por las altas presiones hidrostáticas, mostrando algunas alteraciones fisiológicas inmediatas, pero que en general se manifestaron durante el desarrollo de la maduración postcosecha en el almacenamiento a condiciones controladas. El efecto de las APH fue causado principalmente por el nivel de presión, seguido por el tiempo de presurización y en algunos casos por la interacción de ambos factores. Los daños parecen estar asociados con la descompartamentailización de la estructura celular, la afectación de la permeación de las membranas celulares y la afectación del metabolismo alterando diferentes procesos de síntesis. Estos cambios no inhibieron el proceso de maduración pero modificaron las características de calidad del mismo. El presente estudio permitió entender el efecto de las APH en mango ‘Ataulfo’ en madurez fisiológica. Sin embargo, es necesario estudiar el efecto de las APH en diferentes etapas de la maduración del mango para determinar como se altera la fisiología postcosecha del fruto, cuando se somete a APH verde, en madurez fisiológica o cuando el fruto ha iniciado elclimaterio. También se requiere estudiar el efecto de las APH en las diferentes variedades de mango destinadas a la exportación, ya que los mangos responden en forma diferente a los métodos cuarentenarios y el mango ‘Tommy Atkins’ pareció mostrar mayor resistencia a las APH que la que se reporta en este estudio para el mango ‘Ataulfo’ (Candelario-Rodríguez et al., Citation2009).
Agradecimientos
Los autores agradecen el financiamiento otorgado por el Programa de Mejoramiento del Profesorado (PROMEP), así como a CONACyT a través de las becas para estudios de Maestría en Ciencias para la realización de este trabajo. Asimismo, se agradece la contribución del Sr. Abelardo González Cabrera al aportar los mangos para la realización de este estudio. Se agradece al Fondo Mixto CONACYT – Gobierno del Estado de Tamaulipas por el apoyo para el desarrollo y publicación de este trabajo.
Referencias
- AOAC . 1984 . Official methods of analysis , (13th ed.) , Washington , DC : Association of Official Analytical Chemists .
- Armstrong , J.W. and Mangan , R.L. 2007 . “ Commercial quarantine heat treatments ” . In Heat treatments for postharvest pest control: theory and practice , Edited by: Tang , En J. , Mitcham , E. , Wang , S. and Lurie , S. 311 – 340 . Wallingford , UK : CAB International .
- Asaka , M. , Aoyama , Y. , Ritsuko , N. and Hayashi , R. 1994 . High pressure and temperatura effects on enzyme inactivation in strawberry and orange products . Journal of Food Science , 62 : 85 – 88 .
- Balny , C. and Masson , P. 1993 . Effects of high pressure on proteins . Food Review International , 9 : 611 – 628 .
- Basak , S. and Ramaswamy , H.S. 1996 . Ultrahigh-pressure treatment of orange juice: A kinetic study on inactivation of pectin methyl esterase . Food Research International , 29 : 601 – 607 .
- Butz , P. and Tauscher , B. 1995 . Inactivation of fruit fly eggs by high pressure treatment . Journal of Food Processing and Preservation , 19 : 161 – 164 .
- Camargo , C.A. , Odell , E. and Jirón , L.F. 1996 . Interspecific interactions and host preference of Anastrepha obliqua and Ceratitis capitata (Diptera: Tephritidae), two pests of mango in Central America . The Florida Entomologist , 79 : 266 – 268 .
- Candelario-Rodríguez , H.E. , Hurtado-González , M. , Morales-Castro , J. , Velázquez , G. , Ramírez , J. A. , Loera-Gallardo , J. and Vázquez , M. 2009 . Efficacy of high hydrostatic pressure as aquarantine treatment to improve the quality of mango fruits infested by the Mexican fly . Anastrepha ludens. CyTA Journal of Food , 7 : 135 – 142 .
- Cua , A.H. and Lizada , M.C.C. 1990 . Ethylene production in the Carabao mango (Mangifera indica L) fruit during maturation and ripening . Acta Horticulturae , 269 : 169 – 179 .
- EMEX , A.C. 1998 . Empacadoras de Mango para Exportación. Boletín. Norma de calidad para mango fresco de exportación Elaborada por Comité Técnico Científico de Empacadores de Mango de Exportación, A. C. (EMEX, A. C.) Guadalajara, Jalisco, Mexico. 6 p
- Gazit , Y. , Rossler , Y. , Wang , S. , Tang , J. and Lurie , S. 2004 . Thermal death kinetics of egg and third instar mediterranean fruit fly (Diptera: Tephritidae) . Journal of Economic Entomology , 97 : 1540 – 1546 .
- Guerrero-Beltrán , J.A. , Swanson , B.G. and Barbosa-Canovas , G.V. 2005 . High hydrostatic pressure processing of mango puree containing antibrowning agents . Food Science and Technology International , 11 : 261 – 267 .
- Gupta , D.N. , Lad , B.L. , Chavan , A.S. and Salvi , M.J. 1985 . Enzyme studies on spongy tissue: A physiological ripening disorder inAlphonso mango . Journal of the Majarashtra Agricultural University , 10 : 280 – 282 .
- Hallman , G.J. 2000 . Factors affecting quarantine heat treatment efficacy . Postharvest Biology and Technology , 21 : 95 – 101 .
- Hallman , G.J. and Zhang , Q.H. 1997 . Inhibition of fruit fly (Diptera: Tephritidae) development by pulsed electric field . The Florida Entomologist , 80 : 239 – 248 .
- Hendricks , M. , Ludikhuyze , L. , Van den Broeck , I. and Weemaes , C. 1998 . Effects of high pressure on enzymes related to food quality . Trends in Food Science and Technology , 9 : 197 – 203 .
- Hernández , E. , Orozco , D. , Breceda , S.F. and Domínguez , J. 2007 . Dispersal and longevity of wild and mass-reared Anastrepha ludens and Anastrepha obliqua (Diptera: Tephritidae) . The Florida Entomologist , 90 : 123 – 135 .
- Jolibert , F. , Tonello , C. , Sagegh , P. and Raymond , J. 1994 . Les effets des hautes pressions sur la polyphénol oxydase des fruits . Bioscience Boissons , 251 : 27 – 35 .
- Katrodia , J.S. and Rane , D.A. 1988 . Pattern of distribution of spongy tissue in the affected Alphonso fruit at different locations . Acta Horticulturae , 231 : 873 – 877 .
- Ketsa , S. , Chidtragool , S. , Klein , J.D. and Lurie , S. 1999 . Firmness, pectin components and cell wall hydrolases of mango fruit following low-temperatures stress . Journal of Horticultural Science and Biotechnology , 74 : 685 – 689 .
- Knorr , D. 1995 . “ High-pressure effects on plant derived foods ” . In High-pressure processing of foods , Edited by: Ledward , En D.A. , Johnston , D.E. , Earnshaw , R.G. and Hasting , A.P.M. 125 – 134 . Nottingham : Nottingham University Press .
- Lelièvre , J.M. , Latché , A. , Jones , B. , Bouzayen , M. and Pech , J.C. 1997 . Ethylene and fruit ripening . Plant Physiology , 101 : 727 – 739 .
- Mangan , R.L. and Hallman , G.J. 1998 . “ Temperature treatments forquarantine security new approaches for fresh commodities ” . In Temperature sensitivity in insects and application in integrated pest management , Edited by: Hallman , En G.J. and Denlinger DL , D.L. 201 – 236 . Boulder , CO : Westview Press .
- Mansour , R. , Latché , A. , Vaillant , V. , Pech , J.C. and Reid , M.S. 1986 . Metabolism of 1-aminocyclopropane-1-carboxylic acid in ripening apple fruits . Physiologia Plantarum , 66 : 495 – 502 .
- Mercado-Silva , E. 2011 . “ Ionizing radiation as quarantine treatments in fruits ” . In Advances in post-harvest treatments and fruit quality and safety , Edited by: Ramírez , En J.A. and Vázquez , M. 19 – 30 . Hauppauge , NY : Nova Science Publishers .
- Montalvo , E. , Adame , Y. , García , H.S. , Tovar , B. and Mata , M. 2009 . Changes of sugars, b-carotene and firmness of refrigerated Ataulfo mangoes treated with exogenous ethylene . Journal of Agricultural Science , 147 : 193 – 199 .
- Montalvo , E. , García , H.S. , Tovar , B. and Mata , M. 2007 . Application of exogenous ethylene on postharvest ripening of refrigerated “Ataulfo” mangoes . Lebensmittel Wissenschaft und Technologie , 40 : 1466 – 1472 .
- Monzon , M.E. , Biasi , B. , Mitcham , E.J. , Wang , S.J. , Tang , J.M. and Hallman , G. 2007 . Effect of radiofrequency heating on the quality of ‘‘Fuyu’’ persimmon fruit as a treatment for control of the Mexican fruit fly . Hortscience , 42 : 125 – 129 .
- Nagamani , J.E. , Shivashankara , K.S. and Roy , T.K. 2010 . Role of oxidative stress and the activity of ethylene biosynthetic enzymes on the formation of spongy tissue in ’Alphonso’ mango . Journal of Food Science and Technology-Mysore , 47 : 295 – 299 .
- Neven , L. , Follet , P.A. and Raghubeer , E. 2007 . Potential for high hydrostatic pressure processing to control quarantine insects in fruit . Journal of Economy Entomology , 100 : 1499 – 1503 .
- Oey , I. , Lille , M. , Van Loey , A. and Hendrickx , M. 2008 . Effect ofhigh-pressure processing on color, texture and flavour of fruit-and vegetable-based food products: a review . Trends in Food Science and Technology , 19 : 320 – 328 .
- Oliveira-Lima , L.C. , Bosco-Chitarra , A. and Chitarra , M.I.F. 2001 . Changes in amylase activity starch and sugars contents in mango fruits pulp cv. Tommy Atkins with spongy tissue . Brazilian Archives of Biology and Technology , 44 : 59 – 62 .
- Ragazzo-Sánchez , J.A. , Sánchez-Prado , L. , Gutiérrez-Martínez , P. , Luna-Solano , G. , Gómez-Gil , B. and Calderón-Santoyo , M. 2009 . Inhibition of Salmonella spp. isolated from mango using bacteriocin-like produced by lactobacilli . CyTA – Journal of Food , 7 : 181 – 187 .
- Ramírez , J.A. , Castañón-Rodríguez , J.F. , Velázquez , G. , Montoya , P. and Vázquez , M. 2011 . Resistance of West Indian fruit fly Anastrepha oblicua Macquart to quarantine treatments of high hydrostatic pressure combined with cold . Food and Bioprocess Technology , In press. DOI: 10.1007/s11947-011-0550-6
- Rodríguez , S.L. , Ruiz-Altisent , M. and De la Plaza , P.J.L. 1991 . Bruise development and fruit response of pear (c.v. ‘Blanquilla´) under impact conditions . Journal of Food Engineering , 14 : 289 – 301 .
- Sosa-Morales , M.E. , Tiwari , G. , Wang , S. , Tang , J. , García , H.S. and López-Malo , A. 2009 . Dielectric heating as a potential post-harvest treatment of disinfesting mangoes, Part II: Development of RF-based protocols and quality evaluation of treated fruits . Biosystems Engineering , 103 : 287 – 296 .
- Sosa-Morales , M.E. , Valerio-Junco , L. , López-Malo , A. and García , H.S. 2010 . Dielectric properties of foods: reported data in the 21st century and their potential applications . LWT-Food Science and Technology , 43 : 1169 – 1179 .
- SAGARPA . 2007 . Superficie de mango, volumen de producción y rendimiento promedio por municipio y variedad. Subdelegación Agropecuaria. Programa de Fomento Agrícola y Programa de Sanidad Vegetal. Delegación Estatal en Nayarit. Tepic, México. 10 p
- Tangwongchai , R. , Ledward , D.A. and Ames , J.M. 2000 . Effect of high pressure treatment on the texture of cherry tomato . Journal of Agriculture Food Chemistry , 48 : 1434 – 1441 .
- Tiwari , G. , Wang , S. , Birla , S.L. and Tang , J. 2008 . Effect of water-assisted radio frequency heat treatment on the quality of ‘‘Fuyu’’ persimmons . Biosystems Engineering , 100 : 227 – 234 .
- Tovar , B. , García , H.S. and Mata , M. 2001 . Physiology of pre-cut mango. II Evolution of organics acids . Food Research International , 34 : 705 – 714 .
- Varith , J. , Sirikajornjaru , W. and Kiatsiriroat , T. 2007 . Microwave-vapor heat disinfestation on oriental fruit fly eggs in mangoes . Journal of Food Processing and Preservation , 31 : 253 – 269 .
- Velázquez , G. , Candelario , H.E. , Ramírez , J.A. , Montoya , P. , Loera-Gallardo , J. and Vázquez , M. 2010a . An improved quarantine method against the Mexican fruit fly based on high hydrostatic pressure combined with heat to maintain fruit quality . Foodborne Pathogens and Disease , 7 : 493 – 498 .
- Velázquez , G. , Candelario-Rodríguez , H.E. , Ramírez , J.A. , Mangan , J.L. , Loera-Gallardo , J. and Vázquez , M. 2010b . High hydrostatic pressure at low temperature as a quarantine treatment to improve the quality of fruits . Foodborne Pathogens and Disease , 7 : 287 – 292 .
- Wang , S. , Monzon , M. , Johnson , J.A. , Mitcham , E.J. and Tang , J. 2007 . Industrial scale radio frequency treatments for insect control in walnuts II: insect mortality and product quality . Postharvest Biology and Technology , 45 : 247 – 253 .